Beyond Capricornia: Tropical Sea Slugs (Gastropoda, Heterobranchia) Extend Their Distributions into the Tasman Sea



1
National Marine Science Centre, Southern Cross University, Bay Drive, Coffs Harbour, NSW 2450, Australia
2
Marine Ecology Research Centre, Southern Cross University, Lismore, NSW 2458, Australia
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(3), 99; https://doi.org/10.3390/d10030099
Submission received: 7 August 2018
/
Revised: 27 August 2018
/
Accepted: 28 August 2018
/
Published: 4 September 2018
(This article belongs to the Special Issue Climate Change and Human Activities on Coral Reefs)
Abstract
:There is increasing evidence of poleward migration of a broad range of taxa under the influence of a warming ocean. However, patchy research effort, the lack of pre-existing baseline data, and taxonomic uncertainty for some taxa means that unambiguous interpretation of observations is often difficult. Here, we propose that heterobranch sea slugs provide a useful target group for monitoring shifts in distribution. As many sea slugs are highly colourful, popular with underwater photographers and rock-pool ramblers, and found in accessible habitats, they provide an ideal target for citizen scientist programs, such as the Sea Slug Census. This maximises our ability to rapidly gain usable diversity and distributional data. Here, we review records of recent range extensions by tropical species into the subtropical and temperate waters of eastern Australia and document, for the first time in Australian waters, observations of three tropical species of sea slug as well as range extensions for a further six to various locations in the Tasman Sea.
1. Introduction
By far the majority of Indo-Pacific sea slug species are tropical [1] with diversity declining away from the equator. The highest diversity occurs in the tropical western Pacific, and in particular in the area known as the Coral Triangle (bounded by Indonesia, the Philippines, and the Solomon Islands). However, records are extremely patchy both spatially and temporally, with many regions receiving very little recent attention (e.g., eastern Indonesia) [2]. On the Australian east coast, diversity attenuates from the northern Great Barrier Reef (GBR), Queensland (QLD), where approximately 1000 species have been recorded [3], to the southern tip of the mainland with 400 species recorded from Victoria and Bass Strait [4]. The subtropical region of New South Wales (NSW) supports both tropical and temperate taxa representing a biogeographic ecotone, as well as several endemic taxa restricted to this overlap zone [5].
The oceanographic conditions on the central eastern Australian coast, which extends from the southern GBR in southern QLD to northern NSW, are heavily influenced by the East Australian Current (EAC) which delivers warm water from the tropical Coral Sea into the subtropics [6]. Its southward-flowing waters move along the continental slope but are diverted east at South West Rocks, NSW to form an eastward-flowing current: the Tasman Front [7,8], which transports warm water across the northern Tasman Sea to Lord Howe Island and New Zealand [9]. In addition, eddies of the EAC regularly flood the continental shelf in the region. This combination of circulation patterns leads to rapid changes in water temperature, sometimes as high as 7 °C over a 24 h period, as the EAC drives upwelling in coastal waters [8,10].
In the Solitary Islands Marine Park, some 150 km north of the eastward diversion of the EAC, where shallow reefs are present along a cross-shelf gradient, temperature loggers deployed at depths of ~10 m, indicate a consistent gradient of water temperature with offshore sites (8–11 km offshore) experiencing temperatures that are 1–1.5 °C warmer than nearshore sites [8]. This gradient in water temperature is reflected in biological patterns with increasing representation of tropically affiliated species of coral [11], fish [12], and molluscs [13,14] offshore. It is hypothesised that these patterns are maintained by recruitment of entrained pelagic larvae transported southward by the EAC [11,13]. For example, some tropical reef fish are known to be transported six degrees of latitude south of their usual range [15,16,17]. Variability associated with recruitment events (supply, transport, settlement, survival) may lead to substantial changes in the structure of assemblages at the receiving sites [11]. Despite this variability, and in line with predictions of the biological effects of a strengthening EAC and increasing water temperatures, reports of southward range extensions are rapidly increasing.
1.1. An Emerging Trend—Poleward Range Extensions in Sea Slug Distributions
Over the past 10 years, records of sea slug species found poleward of their previously known range have proliferated in the scientific literature [18,19,20,21,22]. There is little doubt, however, that some of the reports simply reflect a lack of previous scientific scrutiny through limited survey effort. For example, the 23 sea slug species listed by Bertsch [21] as being new to the Gulf of California may simply be a product of increased search effort under the objective of generating a regional species list rather than recent introductions. It is also likely that increased effort associated with the development of a regional inventory resulted in the first reports of: Janolus fuscus O’Donoghue, 1924 into the Sea of Japan [23]; four species into southern Brazil [18]; the occurrence of Geitodoris planata (Alder & Hancock, 1846) on the North Sea coast of the United Kingdom [24]; and northern range extensions for nine taxa from temperate Chilean waters into Peru [22].
However, there are also many substantiated records which result from invasion by non-indigenous fauna, or poleward movement due to warming, for example, Spurilla neapolitana (Delle Chiaje, 1841) into Peru from Costa Rica [25] and Bathydoris aioca Marcus & Marcus, 1962 from Baja California, Mexico to Oregon USA [26]. Anomalous warming in the north-eastern Pacific also resulted in range-shifts for 30 mollusc species from California to Oregon [27]. The regional extinction and subsequent slow recovery of Felimare californiensis (Bergh, 1879) in California, putatively due to regional-scale anthropogenic effects rather than broadscale climatic changes [28], illustrates the importance of comprehensive baseline data to help interpret the scale, and likely causes, of subsequent changes.
Given that range extensions are occurring, an important question is how this will affect the ecology of the receiving sites. However, despite general consensus that cumulative impacts may have global consequences [29], data on species-specific impacts are lacking. Indeed, only one published study to date specifically addressed putative impacts of altered population dynamics. Goddard, Gosliner, & Pearse [30] investigated the effects of a poleward range shift by the nudibranch Phidiana hiltoni (O′Donoghue, 1927) in California. They noted a concomitant decline in the abundance of other nudibranch taxa with subsequent dietary analysis and feeding experiments indicating predation by P. hiltoni as a likely cause. On the eastern and southern coasts of Australia, the introduction and rapid spread of the aeolid nudibranch Spurilla braziliana on intertidal rocky reefs [5,31], has anecdotally been linked to a reduction in the abundance of an indigenous aeolid species, Austraeolis ornata (SDAS, MN, pers. obs.), although quantitative data are currently lacking.
Lessepsian migration, the unidirectional migration of marine taxa from the Red Sea to the eastern Mediterranean via the Suez Canal [32], is assumed to be the means by which several sea slugs have shifted their distribution into the eastern Mediterranean Sea [33,34] and southern European shores [34,35]. Whilst regarded as potentially invasive, many may simply be casual or vagrant animals. However, some are clearly invasive, such as the sea hares Syphonota geographica (Adams & Reeve, 1850) and Bursatella leachii De Blainville, 1817 [34,35,36].
