
Oxidative Stress Markers and Sperm DNA Fragmentation in Men Recovered from COVID-19
 ,
,
Abstract
:1. Introduction
2. Results
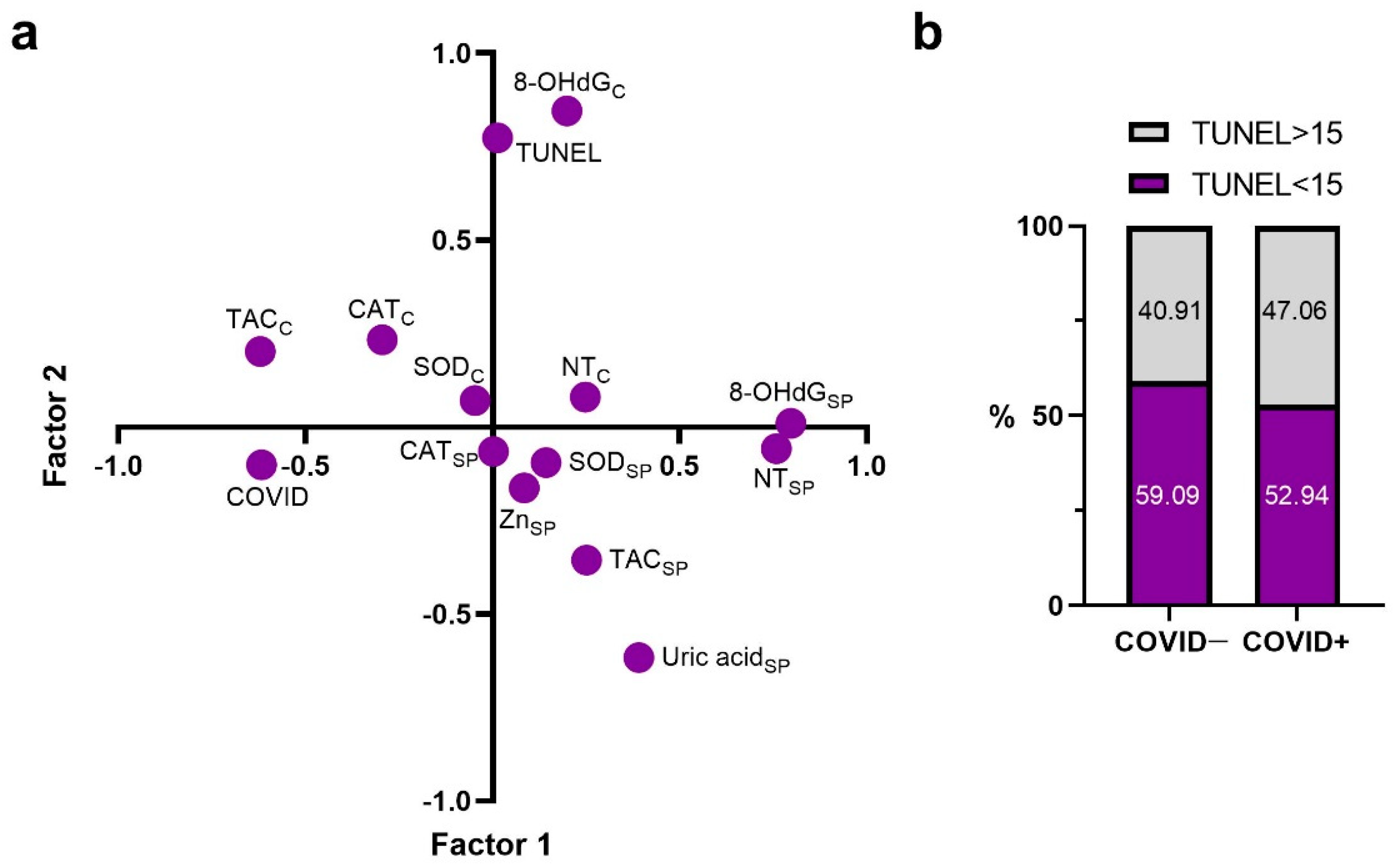
2.1. Principal Components
2.2. Correlations with TUNEL and COVID Factors
2.3. Semen Quality Parameters
2.4. Antioxidant System Components
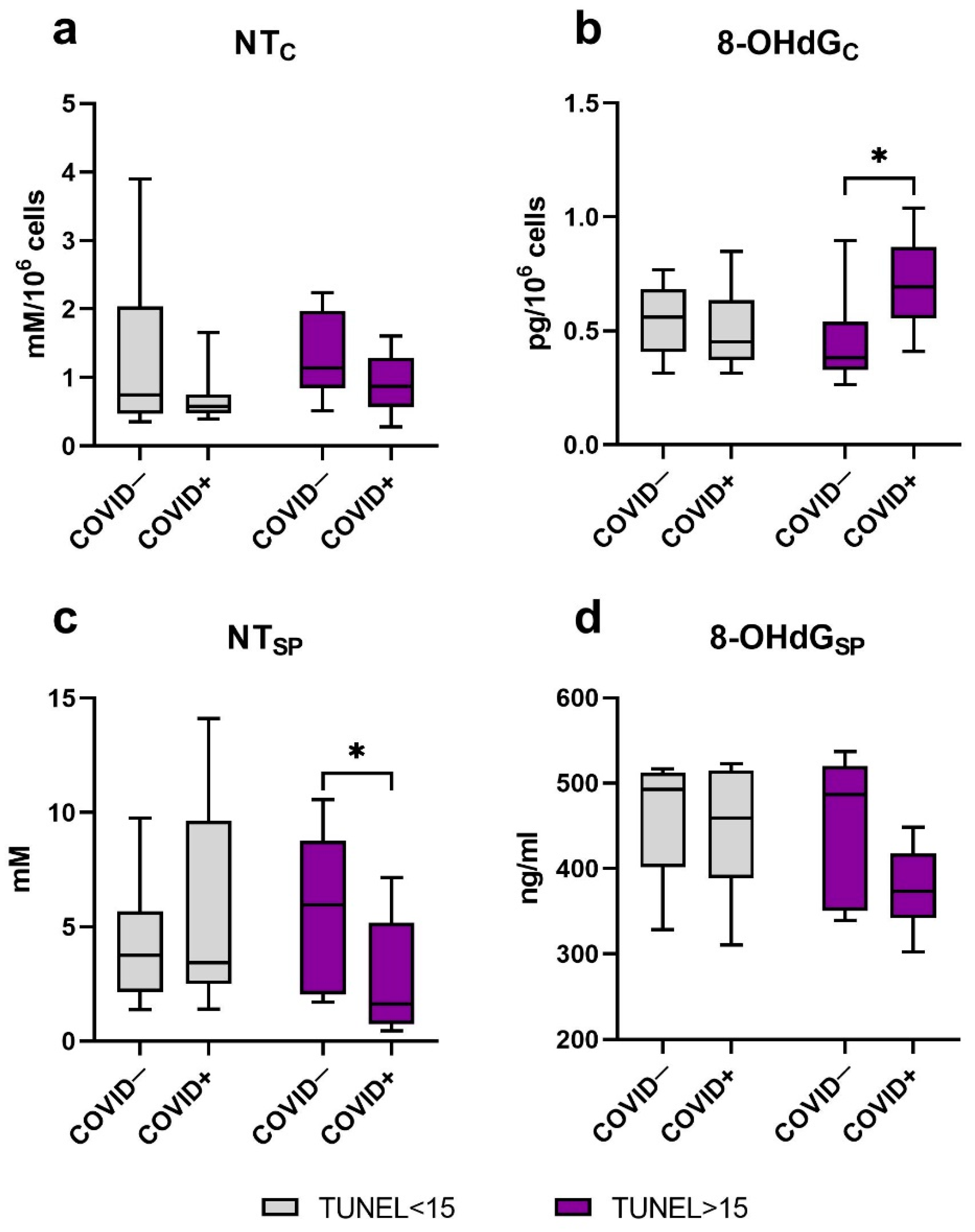
2.5. Oxidative Modification of DNA and Proteins
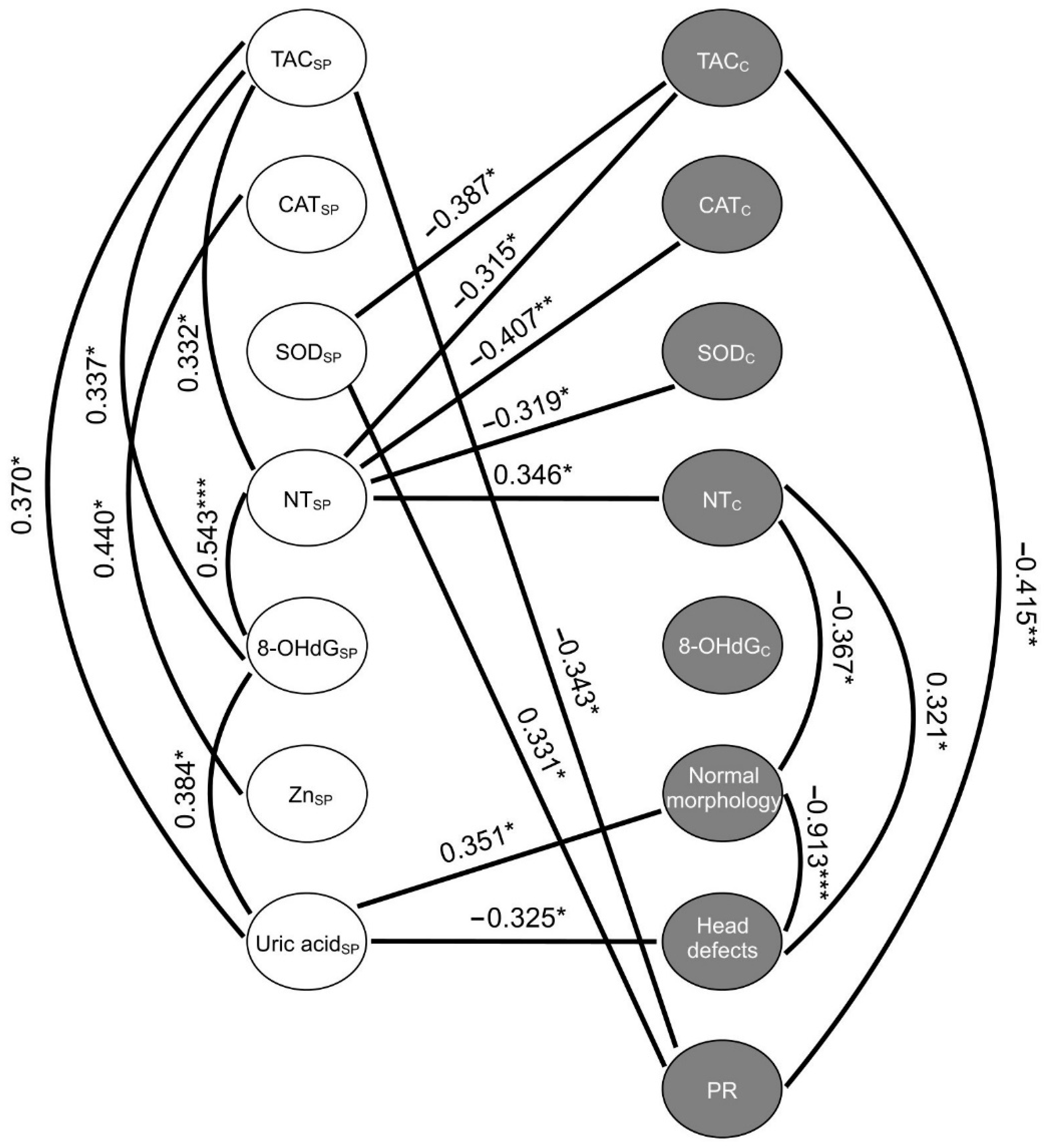
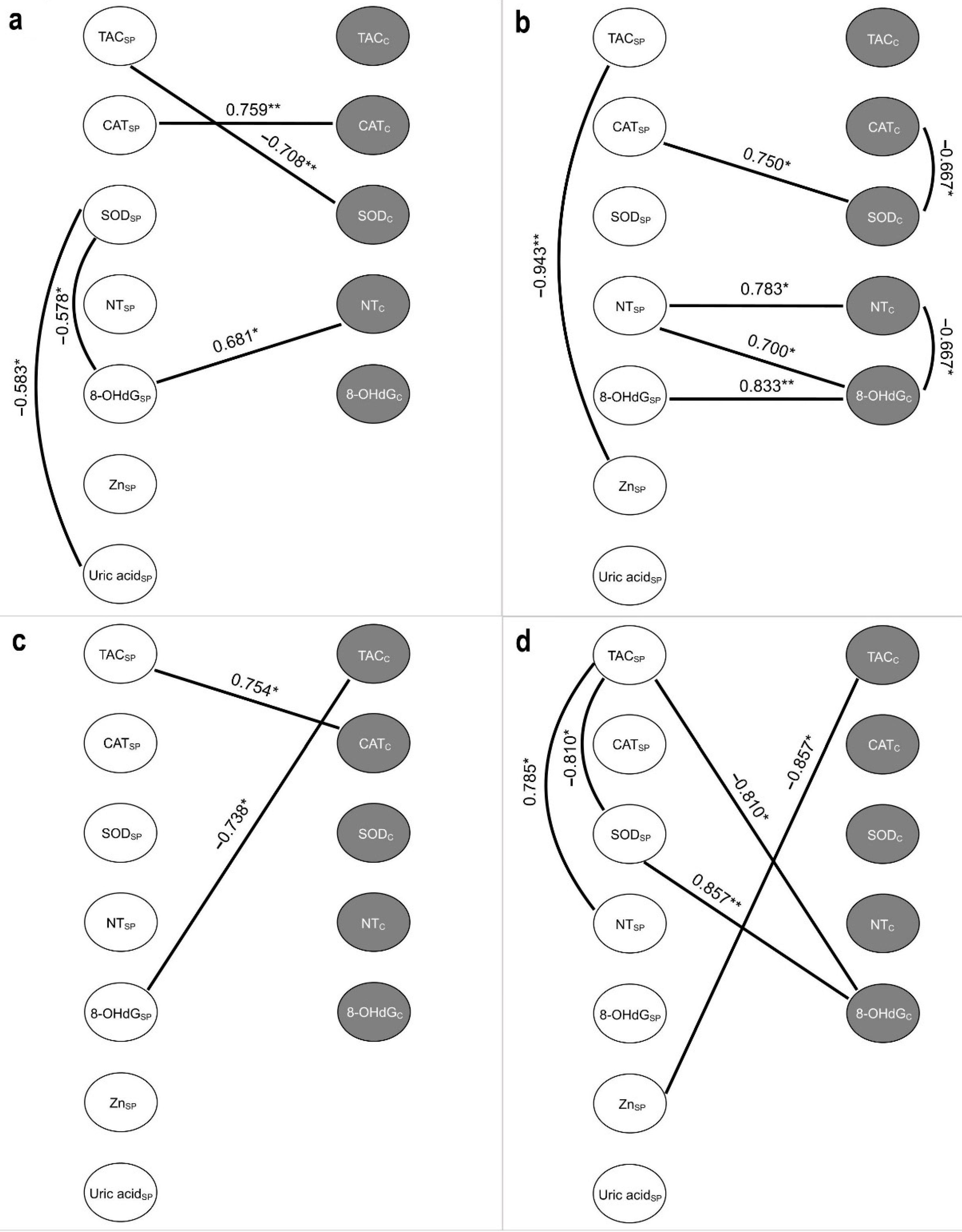
2.6. Correlations between the Studied Variables of Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Semen Analysis and Preparation
4.3. Sperm DNA Fragmentation
4.4. Assessment of Antioxidant Markers
4.4.1. Total Antioxidant Capacity
4.4.2. Superoxide Dismutase Activity
4.4.3. Catalase Activity
4.5. Assessment of Oxidative Modifications of Macromolecules
4.5.1. Nitrotyrosine Content
4.5.2. Oxidative DNA Damage
4.6. Zinc Analysis in Seminal Plasma
4.7. Uric acid Analysis in Seminal Plasma
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Xu, X. scRNA-seq Profiling of Human Testes Reveals the Presence of the ACE2 Receptor, A Target for SARS-CoV-2 Infection in Spermatogonia, Leydig and Sertoli Cells. Cells 2020, 9, 920. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Jin, M.; Bao, P.; Zhao, W.; Zhang, S. Clinical Characteristics and Results of Semen Tests Among Men With Coronavirus Disease 2019. JAMA Netw. Open 2020, 3, e208292. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xiao, X.; Zhang, J.; Zafar, M.I.; Wu, C.; Long, Y.; Lu, W.; Pan, F.; Meng, T.; Zhao, K.; et al. Impaired spermatogenesis in COVID-19 patients. EClinicalMedicine 2020, 28, 100604. [Google Scholar] [CrossRef] [PubMed]
- Paoli, D.; Pallotti, F.; Colangelo, S.; Basilico, F.; Mazzuti, L.; Turriziani, O.; Antonelli, G.; Lenzi, A.; Lombardo, F. Study of SARS-CoV-2 in semen and urine samples of a volunteer with positive naso-pharyngeal swab. J. Endocrinol. Investig. 2020, 43, 1819–1822. [Google Scholar] [CrossRef] [PubMed]
- Saylam, B.; Uguz, M.; Yarpuzlu, M.; Efesoy, O.; Akbay, E.; Çayan, S. The presence of SARS-CoV-2 virus in semen samples of patients with COVID-19 pneumonia. Andrologia 2021, 53, e14145. [Google Scholar] [CrossRef]
- Ma, L.; Xie, W.; Li, D.; Shi, L.; Mao, Y.; Xiong, Y.; Zhang, Y.; Zhang, M. Effect of SARS-CoV-2 infection upon male gonadal function: A single center-based study. medRxiv 2020. [Google Scholar] [CrossRef]
