3. Results
Aphanommata Wollaston, 1873
Aphanommata Wollaston, 1873: 463. Type species Rhyncolus euphorbiarum Wollaston, 1868.
Brachytemnoides Folwaczny, 1973: 155. Type species: Rhyncolus filum Mulsant and Rey, 1859, synonymy in Alonso-Zarazaga, 1989: 325.
Macrancyloides Champion, 1909: 75. Type species: Macrancyloides perlongus Champion, 1909, synonymy by Kuschel in Wibmer and O’Brien, 1986: 5.
Oocorynus Champion, 1909: 76. Type species: Oocorynus corrosus Champion, 1909, synonymy by Kuschel in Wibmer and O’Brien, 1986: 5.
Rhamphocolus Casey, 1892: 702. Type species: Rhamphocolus tenuis Casey, 1892, synonymy by Kuschel in Wibmer and O’Brien, 1986: 5.
Aphanommata Wollaston: Alonso-Zarazaga, 1989: 325 (review).
The genus Aphanommata is readily distinguishable from other genera of the tribe Rhyncolini by the combination of following character states: (1) body in cross section circular; (2) head narrow, conical, tapered anteriad, rostrum longer than wide; (3) eyes not prominent; (4) antennae short and stout, funicule with seven antennomeres; (5) basal margin of pronotum straight; (6) humeri well-defined, elytra parallel-sided; (7) scutellum visible and (8) apical tarsomere expanded to apex.
Aphanommata euphorbiarum (Wollaston, 1867)
Rhyncolus euphorbiarum Wollaston, 1867: 119. Type locality: interior of Santo Antão, 4 syntypes in the Ribeira Fria and 1 syntypus in Ribeira da Babosa.
Aphanommata euphorbiarum: Alonso-Zarazaga, 1989: 327 (redescription, illustration of habitus, lateral view of head and aedeagus).
Type material studied: Lectotype, sex not determined, here designated: (h) euphorbia/round label with red margin (p) Type/(p) T. V. Wollaston Coll., B. M. 1867-82, CAPE VERDE IS./original white label with a green corner. BMNH. Paralectotype, ♂, hardly damaged specimen, aedeagus and some other parts mounted in Euparal: (p) T. V. Wollaston Coll., B. M. 1867-82, CAPE VERDE IS. BMNH.
Remark I. Green corner on the original label is a mark of Wollaston and means that the specimen was collected on the island Santo Antão.
Diagnosis: Smaller species with body length under 3.5 mm, maximum width of elytra 0.96 mm, colour reddish-brown. Antennal scape slightly pedunculate at apex, short, shorter than funicule, funicule with 7 antennomeres; funicular antennomere I slightly longer than wide, about 1.5 times as long as II; antennal club about 1.4 times as long as wide; head and pronotum shiny, finely shagreened, evenly punctate with fine punctures; anterior margin of pronotum slightly narrower than base; eyes oval, flat, about 1.7 times higher than wide, temples short, width of eye equal to distance from margin of eye to pronotal margin (Figure 4f); pronotum widest in posterior third; elytra with 15 rows of punctures of different size, in sutural region cross-cracked, elytral intervals flat.
Differential diagnosis. Aphanommata euphorbiarum is distinguishable from its Old World congeners by the combination of the following character states: (1) body large, more than 3 mm but less than 3.5 mm long (Figure 1a); (2) pronotum with fine punctures, distance between punctures always superior to diameter of punctures (Figure 6c); (3) sides of pronotum evenly rounded (Figure 6c); (4) pronotum smooth (Figure 6c); (5) apex of elytra with very small erect setae (Figure 8a); (6) funicular antennomere I parallel-sided (Figure 1a) and (7) different shape of apex of aedeagus (Alonso-Zarazaga, 1989: 328, Figures 3 and 4). Aphanommata euphorbiarum is readily separated from A. kuscheli sp. nov. and A. strakai sp. nov. by different size, the absence of setae on the apex of elytra and the shape of aedeagus.
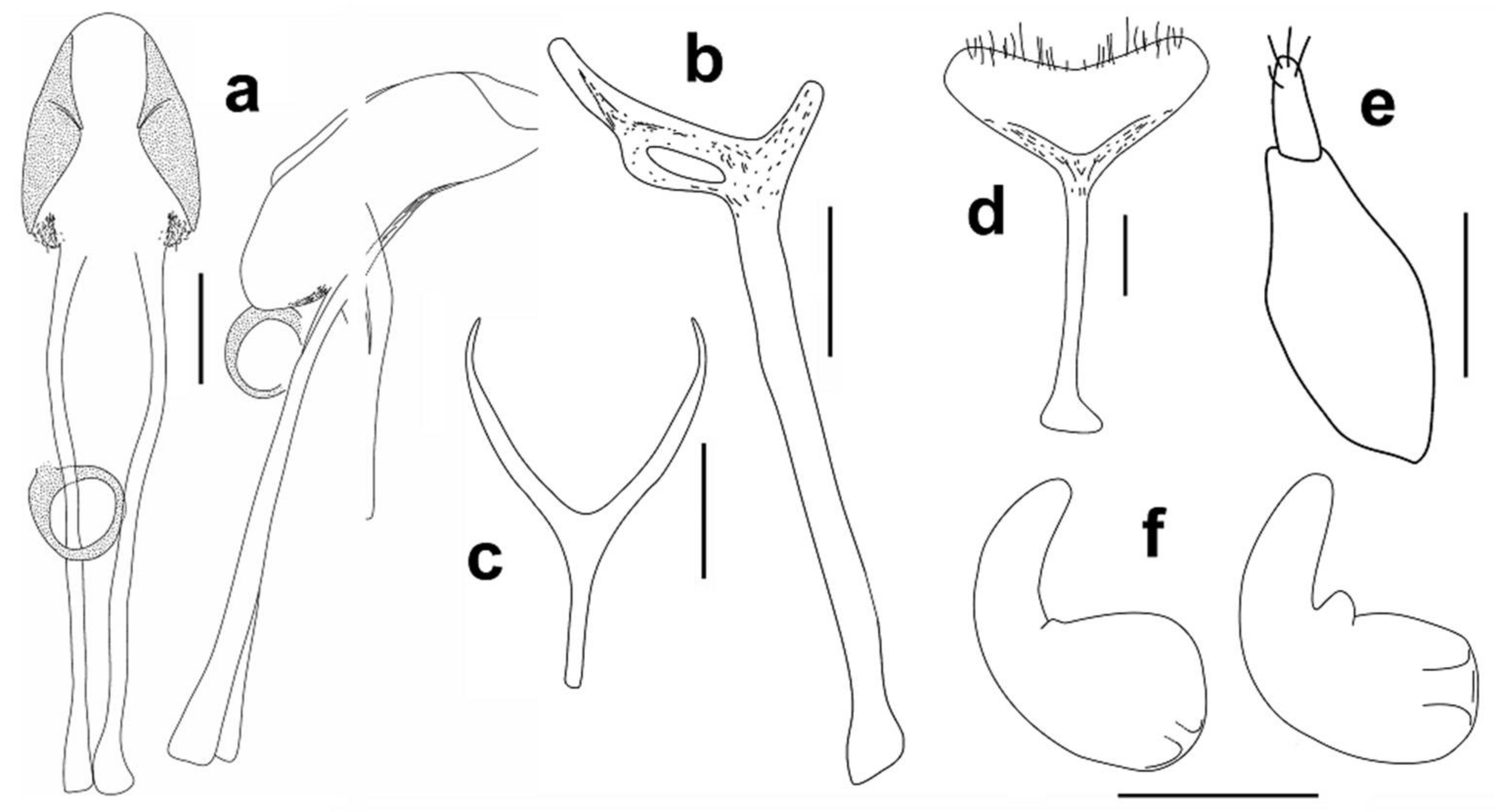
Remark II. The species A. euphorbiarum has been redescribed by Alonso-Zarazaga [7] according to one male from the syntype series and distinguished in the key from A. filum (Mulsant and Rey, 1858) which were in the same work transferred in this genus due to the synonymy of the genus Brachytemnoides Folwaczny, 1973 to Aphanommata. One of the main diagnostic characters cited for A. euphorbiarum was the rostrum with the shallow median sulcus. We have examined 2 syntypes deposited in BMNH and did not find such a sulcus on the rostrum. The rostrum of both specimens examined by us is simple, lacking any trace of the sulcus, with only the sparse puncturation (Figure 4e,f). As the aedeagus illustrated by Alonso-Zarazaga [7] well corresponds with the aedeagus studied by us, we believe that the sulcus on the rostrum could be more an artefact than the case of interspecific variability, but more material will be needed to definitely solve this question. Remark III. A. euphorbiarum has been described according to five specimens sifted by V. Wollaston on two localities in the interior of the island Santo Antão [3:120]. We were searching all syntypes, but it seems that the specimen studied by Alonso-Zarazaga [7] as well as other two syntypes were lost, they are neither in BMNH (Max Barclay, per. comm.) nor in OUMNH (Amoret Spooner, per. comm.) where Wollaston’s collection is deposited. Biology: Syntype series were collected from the decayed stem of the Euphorbia tuckeyana.