1.2. Recent Poleward Range Extensions in Eastern Australia
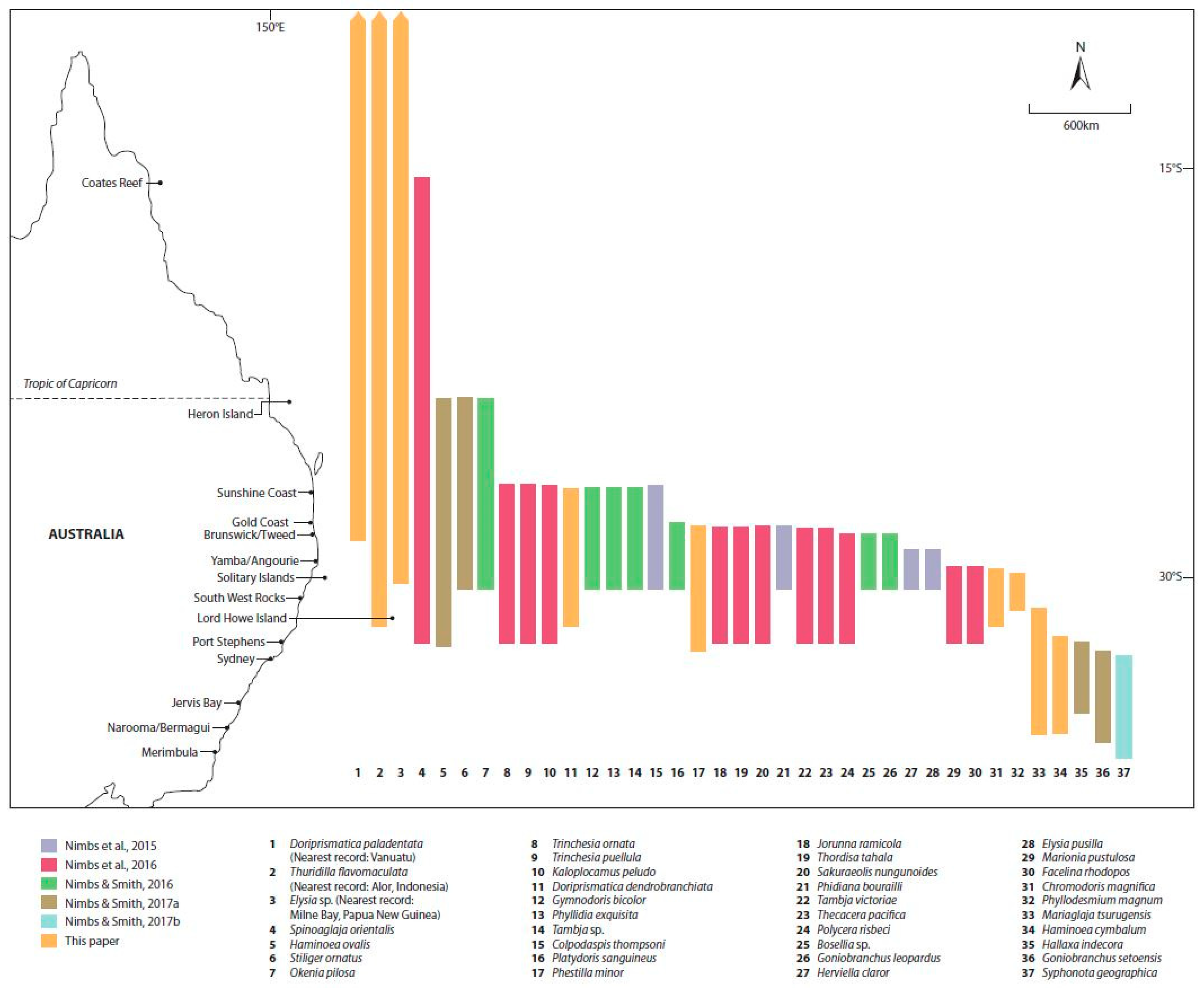
The central eastern Australian coast is a known climate change hot-spot where warming conditions are proving favourable for gradual poleward shifts in southern range limits for several marine taxa (e.g., Harasti; Scott, et al.; Davis [37,38,39]). Since early 2014, we have identified and reported 28 species of sea slug south of their previously known range [5,40,41,42,43] (Figure 1). The distance of such shifts varies from a regional scale at distances of 100s of km to a bioregional scale of >1000 km. The majority of reported range extensions fall within the regional scale: only two of the 28 species were recorded >1000 km poleward of their previous geographical limit.
In this paper, we report nine further range extensions into central and southern NSW and to Lord Howe Island in the central Tasman Sea. The scale of these range shifts varies from <100 km to more than 4000 km and also includes three novel observations for Australian waters. In most cases, these records also represent the southernmost global observations.
2. Materials and Methods
The observations summarised in this paper were made by the authors and well-informed citizen scientists recruited through the Sea Slug Census (SSC) program. The SSC is a volunteer-based project developed by Southern Cross University and volunteers that engages with participants to photographically document sea slug diversity at specific locations (Smith and Davis, unpublished). Observations included here were made during the inaugural Sapphire Coast SSC (April 2017), the 15th Nelson Bay SSC (September 2017) and the second Gold Coast SSC (September 2017).
Sea Slug Census participants are primarily engaged through social media (Facebook https://www.facebook.com/groups/seaslugcensus/) which allows the upload of photos for identification and discussion by other users. Metadata are included for each image, including location, date, and depth. The establishment of a hub for photographic submission and discussion not only provides sharing of recent discoveries, but often uncovers information about the historical presence of species [5]. In this way, previously unpublished observation records for Lord Howe Island were recognised as a result of the development of a social media page for that location. Observations recorded by the authors were made using SCUBA, snorkeling or during searches in intertidal habitats. Photographs were taken using housed Olympus TG-3/5 cameras with iTorch video lights. Voucher specimens were collected in non-sanctuary zones, preserved in 90% ethanol, and deposited at the National Marine Science Centre, Southern Cross University, Coffs Harbour, NSW (NMSC) under NSW Department or Primary Industries permit P14/0014-1.1.
Comparison with published distribution sources and regional lists (e.g., [5]) enables ready detection of species in novel locations. Global distribution data were sourced from the literature and online sources (Table 1). These data include location (latitude and longitude) and date of observation.
Identifications based on photographs alone can be problematic, particularly among groups where there is potential cryptic speciation, such as in the genus Chromodoris [44]. In the absence of genetic analysis, identifications should be regarded as hypotheses. Therefore, we consider our identifications as such and have only included those with which we have reasonable confidence. Additionally, in this paper we use the term ‘range extension′ to mean a potential extension of known distributional range whilst acknowledging that any conclusions are constrained by the distributional data available. For some species, this may be limited to only two or three records.
The familial and generic taxonomy of some species in this paper are in a state of flux at the time of manuscript preparation. Thus, we deferred to the nomenclature listed in World Register of Marine Species [45].
3. Results
Recent Southern Range Shifts in Eastern Australia
- Systematics
- Subclass Heterobranchia
- Order Cephalaspidea Fischer, 1883
- Family Aglajidae Pilsbry, 1895 (1847)
- Genus Mariaglaja Zamora-Silva & Malaquias, 2017
- Mariaglaja tsurugensis (Baba & Abe, 1959) (Figure 2A)
- Synonymy: Chelidonura sandrana Rudman, 1973, Chelidonura babai Gosliner, 1988.
Although highly variable, Mariaglaja tsurugensis is generally characterised by a predominantly black body with yellow spots surrounded by black rings among scattered patches of opaque white. There are, however, occasional specimens that are almost entirely white [60]. The pattern of yellow spots surrounded by white has been variously referred to as ‘floral′ [1,60].
In Australia, M. tsurugensis, has been found on both the east and west coasts (as C. sandrana in Nimbs & Smith [5]), consistent with its broad Indo-Pacific distribution [1,60]. Whilst occasionally abundant on the northern Great Barrier Reef at Lizard Island, QLD [61], it has not been seen elsewhere on the eastern mainland coast except at Angourie (pers. obs.) (specimen collected under permit and deposited at the NMSC [NMSC#0353SDAS]) and the Solitary Islands, NSW [50] (Table 2).
During the inaugural Sapphire Coast SSC on 10 April 2017, a 28-mm long specimen was observed at Blue Pool, Bermagui, NSW. This observation represents a 670 km poleward range extension from its previous southern distribution limit at Lord Howe Island, NSW.
- Family Haminoeidae Pilsbry, 1895
- Genus Haminoea Turton and Kingston [in Carrington], 1830
- Haminoea cymbalum (Quoy and Gaimard, 1832) (Figure 2B)
- Synonymy: Bulla cymbalum Quoy and Gaimard, 1832; Haminea cymbalum (Quoy and Gaimard, 1832); Haminoea simillima Pease, 1868; and, Lamprohaminoea cymbalum (Quoy and Gaimard, 1832) [45].
There are numerous species of Haminoea, many of which are undescribed (M. Malaquias, pers. comm., April 2017). In many cases, distinguishing among species is difficult based on external morphology alone. Indeed, among the European fauna for example, H. orbignyana cannot be reliably identified from its sympatric congeners without examination of the male reproductive system [62]. However, some species do exhibit external features that allow ready identification. Haminoea cymbalum is identified by the presence of orange spots and semi-transparent purple patches on a translucent body. The shell is evident beneath the parapodial lobes and possesses a distinctive ‘fried-egg′ pattern over a darker, often green, background [63].
Haminoea cymbalum has a wide Indo-Pacific distribution, from Mozambique in the west to Japan in the north, to Hawai′i and French Polynesia in the east [59]. On the Australian east coast, it has been found in several locations off Cape York, QLD, throughout the Great Barrier Reef, and south to Port Stephens in central NSW [5] (Table 3). Several specimens were observed on 10 April 2017 at Blue Pool, Bermagui, NSW, by Libby Hepburn and Nick Shaw (Figure 2B). These observations represent a southern range extension of 500 km into the temperate waters of the Tasman Sea from its previous southernmost observation at Port Stephens, NSW.
- Order Sacoglossa
- Family Plakobranchidae Gray, 1840
- Genus Elysia Risso, 1818
- Elysia sp. (Figure 2C)
There are numerous undescribed species in the sacoglossan genus Elysia [1] and, consequently, determining their distributions can be very difficult, particularly as many are morphologically similar or highly cryptic [64]. Nevertheless, some have distinctive, characteristic morphology, and such is the case with Elysia sp. which exhibits a diagnostic mosaic pattern of olive-green patches outlined in black which are replaced with white patches outlined in black along the parapodial margins. No other Elysia is known to exhibit this pattern.
Published records of this species are scarce: indeed, the only existing observation is from Milne Bay in Papua New Guinea [46,47] (Table 4). An observation of a single specimen at North Solitary Island, NSW on 27 May 2017 not only extends the range for this species 2200 km southward into the Tasman Sea, but represents the first Australian, and the most southern, global observation of this species.
- Genus Thuridilla Bergh, 1872
- Thuridilla flavomaculata Gosliner, 1995 (Figure 2D)
The herbivorous Thuridilla flavomaculata is one of the few species in its genus to exhibit scattered yellow spots [65]. An animal collected in the Philippines in 1995 was assigned as the holotype although one paratype was collected in Guam in 1970 [65]. As with many Thuridilla, distribution is generally restricted to the tropics, with observations recorded from the Mariana Islands in the north and Alor, Indonesia in the south. Longitudinally, its range extends from the western Pacific, across the Indian Ocean to Kenya (Table 5).
