Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests

Department of Forestry and Natural Resources, University of Kentucky, Lexington, KY 40546-0073, USA
Diversity 2018, 10(2), 29; https://doi.org/10.3390/d10020029
Submission received: 8 March 2018
/
Revised: 17 April 2018
/
Accepted: 25 April 2018
/
Published: 28 April 2018
(This article belongs to the Special Issue Diversity and Conservation of Bats)
Abstract
:Most eastern North American Myotis roost in forests during summer, with species forming maternity populations, or colonies, in cavities or crevices or beneath the bark of trees. In winter, these bats hibernate in caves and are experiencing overwinter mortalities due to infection from the fungus Pseudogymnoascus destructans, which causes white-nose syndrome (WNS). Population recovery of WNS-affected species is constrained by the ability of survivors to locate habitats suitable for rearing pups in summer. Forests in eastern North America have been severely altered by deforestation, land-use change, fragmentation and inadvertent introduction of exotic insect pests, resulting in shifts in tree distributions and loss of large-diameter canopy-dominant trees. This paper explores patterns in use of tree roosts by species of Myotis across Canada and the United States using meta-data from published sources. Myotis in western Canada, the Northwest, and Southwest selected the largest diameter roost trees and also supported the largest maximum exit counts. Myotis lucifugus, M. septentrionalis and M. sodalis, three species that inhabit eastern forests and which are currently experiencing region-wide mortalities because of WNS, selected roosts with the smallest average diameters. Recovery efforts for bark- and cavity-roosting Myotis in eastern North American forests could benefit from management that provides for large-diameter trees that offer more temporally-stable structures for roosting during the summer maternity season.
1. Introduction
Alterations to forested landscapes across eastern North America from shifting land use practices, fire suppression, and inadvertent introduction of exotic forest pests have led to reduced forest land cover, younger stand ages, and changes in the composition of canopy-dominant trees. Most forests in eastern North America now represent lands recolonized through secondary succession following cessation of agricultural activities [1]. In the Northeast, there has been a large-scale shift to early- and mid-successional tree species away from late-seral canopy dominants [2]. Eastern oak (Quercus) forests have seen a widespread increase in shade-intolerant hardwoods and a decline in shade-intolerant oaks resulting from decades of fire exclusion [3]: an ecological response termed “mesophication” [4]. These forests are further threatened by outbreaks of gypsy moths (Lymatria dispar), which cause mortality of both overstory and young trees in oak species [5]. Several other large, canopy-dominant tree species in eastern North America have experienced mortalities and declines due to introduced forest pests including Canadian hemlock (Tsuga canadensis), American beech (Fagus grandifolia), ash (Fraxinus spp.), and American chestnut (Castanea dentata) [6]. Shifts away from historically available tree species and canopy structures are likely to be accompanied by changes in behavior of bark- and cavity-roosting bats, particularly in selecting roosting and maternity habitats during summer months.
The importance of tall large-diameter dead trees to bark- and cavity-roosting bats in coniferous forests of western North America is well documented [7,8,9]. These “legacy” trees, i.e., roosting sites that remain available on the landscape for generations of bats, are believed: to confer thermal advantages from increased solar exposure [10], to be recognizable to foraging bats within habitat patches while in flight [11], and to persist for longer periods of time than smaller-diameter dead trees [12]. Selection for larger-diameter trees by bats in eastern North American forests is less evident [13], and it is likely that loss of canopy-dominant trees has influenced existing patterns in roost selection by these bat species. Use of multiple roosts with variable patterns of roost-switching, i.e., fission-fusion behavior, has been described in tree-roosting bats [14,15], including Myotis [16,17,18], with evidence for reliance on “central-node” roosts in some populations [18,19,20]. Loss of canopy-dominant tree species from eastern North American forests has likely decreased the availability of large-diameter, long-standing trees, that otherwise could serve as long-lived roosts which provide stability across time and landscape position for summer populations of bark- and cavity-roosting bats.
Most Myotis in eastern North America form maternity populations in cavities or crevices or beneath the bark of trees during the growing season, making these bats ideal species for occupying legacy trees in forests [13]. These bats make migrations of up to several hundred kilometers from maternity sites in late summer/early autumn to caves and mines, where they mate and then overwinter to wait out the period of food shortage by entering deep torpor, i.e., a state of reduced body temperature, to minimize energy loss and stretch energy reserves through the winter. Many cave-hibernating Myotis have experienced recent catastrophic losses in population numbers due to an introduced fungal pathogen, Pseudogymnoascus destructans [21,22,23,24], with range-wide extinction of some species predicted [25,26,27]. The fungus causes structural damage to bat tissues, especially wings, with hyphae that penetrate hair follicles, apocrine glands, and sebaceous glands [28]. The fungus attacks bats while torpid, increasing the frequency of arousal and leading to energy loss, dehydration, emaciation, and immune responses to fungal infection that are lethal to the bats [29,30,31]. Collectively, these effects on hibernating bats have been termed white-nose syndrome (WNS), with recovery efforts of WNS-impacted Myotis now reliant on the ability of overwinter survivors to reproduce and successfully rear young bats to volancy during summer months [32]. The extent to which increased availability of large-diameter trees in eastern North American forests could facilitate reproductive success and recovery in these bats has not been explored. This study examines patterns in use of tree roosts by species of Myotis across Canada and the United States, using meta-data from published sources, and compares use of roosts across species and geographic regions. Two hypotheses were evaluated: Myotis in western North America select larger-diameter and taller trees for roosting than do Myotis in eastern regions due to a greater abundance of older-age forests and, thus, higher availability of larger-diameter live and dead trees; and, tree roosts of Myotis in western North America support larger-numbers of bats in maternity populations than do tree roosts of Myotis in eastern forests, due to a longer temporal stability of roosts combined with a greater volume of available roosting space within roosts.
2. Materials and Methods
To assess geographic patterns in use of tree roosts by Myotis, I surveyed the published literature from 1970 to 2015 for articles referring to tree roosting in North American Myotis. I limited my search to studies occurring north of Mexico. I used Web of Science as the primary search engine to identify relevant literature. I also extracted information from articles in two published symposia, one on forest bats [33] and another specific to the Indiana bat, M. sodalis [34]. I located 63 published sources that had data useful for analysis (Appendix A). Within each article I searched for population-level data on: mean diameter of roost trees (cm), mean height of roost trees (m), and mean size of exit counts from roost trees (No. bats/count). When data were provided on random or adjacent stand conditions, I compared these with values for mean roost-tree diameter and mean roost-tree height. I also identified studies specific to a single roost tree and studies in which the maximum exit count was reported from among a set of roost trees sampled, and collected information on tree diameter (cm), tree height (m), and species of tree from which the maximum exit count was observed. I screened studies to minimize use of data for analysis from populations likely affected by WNS impacts.
Data were partitioned by geographic regions (Figure 1). Western Canada included British Columbia, Alberta, Yukon, Northwest Territories, and the U.S. state of Alaska. Central Canada included Saskatchewan, Manitoba, Nunavut, and Ontario, and eastern Canada covered the remaining provinces east of Ontario. For the U.S., the northwest region was bordered by Montana, Wyoming, and northern California. The southwest region covered southern California to Colorado, and south to Texas. The Midwest region included the states from North Dakota to Oklahoma, northeast to Ohio and Michigan. The southeast region was bordered on the west by Louisiana and Arkansas and to the north by Kentucky and Virginia. The northeast region included all states north of Virginia and east of Ohio. Data were also partitioned among 11 species of Myotis for analysis, including: M. auriculus (southwestern myotis), M. austroriparius (southeastern bat), M. californicus (California myotis), M. evotis (western long-eared bat), M. keenii (Keen’s myotis), M. lucifugus (little brown myotis), M. septentrionalis (northern long-eared bat), Indiana bat, M. thysanodes (fringed myotis), M. volans (long-legged myotis), and M. yumanensis (Yuma myotis).
I compared maximum exit counts, mean exit counts, mean roost tree diameter, and mean roost tree height by geographic region using one-way analysis of variance, with an LSD procedure to identify differences when ANOVAs were significant (PROC GLM, SAS Institute, Cary, NC, USA). Comparisons across Myotis were evaluated similarly for mean roost-tree diameter and mean roost-tree height. To assess the extent to which different Myotis selected roost trees from among those available in the habitat, I calculated the percent difference from surrounding stand-level-conditions for each species, when reported, for both mean roost-tree diameter and mean roost-tree height. Percent differences were then arcsine transformed and evaluated using one-way analysis of variance with an LSD procedure (PROC GLM, SAS Institute, Cary, NC, USA). To assess relationships of maximum exit counts with associated roost tree diameter and roost tree height, and mean exit counts with mean roost tree diameter and mean roost tree height, I performed simple linear and multiple linear regressions combining data across the continent (PROC REG, SAS Institute, Cary, NC, USA). Statistical tests were considered significant when p < 0.05.
