A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT2A Receptor Density in the Pig Brain

 , , and
, , and
Abstract
:1. Introduction
2. Results
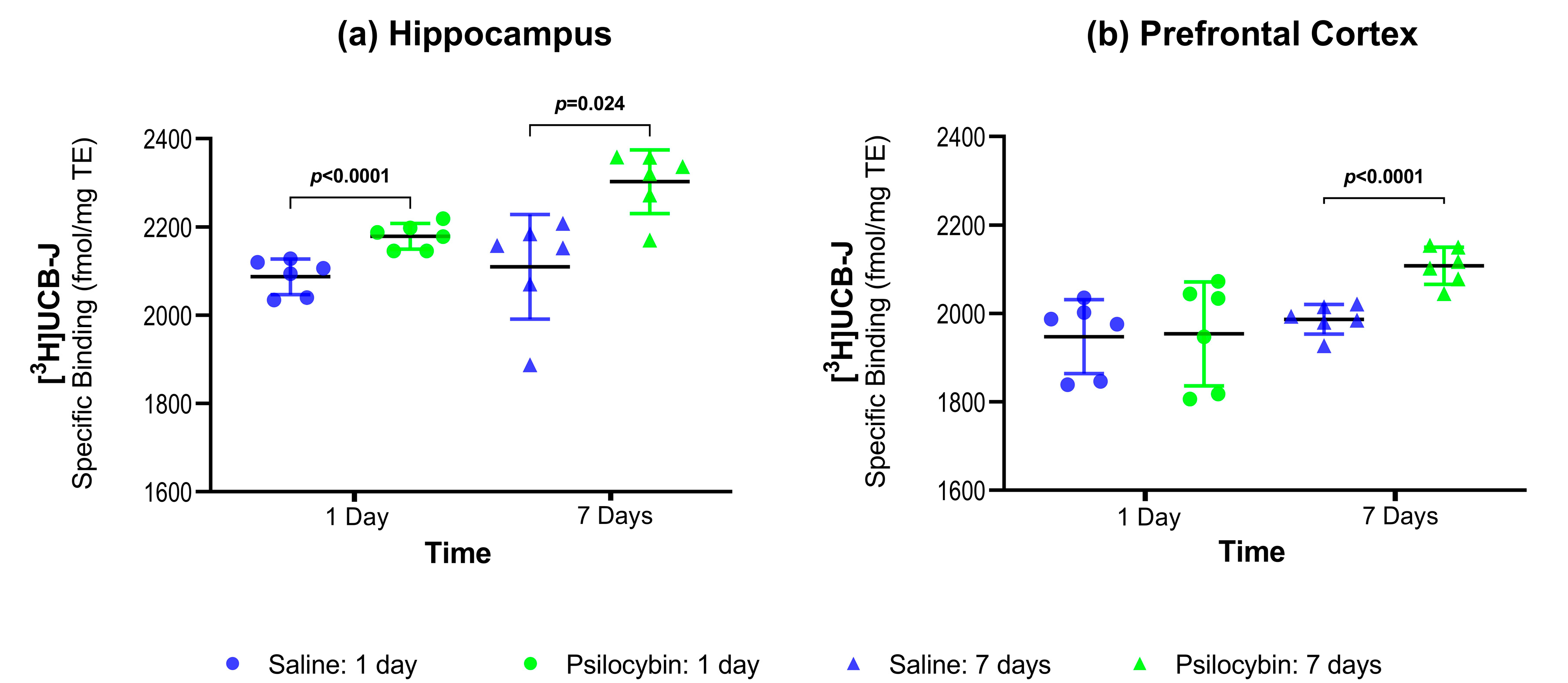
2.1. SV2A Autoradiography
2.2. 5-HT2AR Antagonist Autoradiography:
2.3. 5-HT2AR Agonist Autoradiography