On 28 May 2017, a single individual was observed by Kathryn May in a tidal pool at Lord Howe Island, NSW. This observation extends the range of this species well outside the tropics, some 4250 km to the southeast of the nearest geographic observation at Alor, East Nusa Tenggara, Indonesia.
- Order Nudibranchia
- Family Chromodorididae Bergh, 1891
- Genus Chromodoris Alder & Hancock, 1855
- Chromodoris magnifica (Quoy & Gaimard, 1832) (Figure 2E)
- Synonym: Doris magnifica Quoy & Gaimard, 1832 [45]
There are several species of Chromodoris that exhibit longitudinal black lines and are difficult to identify based on morphology alone [69]. However, C. magnifica, whilst variable in colour, is readily distinguished by the presence of a submarginal orange band [69]. It has a wide distribution in the Indo-Pacific [51] and, in Australia, has been observed several times on the continental east coast from Fitzroy Island in QLD to Cook Island in northern NSW (Table 6).
Although a single observation at Lord Howe Island in 2009 represents the southernmost global record for this taxon, here we report an observation at North Solitary Island, NSW on 18 May 2017 which extends its continental east coast range by 200 km poleward into the subtropical Solitary Islands Marine Park.
- Genus Doriprismatica d’Orbigny, 1839
- Doriprismatica dendrobranchia (Rudman, 1990) (Figure 2F)
- Synonym: Glossodoris dendrobranchia Rudman, 1990 [45]
Doriprismatica dendrobranchia is a high-bodied, robust nudibranch with a brown body and sinuous, white mantle margin [74]. Whilst similar in form, its colour pattern is almost the complete inverse of its more common, sympatric congener, D. atromarginata.
This species is known only from the waters of southern QLD and was described using one animal found at Horseshoe Reef (Swain Reefs) in the southern Great Barrier Reef [75]. During the 27 years since its original description, there have been only two additional observations recorded, from Noosa, QLD in 2004 and recently in 2018 (Table 7). With no other global records, and a range extending across only 450 km of shallow shelf waters, this species appears to be rare and highly restricted.
A specimen photographed by Jasmin Freya at Lord Howe Island at 16 m on 3 February 2016 represents one of only a handful of observations for this taxon. This important record also extends the known range for this rare species some 820 km south and is the first observation in NSW.
Observations of Doriprismatica paladentata are few, potentially an artefact of its close similarity to Doriprismatica atromarginata [77] which, in some locations, can be one of the most frequently observed chromodorid nudibranchs [78]. Thus, D. paladentata may simply have been overlooked where distributions are sympatric. Nevertheless, it can be distinguished from D. atromarginata by the presence of sparse brown spots on the mantle and a light-blue sub-marginal band [77].
Although the type location is Milne Bay, Papua New Guinea, complete records indicate a western Pacific distribution from the Philippines in the north, to Vietnam in the west and Vanuatu in the southeast (Table 8). An observation at Cook Island, NSW on 25 September 2017 at 7 m by Sascha Hofmann as part of the second Gold Coast SSC, not only represents a range extension of 2000 km from its southernmost global observation at Vanuatu in 1999, but also the first observation for this species in Australian waters.
- Family Facelinidae Bergh, 1889
- Genus Phyllodesmium Ehrenberg, 1831
- Phyllodesmium magnum Rudman, 1991 (Figure 3C)
The nudibranch Phyllodesmium magnum is a large corallivorous aeolid which acquires photosynthetic Symbiodinium from its prey, the octocoral Sinularia [80]. These symbionts are sequestered into tissues giving the body and cerata a brown, blotched appearance. In some specimens, the oral tentacles, rhinophores, and cerata have blue or lilac patches (pers. obs.).
This species has a wide Indo-Pacific distribution from South Africa and the Red Sea [49] to Japan, the Philippines, Marshall Islands, Papua New Guinea, Australia [81,82] and east to the Kermadec Islands, New Zealand [83]. In eastern Australia, it has been found in the central Great Barrier Reef south to the Solitary Islands in northern NSW (Table 9).
Although only 75 km south of its known southern limit on the east coast, an observation of Phyllodesmium magnum by Nicola Fraser at 7 m at Ladies Reef near South West Rocks, NSW on 23 May 2016, is important for several reasons. Firstly, it represents a contemporary observation of this species which is rare in NSW. Secondly, it extends the range south of the large marine protected area of the Solitary Islands Marine Park, and lastly, it illustrates the importance of the nearshore reefs of South West Rocks as habitat for a diverse range of taxa, especially molluscs [84].
- Family Trinchesiidae F. Nordsieck, 1972
- Genus Phestilla Bergh, 1874
- Phestilla minor Rudman, 1981 (Figure 3B)
- Synonym: Tenellia minor (Rudman, 1981) [45]
Few nudibranchs prey on scleractinian corals, however, species in the genus Phestilla do so exclusively. Phestilla minor was so named due its diminutive size in comparison to its congeners. It is aeolidiform with rows of four to six cerata arranged in series that slope to the rear. From above, these appear to project postero-horizontally, giving it a characteristic appearance [85]. The cerata themselves are also distinctive with a sub-apical swelling and ‘terminal bulb’ [85].
This species has a wide distribution from Tanzania in the west to Hawai′i in the east and south into the Great Barrier Reef [51,85]. On the Australian east coast, it has been recorded from Lizard Island in northern QLD south to Hastings Point in northern NSW (Table 10).
An observation of a single specimen at Nelson Bay, NSW during the 15th Nelson Bay SSC by Peter Davey extends this range south by 500 km.
4. Discussion
The addition of nine species of heterobranch sea slug that have extended their range southward into subtropical and warm-temperate waters in eastern Australia brings the total, poleward-shifting tally to 37. While there is little doubt that a lack of search effort at some locations, and the cryptic nature of some species, may account for a few of these records [5], by far the majority are for conspicuous species which occur in accessible habitats. This strongly suggests that they represent real shifts in species distribution patterns. Sporadic finds of tropically-affiliated species at southern range extremes is well documented for highly-collectable species with a long historical record of collection or observation (e.g., the Cypraeidae [87]). However, there is also increasing evidence that some species are now regularly found at their new southern limit, suggesting the establishment of populations rather than sporadic recruitment of individuals. For example, since our first observation of a single Tambja sp. at Split Solitary Island in 2016 [42], specimens have been regularly observed at nearby North West Solitary Island (2017, 2018). This is consistent with recent work in temperate waters that modelled the distribution of a number of range-shifting species and suggested not only poleward extension of records, but also a poleward shift in the species′ core range [88].
Of the five major benthic heterobranch orders found in Australian waters [89], four are represented within this list of 37 species: Aplysiida (1), Cephalaspidea (5), Nudibranchia (26), and the Sacoglossa (5). The Pleurobranchida is the only major order that lacks taxa known to have recently shifted poleward in southeastern Australia. Within the numerically largest group, the Nudibranchia, animals from both the major suborders Cladobranchia and Doridina are present, with 9 and 17 species, respectively. This broad taxonomic spread illustrates that range shifts are occurring across much of the Heterobranchia and are not confined to groups that contain potentially invasive taxa, such as the aeolids [28,30], polycerids [90,91], and sea hares [35,40].
The pace of southern migration is difficult to ascertain as there was little consistent, dedicated observation work on the NSW coast prior to 2010. Since the advent of the Nelson Bay SSC in 2013, concentrated research efforts on the north coast, and the phased introduction of regular SSC surveys at other locations, there have been at least five records of range extensions per year since 2014. The highest count for a single year was 13 in 2015 (Table 11).
There is also the suggestion of a seasonal pattern in observations of taxa south of their known range. Thus, most observations were noted in summer (n = 15) with fewer in spring and autumn, and none during winter (Figure 4). This may be partly attributed to the timing of SSC events which mostly occur between September and March, the period of maximum sea slug diversity and abundance (unpublished data). The Nelson Bay SSC program is the only one to incorporate observations in all seasons, but participation rates in winter are usually lower (~40%) than during other seasons. These winter censuses also yielded lower sea slug diversity. Nevertheless, the authors undertake regular diving activities throughout the year and are yet to find new southern records during winter.
The importance of citizen scientist efforts are further evidenced by the number of significant records generated. Of the 37 species with poleward range shifts, 13 were observed by this group with 4 of those observations made during census events. The remaining nine were generated either in the lead-up to censuses or immediately afterwards.
Gathering evidence of climate-related changes in the distribution of species and the ecology of marine habitats is problematic for many reasons [92]. Perhaps the most important relate to the likely slow pace of change in physico-chemical variables, the enormity of the task of keeping track of the distribution of the broader species pool, and predicting which ecological processes to monitor. While engagement with citizen scientists can vastly increase the spatial and temporal scale of efforts to document species distribution, it will arguably be impossible to monitor everything and so a surrogacy approach (e.g., [93,94,95]), targeting a group of organisms that are likely to respond in a way that reflects broader ecosystem change, is clearly needed. There are some clear candidates.