Distribution: Cape Verde (Santo Antão).
Aphanommata kuscheli sp. nov.
Adult material studied
Type material. HOLOTYPE, ♂: CAPE VERDE Isl., 10.X.2013, SAO NICOLAU, W, Mt. Gordo summit, 16.625089, -24.350854, J. Straka and J. Batelka lgt. (p)/windward slopes in rotten wood of Euphorbia tuckeyana (p) (SMNS). PARATYPES: 47 ex.: the same data as holotype. SMNS (18), BMNH (3), OUMNH (2), NMPC (2), CJS (10), CPH (10), CJB (2). Non-type material. 3♀: SAO NICOLAU, W.-S of Cachaco, 13.XI.2011, 16°37′ N, 24°21′ W, J. Straka and J. Batelka lgt., in rotten wood of Agave sisalana. CJS, CPH, CJB.
Remark IV. Specimens from São Nicolau, W-S of Cachaco are not included in the type series due to the lack of a male.
Diagnosis. Antennal scape slightly pedunculate at apex, short, shorter than funicule, funicule with 7 antennomeres; funicular antennomere I slightly longer than wide, about 1.75 times as long as II; antennal club about 1.7 times as long as wide; head and pronotum shiny, finely shagreened, evenly puncturate with fine punctures; anterior margin of pronotum slightly narrower than base; pronotum widest in posterior fourth; elytra with 14–16 rows of punctures of different size, in anterior half cross-cracked, elytral intervals flat.
Description (Figure 2a–c). Body convex, shiny, pitch-black, sometimes elytra brownish, tarsi and antennae brownish (Figure 2a). Length 3.60–4.45 mm, maximum width of elytra 1.08–1.30 mm.
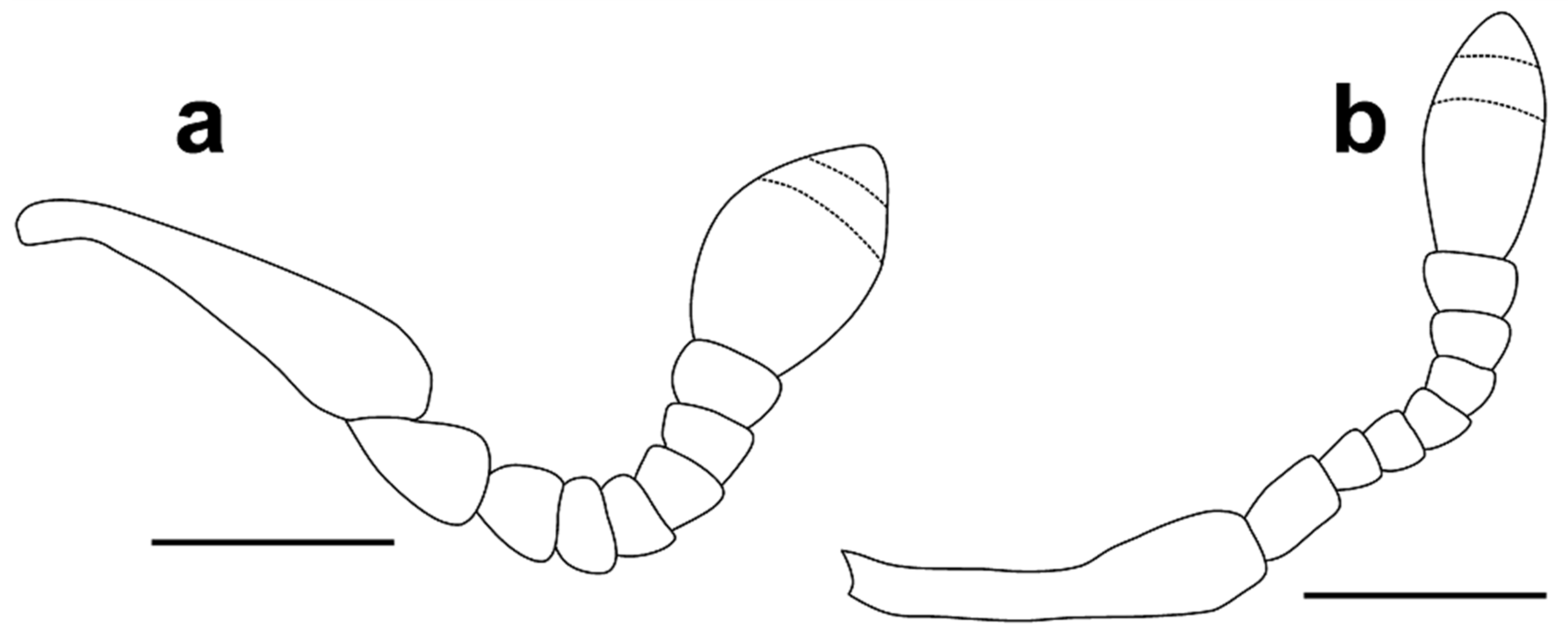
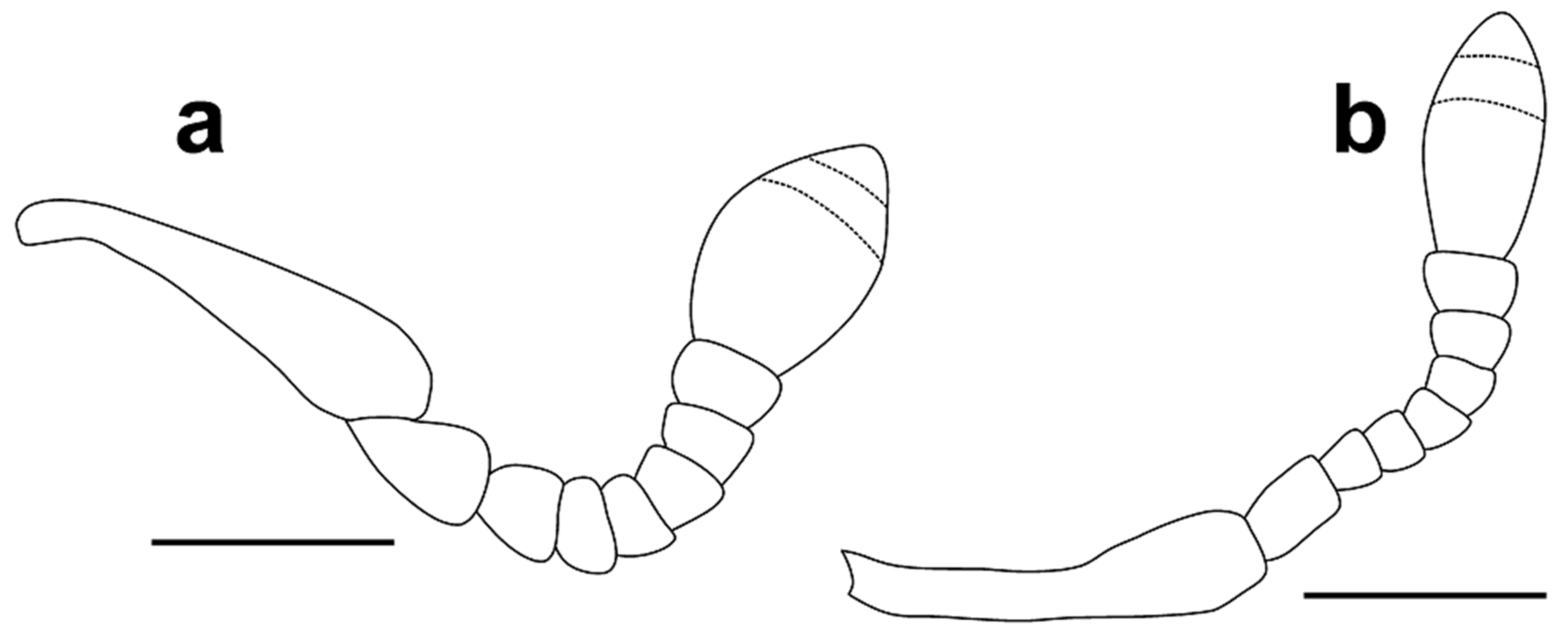
Head 1.15–1.25 times as long as wide, with sparse, fine punctures. Rostrum slightly convex, more than three times as long as head behind eyes (Figure 4a,b). Antennal scrobe lateral, not visible in dorsal view, short, deep, curved downward before eye, the point of antennal insertion located on anterior third of head (Figure 4a,b). Antennae (Figure 5a) stout, with long, sparse setae, antennal club with dense, golden pubescence on apical margin of first and second antennomeres of club; antennal scape short, pedunculate at apex, about 4.20–4.60 times as long as funicular antennomere I and shorter than funicule; funicule with seven antennomeres, funicular antennomere I expanded to apex, 1.40 times as long as wide, slightly longer and wider than II, later about 1.60 times as long as III, III–VII transverse, V–VII subequal, slightly longer than III and IV, antennal club with three antennomeres, first clearly longest, club apically pointed, 1.50–1.60 times as long as wide. Eyes oval, flat, about 1.50 times higher than wide, temples long, width of eye inferior, about 1.5 times, to distance from margin of eye to pronotal margin (Figure 4b).
Pronotum (Figure 6a) with dense, fine, even punctures, pronotal punctures equal in size to punctures on head but puncturation denser, distance between punctures considerably superior to diameter of punctures. Pronotum 1.00–1.05 times as long as wide, 1.15–1.30 times as long as head, widest in posterior fourth, basal margin straight. Scutellum well-defined, oval, wider than long.
Elytra (Figure 7a) 1.80–1.95 times as long as wide, 2.10–2.30 times as long as pronotum, convex, fused, surface smooth and shiny, with about 14–16 striae formed by rows of punctures of different size, distance between punctures varies but always superior to diameter of puncture, in anterior half cross-cracked, elytral intervals flat, humeri prominent, humeral calli well-defined. Hind wings absent.
Prosternum at the same level as meso and metaventrite, finely shagreened, with very few uneven punctures, long, disc depressed in anterior part, depression with transverse furrow, anterior margin finely dentate; procoxae separated by narrow isthmus, hypomerae fused with median part of prosternum; mesoventrite short, about 2.50 times shorter than metaventrite, both finely shagreened, shiny, mesoventrite roughly punctured mainly in anterior part, anterior margin with golden setae, mesoventral process long, wide, truncate, shagreened and punctured, metaventrite evenly punctured, mesocoxae separated, isthmus about as wide as half of coxa, metacoxae strongly separated by large process of first ventrite, puncturation on metaventrite denser than on ventrite I and II, ventrite I about 1.35 times as long as ventrite II, all ventrites shagreened and punctured, posterior part of ventrite I and anterior part of ventrite II medially depressed, ventrites III and IV subequal in length, ventrite V semicircular in shape, more than three times as long as ventrite IV.
Legs (Figure 2a–c) brownish, finely shagreened, tibiae with strongly arcuate uncus and well-defined mucro, fore tibiae with dense, golden setation on inner part of apical half of tibia, apex of mid and hind tibiae decent setae, tarsi with four tarsomeres, tarsomere I long, about as long as II–III combined, III bilobed in apical half, onychium long, about as long as I–III combined, with two free, divaricate, tarsal claws equal.