3. Results
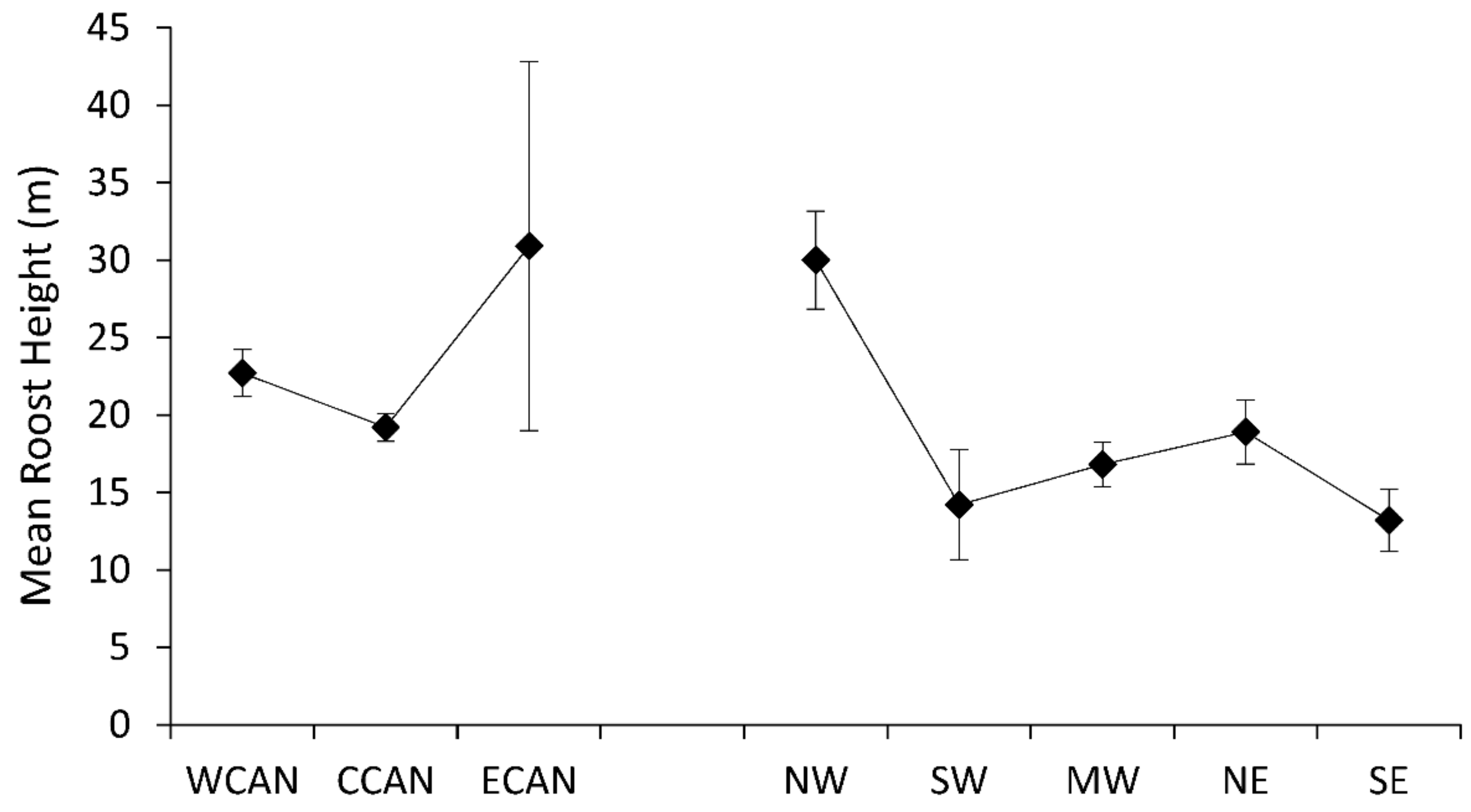
Mean diameter of tree roosts of Myotis demonstrated longitudinal trends from west to east of decreasing roost size in both Canada and the United States (Figure 2). Differences among regions were significant (F7,66 = 6.64, p < 0.0001), with largest average roost diameters for bats in western Canada, the northwest, and southwest regions. Tree roosts of Myotis also exhibited a decreasing trend for height of roost trees from west to east across the United States (F7,54 = 5.39, p < 0.0001); however, data for Canadian populations were reversed with the tallest roost trees recorded in eastern Canada (Figure 3). Variation in roost-tree height was much wider for populations of Myotis in eastern Canada than for all other regions.
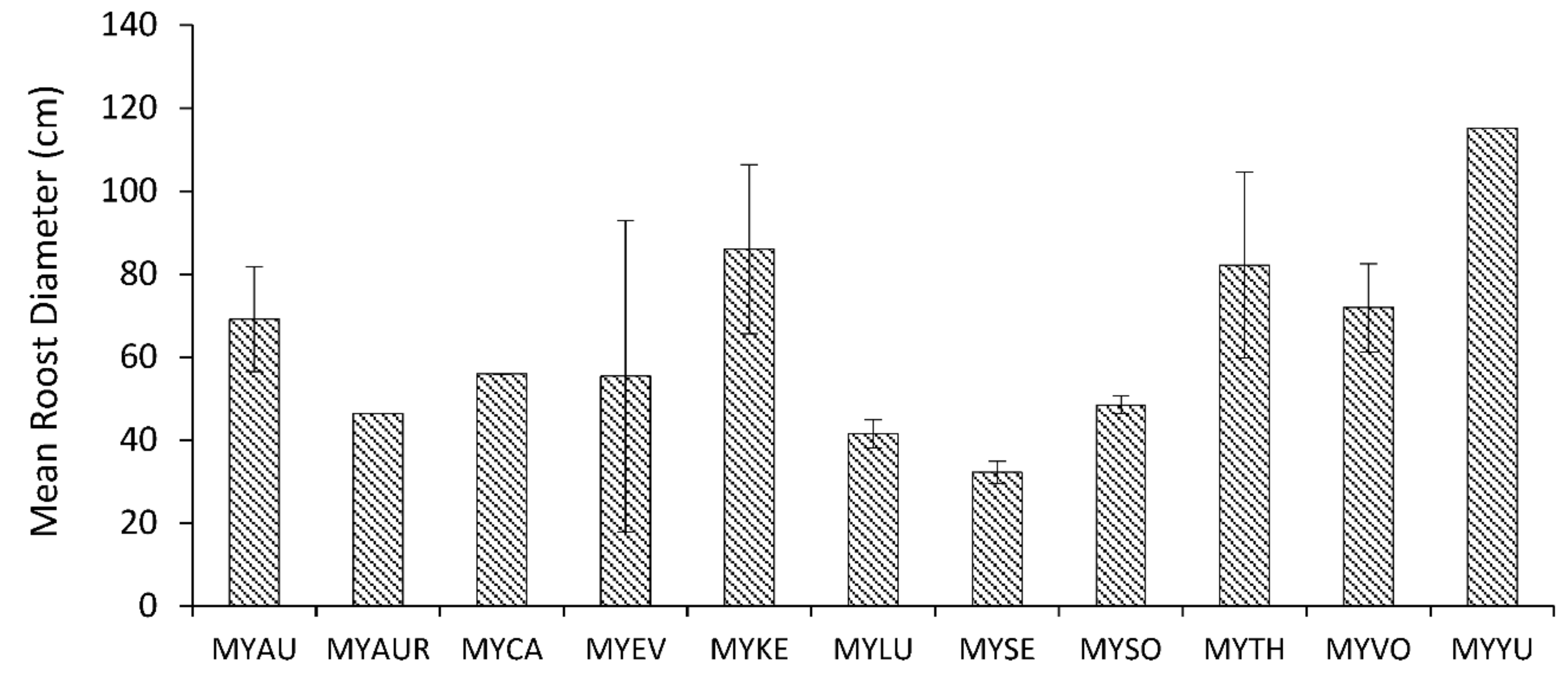
Comparisons among species of Myotis demonstrated differences in mean roost diameter (F10,61 = 7.63, p < 0.0001), but not mean height of roost trees (F9,50 = 1.5, p = 0.1755). Species with the largest roost diameters were Myotis auriculus, M. keenii, M. thysanodes, M. volans, and M. yumanensis (Figure 4); all of these species occur only in western North America. Conversely, populations of M. lucifugus, M. septentrionalis, and M. sodalis, primarily inhabiting eastern forests, used roost trees with the smallest diameters.
Evaluation of differences in roost-tree diameter from surrounding stand conditions demonstrated variation among Myotis (F10,33 = 3.87, p < 0.0015), with the greatest difference observed for M. yumanensis and the least differences observed for M. austroriparius and M. evotis (Table 1). Detectable patterns in difference from stand conditions for height of roost trees were not observed among Myotis (F9,21 = 0.14, p = 0.9979).
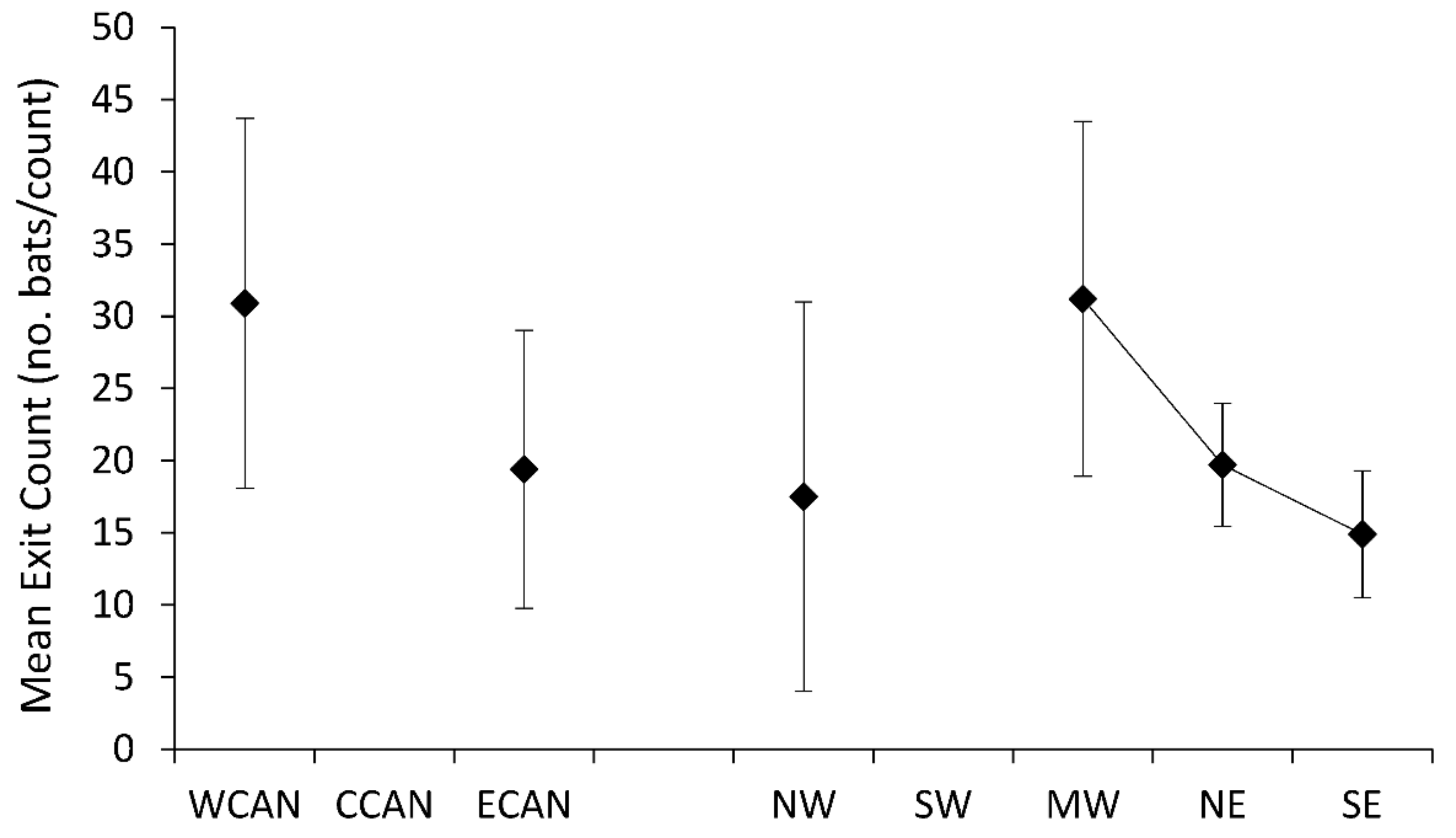
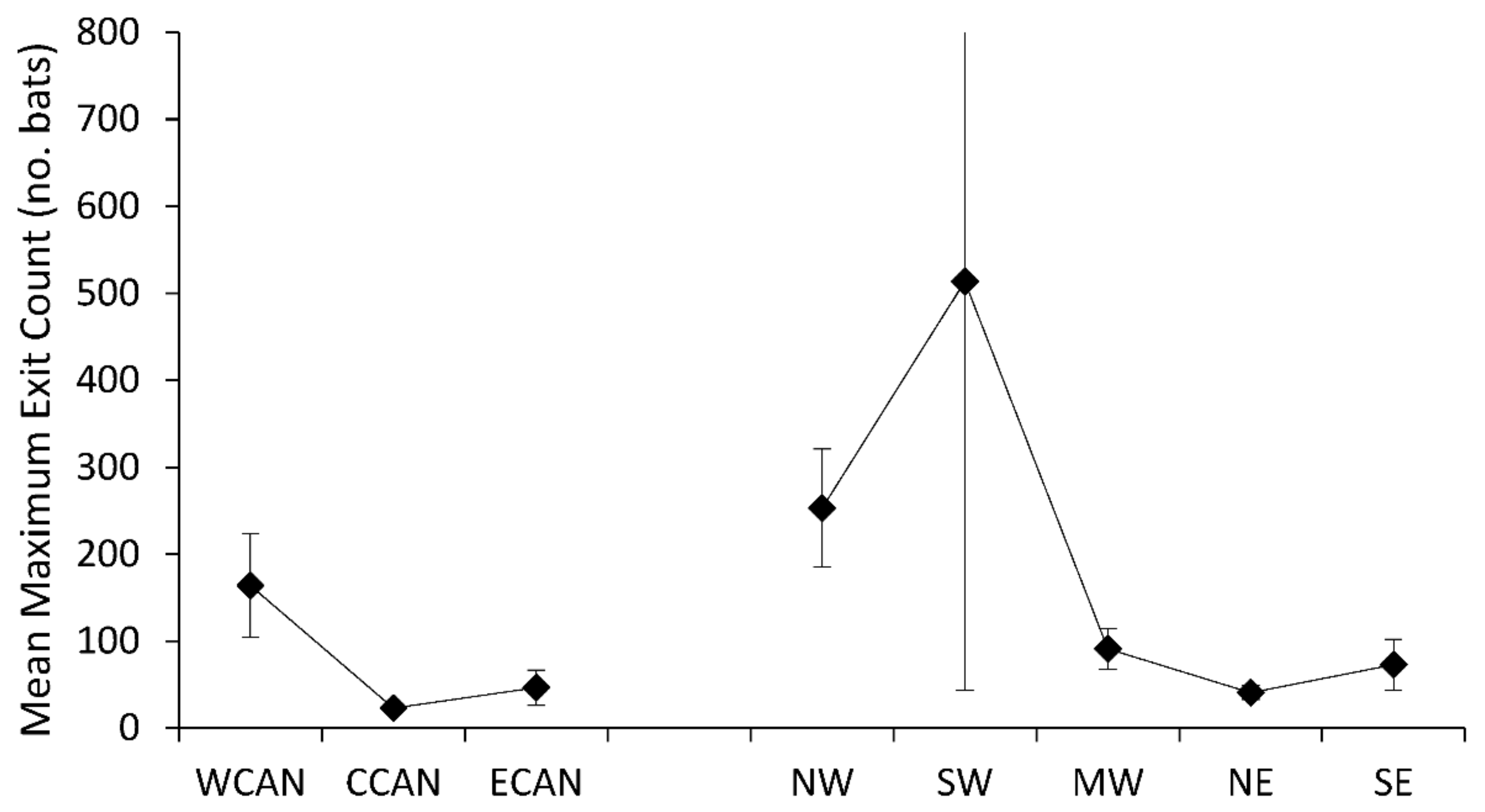
Seventeen species of trees were associated with maximum exit counts of Myotis (Supplementary Table S1), with the largest population sizes recorded for a Myotis sp. in a Pseudotsuga menziesii (Douglas fir, n = 984), for M. volans in a Pinus ponderosa (Ponderosa pine, n = 459), and for M. sodalis in a Populus deltoides (eastern cottonwood, n = 384). Mean maximum exit counts varied regionally (F7,39 = 3.23, p < 0.0084), with highest maximum counts for tree-roosting populations of Myotis in western Canada, the northwest, and southwest regions (Figure 5). Variation in maximum exit counts from tree roosts was greatest for Myotis in southwestern U.S. Mean exit counts of tree-roosting Myotis did not show discernible patterns by geographic region (F5,26 = 0.34, p = 0.8868); however, there were limited data for this metric in two regions, central Canada and southwestern U.S. (Figure 6).
Continent-wide trends in mean maximum exit counts and average exit counts of tree-roosting populations of Myotis demonstrated no clear relationships with roost tree diameter, roost tree height or for models incorporating both characteristics simultaneously (Table 2). Adjusted R2-values failed to exceed 0.13 for any model comparison.
4. Discussion
Findings corroborated the importance of large-diameter trees for summer roosts of Myotis in western North America [7,8,9], with a distinct longitudinal trend from western to eastern populations of decreasing roost-tree diameter. Patterns for height of roost trees were more equivocal, with decreasing height of roost trees from west to east in more southern U.S. populations of Myotis contrasted by increasing height of roost trees from west to east in Canadian populations. Average roost-tree diameter differed among Myotis with western species occupying the largest roosts on average, and eastern species selecting the smallest diameter roosts, especially M. sodalis, eastern and Midwestern populations of M. septentrionalis, and populations of the eastern subspecies of M. lucifugus lucifugus. These three species are all cave-hibernators that are vulnerable to impacts from WNS [25,27,35]. Differences in roost-tree diameter from surrounding stand conditions varied among Myotis, but showed no consistent clinal variation across geographic regions. The geographic difference observed for roost-tree diameter in this study suggests two possibilities. Use of smaller-diameter roosts by eastern North American Myotis represents a difference in preferred roost condition from that of western species, or, is an indication that roost choice in eastern Myotis is constrained by a shortage of available large-diameter trees for roosting in eastern forests.
Average size of bat populations exiting tree roosts did not vary regionally, however, maximum exit counts observed were largest for populations of Myotis in the southwest region, the northwest, and western Canada. Counts of bats exiting tree roosts that exceeded 300 individuals were rare in studies of eastern and midwestern populations [36,37], but were frequently reported for western populations of Myotis [9,38,39,40,41,42]. Interestingly, all maximum counts exceeding 300 bats at roosts of M. lucifugus, a species whose range encompasses both eastern and western regions, occurred in populations from western Canada [41,42]. Thus, large maternity populations are rare in eastern Myotis, but potential exits for at least some species to form large maternity populations in eastern forests with sufficient availability of suitable roosting structures in the form of large-diameter live and dead trees.