Corals are under threat from changing environmental conditions with increasing frequency of bleaching events and subsequent mortality [96]. As habitat formers, changes in coral communities are also likely to have considerable flow-on effects to broader biotic communities. However, while corals are expected to change their distribution patterns, with subtropical reefs forecast to act as refuges for species driven southward under thermal pressure [97], they will remain a feature only of warmer oceans.
Fishes are another likely candidate with considerable evidence of changing distribution patterns [16,88,98]. The importance of fishes as consumers and as key economic resources further underpin their potential. As some families are excellent proxies of broader assemblage patterns, it may be possible to increase the cost-effectiveness of monitoring by focusing primarily on these taxa (pomacentrids and labrids) [99].
While these taxa are undoubtedly useful targets for monitoring, effort is likely to be restricted by the availability of funding and expertise. Corals are notoriously difficult to identify [100] and, while there are some very successful citizen science programs focusing on fish among other taxa (Reef Life Survey, RedMap), access and volunteer capacity can restrict the acquisition of data [88].
In this study, we document the first observations of taxa at novel locations. This provides some focus for efforts to determine the establishment of populations which may herald ecological impacts in the receiving assemblages. For example, regular monitoring enabled Goddard, Gosliner, & Pearse [30] to recognise an increase in abundance of Phidiana hiltoni and the simultaneous collapse of other nudibranch populations in California. Whilst such a dramatic effect has yet to be observed on the Australian east coast, the acquisition of data through regular censuses increases the probability that large or unexpected changes in populations will be rapidly detected.
Through the Sea Slug Census program, we have not only been able to demonstrate the appeal of sea slugs as a target taxon, but also the ability of volunteers to find and record them photographically. Because they are generally very slow moving, photographic records are easy to generate, which facilitates capture of images that allow ready identification of species by scientists. However, apart from their popularity with divers and rock-pool ramblers, there are other reasons to suggest sea slugs as useful surrogates for monitoring ecosystem change. Most have short life cycles and the presence of species is strongly dependent on the availability of specific food sources. This means that populations, and whole assemblages, are likely to respond rapidly to changes in the environment. While this means they may be sensitive to a range of variable factors, increasing knowledge about natural patterns of variation [101,102] will help to distinguish additional effects of changing climate. The data presented here provide ample evidence that distribution patterns are changing rapidly, and that tropical species are able to recruit to warm temperate waters across the continuum of the Australian east coast. This supports a continuation of research effort, supported by volunteers, to further develop their use as an indicator of environmental change.
Author Contributions
M.J.N. and S.D.A.S. conceived the work; M.J.N. and S.D.A.S. carried out the field work; M.J.N. and S.D.A.S. sourced, analysed, and summarised the data; M.J.N. and S.D.A.S. wrote, edited, and reviewed the paper.
Funding
The Sea Slug Census program has been supported by a range of sponsors at different locations and through small grant funding from Hunter Local Land Services. Additional support was provided by the National Marine Science Centre, and the Marine Ecology Research Centre, Southern Cross University, NSW, Australia.
Acknowledgments
The authors are grateful to Libby Hepburn and Nick Shaw for data and images of H. cymbalum and M. tsurugensis taken during the inaugural Sapphire Coast Sea Slug Census, Sascha Hoffman for information and images of D. paladentata taken during the second Gold Coast Sea Slug Census, Peter Davey for data and images of T. minor taken during the 15th Nelson Bay Sea Slug Census, Jasmin Freya for data and images of D. dendrobranchiata taken at Lord Howe Island, and Kathryn May for data and images of T. flavomacula taken at Lord Howe Island. We also extend our thanks to Richard Willan from the Museum and Art Gallery of the Northern Territory for assistance with identifications of species, and four anonymous reviewers for constructive feedback that resulted in improvements to this paper. Kathryn James produced Figure 1.
Conflicts of Interest
The authors declare no conflict of interest. The sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gosliner, T.M.; Valdés, Á.; Behrens, D.W. Nudibranch and Sea Slug Identification: Indo-Pacific; New World Publications: Jacksonville, FL, USA, 2015. [Google Scholar]
- Kaligis, F.; Eisenbarth, J.H.; Schillo, D.; Dialao, J.; Schäberle, T.F.; Böhringer, N.; Bara, R.; Reumschüssel, S.; König, G.M.; Wägele, H. Second survey of heterobranch sea slugs (Mollusca, Gastropoda, Heterobranchia) from Bunaken National Park, North Sulawesi, Indonesia—How much do we know after 12 years? Mar. Biodivers. Rec. 2018, 11, 2. [Google Scholar] [CrossRef]
- Rudman, W.; Willan, R. Fauna of Australia: Opisthobranchia. In Mollusca: The Southern Synthesis; CSIRO: Melbourne, Australia, 1998. [Google Scholar]
- Burn, R. A checklist and bibliography of the Opisthobranchia (Mollusca: Gastropoda) of Victoria and the Bass Strait area, southeastern Australia. Mus. Vic. Sci. Rep. 2006, 10, 7–13. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Smith, S.D.A. An illustrated inventory of the sea slugs of New South Wales, Australia (Gastropoda: Heterobranchia). Proc. R. Soc. Vic. 2017, 128, 44–113. [Google Scholar] [CrossRef]
- Ridgway, K.R. Long-term trend and decadal variability of the southward penetration of the East Australian Current. Geophys. Res. Lett. 2007, 34, 13. [Google Scholar] [CrossRef]
- Suthers, I.M.; Young, J.W.; Baird, M.E.; Roughan, M.; Everett, J.D.; Brassington, G.B.; Byrne, M.; Condie, S.A.; Hartog, J.R.; Hassler, C.S. The strengthening East Australian Current, its eddies and biological effects—An introduction and overview. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 538–546. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Davies, P.L.; Jordan, A.; Smith, S.D.A. Variation in sea temperature and the East Australian Current in the Solitary Islands region between 2001–2008. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 616–627. [Google Scholar] [CrossRef]
- Denham, R.N.; Crook, F.G. The Tasman front. N. Z. J. Mar. Freshw. 1976, 10, 15–30. [Google Scholar] [CrossRef]
- Roughan, M.; Middleton, J.H. A comparison of observed upwelling mechanisms off the east coast of Australia. Cont. Shelf Res. 2002, 22, 2551–2572. [Google Scholar] [CrossRef] [Green Version]
- Harriott, V.J.; Smith, S.D.A.; Harrison, P.L. Patterns of coral community structure of subtropical reefs in the Solitary-Islands Marine Reserve, Eastern Australia. Mar. Ecol. Prog. Ser. 1994, 109, 67–76. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Jordan, A.; Smith, S.D.A. Biogeographical and cross-shelf patterns of reef fish assemblages in a transition zone. Mar. Biodivers. 2010, 40, 181–193. [Google Scholar] [CrossRef]
- Smith, S.D.A. Growth and population dynamics of the giant clam Tridacna maxima (Röding) at its southern limit of distribution in coastal, subtropical eastern Australia. Molluscan Res. 2011, 31, 37–41. [Google Scholar]
- Harrison, M.A.; Smith, S.D.A. Cross-shelf variation in the structure of molluscan assemblages on shallow, rocky reefs in subtropical, eastern Australia. Mar. Biodivers. 2012, 42, 203–216. [Google Scholar] [CrossRef]
- Booth, D.J.; Figueira, W.F.; Gregson, M.A.; Brown, L.; Beretta, G. Occurrence of tropical fishes in temperate southeastern Australia: Role of the East Australian Current. Estuar. Coast. Shelf Sci. 2007, 72, 102–114. [Google Scholar] [CrossRef]
- Figueira, W.F.; Booth, D.J. Increasing ocean temperatures allow tropical fishes to survive overwinter in temperate waters. Glob. Chang. Biol. 2010, 16, 506–516. [Google Scholar] [CrossRef]
- Fowler, A.M.; Parkinson, K.; Booth, D.J. New poleward observations of 30 tropical reef fishes in temperate southeastern Australia. Mar. Biodives. 2017, 1–6. [Google Scholar] [CrossRef]