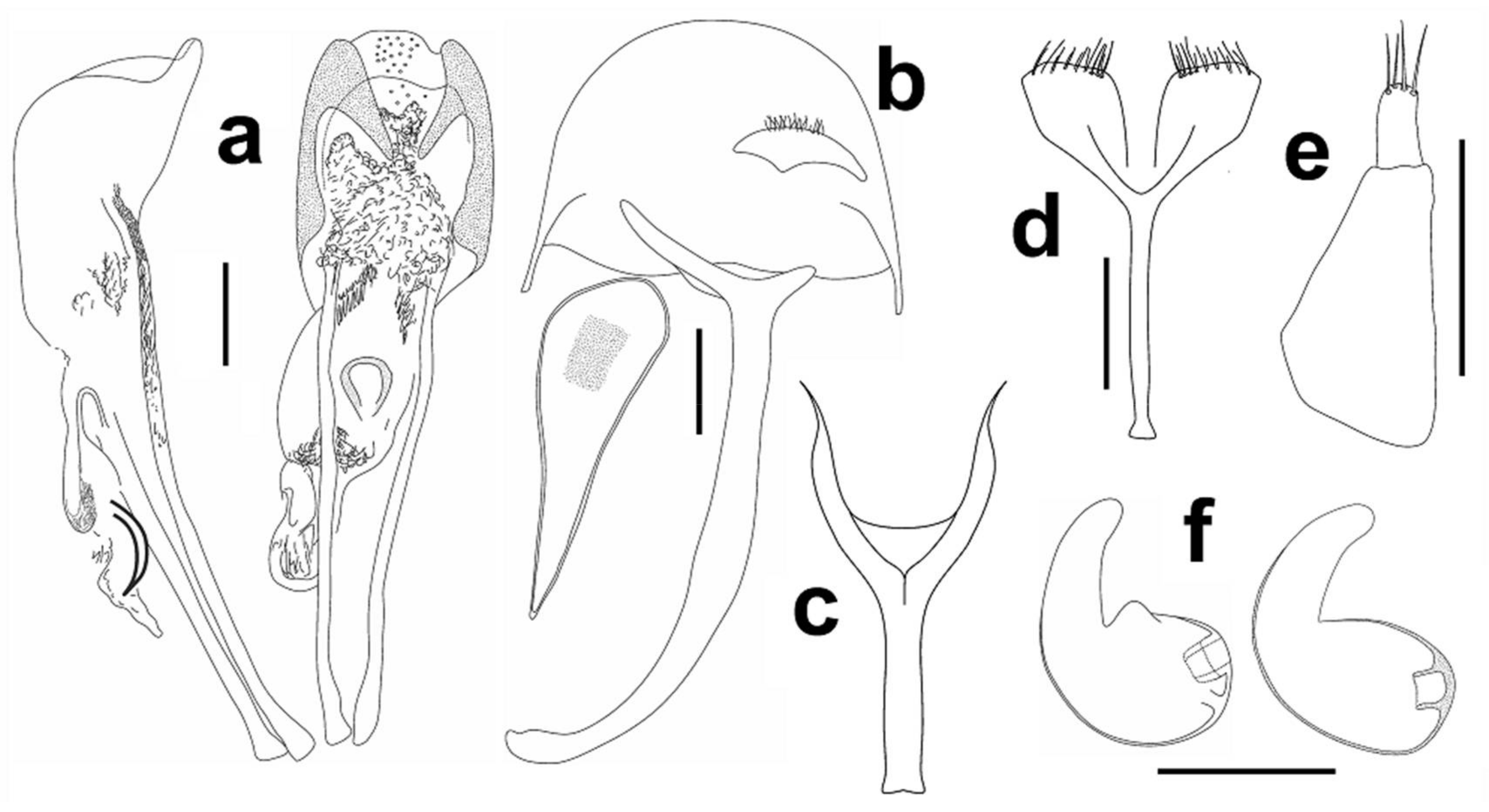
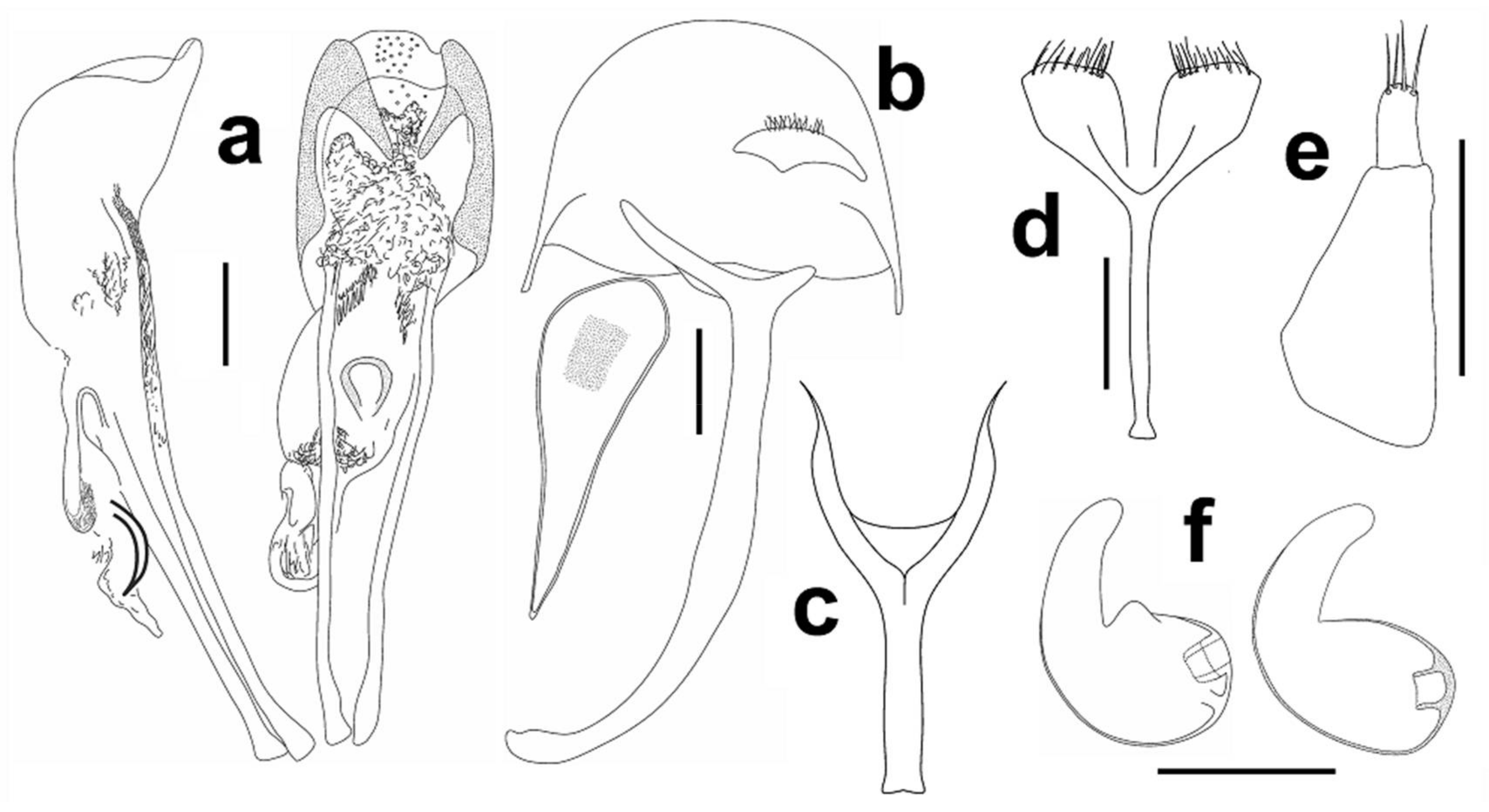
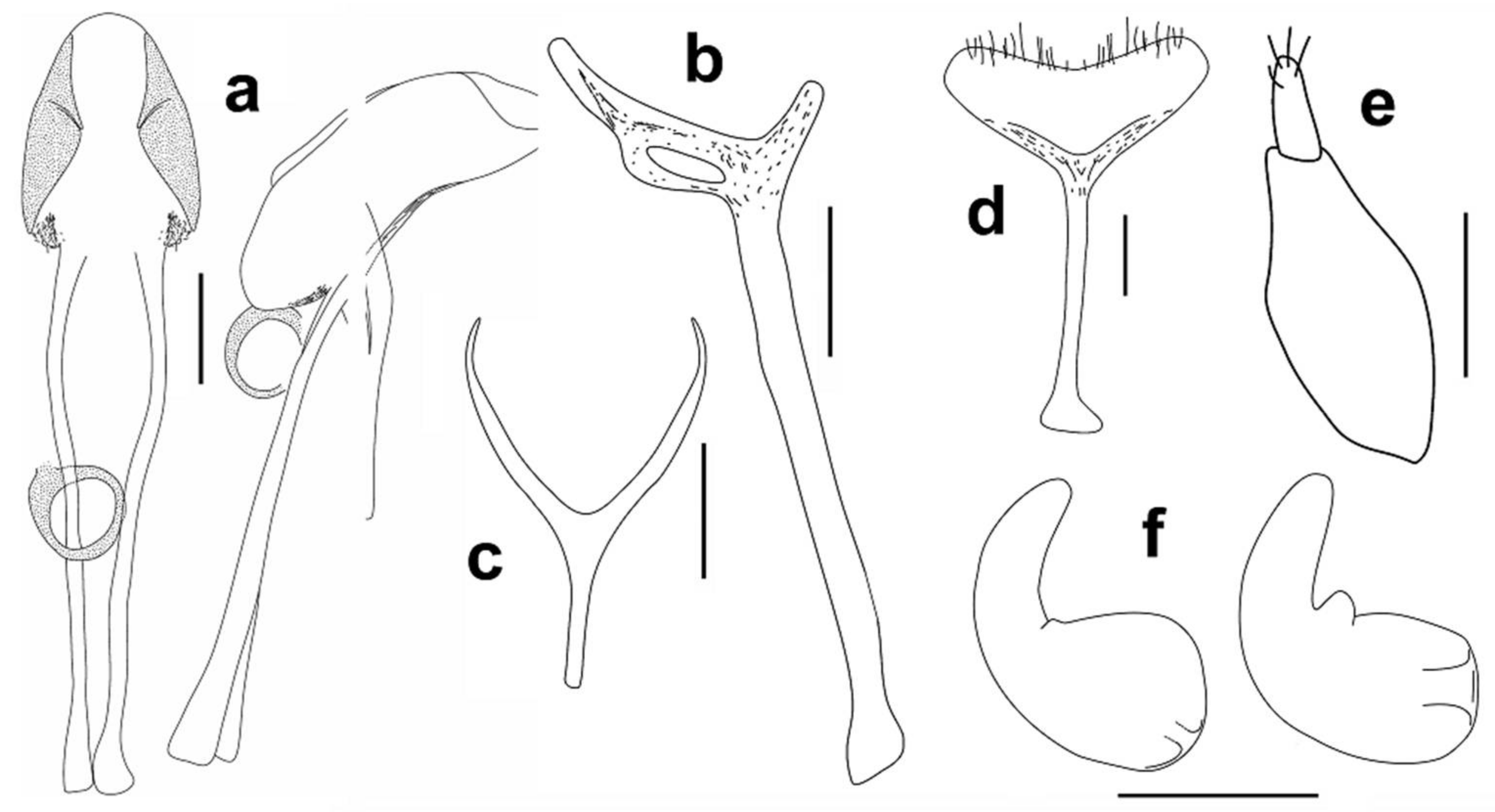
Male genitalia. Aedeagus (Figure 3a), male sternite 9 (spiculum gastrale), hemisternite and rectal loop (Figure 3b).
Female genitalia. Spermatheca (Figure 3f), female sternite 8 (spiculum ventral) (Figure 3d), gonocoxite (Figure 3e).
Sexual dimorphism. Ventrites I and II in females simple, lacking median depression.
Differential diagnosis. Aphanommata kuscheli sp. nov. is distinguishable from its Old World congeners by the combination of the following character states: (1) body large, more than 4 mm long; (2) pronotum with fine punctures, distance between punctures always superior to diameter of punctures (Figure 6a); (3) sides of pronotum evenly rounded (Figure 6a); (4) pronotum shagreened (Figure 6a); (5) apex of elytra with any erect setae (Figure 7a); (6) funicular