Bark- and cavity-roosting Myotis reuse roost trees over multiple seasons [13,43,44], with populations of Myotis in Europe known to inhabit the same roost tree in summer for up to 16 years [45]. This longevity is constrained, however, by how long it takes for standing dead trees to fall [46]. Survivorship of standing dead trees varies by species and size [47,48], with trees larger in diameter and shorter in height persisting longer [47,49]. Half-lives of tree roosts of M. volans in coniferous forests in the northwest U.S. were <3 years, with only 4.3% of roosts remaining upright after 10-years post-discovery [12]. Suitability of tree roosts for reuse by M. californicus in western Canada also declined rapidly post-discovery [44]. These patterns would indicate that an increase in the availability of large-diameter trees could help in maintaining connectivity and persistence of maternity populations of Myotis at local scales, as these trees are more likely to persist and remain standing for longer periods [12,47,49]. Loss of roosts results in changes in behavior of some species of bats [50,51], with M. lucifugus know to abandon local roosting areas with loss of access to a significant building roost [52].
To an extent, bark- and cavity-roosting bats are preadapted to seasonal and annual shifts in availability of tree roosts. These species form social units of varying relatedness in which individual members move among roost trees and associated population sub-groups, or fission-fusion behavior [14,53]. Fission-fusion behavior is well documented in North American bats, including Myotis [16,17,18], with highly-centralized, scale-free roost networks centered around a central-node roost tree typical of some maternity populations [18,19]. A central-node roost serves as the focal site of a maternity population, within which all members of the colony roost at some point in time [54]. Central-node roosts are commonly enclosed on the landscape by a number of peripheral or satellite roosts, each used by only a subset of bats in the colony. In Myotis, loss of central-node or peripheral roosts has resulted in minimal impact to roosting patterns as bats remained in the same roosting areas [18,55]; however, increases in distances moved and changes in social behavior were observed in M. septentrionalis following removal of roosts [55].
Declines in the abundance and distribution of American chestnut, due to the introduced blight fungus (Cryphonectria parasitica), certainly represented a significant change to canopy structures and deadwood biomass in eastern North American forests where this species once thrived. American chestnut had a distribution from New England to the southern Appalachian Mountains where it represented >50% of the timber volume in stands on well-drained slopes [56,57]. The largest tree in eastern North American forests, American chestnut achieved heights of 40 m and diameters of 3 m, and lived as long as 600 years [58]. American chestnut has thick bark in older trees and decay-resistant wood [59,60,61], contributing to the potential of this tree as a long-term habitat structure for use by wildlife as shelter [62,63]. The extent to which bark- and cavity-roosting bats used American chestnut for roosting is unknown, however, the size and longevity of these trees likely made them a desirable choice of bats as roosting habitat. American chestnut trees occurred at an estimated historic population size of 4.2 billion trees [64]. Loss of American chestnut likely resulted in a pulse of dead wood legacy structures that have now all but disappeared from forested landscapes in eastern North America where the species occurred as a canopy dominant. Impacts to roosting bats were likely, as these trees may have served as temporally stable, central-node roost trees for populations of bark- and cavity-roosting Myotis bats. Chestnut trees were once distributed across >800,000 km2 of eastern North American forests [60], and the disappearance of American chestnuts from the landscape may have required populations of bats to roost more frequently in alternate trees, most of which do not achieve the same size or persist for as long as American chestnut did before population collapse.
Limited data exist for survivorship of bark- and cavity-roosting bats [65,66], including North American Myotis, with existing studies of survivorship in these bats having focused primarily on populations inhabiting caves [67,68]. Implicit to benefits of large-diameter trees for roosting in bats is the assumption that use of these structures, especially by larger colonies, will lead to enhanced survival and fitness of individuals within the colony. Studies in New Zealand have demonstrated higher survival rates for populations of Chalinolobus tuberculatus roosting in large-diameter, well insulated, tree roosts, compared with bats occupying poorly insulated, roosts in smaller-diameter stems [69,70]. Loss of 97% of the original forest in some areas was responsible for a decline in large-diameter trees, leading to use of poor quality roosting habitat by this bat following disturbance [71]. Although limited in scope, these data would indicate that disease, fragmentation, and loss of forests in eastern North America, and the concomitant decline in availability of large-diameter trees, may have produced similar responses in summer populations of bark- and cavity-roosting Myotis bats. Regardless, a full understanding of the effects of roost trees on bark- and cavity-roosting bats also necessitates continued research into the importance or limitations of smaller-diameter trees as roosts for bats, focusing on reproductive and survivorship rates of both solitary-roosting individuals and small colonies, and the role of these smaller-diameter trees in maintaining connectivity of roost networks of bat colonies.