- Padula, V.; Bahia, J.; Vargas, C.; Lindner, A. Mollusca, Nudibranchia: New records and southward range extensions in Santa Catarina, southern Brazil. Check List 2011, 7, 806–808. [Google Scholar] [CrossRef]
- Tibirica, Y.; Pola, M.; Cervera, J.L. Astonishing diversity revealed: An annotated and illustrated inventory of Nudipleura (Gastropoda: Heterobranchia) from Mozambique. Zootaxa 2017, 4359, 1–133. [Google Scholar] [CrossRef] [PubMed]
- Richards, Z.; Kirkendale, L.; Moore, G.; Hosie, A.; Huisman, J.; Bryce, M.; Marsh, L.; Bryce, C.; Hara, A.; Wilson, N. Marine biodiversity in temperate Western Australia: Multi-taxon surveys of Minden and Roe reefs. Diversity 2016, 8, 7. [Google Scholar] [CrossRef]
- Bertsch, H. Biodiversity in La Reserva de la Biósfera Bahía de los Ángeles y Canales de Ballenas y Salsipuedes: naming of a new genus, range extensions and new records, and species list of Heterobranchia (Mollusca: Gastropoda), with comments on biodiversity conservation. Festivus 2014, 46, 158–177. [Google Scholar]
- Schrödl, M.; Hooker, Y. Sea slugs of Peru: Peruvian-Chilean faunal elements. Spixiana 2014, 37, 45–59. [Google Scholar]
- Chichvarkhin, A.Y.; Chichvarkhina, O.V.; Kartavtsev, Y.P. Janolus fuscus O’Donoghue, 1924 (Gastropoda: Proctonotidae), a species of a nudibranch family that is new to the marine fauna of Russia. Russ. J. Mar. Biol. 2016, 42, 20–25. [Google Scholar] [CrossRef]
- Whittaker, D.E. On the occurrence of the nudibranch Geitodoris planata in the North Sea and its addition to the Yorkshire marine fauna, with a note on recent new species. Naturalist 2013, 138, 82. [Google Scholar]
- Uribe, R.A.; Pacheco, A.S. First record of Spurilla neapolitana (Mollusca: Nudibranchia: Aeolidiidae) on the central coast of Peru (Humboldt Current Upwelling Ecosystem). Mar. Biodivers. Rec. 2012, 5. [Google Scholar] [CrossRef]
- Valdés, Á.; Bertsch, H. Redescription and range extension of Bathydoris aioca Marcus, & Marcus, 1962 (Nudibranchia: Gnathodoridoidea). Veliger-Berkeley 2000, 43, 172–178. [Google Scholar]
- Goddard, J.H.R.; Treneman, N.; Pence, W.E.; Mason, D.E.; Dobry, P.M.; Green, B.; Hoover, C. Nudibranch range shifts associated with the 2014 warm anomaly in the Northeast Pacific. Bull. South. Calif. Acad. Sci. 2016, 115, 15–40. [Google Scholar] [CrossRef]
- Goddard, J.H.R.; Schaefer, M.C.; Hoover, C.; Valdés, Á. Regional extinction of a conspicuous dorid nudibranch (Mollusca: Gastropoda) in California. Mar. Biol. 2013, 160, 1497–1510. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, J.H.R.; Gosliner, T.M.; Pearse, J.S. Impacts associated with the recent range shift of the aeolid nudibranch Phidiana hiltoni (Mollusca, Opisthobranchia) in California. Mar. Biol. 2011, 158, 1095–1109. [Google Scholar] [CrossRef] [PubMed]
- Bridle, T. Spurilla braziliana—A new sea slug in South Australia. South Aust. Nat. 2017, 91, 29–33. [Google Scholar]
- Por, F.D. Lessepsian Migration: The Influx of Red Sea Biota into the Mediterranean by Way of the Suez Canal; Springer Science & Business Media: Berlin, Germany, 2012; Volume 23. [Google Scholar]
- Çevik, C.E.M.; Özturk, B. A new Lessepsian mollusc Hypselodoris infucata (Ruppell and Leuckart, 1828) (Gastropoda: Nudibranchia) for the coasts of Turkey. Turk. J. Zool. 2000, 25, 27–30. [Google Scholar]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Moles, J.; Mas, G.; Figueroa, I.; Fernández-Vilert, R.; Salvador, X.; Giménez, J. As fast as a hare: Colonization of the heterobranch Aplysia dactylomela (Mollusca: Gastropoda: Anaspidea) into the western Mediterranean Sea. Cah. Biol. Mar. 2017, 2017 58, 341–345. [Google Scholar]
- Gavagnin, M.; Carbone, M.; Nappo, M.; Mollo, E.; Roussis, V.; Cimino, G. First chemical study of anaspidean Syphonota geographica: Structure of degraded sterols aplykurodinone-1 and -2. Tetrahedron 2005, 61, 617–621. [Google Scholar] [CrossRef]
- Harasti, D. Range extension and first occurrence of the thorny seahorse Hippocampus histrix in New South Wales, Australia. Mar. Biodivers. Rec. 2015, 8. [Google Scholar] [CrossRef]
- Scott, A.; Harasti, D.; Davis, T.; Smith, S.D.A. Southernmost records of the host sea anemone, Stichodactyla haddoni, and associated commensal shrimps in a climate change hotspot. Mar. Biodivers. 2015, 45. [Google Scholar] [CrossRef]
- Davis, T.R. First records of three fishes, and southern records of a further four fishes, from New South Wales, Australia. Check List 2016, 12, 2008. [Google Scholar] [CrossRef] [Green Version]
- Nimbs, M.J.; Smith, S.D.A. Revision of the southern distribution limit for the tropical marine herbivore Syphonota geographica (A. Adams & Reeve, 1850) (Heterobranchia: Aplysiidae) in a global climate change hot-spot. Aust. Zool. 2017, 38. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Willan, R.C.; Smith, S.D.A. Range extensions for heterobranch sea slugs (formerly opisthobranch) belonging to the families Diaphanidae, Plakobranchidae and Facelinidae on the Eastern Coast of Australia. Mar. Biodivers. Rec. 2015, 8, e76. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Smith, S.D.A. Welcome strangers: Southern range extensions for seven heterobranch sea slugs (Mollusca: Gastropoda) on the subtropical east Australian coast, a climate change hot spot. Reg. Stud. Mar. Sci. 2016, 8, 27–32. [Google Scholar] [CrossRef]
- Nimbs, M.J.; Willan, R.C.; Larkin, M.; Davis, T.R.; Smith, S.D.A. Southern range extensions for twelve heterobranch sea slugs (Gastropoda: Heterobranchia) on the eastern coast of Australia. Mar. Biodivers. Rec. 2016, 9. [Google Scholar] [CrossRef]
- Layton, K.K.; Gosliner, T.M.; Wilson, N.G. Flexible colour patterns obscure identification and mimicry in Indo-Pacific Chromodoris nudibranchs (Gastropoda: Chromodorididae). Mol. Phylogenet. Evol. 2018, 124, 27–36. [Google Scholar] [CrossRef] [PubMed]
- WoRMS Editorial Board. World Register of Marine Species. 2018. Available online: http://www.marinespecies.org at VLIZ (accessed on 4 August 2018).
- Coleman, N. Nudibranchs Encyclopedia; Neville Coleman’s Underwater Geographic Pty, National Library of Australia: Springwood, Australia, 2008; ISBN 9780947325411. [Google Scholar]
- Coleman, N. Nudibranchs Encyclopedia: Catalogue of Asia/Indo Pacific Sea Slugs, 2nd ed.; Neville Coleman’s Underwater Geographic: Springwood, Australia, 2008. [Google Scholar]
- Cobb, G.; Willan, R.C. Undersea Jewels: A Colour Guide to Nudibranchs; ABRS: Canberra, Australia, 2006. [Google Scholar]
- Debelius, H.; Kuiter, R.H. Nudibranchs of the World; Ikan-Unterwasserarchiv: Frankfurt, Germany, 2007; ISBN 3939767069. [Google Scholar]
- Buchanan, C. Species List and Semi-Quantitative Abundance from Buchanan (1989) A Reference List of Opisthobranch Molluscs from the Solitary Islands and Adjacent Coast; Solitary Islands Underwater Research Group: Coffs Harbour Jetty, Australia, 1989. [Google Scholar]
- Pixel, N. Available online: http://www.nudipixel.net (accessed on 3 October 2017).
- Atlas of Living Australia. 2015. Available online: http://www.ala.org.au (accessed on 31 March 2017).
- Rudman, W.B. Sea Slug Forum. Available online: http://www.seaslugforum.net/find/chelfulv (accessed on 6 October 2017).
- Ocean Biogeographic Information System (OBIS). Intergovernmental Oceanographic Commission of UNESCO. Available online: http://www.iobis.org (accessed on 3 October 2017).
- Cobb, G.; Mullins, D. Nudibranchs: Sunshine Coast QLD & Tasmania Australia—Thordisa tahala. Available online: http://www.nudibranch.com.au/pages/CapThorClown-044w.htm (accessed on 7 October 2017).
- Köhler, E. “Doriprismatica” paladentata (Rudman, 1986). Available online: https://www.philippine-sea-slugs.com/Nudibranchia/Euctenidiacea/Doriprismatica_paladentata_02.htm (accessed on 6 October 2017).
- Clarke, L.; Buckland, P. Julian Rocks.net. Available online: http://www.julianrocks.net/index.php (accessed on 19 October 2017).
- Riek, D. Sea Slugs and other Marine Invertebrates of the Tweed-Byron Coast, Australia. Available online: http://www.roboastra.com (accessed on 17 October 2017).
- GBIF.org. GBIF Home Page. Available online: http://www.gbif.org (accessed on 20 April 2017).
- Rudman, W.B. Chelidonura sandrana. Available online: http://www.seaslugforum.net/find/chelsand (accessed on 20 April 2017).
- Rudman, W.B. Re: Chelidonura sandrana—Feeding. Available online: http://www.seaslugforum.net/find/8940 (accessed on 20 April 2017).