- Hajizadeh Maleki, B.; Tartibian, B. COVID-19 and male reproductive function: A prospective, longitudinal cohort study. Reproduction 2021, 161, 319–331. [Google Scholar] [CrossRef]
- Xie, Y.; Mirzaei, M.; Kahrizi, M.S.; Shabestari, A.M.; Riahi, S.M.; Farsimadan, M.; Roviello, G. SARS-CoV-2 effects on sperm parameters: A meta-analysis study. J. Assist. Reprod. Genet. 2022, 39, 1555–1563. [Google Scholar] [CrossRef]
- Ruan, Y.; Hu, B.; Liu, Z.; Liu, K.; Jiang, H.; Li, H.; Li, R.; Luan, Y.; Liu, X.; Yu, G.; et al. No detection of SARS-CoV-2 from urine, expressed prostatic secretions, and semen in 74 recovered COVID-19 male patients: A perspective and urogenital evaluation. Andrology 2020, 9, 99–106. [Google Scholar] [CrossRef]
- Hu, B.; Liu, K.; Ruan, Y.; Wei, X.; Wu, Y.; Feng, H.; Deng, Z.; Liu, J.; Wang, T. Evaluation of mid- and long-term impact of COVID-19 on male fertility through evaluating semen parameters. Transl. Androl. Urol. 2022, 11, 159–167. [Google Scholar] [CrossRef]
- Shi, Q.-H.; Weng, J.-P.; Guo, T.-H.; Sang, M.-Y.; Bai, S.; Ma, H.; Wan, Y.-Y.; Jiang, X.-H.; Zhang, Y.-W.; Xu, B.; et al. Semen parameters in men recovered from COVID-19. Asian J. Androl. 2021, 23, 479. [Google Scholar] [CrossRef]
- Donders, G.G.G.; Bosmans, E.; Reumers, J.; Donders, F.; Jonckheere, J.; Salembier, G.; Stern, N.; Jacquemyn, Y.; Ombelet, W.; Depuydt, C.E. Sperm quality and absence of SARS-CoV-2 RNA in semen after COVID-19 infection: A prospective, observational study and validation of the SpermCOVID test. Fertil. Steril. 2022, 117, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Bronson, R.; Smith, T.B.; De Iuliis, G.N. The source and significance of DNA damage in human spermatozoa; a commentary on diagnostic strategies and straw man fallacies. Mol. Hum. Reprod. 2013, 19, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Greco, E.; Scarselli, F.; Iacobelli, M.; Rienzi, L.; Ubaldi, F.; Ferrero, S.; Franco, G.; Anniballo, N.; Mendoza, C.; Tesarik, J. Efficient treatment of infertility due to sperm DNA damage by ICSI with testicular spermatozoa. Hum. Reprod. 2005, 20, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Moskovtsev, S.I.; Jarvi, K.; Mullen, J.B.M.; Cadesky, K.I.; Hannam, T.; Lo, K.C. Testicular spermatozoa have statistically significantly lower DNA damage compared with ejaculated spermatozoa in patients with unsuccessful oral antioxidant treatment. Fertil. Steril. 2010, 93, 1142–1146. [Google Scholar] [CrossRef]