Maternity habitat that allows for gestation, parturition, and the successful rearing of young to sexual maturity is fundamental to the long-term conservation of tree-roosting Myotis affected by WNS [72,73]. Results from this study show that eastern Myotis currently rely on smaller-diameter trees for use as maternity roosts compared to western North American species and live in colonies that achieve smaller maximum population sizes. Through use of retention forestry methods, that provide for uncut forested habitats in varying patch sizes and spatial arrangements [74], development of landscapes supporting higher densities of large-diameter trees could lead to increased stability and long-term enhancements in available roosting habitat of bark- and cavity-roosting bats in forests of eastern North America. Natural disturbance-based silviculture is increasingly being applied with the intent of creating habitat conditions comparable to those resulting from natural disturbance events [75], while enhancing forest resilience and ability of forests to respond to environmental shifts [76,77]. Nevertheless, forest management which promotes large-diameter trees, with potential to remain standing for longer intervals, should create opportunities for formation of larger maternity populations of bats, potentially leading to improved survivorship, more favorable internal micro-climates, and increased benefits from social thermoregulation [70,78,79].
5. Conclusions
Concerted efforts are underway to develop blight resistant chestnut trees and restore this tree as a canopy dominant in eastern North American forests through genetic engineering, inter- and intra-specific breeding programs, and use of biocontrol with hypovirulence [80]. In turn, recent advances in the study of the Pseudogymnoascus fungus responsible for WNS have identified potential control strategies to reduce impacts to hibernating bats including use of ultraviolet light and microbial inhibitors [31,81]. Perhaps one day bark- and cavity-roosting bats in eastern North America will occupy American chestnut trees as roosting sites, but until then management efforts to increase availability of large-diameter trees via other canopy-dominant tree species could help facilitate reproduction and population recovery of bark-and cavity-roosting Myotis impacted by WNS. As a starting point, the U.S. Forest Service manages 6 million hectares of forests within the former range of the American chestnut [82], and silvicultural efforts to promote green-tree retention strategies on these public lands could begin the process of returning large-diameter trees to eastern forests. The positive impacts of legacy trees can be even more pronounced in plantation forests, where biological reserves occur within an expanse of exotic tree plantations [83], and in disturbed and fragmented forests [84,85]. Regardless, management that fosters development of large-diameter trees will not be implemented without problems associated with balancing tree recruitment against mortality [86], and effects of climate change that could lead to geographic shifts in the distribution of canopy-dominant tree species over time [87].
Supplementary Materials
The following are available online at https://www.mdpi.com/1424-2818/10/2/29/s1, Table S1: Supplemental file of legacy structure data base.
Funding
This research received no external funding.
Acknowledgments
This manuscript (KAES No.: 18-09-031) is connected with a project of the Kentucky Agricultural Experiment Station and is published with support and approval of the director.
Conflicts of Interest
The author declares no conflict of interest.
Appendix A. Literature Sources Used in Analyses of Roosting Preferences of Myotis in North America
| Baker, M.D.; Lacki, M.J. Day-roosting habitat of female long-legged Myotis in ponderosa pine forests. J. Wildl. Manag. 2006, 70, 207–215. |
| Bergeson, S.M.; Carter, T.C.; Whitby, M.D. Adaptive roosting gives little brown bats an advantage over endangered Indiana bats. Am. Midl. Nat. 2015, 174, 321–330. |
| Bernardos, D.A.; Chambers, C.L.; Rabe, M.J. Selection of gambel oak roosts by southwestern Myotis in ponderosa pine-dominated forests, northern Arizona. J. Wildl. Manag. 2004, 68, 595–601. |
| Boland, J.L.; Hayes, J.P.; Smith, W.P.; Huso, M.H. Selection of day-roosts by Keen’s Myotis (Myotis keenii) at multiple spatial scales. J. Mammal. 2009, 90, 222–234. |
| Brandebura, S.C.; Pannkuk, E.L.; Risch, T.S. Indiana bat (Myotis sodalis) maternity colonies in Arkansas. Southeast. Nat. 2011, 10, 529–532. |
| Brigham, R.M.; Vonhof, M.J.; Barclay, R.M.R.; Gwilliam, J.C. Roosting behavior and roost-site preferences of forest-dwelling California bats (Myotis californicus). J. Mammal. 1997, 78, 1231–1239. |
| Britzke, E.R.; Harvey, M.J.; Loeb, S.C. Indiana bat, Myotis sodalis, maternity roosts in the southern United States. Southeast. Nat. 2003, 2, 235–242. |
| Britzke, E.R.; Hicks, A.C.; von Oettingen, S.L.; Darling, S.R. Description of spring roost trees used by female Indiana bats (Myotis sodalis) in the Lake Champlain Valley of Vermont and New York. Am. Midl. Nat. 2006, 155, 181–187. |
| Broders, H.G.; Forbes, G.J. Interspecific and intersexual variation in roost-site selection of northern long-eared and little brown bats in the Greater Fundy National Park ecosystem. J. Wildl. Manag. 2004, 68, 602–610. |
| Callahan, E.V.; Drobney, R.D.; Clawson, R.L. Selection of summer roosting sites by Indiana bats (Myotis sodalis) in Missouri. J. Mammal. 1997, 78, 818–825. |
| Carter, T.C.; Feldhamer, G.A. Roost tree use by maternity colonies of Indiana bats and northern long-eared bats in southern Illinois. For. Ecol. Manag. 2005, 219, 259–268. |
| Carver, B.D.; Ashley, N. Roost tree use by sympatric Rafinesque’s big-eared bats (Corynorhinus rafinesquii) and southeastern Myotis (Myotis austroriparius). Am. Midl. Nat. 2008, 160, 364–373. |
| Clement, M.J.; Castleberry, S.B. Divergent roosting habits of Rafinesque’s big-eared bat and southeastern Myotis during winter floods. Am. Midl. Nat. 2013, 170, 158–170. |
| Cope, J.B.; Richter, A.R.; Mills, R.S. A summer concentration of the Indiana bat, Myotis sodalis, in Wayne County, Indiana. Proc. Ind. Acad. Sci. 1974, 83, 482–484. |
| Crampton, L.H.; Barclay, R.M.R. 1996. Habitat selection by bats in fragmented and unfragmented aspen mixedwood stands of different ages. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 238−259. |
| Crampton, L.H.; Barclay, R.M.R. Selection of roosting and foraging habitat by bats in different-aged aspen mixedwood stands. Conserv. Biol. 1998, 12, 1347–1358. |
| Cryan, P.M.; Bogan, M.A.; Yanega, G.M. Roosting habits of four bat species in the Black Hills of South Dakota. Acta Chiropterol. 2001, 3, 43–52. |
| Evelyn, M.J.; Stiles, D.A.; Young, R.A. Conservation of bats in suburban landscapes: Roost selection by Myotis yumanensis in a residential area in California. Biol. Conserv. 2004, 115, 463–473. |
| Fleming, H.L.; Jones, J.C.; Belant, J.L.; Richardson, D.M. Multi-scale roost site selection by Rafinesque’s big-eared bat (Corynorhinus rafinesquii) and southeastern Myotis (Myotis austroriparius) in Mississippi. Am. Midl. Nat. 2013, 169, 43–55. |
| Ford, W.M.; Owen, S.F.; Edwards, J.W.; Rodrigue, J.L. Robinia pseudoacacia (black locust) as day-roosts of male Myotis septentrionalis (northern bats) on the Fernow Experimental Forest, West Virginia. Northeast. Nat. 2006, 13, 15–24. |
| Foster, R.W.; Kurta, A. Roosting ecology of the northern bat (Myotis septentrionalis) and comparisons with the endangered Indiana bat (Myotis sodalis). J. Mammal. 1999, 80, 659–672. |
| Garroway, C.J.; Broders, H.G. Nonrandom association patterns at northern long-eared bat maternity roosts. Can. J. Zool. 2007, 85, 956–964. |
| Garroway, C.J.; Broders, H.G. Day roost characteristics of northern long-eared bats (Myotis septentrionalis) in relation to female reproductive status. Ecoscience 2008, 15, 89–93. |
| Gooding, G.; Langford, J.R. Characteristics of tree roosts of Rafinesque’s big-eared bat and southeastern bat in northeastern Louisiana. Southwest. Nat. 2004, 49, 61–67. |
| Grindal, S.D. Habitat use by bats, Myotis spp., in western Newfoundland. Can. Field-Nat. 1998, 113, 258–263. |
| Hofmann, J.E.; Gardner, J.E.; Krejca, J.K.; Garner, J.D. Summer records and a maternity roost of the southeastern Myotis (Myotis austroriparius) in Illinois. Trans. Il. State Acad. Sci. 1999, 92, 95–107. |
| Humphrey, S.R.; Richter, A.R.; Cope, J.B. Summer habitat and ecology of the endangered Indiana bat, Myotis sodalis. J. Mammal. 1977, 58, 334–346. |
| Johnson, J.B.; Gates, J.E. Observations of Indiana Myotis roosting and foraging behavior in Carroll County, Maryland. Md. Nat. 2009, 50, 11–30. |
| Johnson, J.B.; Ford, W.M.; Rodrigue, J.L.; Edwards, J.W.; Johnson, C.M. Roost selection by male Indiana Myotis following forest fires in central Appalachian hardwood forests. J. Fish Wildl. Manag. 2010, 1, 111–121. |
| Johnson, J.B.; Ford, W.M.; Edwards, J.W. Roost networks of northern Myotis (Myotis septentrionalis) in a managed landscape. For. Ecol. Manag. 2012, 266, 223–231. |
| Jung, T.S., Thompson, I.D.; Titman, R.D. Roost site selection by forest-dwelling male Myotis in central Ontario, Canada. For. Ecol. Manag. 2004, 202, 325–335. |
| Kalcounis, M.C.; Hecker, K.R. Intraspecific variation in roost-site selection by little brown bats (Myotis lucifugus). In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 81–90. |