- Malaquias, M.A.E.; Cervera, J.L. The genus Haminoea (Gastropoda: Cephalaspidea) in Portugal, with a review of the European species. J. Molluscan Stud. 2006, 72, 89–103. [Google Scholar] [CrossRef]
- Rudman, W.B. Haminoea cymbalum in Southern Queensland. Available online: http://www.seaslugforum.net (accessed on 20 April 2017).
- Carmona, L.; Malaquias, M.A.E.; Gosliner, T.M.; Pola, M.; Cervera, J.L. Amphi-Atlantic distributions and cryptic species in sacoglossan sea slugs. J. Molluscan Stud. 2011, 77, 401–412. [Google Scholar] [CrossRef]
- Gosliner, T.M. The genus Thuridilla (Opisthobranchia: Elysiidae) from the tropical Indo-Pacific, with a revision of the phylogeny and systematics of the Elysiidae. Proc. Calif. Acad. Sci. 1995, 49, 1–54. [Google Scholar]
- Urasawa, T. Thuridilla flavomaculata from the Mariana Islands. Sea Slug Forum. Available online: http://www.seaslugforum.net/showall/thurflav (accessed on 3 October 2017).
- Bala, K. Thuridilla flavomaculata from Bunaken, Indonesia. Available online: http://www.seaslugforum.net/find/15555 (accessed on 3 October 2017).
- Warren, L. Thuridilla flavomaculata from Sulawesi. Available online: http://www.seaslugforum.net/find/2804 (accessed on 3 October 2017).
- Rudman, W.B. Chromodoris magnifica (Quoy & Gaimard, 1832). Available online: http://www.seaslugforum.net/showall/chrmag (accessed on 3 October 2017).
- Bryce, C. Chromodoris magnifica from W. Australia. Available online: http://www.seaslugforum.net/find/805 (accessed on 3 October 2017).
- Reef Life Survey. Available online: http://reeflifesurvey.com (accessed on 23 August 2018).
- Marshall, J.G.; Willan, R.C. Nudibranchs of Heron Island, Great Barrier Reef: A Survey of the Opisthobranchia (Sea Slugs) of Heron and Wistari Reefs; Backhuys: Leiden, The Netherlands, 1999; ISBN 9057820331. [Google Scholar]
- Atlas of Living Australia. Human observation of Chromodoris magnifica Recorded on 2013-06-16. Available online: https://biocache.ala.org.au/occurrences/defbe9d3-3638-49b8-a699-b4f8ea5abb11 (accessed on 3 October 2017).
- Rudman, W.B. Glossodoris dendrobranchia Rudman. 1990. Available online: http://www.seaslugforum.net/factsheet/glosdend (accessed on 5 October 2017).
- Rudman, W.B. The Chromodorididae (Opisthobranchia: Mollusca) of the Indo-West Pacific: further species of Glossodoris, Thorunna and the Chromodoris aureomarginata colour group. Zool. J. Linn. Soc. 1990, 100, 263–326. [Google Scholar] [CrossRef]
- Cobb, G. Glossodoris dendrobranchia in sthn Queensland. Available online: http://www.seaslugforum.net/find/12235 (accessed on 5 October 2017).
- Rudman, W.B. Glossodoris paladentatus Rudman. 1986. Available online: http://www.seaslugforum.net/showall/glospala (accessed on 6 October 2017).
- Rudman, W.B. Glossodoris atromarginata. Available online: http://www.seaslugforum.net/find/glosatro (accessed on 6 October 2017).
- Rudman, W.B. The Chromodorididae (Opisthobranchia: Mollusca) of the Indo-West Pacific: The genus Glossodoris Ehrenbergh (=Casella, H. & A. Adams). Zool. J. Linn. Soc. 1986, 86, 101–184. [Google Scholar]
- Rudman, W.B. Phyllodesmium magnum Rudman. 1991. Available online: http://www.seaslugforum.net/showall/phylmagn (accessed on 7 October 2017).
- Rudman, W.B. The anatomy and biology of alcyonarian-feeding aeolid opisthobranch molluscs and their development of symbiosis with zooxanthellae. Zool. J. Linn. Soc. 1981, 72, 219–262. [Google Scholar] [CrossRef] [Green Version]
- Rudman, W.B. Further studies on the taxonomy and biology of the octocoral-feeding genus Phyllodesmium Ehrenberg, 1831 (Nudibranchia: Aeolidoidea). J. Molluscan Stud. 1991, 57, 167–203. [Google Scholar] [CrossRef]
- Brook, F.J.; Marshall, B.A. Checklist of benthic coastal marine Mollusca from the northern Kermadec Islands. Appendix in: Brook, F.J., The coastal molluscan fauna of northern Kermadec Islands, Southwest Pacific Ocean. J. R. Soc. New Zeal. 1998, 28, 210–233. [Google Scholar] [CrossRef]
- Smith, S.D.A.; Rule, M.J.; Harrison, M.; Dalton, S.J. Monitoring the sea change: Preliminary assessment of the conservation value of nearshore reefs, and existing impacts, in a high-growth, coastal region of subtropical eastern Australia. Mar. Poll. Bull. 2008, 56, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Rudman, W.B. Further studies on the anatomy and ecology of opisthobranch molluscs feeding on the scleractinian coral Porites. Zool. J. Linn. Soc. 1981, 71, 373–412. [Google Scholar] [CrossRef]
- Rudman, W.B. Sea Slug Forum: Phestilla minor. Available online: http://www.seaslugforum.net/find/phesmino (accessed on 7 October 2017).
- Tarrant, D. Cowries of New South Wales; Regal Printing: Hong Kong, China, 2016. [Google Scholar]
- Champion, C.; Hobday, A.J.; Tracey, S.R.; Pecl, G.T. Rapid shifts in distribution and high-latitude persistence of oceanographic habitat revealed using citizen science data from a climate change hotspot. Glob. Chang. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nimbs, M.J. NudiKey: An illustrated, interactive identification key to the families of Australian heterobranch sea-slugs (Mollusca: Gastropoda). Aust. Zool. 2017, 38, 537–546. [Google Scholar] [CrossRef]
- Giacobbe, S.; De Matteo, S. The potentially invasive opisthobranch Polycera hedgpethi Er. Marcus, 1964 (Gastropoda Nudibranchia), introduced in a Mediterranean coastal lagoon. Biodivers. J. 2013, 4, 359–364. [Google Scholar]
- Wilson, N.G. New record of the nudibranch Polycera hedgpethi Er. Marcus, 1964, in South Australia, with a discussion on its occurrence in Australia. Rec. West. Aust. Mus. 2006, 140, 137–140. [Google Scholar] [CrossRef]
- Przeslawski, R.; Ahyong, S.; Byrne, M.; Woerheide, G.; Hutchings, P.A.T. Beyond corals and fish: The effects of climate change on noncoral benthic invertebrates of tropical reefs. Glob. Chang. Biol. 2008, 14, 2773–2795. [Google Scholar] [CrossRef]
- Balmford, A.; Green, M.J.B.; Murray, M.G. Using higher-taxon richness as a surrogate for species richness: I. Regional tests. Proc. R. Soc. Lond. B 1996, 263, 1267–1274. [Google Scholar] [CrossRef]
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Smith, S.D.A. Rapid assessment of invertebrate biodiversity on rocky shores: Where there’s a whelk there’s a way. Biodivers. Conserv. 2005, 14, 3565–3576. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373. [Google Scholar] [CrossRef] [PubMed]
- Beger, M.; Sommer, B.; Harrison, P.L.; Smith, S.D.A.; Pandolfi, J.M. Conserving potential coral reef refuges at high latitudes. Divers. Distrib. 2014, 20, 245–257. [Google Scholar] [CrossRef]
- Last, P.R.; White, W.T.; Gledhill, D.C.; Hobday, A.J.; Brown, R.; Edgar, G.J.; Pecl, G. Long-term shifts in abundance and distribution of a temperate fish fauna: a response to climate change and fishing practices. Glob. Ecol. Biogeogr. 2011, 20, 58–72. [Google Scholar] [CrossRef]
- Malcolm, H.A.; Smith, S.D.A. Objective selection of surrogate families to describe reef fish assemblages in a subtropical marine park. Biodivers. Conserv. 2010, 19, 3611–3618. [Google Scholar] [CrossRef]
- Veron, J.E.N. Corals in Space and Time: The Biogeography and Evolution of the Scleractinia; Cornell University Press: New York, NY, USA, 1995; ISBN 0801482631. [Google Scholar]
- Smith, S.D.A.; Nimbs, M.J. Quantifying temporal variation in heterobranch (Mollusca: Gastropoda) sea slug assemblages: Tests of alternate models. Molluscan Res. 2017, 37. [Google Scholar] [CrossRef]
- Larkin, M.; Smith, S.D.A.; Willan, R.C.; Davis, T.R. Diel and seasonal variation in heterobranch sea slug assemblages within an embayment in temperate eastern Australia. Mar. Biodivers. 2017. [Google Scholar] [CrossRef]
Figure 1.