- Saleh, R.A.; Agarwal, A. Oxidative stress and male infertility: From research bench to clinical practice. J. Androl. 2002, 23, 737–752. [Google Scholar]
- Moghimi, N.; Eslami Farsani, B.; Ghadipasha, M.; Mahmoudiasl, G.-R.; Piryaei, A.; Aliaghaei, A.; Abdi, S.; Abbaszadeh, H.-A.; Abdollahifar, M.-A.; Forozesh, M. COVID-19 disrupts spermatogenesis through the oxidative stress pathway following induction of apoptosis. Apoptosis 2021, 26, 415–430. [Google Scholar] [CrossRef]
- Uribe, P.; Boguen, R.; Treulen, F.; Sánchez, R.; Villegas, J.V. Peroxynitrite-mediated nitrosative stress decreases motility and mitochondrial membrane potential in human spermatozoa. MHR Basic Sci. Reprod. Med. 2015, 21, 237–243. [Google Scholar] [CrossRef]
- Cassina, A.; Silveira, P.; Cantu, L.; Montes, J.M.; Radi, R.; Sapiro, R. Defective Human Sperm Cells Are Associated with Mitochondrial Dysfunction and Oxidant Production1. Biol. Reprod. 2015, 93. [Google Scholar] [CrossRef]
- Aktan, G.; Doğru-Abbasoğlu, S.; Küçükgergin, C.; Kadıoğlu, A.; Özdemirler-Erata, G.; Koçak-Toker, N. Mystery of idiopathic male infertility: Is oxidative stress an actual risk? Fertil. Steril. 2013, 99, 1211–1215. [Google Scholar] [CrossRef]
- Kalezic, A.; Macanovic, B.; Garalejic, E.; Korac, A.; Otasevic, V.; Korac, B. Level of NO/nitrite and 3-nitrotyrosine in seminal plasma of infertile men: Correlation with sperm number, motility and morphology. Chem.-Biol. Interact. 2018, 291, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Hammadeh, M.; Hamad, M.; Montenarh, M.; Fischer-Hammadeh, C. Protamine contents and P1/P2 ratio in human spermatozoa from smokers and non-smokers. Hum. Reprod. 2010, 25, 2708–2720. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.K.; Zieschang, J.-A.; Clark, A.M. Oxidative deoxyribonucleic acid damage in sperm has a negative impact on clinical pregnancy rate in intrauterine insemination but not intracytoplasmic sperm injection cycles. Fertil. Steril. 2011, 96, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Micillo, A.; Vassallo, M.R.C.; Cordeschi, G.; D’Andrea, S.; Necozione, S.; Francavilla, F.; Francavilla, S.; Barbonetti, A. Semen leukocytes and oxidative-dependent DNA damage of spermatozoa in male partners of subfertile couples with no symptoms of genital tract infection. Andrology 2016, 4, 808–815. [Google Scholar] [CrossRef]