antennomere I expanded to apex (Figure 5a); (7) different shape of apex of aedeagus (Figure 3a); and (8) apex of cornu on spermatheca slightly bent (Figure 3f). Aphanommata kuscheli sp. nov. is readily separated from A. strakai sp. nov. by the different surface of pronotum, the shape of antenna, aedeagus and spermatheca; and from A. euphorbiarum by different size, the absence of setae on the apex of elytra and the shape of aedeagus.
Etymology. Patronymic, named after Guillermo Kuschel, an eminent Curculionidae worker who has also contributed significantly to knowledge of the subfamily Cossoninae.
Description of mature larva
Material examined. 2 mature larvae: Cape Verde Isl., São Nicolau W—Mt. Gordo Summit, 10.x.2013, 16.625089 N, 24.35084 W, J. Straka and J. Batelka lgt., windward slopes, in rotten wood of Euphorbia tuckeyana Steud. (Euphorbiaceae).
Measurements (in mm). Body length: 5.48–6.15. The widest body part (abdominal segments II–VI) 1.47. Head width: 0.87–0.96.
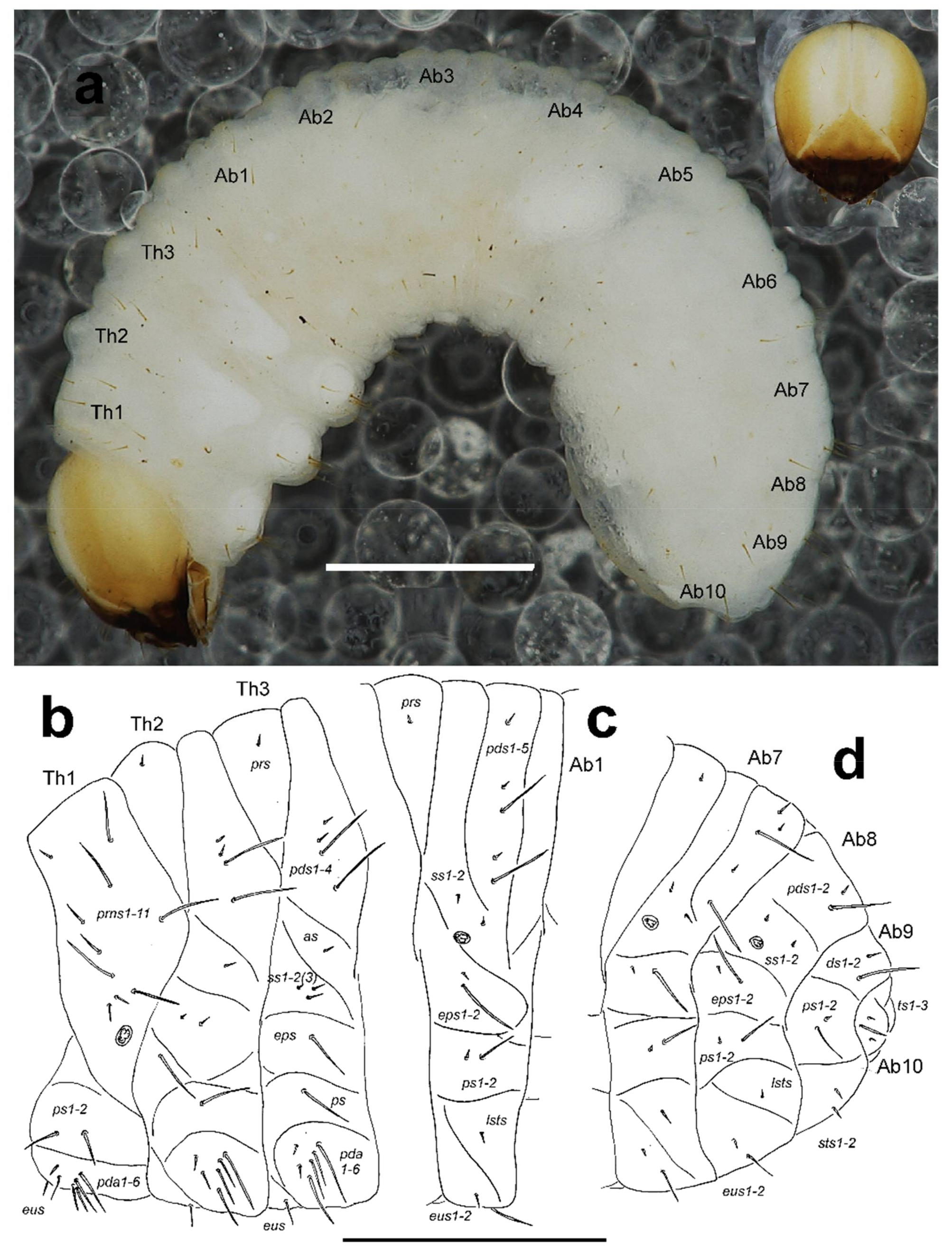
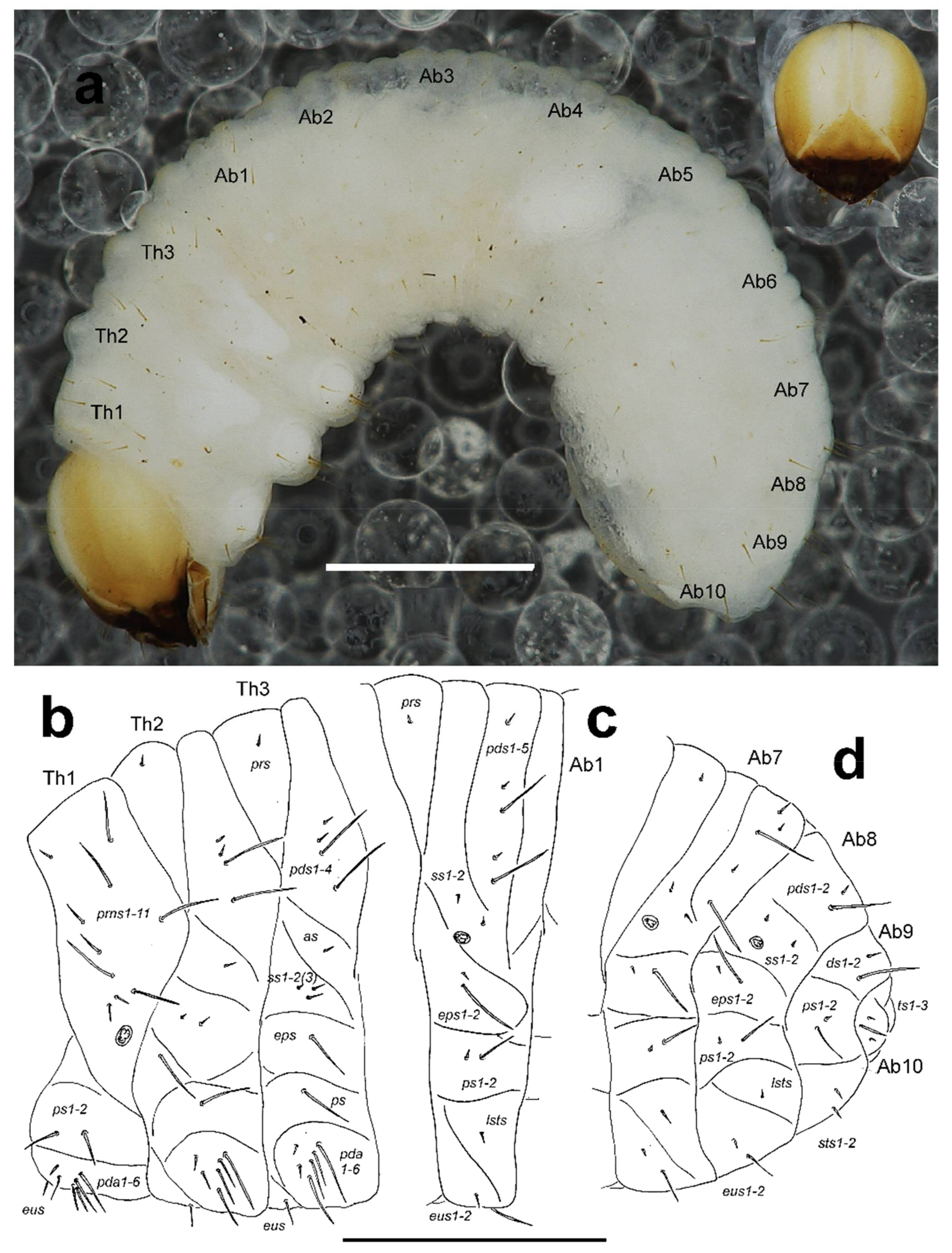
General. Body elongate, slightly curved, rounded in cross section (Figure 10a).
Colouration. Dark yellow to pale brown head around suture white (Figure 10a). All thoracic and abdominal segments white and dorsum with fine speckling (Figure 10a).
Vestiture. Setae on body very long to very short, orange and clearly visible.
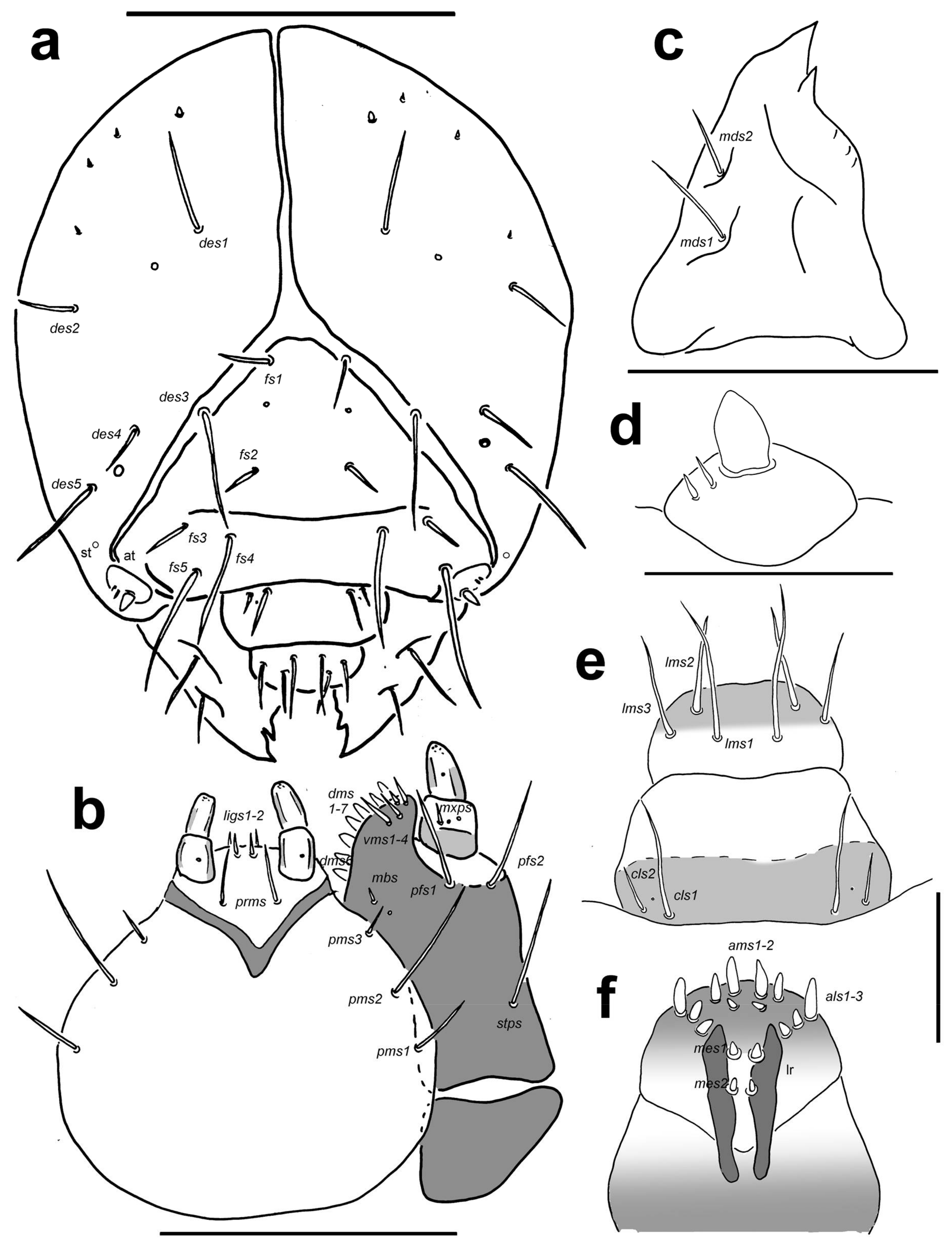
Head capsule (Figure 9a). Head suboval and distinctly rounded laterally, endocarinal line absent. Frontal sutures on head very broad and well visible, extended to antennae. One stemmata (st) feebly visible, in the form of a minute pigmented spot with convex cornea, located on each side anterolaterally, behind antenna. Setae on head piliform, varying in length, from very long to short. Des1 and des2 located behind the middle part of the central part of epicranium, very long des1 near to the middle part of epicranium, and medium des2 near to side of epicranium, very long des3 located anteriorly on epicranium in or very close to frontal suture, medium des4 close to des3, very long des5 located anterolaterally (Figure 9a); des2 and des4 as long as third of length of remaining three des. Dorsal part of epicranium with 2 sensilla; one between des1 and des2, and next one close to des4. Fs1 of medium length located posteriomedially, and almost in the frontal suture, fs2 located medially, fs3 and fs4 located anterolaterally; and fs5 close to antennae; fs1, fs2 and fs4 as long as des4, and fs3 and fs5 very long as long as des5 (Figure 9a). Les1 and les2 as long as des1; both ves short, as long as third length of les. Epicranial area with 3–4 pes.