| Kiser, J.D.; MacGregor, J.R.; Bryan, H.D.; Howard, A. Use of concrete bridges as nightroosts. In The Indiana Bat: Biology and Management of An Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 208–215. |
| Kurta, A.; King, D.; Teramino, J.A.; Stribley, J.M.; Williams, K.J. Summer roosts of the endangered Indiana bat (Myotis sodalis) on the northern edge of its range. Am. Midl. Nat. 1993, 129, 132–138. |
| Kurta, A.; Kath, J.; Smith, E.L.; Foster, R.; Orick, M.W.; Ross, R. A maternity roost of the endangered Indiana bat (Myotis sodalis) in an unshaded, hollow, sycamore tree (Platanus occidentalis). Am. Midl. Nat. 1993b, 130, 405–407. |
| Kurta, A.; Williams, K.J.; Mies, R. Ecological, behavioural, and thermal observations of a peripheral population of Indiana bats (Myotis sodalis). In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 102–117. |
| Kurta, A.; Murray, S.W.; Miller, D.H. Roost selection and movements across the summer landscape. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 118–129. |
| Lacki, M.J.; Baker, M.D. Day roosts of female fringed Myotis (Myotis thysanodes) in xeric forests of the Pacific Northwest. J. Mammal. 2007, 88, 967–973. |
| Lacki, M.J.; Schwierjohann, J.H. Day-roost characteristics of northern bats in mixed mesophytic forest. J. Wildl. Manag. 2001, 65, 482–488. |
| Lacki, M.J.; Cox, D.R.; Dodd, L.E.; Dickinson, M.B. Response of northern bats (Myotis septentrionalis) to prescribed fires in eastern Kentucky forests. J. Mammal. 2009, 90, 1165–1175. |
| Lacki, M.J.; Baker, M.D.; Johnson, J.S. Geographic variation in roost-site selection of long-legged Myotis in the Pacific Northwest. J. Wildl. Manag. 2010, 74, 1218–1228. |
| Menzel, M.A.; Owen, S.F.; Ford, W.M.; Edwards, J.W.; Wood, P.B.; Chapman, B.R.; Miller, K.V. Roost tree selection by northern long-eared bat (Myotis septentrionalis) maternity colonies in an industrial forest of the central Appalachian mountains. For. Ecol. Manag. 2002, 155, 107–114. |
| Olson, C.R.; Barclay, R.M.R. Concurrent changes in group size and roost use by reproductive female little brown bats (Myotis lucifugus). Can. J. Zool. 2013, 91, 149–155. |
| Ormsbee, P.C. Characteristics, use, and distribution of day roosts selected by female Myotis volans (long-legged Myotis) in forested habitat of the Central Oregon Cascades. In Bats and Forests Symposium, Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 124–131. |
| Ormsbee, P.C.; McComb, W.C. Selection of day roosts by female long-legged Myotis in the central Oregon Cascade Range. J. Wildl. Manag. 1998, 62, 596–603. |
| Parsons, S.; Lewis, K.J.; Psyllakis, J.M. Relationships between roosting habitat of bats and decay of aspen in the sub-boreal forests of British Columbia. For. Ecol. Manag. 2003, 177, 559–570. |
| Perry, R.W.; Thill, R.E. Roost selection by male and female northern long-eared bats in a pine-dominated landscape. For. Ecol. Manag. 2007, 247, 220–226. |
| Psyllakis, J.M.; Brigham, R.M. Characteristics of diurnal roosts used by female Myotis bats in sub-boreal forests. For. Ecol. Manag. 2006, 223, 93–102. |
| Rabe, M.J.; Morrell, T.E.; Green, H.; deVos, J.C., Jr.; Miller, C.R. Characteristics of ponderosa pine snag roosts used by reproductive bats in northern Arizona. J. Wildl. Manag. 1998, 62, 612–621. |
| Rancourt, S.J.; Rule, M.I.; O’Connell, M.A. Maternity roost site selection of long-eared Myotis, Myotis evotis. J. Mammal. 2005, 86, 77–84. |
| Sasse, D.B.; Pekins, P.J. Summer roosting ecology of northern long-eared bats (Myotis septentrionalis) in the White Mountain National Forest. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests, Victoria, BC, Canada, 1996; pp. 91–101. |
| Silvis, A.; Ford, W.M.; Britzke, E.R.; Beane, N.R.; Johnson, J.B. Forest succession and maternity day roost selection by Myotis septentrionalis in a mesophytic hardwood forest. Inter. J. For. Res. 2012, doi:10.1155/2012/148106. |
| Silvis, A.; Kniowski, A.B.; Gehrt, S.D.; Ford, W.M. Roosting and foraging social structure of the endangered Indiana bat (Myotis sodalis). PLoS ONE 2014, 9, e96937, doi:10.1371/journal.pone.0096937. |
| Silvis, A.; Ford, W.M.; Britzke, E.R.; Johnson, J.B. Association, roost use and simulated disruption of Myotis septentrionalis maternity colonies. Behav. Processes 2014, 103, 283–290. |
| Silvis, A.; Ford, W.M.; Britzke, E.R. Effects of hierarchical roost removal on northern long-eared bat (Myotis septentrionalis) maternity colonies. PLoS ONE 2015, 10, e0116356, doi:10.1371/journal.pone.0116356. |
| Timpone, J.C.; Boyles, J.G.; Murray, K.L.; Aubrey, D.P.; Robbins, L.W. Overlap in roosting habits of Indiana bats (Myotis sodalis) and northern bats (Myotis septentrionalis). Am. Midl. Nat. 2010, 163, 115–123. |
| Veilleux, J.P.; Whitaker, J.O., Jr.; Vincent, E.A. Mammals of the Newport Chemical Depot, Vermillion County, Indiana. Proc. Ind. Acad. Sci. 1998, 107, 91–104. |
| Vonhof, M.J.; Gwilliam, J.C. Intra- and interspecific patterns of day roost selection by three species of forest-dwelling bats in southern British Columbia. For. Ecol. Manag. 2007, 252, 165–175. |
| Waldien, D.L.; Hayes, J.P.; Arnett, E.B. Day-roosts of female long-eared Myotis in western Oregon. J. Wildl. Manag. 2000, 64, 785–796. |
| Watrous, K.S.; Donovan, T.M.; Mickey, R.M.; Darling, S.R.; Hicks, A.C.; von Oettingen, S.L. Predicting minimum habitat characteristics for the Indiana bat in the Champlain Valley. J. Wildl. Manag. 2006, 70, 1228–1237. |
| Weller, T.J.; Zabel, C.J. Characteristics of fringed Myotis day roosts in northern California. J. Wildl. Manag. 2001, 65, 489–497. |
| Whitaker, J.O., Jr.; Brack, V., Jr. Distribution and summer ecology in Indiana. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 48–54. |
| Whitaker, J.O., Jr.; Sparks, D.W. Roosts of Indiana bats (Myotis sodalis) near the Indianapolis International Airport (1997–2001). Proc. Ind. Acad. Sci. 2008, 117, 193–202. |
References
- Holmes, M.A.; Matlack, G.R. Agricultural history drives structure and tree species composition of second growth forest over 100 years in southeastern Ohio, USA. J. Veg. Sci. 2017, 28, 736–746. [Google Scholar] [CrossRef]
- Thompson, J.R.; Carpenter, D.N.; Cogbill, C.V.; Foster, D.R. Four centuries of change in northeastern United States forests. PLoS ONE 2013, 8, e72540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varner, J.M.; Arthur, M.A.; Clark, S.L.; Dey, D.C.; Hart, J.L.; Schweitzer, C.J. Fire in eastern North American oak ecosystems: Filling the gaps. Fire Ecol. 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M. Invasive forest defoliator contributed to the impending downward trend of oak dominance in eastern North America. Forestry 2016, 89, 284–289. [Google Scholar] [CrossRef]
- Pautasso, M.; Schlegel, M.; Holdenrieder, O. Forest health in a changing world. Microb. Ecol. 2015, 69, 826–842. [Google Scholar] [CrossRef] [PubMed]
- Betts, B. Roosts used by maternity colonies of silver-haired bats in northeastern Oregon. J. Mammal. 1998, 79, 643–650. [Google Scholar] [CrossRef]
- Mazurek, M.J.; Zielinski, W.J. Individual legacy trees influence vertebrate wildlife diversity in commercial forests. For. Ecol. Manag. 2004, 193, 321–334. [Google Scholar] [CrossRef]
- Baker, M.D.; Lacki, M.J. Day-roosting habitat of female long-legged myotis in ponderosa pine forests. J. Wildl. Manag. 2006, 70, 207–215. [Google Scholar] [CrossRef]
- Jung, T.S.; Thompson, I.D.; Titman, R.D.; Applejohn, A.P. Habitat selection by forest bats in relation to mixed-wood stand types and structure in central Ontario. J. Wildl. Manag. 1999, 63, 1306–1319. [Google Scholar] [CrossRef]
- Campbell, L.A.; Hallett, J.G.; O’Connell, M.A. Conservation of bats in managed forests: Use of roosts by Lasionycteris noctivagans. J. Mammal. 1996, 77, 976–984. [Google Scholar] [CrossRef]
- Lacki, M.J.; Baker, M.D.; Johnson, J.S. Temporal dynamics of roost snags of long-legged myotis in the Pacific Northwest, USA. J. Wildl. Manag. 2012, 76, 1310–1316. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Kurta, A. Ecology and behavior of bats roosting in tree cavities and under bark. In Bats in Forests: Conservation and Management; Lacki, M.J., Hayes, J.P., Kurta, A., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 17–59. [Google Scholar]
- Kerth, G.; König, B. Fission, fusion and nonrandom associations in female Bechstein’s bats (Myotis bechsteinii). Behaviour 1999, 136, 1187–1202. [Google Scholar] [CrossRef]
- Willis, C.K.R.; Brigham, R.M. Roost switching, roost sharing and social cohesion: Forest-dwelling big brown bats, Eptesicus fuscus, conform to the fission-fusion model. Anim. Behav. 2004, 68, 495–505. [Google Scholar] [CrossRef]
- Garroway, C.J.; Broders, H.G. Nonrandom association patterns at northern long-eared bat maternity roosts. Can. J. Zool. 2007, 85, 956–964. [Google Scholar] [CrossRef]
- Patriquin, K.J.; Leonard, M.L.; Broders, H.G.; Garroway, C.J. Do social networks of female northern long-eared bats vary with reproductive period and age? Behav. Ecol. Sociobiol. 2010, 64, 899–913. [Google Scholar] [CrossRef]
- Silvis, A.; Kniowski, A.B.; Gehrt, S.D.; Ford, W.M. Roosting and foraging social structure of the endangered Indiana bat (Myotis sodalis). PLoS ONE 2014, 9, e96937. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.B.; Ford, W.M.; Edwards, J.W. Roost networks of northern myotis (Myotis septentrionalis) in a managed landscape. For. Ecol. Manag. 2012, 266, 223–231. [Google Scholar] [CrossRef]