- Aitken, R.J.; De Iuliis, G.N.; Finnie, J.M.; Hedges, A.; McLachlan, R.I. Analysis of the relationships between oxidative stress, DNA damage and sperm vitality in a patient population: Development of diagnostic criteria. Hum. Reprod. 2010, 25, 2415–2426. [Google Scholar] [CrossRef]
- Berby, B.; Bichara, C.; Rives-Feraille, A.; Jumeau, F.; Pizio, P.D.; Sétif, V.; Sibert, L.; Dumont, L.; Rondanino, C.; Rives, N. Oxidative Stress Is Associated with Telomere Interaction Impairment and Chromatin Condensation Defects in Spermatozoa of Infertile Males. Antioxidants 2021, 10, 593. [Google Scholar] [CrossRef]
- Muratori, M.; Tamburrino, L.; Marchiani, S.; Cambi, M.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E. Investigation on the Origin of Sperm DNA Fragmentation: Role of Apoptosis, Immaturity and Oxidative Stress. Mol. Med. 2015, 21, 109–122. [Google Scholar] [CrossRef]
- Falahieh, F.M.; Zarabadipour, M.; Mirani, M.; Abdiyan, M.; Dinparvar, M.; Alizadeh, H.; Paktinat, S.; Hosseinirad, H. Effects of moderate COVID-19 infection on semen oxidative status and parameters 14 and 120 days after diagnosis. Reprod. Fertil. Dev. 2021, 33, 683–690. [Google Scholar] [CrossRef]
- Zhang, H.; He, J.; Zhao, Y.; Wu, Q.; Zou, T.; Sun, J.; Zhu, H.; Wang, X.; Sun, F.; Xing, J.; et al. Effect of glutathione S-transferase gene polymorphisms on semen quality in patients with idiopathic male infertility. J. Int. Med. Res. 2021, 49, 03000605211061045. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Coskun, S.; Al-Rouqi, R.; Al-Rajudi, T.; Eltabache, C.; Abduljabbar, M.; Al-Hassan, S. Oxidative stress and DNA damage status in couples undergoing in vitro fertilization treatment. Reprod. Fertil. 2021, 2, 117–139. [Google Scholar] [CrossRef]
- Alahmar, A.T.; Singh, R. Comparison of the effects of coenzyme Q10 and Centrum multivitamins on semen parameters, oxidative stress markers, and sperm DNA fragmentation in infertile men with idiopathic oligoasthenospermia. Clin. Exp. Reprod. Med. 2022, 49, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Cao, X.; Pang, D.; Li, C.; Luo, Q.; Zou, Y.; Feng, B.; Li, L.; Cheng, A.; Chen, Z. Is male infertility associated with increased oxidative stress in seminal plasma? A-meta analysis. Oncotarget 2018, 9, 24494–24513. [Google Scholar] [CrossRef]