Antennae located at the end of the frontal suture on each side, membranous and conical basal article bearing one conical triangular sensorium located centrally; basal membranous article with 2(3) very short basiconic sensillae (Figure 9d).
Clypeus (Figure 9e) slightly trapezium-shaped, anterior margin of clypeus slightly concave; approximately 2 times as wide as long; cls1 very long, placed posteromedially, cls2 of medium length to long, localized posterolaterally; 1 sensillum clss placed between cls1 and cls2.
Mouth parts. Labrum (Figure 9e) also slightly trapezium-shaped, approximately twice as wide as long, with 3 piliform lms, of different lengths; lms1 very long, lms2 and lms3 as long as two third of length of lms1; all lms protrudes well over the anterior margin of labrum; lms1 placed medially in the central part of labrum, lms2 located anteromedially and lms3 located anterolaterally; anterior margin bi-sinuate. Epipharynx (Figure 9f) with 3 finger-like als, als1 more than twice as long as als2 and als3, all in line to labral rods; with 3 ams piliform, ams1 as long as als1, ams2 as long as half or one third length of ams1, and ams3 very short as als2 and als3; and with 2 very short to minute, finger-like mes; labral rods (lr) narrow, elongate, parallel. Mandibles (Figure 9c) slightly curved, distinctly broad, with divided apex; bifid, teeth of unequal length; slightly truncate; mds1 very long, and mds2 long, piliform, located in distinct holes. Maxilla (Figure 9b) stipes with 1 stps, 2 pfs and 1 mbs with one sensillum close to mbs, stps and pfs1–2 very long, mbs minute to very short; mala with 7 finger-like dms; 4 vms, different in length, one very short seta, one short and two as long as dms; 2 vms distinctly shorter than dms. Maxillary palpi with two palpomeres; basal palpomere with 1 relatively long mxps and two sensilla; length ratio of basal and distal palpomeres: 1:1; distal palpomere with one sensillum and a group of conical, cuticular apical processes. Prelabium (Figure 9b) oval-shaped and feebly elongate, with 1 long prms; ligula with sinuate margin, 2 relatively long to short ligs, and one sensillum; premental sclerite feebly visible. Labial palpi with two palpomeres; length ratio of basal and distal palpomeres, basal palpomere much wider than distal: 1:1.3; distal palpomere elongate, with one sensillum and short, cuticular apical processes; basal palpomere with 1 dorsal sensillum.
Postlabium (Figure 9b) with 3 pms, all pms located laterally; pms1 and pms2 very long, but pms1 as long as two third of pms2, pms3 long, more than 2 times shorter than pms1.
Thorax. Prothorax smaller than meso- and metathorax. Spiracle bicameral, placed between the pro- and mesothorax (see Skuhrovec et al. [
9]). Prothorax (Figure 10b) with 10 short to very long
prns (3 placed apically (one short and 2 very long); 1 or 2 very long and 2 relatively long medially; 4 form a group close to spiracle, 2 very long, and 2 short); 2 very long to long
ps; and 1 relatively long
eus. Mesothorax (Figure 10b) with 1 short to very short
prs, 4
pds different in length (order: very short, very short, very long, very long); 1 short to very short
as; 1 very short and 1–2 short to relatively long
ss; 1 very long
eps; 1 very long
ps; and 1 long to very long
eus. Chaetotaxy of metathorax (Figure 10b) almost identical to that of mesothorax. Each pedal area of thoracic segments well separated, with 4 very long and 2 relatively long to short
pda.
Abdomen. Abdominal segments I–VI of almost equal length, remaining abdominal segments decreasing in width gradually posterad. Abdominal segment X reduced to four anal lobes of unequal size, the dorsal being distinctly the largest, the lateral pair equal in size, and the ventral lobe very small. Anus located terminally; ambulatory ampullae bilobate to circular. Spiracles bicameral, the eight abdominal spiracles located medially, close to the anterior margin of abdominal segments I–VIII. Abdominal segments I–VII (Figure 10c) with 1 very short to short prs; 5 pds different in length (order: short, very short, very long, very short, very long); 2 short ss; 1 very short and 1 very long eps; 1 very short and 1 very long ps; 1 relatively long lsts; and 1 relatively long and 1 very long eus. Abdominal segment VIII (Figure 10d) with 2 pds different in length (order: short, very long); 2 short ss; 1 very short and 1 very long eps; 1 very short and 1 very long ps; 1 relatively long lsts; and 1 relatively long and 1 very long eus. Abdominal segment IX (Figure 10d) with 1 short to relatively long and 1 very long ds, located close to posterior margin of segment; 1 very long and 1 very short ps; and 2 relatively long to short sts. Abdominal segment X (Figure 10d) with 3 ts, 1 long and 2 very short to minute.
Collection circumstances. Imagines and larva collected in 2011 south-west of Cachaco were extracted from the outer layer of the rotten stem of Agave sisalana, growing by the mountain trail circa 1050 m a.s.l. More specimens of the species were obtained in 2013 from the rotten stem of Euphorbia tuckeyana on the northern slope of Monte Gordo, not far from the summit, some 1200 m a.s.l. The stem of Euphorbia was much eaten by larvae of the species, imagines were in the larval galleries and pupal chambers.
Distribution: Cape Verde (São Nicolau)
Aphanommata strakai sp. nov.
(Figures 4c,d, 5b, 6b, 7b, 11a–c and 12a–f)
Type material studied. HOLOTYPE, ♂: CAPE VERDE Isl., FOGO—Chã des Caldeiras 8.–9.X.2009, J. Straka and J. Batelka lgt. (p) (SMNS). PARATYPES: 9 ex.: the same data as holotype. SMNS (1), BMNH (1), OUMNH (1), NMPC (1), CJS (2), CPH (2), CJB (1).