Summary of recent southward range extensions for sea slugs in southeastern Australia.

Figure 2.
(A) Mariaglaja tsurugensis, Angourie Pool, NSW, 21 September 2017. Photo: S.D.A. Smith. (B) Haminoea cymbalum at Blue Pool, Bermagui, NSW on 10 March 2017. Photo: N. Shaw. (C) First Australian observation of Elysia sp. at North Solitary Island, NSW, 27 May 2017 Photo: M. Nimbs. (D) Thuridilla flavomaculata at Lord Howe Island, NSW, 28 May 2017. Photo: K. May. (E) Chromodoris magnifica at North Solitary Island, NSW, 18 May 2017. Photo: M. Nimbs. (F) Southernmost record of the Australian east coast endemic Doriprismatica dendrobranchia, Lord Howe Island, NSW, 15 January 2017. Photo: J. Freya.
Figure 2.
(A) Mariaglaja tsurugensis, Angourie Pool, NSW, 21 September 2017. Photo: S.D.A. Smith. (B) Haminoea cymbalum at Blue Pool, Bermagui, NSW on 10 March 2017. Photo: N. Shaw. (C) First Australian observation of Elysia sp. at North Solitary Island, NSW, 27 May 2017 Photo: M. Nimbs. (D) Thuridilla flavomaculata at Lord Howe Island, NSW, 28 May 2017. Photo: K. May. (E) Chromodoris magnifica at North Solitary Island, NSW, 18 May 2017. Photo: M. Nimbs. (F) Southernmost record of the Australian east coast endemic Doriprismatica dendrobranchia, Lord Howe Island, NSW, 15 January 2017. Photo: J. Freya.

Figure 3.
(A) Doriprismatica paladentata Cook Island, NSW, 24 September 2017. Photo: S. Hofmann. (B) Phestilla minor, Nelson Bay, NSW, 9 September 2017. Photo: P. Davey. (C) Phyllodesmium magnum at the southern limit of its range, South-West Rocks, NSW, 23 May 2016. Photo: M. Nimbs.
Figure 3.
(A) Doriprismatica paladentata Cook Island, NSW, 24 September 2017. Photo: S. Hofmann. (B) Phestilla minor, Nelson Bay, NSW, 9 September 2017. Photo: P. Davey. (C) Phyllodesmium magnum at the southern limit of its range, South-West Rocks, NSW, 23 May 2016. Photo: M. Nimbs.

Figure 4.
Number of published observations that represent range extension records by season (austral) from 2010 to 2017 (n = 37).
Figure 4.
Number of published observations that represent range extension records by season (austral) from 2010 to 2017 (n = 37).
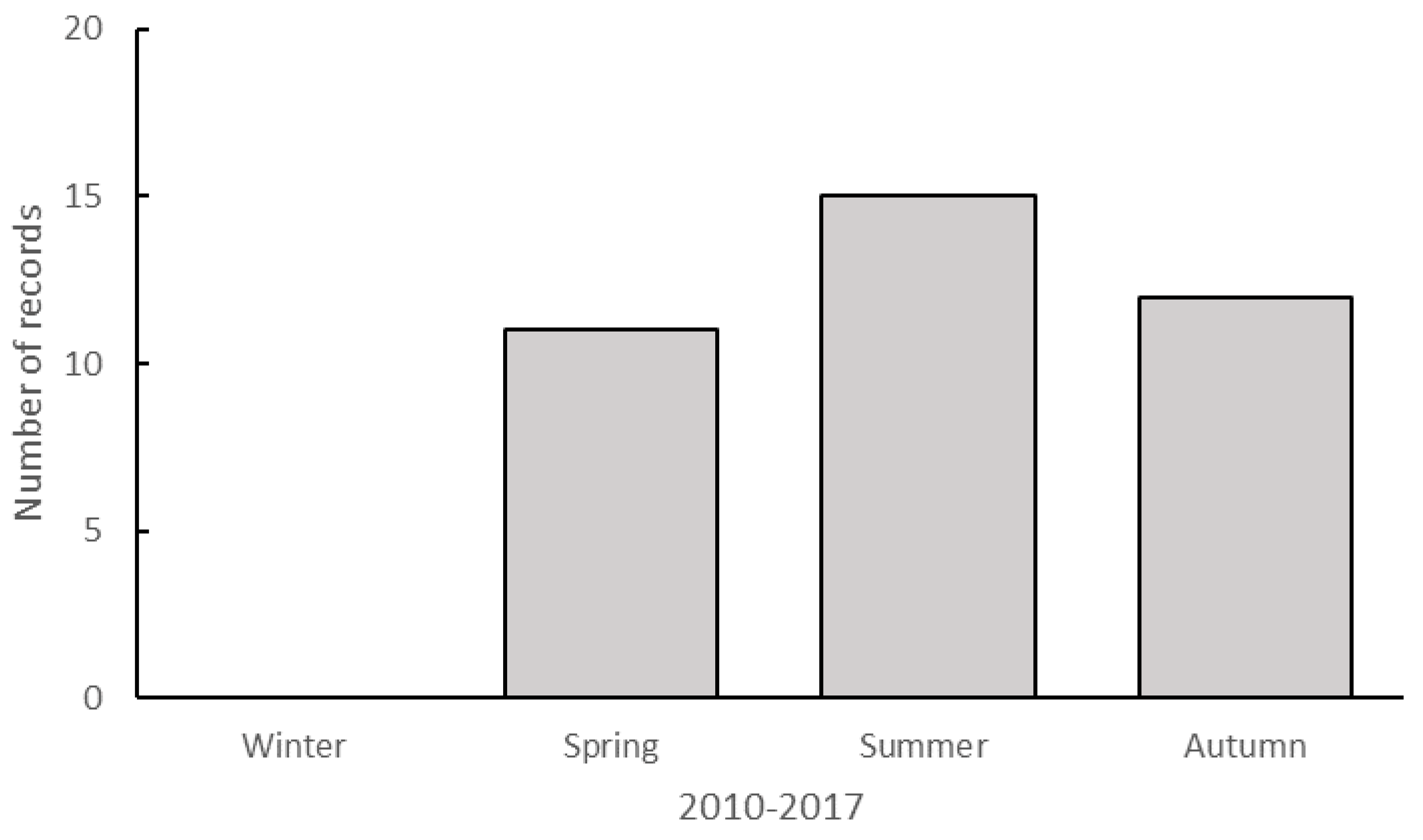

{kind=link}
{kind=link}
{kind=link}
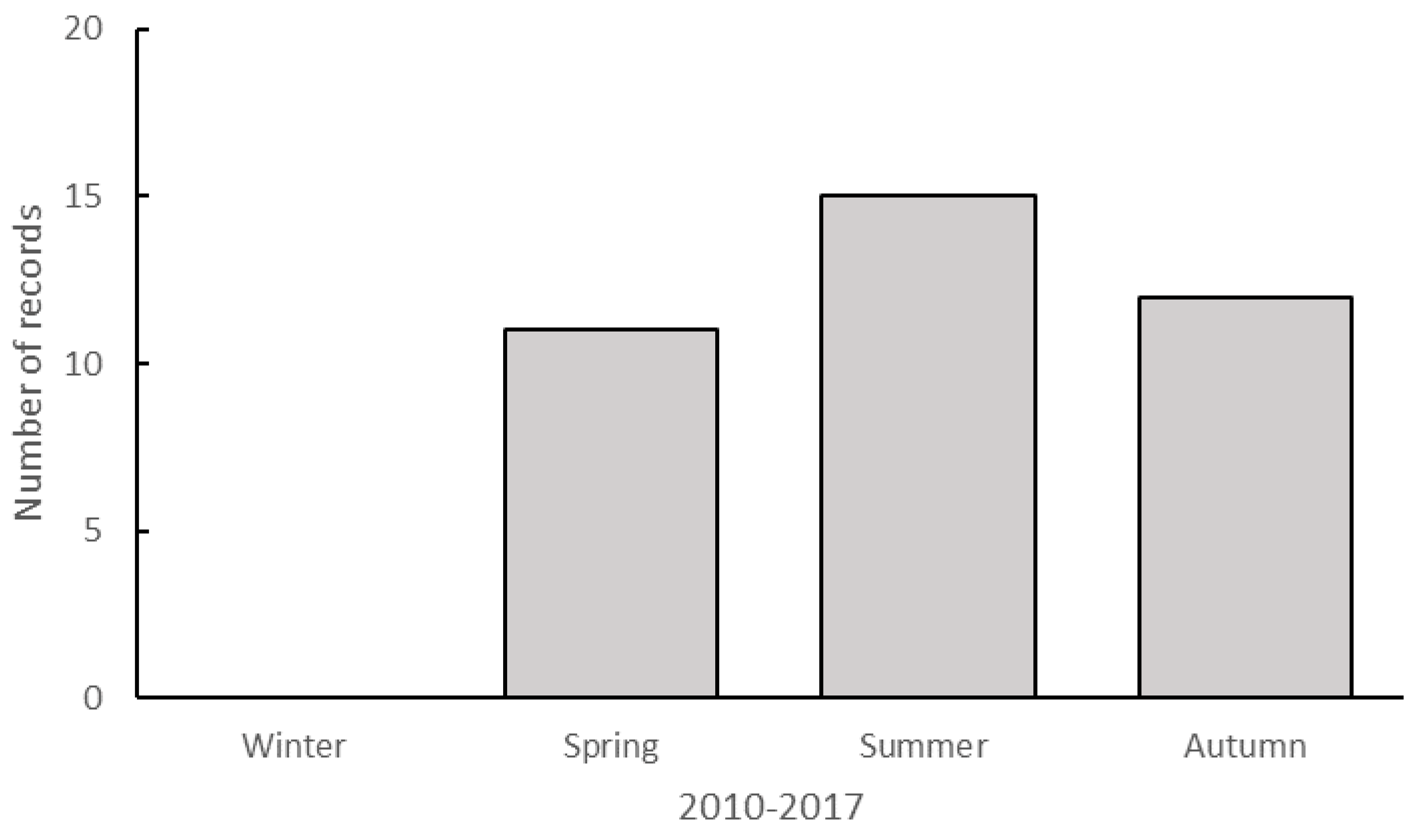{kind=link}
Table 1.