- Kehinde, E.; Anim, J.; Oriowo, M.; Memon, A.; Omu, A.; Al-Azemi, M.; Al-Maghrebi, M.; Mathew, C.; Omu, F. Molecular basis for the effects of zinc deficiency on spermatogenesis: An experimental study in the Sprague-dawley rat model. Indian J. Urol. 2015, 31, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dong, X.; Hu, X.; Long, Z.; Wang, L.; Liu, Q.; Sun, B.; Wang, Q.; Wu, Q.; Li, L. Zinc levels in seminal plasma and their correlation with male infertility: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 9932. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Zaman, S.; Sajjad, M.; Shoaib, M.; Gilani, G. Assessment of the level of trace element zinc in seminal plasma of males and evaluation of its role in male infertility. Int. J. Appl. Basic Med. Res. 2011, 1, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.S. COVID-19 pandemic: Can zinc supplementation provide an additional shield against the infection? Comput. Struct. Biotechnol. J. 2021, 19, 1371–1378. [Google Scholar] [CrossRef]
- Huang, L.; Yao, G.; Huang, G.; Jiang, C.; Li, L.; Liao, L.; Yuan, G.; Shang, L.; Xu, W. Association of Zinc deficiency, oxidative stress and increased double-stranded DNA breaks in globozoospermic infertile patients and its implication for the assisted reproductive technique. Transl. Androl. Urol. 2021, 10, 1088–1101. [Google Scholar] [CrossRef]
- Rhemrev, J.P.; van Overveld, F.W.; Haenen, G.R.; Teerlink, T.; Bast, A.; Vermeiden, J.P. Quantification of the nonenzymatic fast and slow TRAP in a postaddition assay in human seminal plasma and the antioxidant contributions of various seminal compounds. J. Androl. 2000, 21, 913–920. [Google Scholar]
- Aitken, R.J.; Ryan, A.L.; Baker, M.A.; McLaughlin, E.A. Redox activity associated with the maturation and capacitation of mammalian spermatozoa. Free Radic. Biol. Med. 2004, 36, 994–1010. [Google Scholar] [CrossRef]
- Becker, B.F. Towards the physiological function of uric acid. Free Radic. Biol. Med. 1993, 14, 615–631. [Google Scholar] [CrossRef]
- Ward, W.S.; Guz, J.; Gackowski, D.; Foksinski, M.; Rozalski, R.; Zarakowska, E.; Siomek, A.; Szpila, A.; Kotzbach, M.; Kotzbach, R.; et al. Comparison of Oxidative Stress/DNA Damage in Semen and Blood of Fertile and Infertile Men. PLoS ONE 2013, 8, e68490. [Google Scholar] [CrossRef]