- Johnson, J.S.; Kropczynski, J.N.; Lacki, M.J.; Langlois, G.D. Social networks of Rafinesque’s big-eared bats (Corynorhinus rafinesquii) in bottomland hardwood forests. J. Mammal. 2012, 93, 1545–1558. [Google Scholar] [CrossRef]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.; Darling, S.R.; Gargas, A.; Niver, R.; et al. Bat white-nose syndrome: An emerging fungal pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef] [PubMed]
- Gargas, A.; Trest, M.T.; Christensen, M.; Volk, T.J.; Blehert, D.S. Geomyces destructans sp. nov associated with bat white-nose syndrome. Mycotaxon 2009, 108, 147–154. [Google Scholar] [CrossRef]
- Frick, W.F.; Puechmaille, S.J.; Hoyt, J.R.; Nickel, B.A.; Langwig, K.E.; Foster, J.T.; Barlow, K.E.; Bartonička, T.; Feller, D.; Haarsma, A.J. Disease alters macroecological patterns of North American bats. Glob. Ecol. Biogeogr. 2015, 24, 741–749. [Google Scholar] [CrossRef]
- Turner, G.G.; Reeder, D.M.; Coleman, J.T.H. A five-year assessment of mortality and geographic spread of white-nose syndrome in North American bats and a look to the future. Bat Res. News 2011, 52, 13–27. [Google Scholar]
- Frick, W.F.; Pollock, J.F.; Hicks, A.C.; Langwig, K.E.; Reynolds, D.S.; Turner, G.G.; Butchkoski, C.M.; Kunz, T.H. An emerging disease causes regional population collapse of a common North American bat species. Science 2010, 329, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Thogmartin, W.E.; King, R.A.; McKann, P.C.; Szymanski, J.A.; Pruitt, L. Population-level impact of white-nose syndrome on the endangered Indiana bat. J. Mammal. 2012, 93, 1086–1098. [Google Scholar] [CrossRef]
- Thogmartin, W.E.; Sanders-Reed, C.A.; Szymanski, J.A.; McKann, P.C.; Pruitt, L.; King, R.A.; Runge, M.C.; Russell, R.E. White-nose syndrome is likely to extirpate the endangered Indiana bat over large parts of its range. Biol. Conserv. 2013, 160, 162–172. [Google Scholar] [CrossRef]
- Moore, M.S.; Field, K.A.; Behr, M.J.; Turner, G.G.; Furze, M.E.; Stern, D.W.F.; Allegra, P.R.; Bouboulis, S.A.; Musante, C.D.; Vodzak, M.E.; et al. Energy conserving thermoregulatory patterns and lower disease severity in a bat resistant to the impacts of white-nose syndrome. J. Comp. Physiol. B 2017. [Google Scholar] [CrossRef] [PubMed]
- Verant, M.L.; Meteyer, C.U.; Speakman, J.R.; Cryan, P.M.; Lorch, J.M.; Blehert, D.S. White-nose syndrome initiates a cascade of physiologic disturbances in the hibernating bat host. BMC Physiol. 2014, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meteyer, C.U.; Barber, D.; Mandl, J.N. Pathology in euthermic bats with white nose syndrome suggests a natural manifestation of immune reconstitution inflammatory syndrome. Virulence 2012, 3, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, E.W.; Mack, J.N.; White, G.P.; Avis, T.J.; Smith, M.L. Microbial inhibitors of the fungus Pseudogymnoascus destructans, the causal agent of white-nose syndrome in bats. PLoS ONE 2017, 12, e0179770. [Google Scholar] [CrossRef] [PubMed]
- Lacki, M.J.; Dodd, L.E.; Toomey, R.S.; Thomas, S.C.; Couch, Z.L.; Nichols, B.S. Temporal changes in body mass and body condition of cave-hibernating bats during staging and swarming. J. Fish Wildl. Manag. 2015, 6, 360–370. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Brigham, R.M. (Eds.) Bats and Forests Symposium; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; p. 292.
- Kurta, A.; Kennedy, J. The Indiana Bat: Biology and Management of an Endangered Species; Bat Conservation International: Austin, TX, USA, 2002; p. 253. [Google Scholar]
- U.S. Department of the Interior. Final register for endangered and threatened wildlife and plants: Threatened species status for the northern long-eared bat with 4(d) rule. Fed. Regist. 2015, 80, 17973–18033. [Google Scholar]
- Kiser, J.D.; MacGregor, J.R.; Bryan, H.D.; Howard, A. Use of concrete bridges as night roosts. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 208–215. [Google Scholar]
- Fleming, H.L.; Jones, J.C.; Belant, J.L.; Richardson, D.M. Multi-scale roost site selection by Rafinesque’s big-eared bat (Corynorhinus rafinesquii) and southeastern myotis (Myotis austroriparius) in Mississippi. Am. Midl. Nat. 2013, 169, 43–55. [Google Scholar] [CrossRef]
- Ormsbee, P.C. Characteristics, use, and distribution of day roosts selected by female Myotis volans (long-legged myotis) in forested habitat of the Central Oregon Cascades. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 124–131. [Google Scholar]
- Ormsbee, P.C.; McComb, W.C. Selection of day roosts by female long-legged Myotis in the central Oregon Cascade Range. J. Wildl. Manag. 1998, 62, 596–603. [Google Scholar] [CrossRef]
- Rabe, M.J.; Morrell, T.E.; Green, H.; deVos, J.C., Jr.; Miller, C.R. Characteristics of ponderosa pine snag roosts used by reproductive bats in northern Arizona. J. Wildl. Manag. 1998, 62, 612–621. [Google Scholar] [CrossRef]
- Psyllakis, J.M.; Brigham, R.M. Characteristics of diurnal roosts used by female Myotis bats in sub-boreal forests. For. Ecol. Manag. 2006, 223, 93–102. [Google Scholar] [CrossRef]
- Olson, C.R.; Barclay, R.M.R. Concurrent changes in group size and roost use by reproductive female little brown bats (Myotis lucifugus). Can. J. Zool. 2013, 91, 149–155. [Google Scholar] [CrossRef]
- Foster, R.W.; Kurta, A. Roosting ecology of the northern bat (Myotis septentrionalis) and comparisons with the endangered Indiana bat (Myotis sodalis). J. Mammal. 1999, 80, 659–672. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Brigham, R.M. Year-to-year reuse of tree-roosts by California bats (Myotis californicus) in southern British Columbia. Am. Midl. Nat. 2001, 146, 80–85. [Google Scholar] [CrossRef]
- Lučan, R.K.; Hanák, V.; Horáček, I. Long-term re-use of tree roosts by European forest bats. For. Ecol. Manag. 2009, 258, 1301–1306. [Google Scholar] [CrossRef]
- Angers, V.A.; Drapeau, P.; Bergeron, Y. Snag degradation pathways of four North American boreal tree species. For. Ecol. Manag. 2010, 259, 246–256. [Google Scholar] [CrossRef]
- Everett, R.; Lehmkuhl, J.; Schellhaas, R.; Ohlson, P.; Keenum, D.; Riesterer, H.; Spurbeck, D. Snag dynamics in a chronosequence of 26 wildfires on the east slope of the cascade range in Washington state, USA. Int. J. Wildland Fire 1999, 9, 223–234. [Google Scholar] [CrossRef]
- DeLong, S.C.; Sutherland, G.D.; Daniels, L.D.; Heemskerk, B.H.; Storaunet, K.O. Temporal dynamics of snags and development of snag habitats in wet spruce−fir stands in east-central British Columbia. For. Ecol. Manag. 2008, 255, 3613–3620. [Google Scholar] [CrossRef]
- Klingsporn, S.; Webster, C.R.; Bump, J.K. Influence of legacy-tree retention on group-selection opening persistence. For. Ecol. Manag. 2012, 286, 121–128. [Google Scholar] [CrossRef]
- Borkin, K.M.; O’Donnell, C.; Parsons, S. Bat colony size reduction coincides with clear-fell harvest operations and high rates of roost loss in plantation forest. Biodivers. Conserv. 2011, 20, 3537–3548. [Google Scholar] [CrossRef]
- Chaverri, G.; Kunz, T.H. Response of a specialist bat to the loss of a critical resource. PLoS ONE 2011, 6, e28821. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.L.; Fenton, M.B. Responses of little brown myotis to exclusion and to bat houses. Wildl. Soc. Bull. 1994, 22, 8–14. [Google Scholar]
- Kerth, G. Causes and consequences of sociality in bats. BioScience 2008, 58, 737–746. [Google Scholar] [CrossRef]
- Johnson, J.S.; Kropczynski, J.N.; Lacki, M.J. Social network analysis and the study of sociality in bats. Acta Chiropterol. 2013, 15, 1–17. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R. Effects of hierarchical roost removal on northern long-eared bat (Myotis septentrionalis) maternity colonies. PLoS ONE 2015, 10, e0116356. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, H.J.; Nelson, C.D.; Scrivani, J.A.; Jacobs, D.F. Consequences of shifts in abundance and distribution of American chestnut for restoration of a foundation forest tree. Forests 2016, 7, 4. [Google Scholar] [CrossRef]
- Paillet, F.L. Chestnut: History and ecology of a transformed species. J. Biogeogr. 2002, 29, 1517–1530. [Google Scholar] [CrossRef]
- Wang, G.G.; Hu, H. The replacements of American chestnut: A range-wide assessment based on data from forest inventory and published studies. In Proceedings of the 17th Biennial Southern Silvicultural Research Conference; Gordon, H.A., Conner, K.F., Haywood, J.D., Eds.; General Technical Report SRS-203; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2015; pp. 513–515. [Google Scholar]
- De Bruijn, A.; Gustafson, E.J.; Kashian, D.M.; Dalgleish, H.J.; Sturtevant, B.R.; Jacobs, D.F. Decomposition rates of American chestnut (Castanea dentata) wood and implications for coarse woody debris pools. Can. J. For. Res. 2014, 44, 1575–1585. [Google Scholar] [CrossRef]
- Faison, E.K.; Foster, D.R. Did American chestnut really dominate the eastern forest? Arnoldia 2014, 72, 18–32. [Google Scholar]
- Seiler, J.; Jensen, E.; Niemiera, A.; Peterson, J. American chestnut Fagaceae Castanea dentata (Marsh.) Borkh., Ag Handbook 654, Virginia Tech Department of Forest Resources and Environmental Conservation. 2018. Available online: http://dendro.cnre.vt.edu/dendrology/syllabus/factsheet.cfm?ID=21 (accessed on 23 February 2018).