Diagnosis. Antennal scape slightly pedunculate at apex, short, shorter than funicule, funicule with seven antennomeres; funicular antennomere I about 1.60 times as long as wide, almost parallel-sided, about 1.75 times as long as II; antennal club about twice as long as wide; head and pronotum shiny, head finely shagreened, pronotom not shagreened, evenly puncturate with fine punctures, punctures on pronotum bigger; elytra with 14–15 rows of punctures of different size, finely cross-cracked in anterior half only close to sutura, elytral intervals flat; anterior margin of pronotum slightly narrower than base; pronotum widest in posterior fourth.
Description (
Figure 11a–c)
. Body convex, shiny, reddish-brown to dark reddish-brown, tarsi and antennae of the same colour (Figure 11a–c). Length 4.00–4.25 mm, maximal width of elytra 1.10–1.20 mm.
Head 1.15–1.20 times as long as wide, with sparse, fine punctures. Rostrum slightly convex, more than three times as long as head behind eyes (Figure 4c,d). Antennal scrobe lateral, not visible in dorsal view, short, deep, curved downward before eye, the point of antennal insertion located on anterior third of head (Figure 4c,d). Antennae (Figure 5b) stout, with long, sparse setae, antennal club with dense, golden pubescence on apical margin of first and second antennomeres of club; antennal scape short, pedunculate at apex, about 4.65–4.75 times as long as funicular antennomere I and shorter than funicule; funicule with seven antennomeres, funicular antennomere I parallel-sided, 1.60 times as long as wide and 1.75 times as long as II, later about 1.30 times as long as III, III–VII transverse, V–VI subequal, VII about 1.2 times as wide as VI, antennal club with three antennomeres, first clearly longest, club apically pointed, 1.40–1.50 times as long as wide. Eyes oval, flat, about 1.2–1.3 times higher than wide, temples long, width of eye inferior, about 1.33 times, to distance from margin of eye to pronotal margin (Figure 4d).
Pronotum (Figure 6b) with dense, fine, even punctures, pronotal punctures larger and denser than punctures on head, distance between punctures varies considerably, from inferior, equal to clearly superior to diameter of punctures, pronotum 1.00–1.05 times as long as wide, 1.10–1.25 times as long as head, widest in posterior fourth, basal margin straight. Scutellum well-defined, oval, wider than long.
Elytra (Figure 7b) 1.95–2.05 times as long as wide, 2.30–2.45 times as long as pronotum, convex, fused, surface smooth and shiny, with about 14–16 striae formed by punctures of different size, distance between punctures varies but always superior to diameter of puncture, in anterior half cross-cracked only close to suture, elytral intervals flat, humeri prominent, humeral calli well-defined. Hind wings absent.
Prosternum at the same level as meso and metaventrite, finely shagreened, with uneven punctures, long, disc depressed in anterior part, depression with transverse furrow, anterior margin finely dentate; procoxae separated by narrow isthmus, hypomerae fused with median part of prosternum; mesoventrite long, about 1.75 times shorter than metaventrite, both finely shagreened, shiny, mesoventrite roughly punctured mainly in anterior part, anterior margin with golden setae, mesoventral process long, wide, truncate, shagreened and punctured, metaventrite sparsely unevenly punctured, mesocoxae separated, isthmus about as wide as half of coxa, metacoxae strongly separated by large process of first ventrite, puncturation on metaventrite about as dense as on ventrite I and II, ventrite I about 1.30 times as long as ventrite II, all ventrites shagreened and punctured, posterior part of ventrite I and anterior part of ventrite II medially depressed, ventrites III and IV subequal in length, ventrite V semicircular in shape, more than three times as long as ventrite IV.
Legs (Figure 11a–c) brownish, finely shagreened, tibiae with strongly arcuate uncus and well-defined mucro, fore tibiae with dense, golden setation on inner part of apical half of tibia, apex of mid and hind tibiae decent setae, tarsi with four tarsomeres, tarsomere I long, about as long as II–III combined, III bilobed in apical half, onychium long, about as long as I–III combined, with two free, divaricate, tarsal claws equal.
Male genitalia. Aedeagus (
Figure 12a), male sternite 9 (spiculum gastrale) (Figure 12b).
Female genitalia. Spermatheca (Figure 12f), female sternite 8 (spiculum ventral) (Figure 12d), gonocoxite (Figure 12e).
Sexual dimorphism. Ventrites I and II in females simple, lacking median depression.
Differential diagnosis. Aphanommata strakai sp. nov. is distinguishable from its Old World congeners by the combination of the following character states: (1) body large, more than 4 mm long; (2) pronotum with fine punctures, distance between punctures always superior to diameter of punctures (Figure 6b); (3) sides of pronotum evenly rounded (Figure 6b); (4) pronotum smooth (Figure 6b); (5) apex of elytra with any erect setae (Figure 7b); (6) funicular antennomere I parallel-side (Figure 5b); (7) different shape of apex of aedeagus (Figure 12a); and (8) apex of cornu on spermatheca straight (Figure 12f). Aphanommata strakai sp. nov. is readily separated from A. kuscheli sp. nov. by the different surface of pronotum, the shape of antenna, aedeagus and spermatheca; and from A. euphorbiarum by the different size, the absence of setae on the apex of elytra and the shape of aedeagus.
Etymology. The species name is dedicated to the Czech entomologist Jakub Straka (Praha, Charles University) in appreciation of his friendship and his energetic field research on the Cape Verde Curculionidae.
Collection circumstances. All adults were collected under the bark of dead Euphorbia tuckeyana on the floor of the caldera of the Fogo volcano, about 2 km SW of Portela village, circa 1710 m a.s.l.
Distribution. Cape Verde (Fogo).
Key to Aphanommata species of Old World
1 Small species, body length 2.30–2.80 mm (Figure 1c,d). Pronotum with rough, dense punctures, the distance between punctures inferior to the diameter of punctures (Figure 6d). Sides of pronotum parallel (Figure 6d). Distribution: known from southern Europe, and Algeria. ……………………………………………………………………………… A. filum (Mulsant and Rey)
- Larger species, body length more than 3 mm (Figure 1a,b; Figure 2a–c; Figure 11a–c). Pronotum with fine punctures, distance between punctures always superior to the diameter of punctures (Figure 6a–c). Sides of pronotum evenly rounded (Figure 6a–c). Distribution: only on the Cape Verde islands. ………………………………………………………………………………………… 2
2 Smaller species, body length less than 3.5 mm (Figure 1a,b). The apex of elytra with small erect setae (Figure 8a). ………………………………………………………… A. euphorbiarum (Wollaston)
- Larger species, body length more than 4.0 mm (Figure 2a–c; Figure 11a–c). The apex of elytra lacking erect setae (Figure 7a,b). ……………………………………………………………………… 3
3 Pronotum shagreened (Figure 6a). Funicular antennomere I expanded to apex, 1.40 times as long as wide, slightly longer and wider than II, later about 1.60 times as long as III (Figure 5a). The apex of cornu on spermatheca slightly bent (Figure 3f). ………………………… A. kuscheli sp. nov.
- Pronotum smooth, not shagreened (Figure 6b). Funicular antennomere I parallel-sided, 1.60 times as long as wide and 1.75 times as long as II, later about 1.30 times as long as III (Figure 5b). The apex of cornu on spermatheca straight (Figure 12f). ……………………… A. strakai sp. nov.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}