- Buhling, K.; Schumacher, A.; Eulenburg, C.Z.; Laakmann, E. Influence of oral vitamin and mineral supplementation on male infertility: A meta-analysis and systematic review. Reprod. BioMedicine Online 2019, 39, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchakulla, M.; Soni, Y.; Patel, P.; Parekh, N.; Ramasamy, R. A Systematic Review and Evidence-based Analysis of Ingredients in Popular Male Fertility Supplements. Urology 2020, 136, 133–141. [Google Scholar] [CrossRef]
- Fraga, C.G.; Motchnik, P.A.; Shigenaga, M.K.; Helbock, H.J.; Jacob, R.A.; Ames, B.N. Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc. Natl. Acad. Sci. USA 1991, 88, 11003–11006. [Google Scholar] [CrossRef] [PubMed]
- Vani, K.; Kurakula, M.; Syed, R.; Alharbi, K. Clinical Relevance of Vitamin C Among Lead-Exposed Infertile Men. Genet. Test. Mol. Biomark. 2012, 16, 1001–1006. [Google Scholar] [CrossRef]
- Martínez-Soto, J.C.; Domingo, J.C.; Cordobilla, B.; Nicolás, M.; Fernández, L.; Albero, P.; Gadea, J.; Landeras, J. Dietary supplementation with docosahexaenoic acid (DHA) improves seminal antioxidant status and decreases sperm DNA fragmentation. Syst. Biol. Reprod. Med. 2016, 62, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Omu, A.E.; Al-Azemi, M.K.; Kehinde, E.O.; Anim, J.T.; Oriowo, M.A.; Mathew, T.C. Indications of the Mechanisms Involved in Improved Sperm Parameters by Zinc Therapy. Med. Princ. Pract. 2008, 17, 108–116. [Google Scholar] [CrossRef]
- Mathieu d’Argent, E.; Ravel, C.; Rousseau, A.; Morcel, K.; Massin, N.; Sussfeld, J.; Simon, T.; Antoine, J.-M.; Mandelbaume, J.; Daraï, E.; et al. High-Dose Supplementation of Folic Acid in Infertile Men Improves IVF-ICSI Outcomes: A Randomized Controlled Trial (FOLFIV Trial). J. Clin. Med. 2021, 10, 1876. [Google Scholar] [CrossRef]
- Boonyarangkul, A.; Vinayanuvattikhun, N.; Chiamchanya, C.; Visutakul, P. Comparative Study of the Effects of Tamoxifen Citrate and Folate on Semen Quality of the Infertile Male with Semen Abnormality. J. Med. Assoc. Thai 2015, 98, 1057–1063. [Google Scholar]
- Jannatifar, R.; Parivar, K.; Roodbari, N.H.; Nasr-Esfahani, M.H. Effects of N-acetyl-cysteine supplementation on sperm quality, chromatin integrity and level of oxidative stress in infertile men. Reprod. Biol. Endocrinol. 2019, 17, 24. [Google Scholar] [CrossRef]