- Dalgleish, H.J.; Swihart, R.K. American chestnut past and future: Implications of restoration for resource pulses and consumer populations of eastern U.S. forests. Restor. Ecol. 2012, 20, 490–497. [Google Scholar] [CrossRef]
- Davis, D.E. Historical significance of American chestnut to Appalachian culture and ecology. In Restoration of American Chestnut to Forest Lands-Proceedings of a Conference and Workshop; Steiner, K.C., Carlson, J.E., Eds.; Natural Resources Report NPS/NCR/CUE/NRR—2006/001; National Park Service: Washington, DC, USA, 2006; pp. 53–60. [Google Scholar]
- Gravatt, F. Chestnut blight in Asia and North America. Unasylva 1949, 3, 2–7. [Google Scholar]
- O’Shea, T.J.; Ellison, L.E.; Stanley, T.R. Survival estimation in bats: Historical overview, critical appraisal, and suggestions for new approaches. In Sampling Rare or Elusive Species: Concepts, Designs, and Techniques foe Estimating Population Parameters; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 297–336. [Google Scholar]
- O’Donnell, C.F.J. Population dynamics and survivorship in bats. In Ecological and Behavioral Methods for the Study of Bats, 2nd ed.; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 158–176. [Google Scholar]
- Foster, G.; Humphrey, S.R.; Humphrey, P.P. Survival rate of young southeastern brown bats, Myotis austroriparius, in Florida. J. Mammal. 1978, 59, 299–304. [Google Scholar] [CrossRef]
- Stevenson, D.E.; Tuttle, M.D. Survivorship in the endangered gray bat (Myotis grisescens). J. Mammal. 1981, 62, 244–257. [Google Scholar] [CrossRef]
- Sedgeley, J.A.; O’Donnell, C.F.J. Roost use by long-tailed bats in South Canterbury: Examining predictions of roost-site selection in a highly fragmented landscape. N. Z. J. Ecol. 2004, 28, 1–18. [Google Scholar]
- O’Donnell, C.F.J.; Sedgeley, J.A. Causes and consequences of tree-cavity roosting in a temperate bat, Chalinolobus tuberculatus, from New Zealand. In Functional and Evolutionary Ecology of Bats; Zubaid, A., McCracken, G.F., Kunz, T.H., Eds.; Oxford University Press, Inc.: New York, NY, USA, 2006; pp. 308–328. [Google Scholar]
- O’Donnell, C.F.J. Distribution, Status and Conservation of Long-Tailed Bat (Chalinolobus tuberculatus) Communities in Canterbury, New Zealand; Environment Canterbury Report U00/38; Environment Canterbury: Christchurch, New Zealand, 2000. [Google Scholar]
- Brigham, R.M. Bats in forests: What we know and what we need to learn. In Bats in Forests: Conservation and Management; Lacki, M.J., Hayes, J.P., Kurta, A., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 1–15. [Google Scholar]
- Jachowski, D.S.; Rota, C.T.; Dobony, C.A.; Ford, W.M.; Edwards, J.W. Seeing the forest through the trees: Considering roost-site selection at multiple spatial scales. PLoS ONE 2015, 11, e0150011. [Google Scholar]
- Mori, A.S.; Kitagawa, R. retention forestry as a major paradigm for safeguarding forest biodiversity in productive landscapes: A global meta-analysis. Biol. Conserv. 2014, 175, 65–73. [Google Scholar] [CrossRef]
- Cowden, M.M.; Hart, J.L.; Schweitzer, C.J.; Dey, D.C. Effects of intermediate-scale wind disturbance on composition, structure, and succession in Quercus stands: Implications for natural disturbance-based silviculture. For. Ecol. Manag. 2014, 330, 240–251. [Google Scholar] [CrossRef]
- Puettmann, K. Silvicultural challenges and options in the context of global change: ‘simple’ fixes and opportunities for new management approaches. J. For. 2011, 109, 321–331. [Google Scholar]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Lacki, M.J.; Johnson, J.S.; Baker, M.D. Temperatures beneath bark of dead trees used as roosts by Myotis volans in forests of the Pacific Northwest, USA. Acta Chiropter. 2013, 15, 143–151. [Google Scholar] [CrossRef]
- Willis, C.K.R.; Brigham, R.M. Social thermoregulation exerts more influence than microclimate on forest roost preferences by a cavity-dwelling bat. Behav. Ecol. Sociobiol. 2007, 62, 97–108. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Dalgleish, H.J.; Nelson, C.D. A conceptual framework for restoration of threatened plants: The effective model of American chestnut (Castanea dentata) restoration. New Phytol. 2013, 197, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.M.; Drees, K.P.; Foster, J.T.; Lindner, D.L. Extreme sensitivity to ultraviolet light in the fungal pathogen causing white-nose syndrome of bats. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.L.; Schlarbaum, S.E.; Pinchot, C.C.; Anagnostakis, S.L.; Saunders, M.R.; Thomas-Van Gundy, M.; Schaberg, P.; McKenna, J.; Bard, J.F.; Berrang, P.C.; et al. Reintroduction of American chestnut in the National Forest system. J. For. 2014, 112, 502–512. [Google Scholar] [CrossRef]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar] [CrossRef]
- Mori, A.S.; Tatsumi, S.; Gustafsson, L. Landscape properties affect biodiversity response to retention approaches in forestry. J. Appl. Ecol. 2017, 54, 1627–1637. [Google Scholar] [CrossRef]
- Manning, A.D.; Fischer, J.; Lindenmayer, D.B. Scattered trees are keystone structures—Implications for conservation. Biol. Conserv. 2006, 132, 311–321. [Google Scholar] [CrossRef]
- Hallinger, M.; Johansson, V.; Schmalholz, M.; Sjöberg, S.; Ranius, T. Factors driving tree mortality in retained forest fragments. For. Ecol. Manag. 2016, 368, 163–172. [Google Scholar] [CrossRef]
- Périé, C.; de Blois, S. Dominant forest tree species are potentially vulnerable to climate change over large portions of their range even at high latitudes. PeerJ 2016, 4, e2218. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic map of North America minus Mexico, with regional groupings of states and provinces used for analyses of tree roosts of Myotis bats. Abbreviations for regions are: WCAN (western Canada), CCAN (central Canada), ECAN (eastern Canada), NW (Northwest), SW (Southwest), MW (Midwest), NE (Northeast), and SE (Southeast).
Figure 1.
Schematic map of North America minus Mexico, with regional groupings of states and provinces used for analyses of tree roosts of Myotis bats. Abbreviations for regions are: WCAN (western Canada), CCAN (central Canada), ECAN (eastern Canada), NW (Northwest), SW (Southwest), MW (Midwest), NE (Northeast), and SE (Southeast).

Figure 2.
Mean diameter (SE) of roost trees of Myotis by region across Canada and the United States.
Figure 2.
Mean diameter (SE) of roost trees of Myotis by region across Canada and the United States.
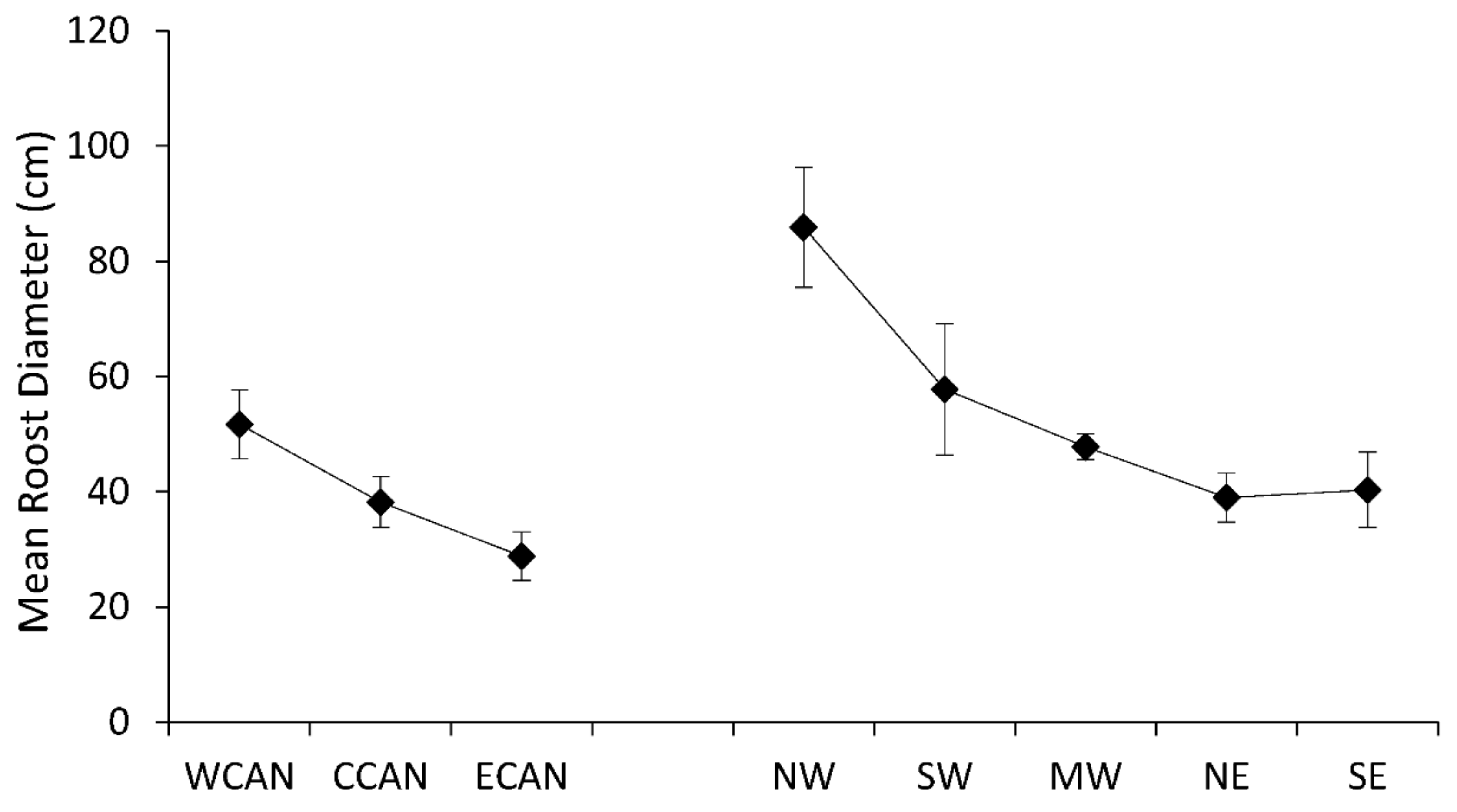
Figure 3.
Mean height (SE) of roost trees of Myotis by region across Canada and the United States.