Summary of source documents for global distribution data.
| Source Type | Reference |
|---|---|
| Published literature | Coleman [46,47] |
| Nimbs & Smith [5] | |
| Gosliner et al. [1] | |
| Burn [4] | |
| Cobb & Willan [48] | |
| Debelius & Kuiter [49] | |
| Buchanan [50] | |
| Electronic sources | NudiPixel [51] |
| Atlas of Living Australia [52] | |
| Sea Slug Forum [53] | |
| OBIS [54] | |
| Cobb & Mullins [55] | |
| Köhler [56] | |
| Clarke & Buckland [57] | |
| Riek [58] | |
| GBIF [59] |
Table 2.
Distribution records of Mariaglaja tsurugensis from eastern Australia.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Lizard Island, QLD | 1979 | 14°40′24″ S | 145°27′05″ E | [61] |
| Angourie, NSW | 2017 | 29°28′53″ S | 153°21′54″ E | This paper |
| Solitary Islands, NSW | 1988 | 30°12′13″ S | 153°16′52″ E | [50] |
| Lord Howe Island, NSW | 1979 | 31°33′20″ S | 159°04′56″ E | [46] |
| Bermagui, NSW | 2017 | 36°25′22″ S | 150°05′13″ E | This paper |
Table 3.
Distribution records of Haminoea cymbalum from eastern Australia.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Great Detached Reef | 1981 | 11°44′23″ S | 144°03′46″ E | [52] |
| Lizard Island, QLD | 2004 | 14°57′23″ S | 145°44′05″ E | [52] |
| Marble Island, QLD | 2002 | 21°59′03″ S | 150°09′21″ E | [52] |
| Saumarez Reef, QLD | 1983 | 21°50′00″ S | 153°39′59″ E | [52] |
| Heron Island, QLD | 1981 | 23°26′35″ S | 151°54′35″ E | [52] |
| Gold Coast, QLD | 2007 | 28°03′12″ S | 153°26′27″ E | [47] |
| Solitary Islands, NSW | 2015 | 30°12′13″ S | 153°16′52″ E | [5] |
| South West Rocks, NSW | 2014 | 30°55′49″ S | 153°05′33″ E | [52] |
| Port Stephens, NSW | 1987 | 32°43′54″ S | 152°32′00″ E | [52] |
| Bermagui, NSW | 2017 | 36°25′22″ S | 150°05′13″ E | This paper |
Table 4.
Global distribution records of Elysia sp.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Milne Bay, Papua New Guinea | 2007 | 10°24′21″ S | 150°36′36″ E | [46,47] |
| North Solitary Island, NSW | 2017 | 29°55′30″ S | 153°23′20″ E | This paper |
Table 5.
Selected global distribution records of Thuridilla flavomaculata.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Saipan, N. Mariana Islands | 2006 | 15°09′15″ N | 145°45′13″ E | [66] |
| Guam | 2010 | 13°26′32″ N | 144°51′35″ E | [65] |
| Marshall Islands | 1995 | 07°07′53″ N | 171°11′04″ E | [65] |
| Bunaken, Indonesia | 2006 | 01°37′51″ N | 124°45′42″ E | [67] |
| Lembeh, Indonesia | 2009 | 01°26′43″ N | 125°12′56″ E | [51] |
| Tukang Besi, Indonesia | 2000 | 05°27′17″ S | 123°39′30″ E | [68] |
| Alor, Indonesia | 2010 | 08°15′01″ S | 124°29′37″ E | [51] |
| Malindi, Kenya | 2010 | 03°13′34″ S | 040°08′38″ E | [51] |
| Lord Howe Island, NSW | 2017 | 31°33′45″ S | 159°05′36″ E | This paper |
Table 6.
Selected Australian distribution records of Chromodoris magnifica.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Cartier & Ashmore Reefs, WA | 1999 | 12°15′26″ S | 123°00′01″ E | [70] |
| Fitzroy Island, QLD | 2015 | 16°55′18″ S | 145°59′00″ E | [71] |
| Heron Island, QLD | 1999 | 23°26′35″ S | 151°54′38″ E | [72] |
| Gneering Shoals, QLD | 2012 | 26°39′00″ S | 153°11′00″ E | [48] |
| Cook Island, NSW | 2013 | 28°11′46″ S | 153°34′35″ E | [73] |
| North Solitary Island, NSW | 2017 | 29°55′50″ S | 153°23′29″ E | This paper |
| Lord Howe Island, NSW | 2009 | 31°33′45″ S | 159°05′36″ E | [51] |
Table 7.
Australian distribution records of Doriprismatica dendrobranchia.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Swain Reefs, QLD | 1990 | 21°45′18″ S | 152°22′20″ E | [74] |
| Noosa, QLD | 2004, 2018 | 26°21′33″ S | 153°06′46″ E | [76] |
| Lord Howe Island, NSW | 2017 | 31°33′45″ S | 159°05′36″ E | This paper |
Table 8.
Global distribution records of Doriprismatica paladentata.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Anilao, Philippines | 2010 | 13°42′45″ N | 120°51′10″ E | [56] |
| Vietnam | 2005 | 11°12′06″ N | 109°08′41″ E | [51] |
| Banda Islands, Indonesia | 2010 | 04°43′09″ S | 129°59′35″ E | [51] |
| Milne Bay, PNG | 1982 | 10°02′45″ S | 150°05′02″ E | [79] |
| Espiritu Santo Is., Vanuatu | 1999 | 15°15′59″ S | 166°40′44″ E | [53] |
| Cook Island, NSW | 2017 | 28°11′46″ S | 153°34′35″ E | This paper |
Table 9.
Eastern Australian distribution records of Phyllodesmium magnum.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Keppel Island, QLD | 1969 | 23°10′30″ S | 150°55′38″ E | [46,47] |
| Heron Island, QLD | 1999 | 23°26′35″ S | 151°54′38″ E | [72] |
| Gneering Shoals, QLD | 2011 | 26°39′00″ S | 153°11′00″ E | [48] |
| Cook Island, NSW | 1977 | 28°11′46″ S | 153°34′35″ E | [46] |
| Julian Rocks, NSW | n.d. | 28°36′42″ S | 153°37′44″ E | [57] |
| Solitary Islands, NSW | 1988 | 30°04′03″ S | 153°20′15″ E | [50] |
| South West Rocks, NSW | 2016 | 30°55′49″ S | 153°05′33″ E | This paper, [5] |
Table 10.
Eastern Australian distribution records of Phestilla minor.
| Location | Date | Latitude | Longitude | Reference |
|---|---|---|---|---|
| Lizard Island, QLD | 1979 | 14°40′24″ S | 145°27′05″ E | [86] |
| Mooloolaba, QLD | 2004 | 26°40′32″ S | 153°08′12″ E | [55] |
| Hastings Point, NSW | 2006 | 28°21′40″ S | 153°34′50″ E | [58] |
| Port Stephens, NSW | 2017 | 32°43′54″ S | 152°32′00″ E | This paper |
Table 11.
Range extension observations by calendar year.
| Year | |||||||
| 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 |
| 1 | - | - | 3 | 5 | 13 | 7 | 8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nimbs, M.J.; Smith, S.D.A. Beyond Capricornia: Tropical Sea Slugs (Gastropoda, Heterobranchia) Extend Their Distributions into the Tasman Sea. Diversity 2018, 10, 99. https://doi.org/10.3390/d10030099
AMA Style
Nimbs MJ, Smith SDA. Beyond Capricornia: Tropical Sea Slugs (Gastropoda, Heterobranchia) Extend Their Distributions into the Tasman Sea. Diversity. 2018; 10(3):99. https://doi.org/10.3390/d10030099
Chicago/Turabian StyleNimbs, Matt J., and Stephen D. A. Smith. 2018. "Beyond Capricornia: Tropical Sea Slugs (Gastropoda, Heterobranchia) Extend Their Distributions into the Tasman Sea" Diversity 10, no. 3: 99. https://doi.org/10.3390/d10030099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.