- Lipovac, M.; Bodner, F.; Imhof, M.; Chedraui, P. Comparison of the effect of a combination of eight micronutrients versus a standard mono preparation on sperm parameters. Reprod. Biol. Endocrinol. 2016, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- Gual-Frau, J.; Abad, C.; Amengual, M.J.; Hannaoui, N.; Checa, M.A.; Ribas-Maynou, J.; Lozano, I.; Nikolaou, A.; Benet, J.; García-Peiró, A.; et al. Oral antioxidant treatment partly improves integrity of human sperm DNA in infertile grade I varicocele patients. Hum. Fertil. 2015, 18, 225–229. [Google Scholar] [CrossRef]
- Iovine, C.; Mottola, F.; Santonastaso, M.; Finelli, R.; Agarwal, A.; Rocco, L. In vitro ameliorative effects of ellagic acid on vitality, motility and DNA quality in human spermatozoa. Mol. Reprod. Dev. 2021, 88, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Finelli, R.; Selvam, M.K.P.; Leisegang, K.; Majzoub, A.; Tadros, N.; Ko, E.; Parekh, N.; Henkel, R.; Durairajanayagam, D.; et al. A Global Survey of Reproductive Specialists to Determine the Clinical Utility of Oxidative Stress Testing and Antioxidant Use in Male Infertility. World J. Men’s Health 2021, 39, 470–488. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Majzoub, A. Best laboratory practices and therapeutic interventions to reduce sperm DNA damage. Andrologia 2020, 53, e13736. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Rs | |
|---|---|---|
| TUNEL | Months after COVID-19 | |
| Months after COVID−19 | −0.500 * | 1.000 |
| Age (years) | −0.033 | −0.064 |
| BMI, kg/m2 | −0.037 | 0.089 |
| PR | −0.281 | −0.269 |
| NP | −0.288 | 0.338 |
| IM | 0.398 * | 0.184 |
| Round cells | 0.343 * | −0.239 |
| Normal morphology | −0.559 *** | 0.447 |
| Mild degree of pathology | −0.443 ** | 0.523 * |
| Head defects | 0.481 ** | −0.534 * |
| Midpiece defects | −0.244 | 0.199 |
| Tail defects | −0.136 | 0.674 ** |
| TACC | 0.232 | 0.231 |
| TACSP | −0.095 | 0.751 *** |
| NTC | 0.340 * | −0.012 |
| NTSP | −0.114 | 0.401 |
| CATC | 0.385 * | −0.288 |
| CATSP | 0.023 | −0.004 |
| SODC | 0.070 | −0.402 |
| SODSP | −0.442 ** | −0.354 |
| 8−OHdGC | 0.179 | −0.391 |
| 8OHdGSP | −0.153 | 0.031 |
| Uric acidSP | −0.414 ** | 0.216 |
| ZnSP | −0.279 | 0.366 |
| Parameters | COVID−, TUNEL<15 | COVID+, TUNEL<15 | COVID−, TUNEL>15 | COVID+, TUNEL>15 |
|---|---|---|---|---|
| Progressive motility (PR, %) | 60.00 [49.00–70.00] | 62.00 [56.00–67.00] | 53.00 [41.00–63.00] | 62.00 [49.00–68.00] |