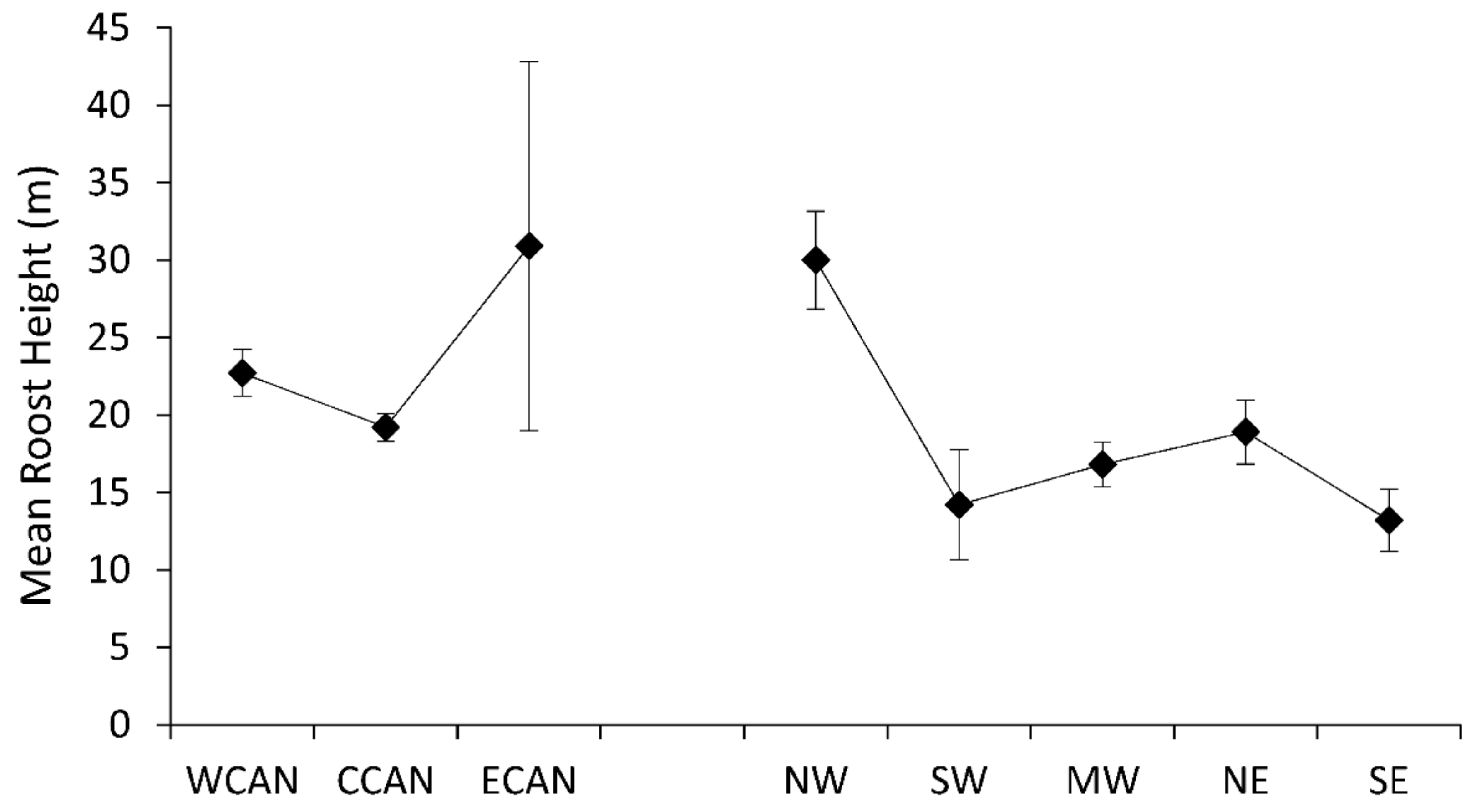
Figure 4.
Mean diameter (SE) of tree roosts of Myotis in Canada and the United States. Abbreviations for species are: MYAU (Myotis auriculus), MYAUR (M. austroriparius), MYCA (M. californicus), MYEV (M. evotis), MYKE (M. keenii), MYLU (M. lucifugus), MYSE (M. septentrionalis), MYSO (M. sodalis), MYTH (M. thysanodes), MYVO (M. volans), and MYYU (M. yumanensis).
Figure 4.
Mean diameter (SE) of tree roosts of Myotis in Canada and the United States. Abbreviations for species are: MYAU (Myotis auriculus), MYAUR (M. austroriparius), MYCA (M. californicus), MYEV (M. evotis), MYKE (M. keenii), MYLU (M. lucifugus), MYSE (M. septentrionalis), MYSO (M. sodalis), MYTH (M. thysanodes), MYVO (M. volans), and MYYU (M. yumanensis).

Figure 5.
Mean maximum exit counts from tree roosts of Myotis across Canada and the United States.
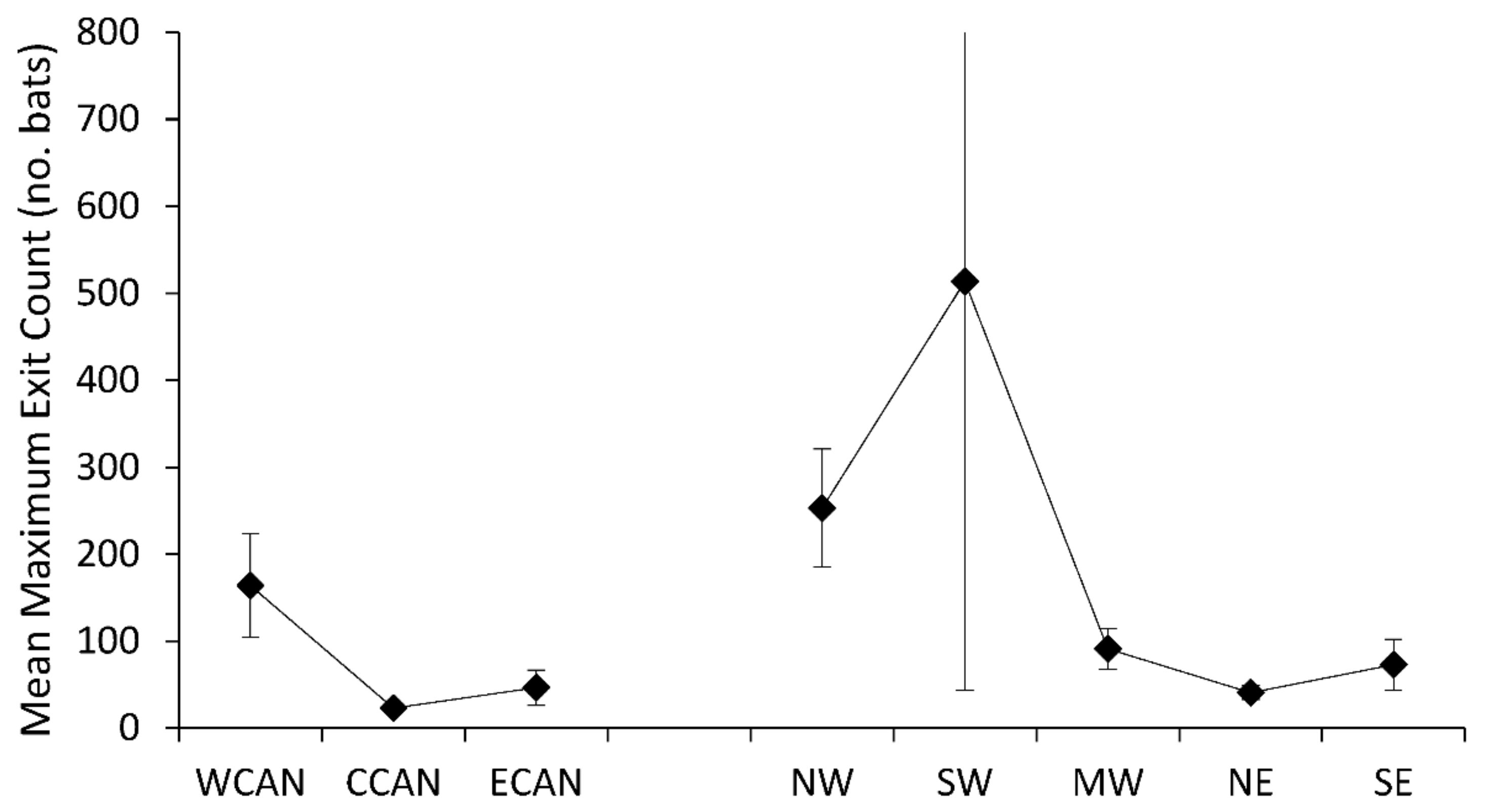
Figure 6.
Mean number of bats exiting tree roosts of Myotis across Canada and the United States.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mean (±SE) difference in diameter (%) and height (%) of roost trees from surrounding stand-level conditions for populations of Myotis bats in North America.
Table 1.
Mean (±SE) difference in diameter (%) and height (%) of roost trees from surrounding stand-level conditions for populations of Myotis bats in North America.
| % Difference | % Difference | |||
|---|---|---|---|---|
| Bat Species | na | in Diameter | n | in Height |
| M. austroriparius | 4 | 0.9 (0.25) | 2 | 0.85 (0.6) |
| M. auriculus | 1 | 1.06 (–) | 1 | 1.3 (–) |
| M. californicus | 2 | 1.77 (0.23) | 2 | 1.58 (0.005) |
| M. evotis | 1 | 0.83 (–) | 1 | 0.75 (–) |
| M. keenii | 2 | 1.7 (0.4) | ||
| M. lucifugus | 4 | 1.12 (0.06) | 3 | 1.1 (0.06) |
| M. septentrionalis | 14 | 1.33 (0.12) | 11 | 2.02 (0.84) |
| M. sodalis | 8 | 1.72 (0.23) | 3 | 1.73 (0.38) |
| M. thysanodes | 3 | 1.41 (0.08) | 3 | 1.62 (0.51) |
| M. volans | 4 | 1.29 (0.09) | 4 | 1.69 (0.26) |
| M. yumanensis | 1 | 3.74 (–) | 1 | 2.02 (–) |
a Indicates number of populations.
Table 2.
Linear regressions of maximum exit counts (No. bats) and mean exit counts (No. bats/count) with characteristics of tree roosts of Myotis across Canada and the United States.
Table 2.
Linear regressions of maximum exit counts (No. bats) and mean exit counts (No. bats/count) with characteristics of tree roosts of Myotis across Canada and the United States.
| Adj-R2 | Parameter | β | SE | F | p | |
|---|---|---|---|---|---|---|
| Maximum | 0.0162 | Roost diameter | −1.347 | 1.23 | 1.2 | 0.297 |
| 0.0771 | Roost height | 8.748 | 7.35 | 1.42 | 0.299 | |
| 0.1221 | Roost diameter | −1.825 | 1.66 | 1.1 | 0.352 | |
| Roost height | 12.061 | 7.78 | 1.55 | 0.218 | ||
| Mean | −0.0021 | Mean diameter | 0.223 | 0.23 | 0.94 | 0.341 |
| −0.0155 | Mean height | 0.451 | 0.55 | 0.66 | 0.424 | |
| −0.0662 | Mean diameter | 0.016 | 0.38 | 0.04 | 0.967 | |
| Mean height | 0.421 | 0.92 | 0.46 | 0.653 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lacki, M.J. Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests. Diversity 2018, 10, 29. https://doi.org/10.3390/d10020029
AMA Style
Lacki MJ. Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests. Diversity. 2018; 10(2):29. https://doi.org/10.3390/d10020029
Chicago/Turabian StyleLacki, Michael J. 2018. "Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests" Diversity 10, no. 2: 29. https://doi.org/10.3390/d10020029
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.