| Nonprogressive motility (NP, %) | 9.00 [8.00–11.00] | 10.00 [7.00–12.00] | 7.00 [6.00–11.00] | 7.50 [6.50–9.50] |
| Immotility (IM, %) | 27.00 [21.00–35.00] | 26.00 [21.00–29.00] | 40.00 [26.00–46.00] | 28.50 [24.00–44.00] |
| Round cells (106 cells/mL) | 0.20 [0.00–1.00] | 0.20 [0.10–1.40] | 0.20 [0.20–0.40] | 1.55 [0.60–3.00] * |
| Normal morphology (%) | 4.00 [3.00–4.00] | 3.00 [3.00–5.00] | 3.00 [2.00–3.00] | 2 [1.00–3.00] |
| Head defects (%) | 89.00 [87.00–93.00] | 89.00 [87.00–93.00] | 93.00 [91.00–95.00] | 93.00 [92.50–95.50] |
| Midpiece defects (%) | 1.00 [0.00–2.50] | 1.00 [1.00–1.00] | 1.00 [0.00–1.00] | 1.00 [0.00–1.50] |
| Tail defects (%) | 0.50 [0.00–1.00] | 1.00 [0.50–1.00] | 0.00 [0.00–1.00] | 0.0 [0.00–1.50] |
| Parameters | COVID−, TUNEL<15 | COVID+, TUNEL<15 | COVID−, TUNEL>15 | COVID+, TUNEL>15 |
|---|---|---|---|---|
| n | 13 | 9 | 9 | 8 |
| Age (years) | 36.00 [33.00–38.00] | 35.00 [34.00–38.00] | 34.00 [32.00–37.00] | 34.50 [34.00–36.50] |
| BMI | 25 [23–25] | 23 [23,24] | 23 [22–24.5] | 24 [23–25] |
| Volume (ml) | 3.50 [3.00–4.00] | 4.00 [3.00–6.50] | 3.00 [2.50–5.00] | 3.50 [2.75–3.75] |
| Sperm concentration (mln/mL) | 84.00 [67.00–176.00] | 98.00 [93.00–146.00] | 70.00 [49.00–135.00] | 72.00 [59.00–111.50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shcherbitskaia, A.D.; Komarova, E.M.; Milyutina, Y.P.; Ishchuk, M.A.; Sagurova, Y.M.; Safaryan, G.K.; Lesik, E.A.; Gzgzyan, A.M.; Bespalova, O.N.; Kogan, I.Y. Oxidative Stress Markers and Sperm DNA Fragmentation in Men Recovered from COVID-19. Int. J. Mol. Sci. 2022, 23, 10060. https://doi.org/10.3390/ijms231710060
Shcherbitskaia AD, Komarova EM, Milyutina YP, Ishchuk MA, Sagurova YM, Safaryan GK, Lesik EA, Gzgzyan AM, Bespalova ON, Kogan IY. Oxidative Stress Markers and Sperm DNA Fragmentation in Men Recovered from COVID-19. International Journal of Molecular Sciences. 2022; 23(17):10060. https://doi.org/10.3390/ijms231710060
Chicago/Turabian StyleShcherbitskaia, Anastasiia D., Evgeniia M. Komarova, Yulia P. Milyutina, Mariia A. Ishchuk, Yanina M. Sagurova, Galina K. Safaryan, Elena A. Lesik, Alexander M. Gzgzyan, Olesya N. Bespalova, and Igor Y. Kogan. 2022. "Oxidative Stress Markers and Sperm DNA Fragmentation in Men Recovered from COVID-19" International Journal of Molecular Sciences 23, no. 17: 10060. https://doi.org/10.3390/ijms231710060