2.4. Antagonist vs. Agonist Radioligand for 5-HT2AR Density
2.5. Plasma Psilocin
3. Discussion
4. Materials and Methods
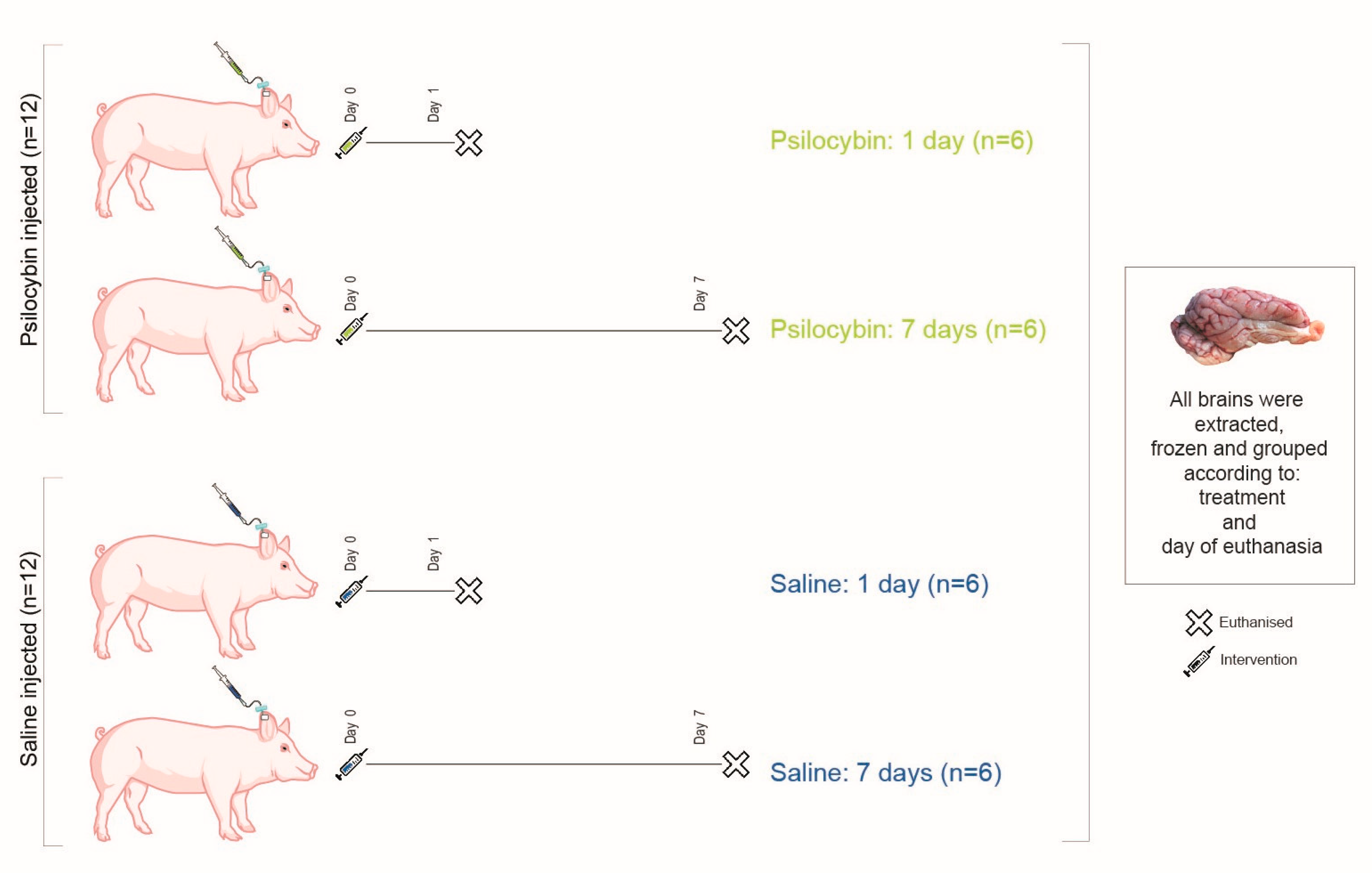
4.1. Animals and Drug Dosage
4.2. Ethical Statement
4.3. Study Design
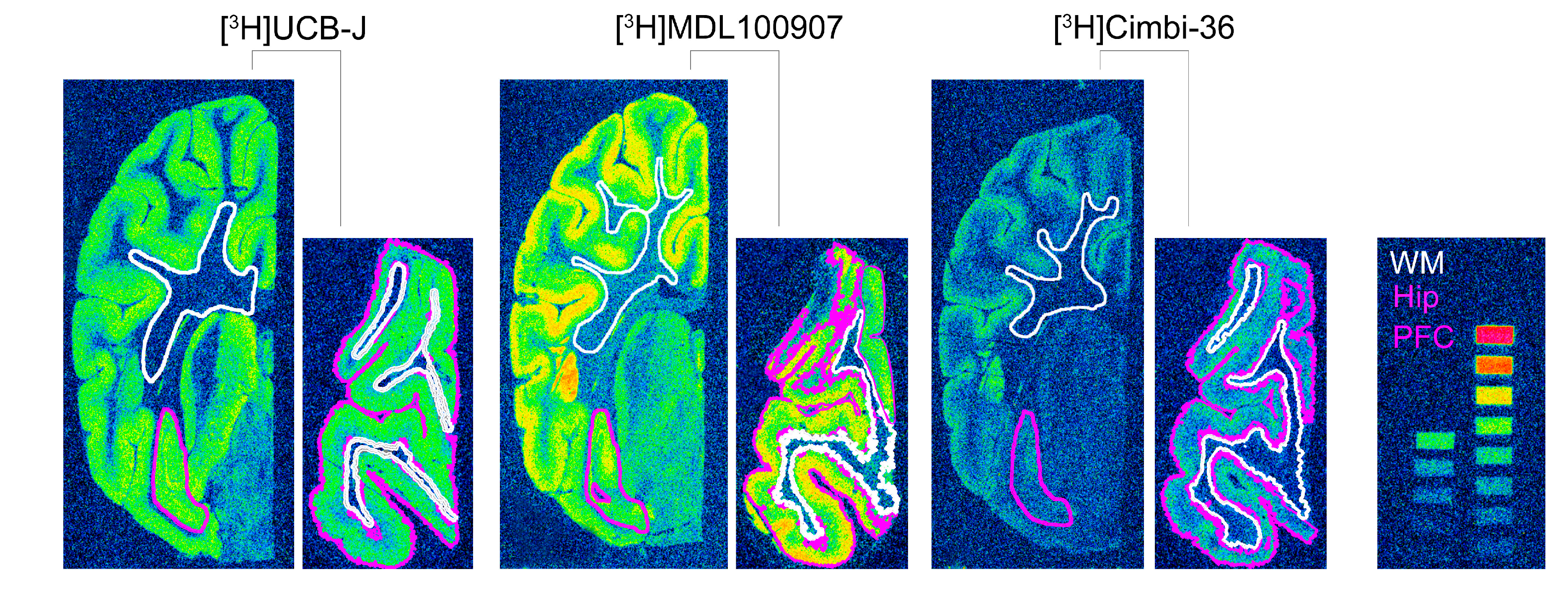
4.4. Autoradiography
4.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HT | 5-Hydroxytryptamine (serotonin) |
| 5-HT1AR | 5-Hydroxytryptamine (serotonin) 1A Receptor |
| 5-HT2AR | 5-Hydroxytryptamine (serotonin) 2A Receptor |
| 5-HT2CR | 5-Hydroxytryptamine (serotonin) 2C Receptor |
| ARRIVE | Animal Research: Reporting of In Vivo Experiments |
| BDNF | Brain-Derived Neurotropic Factor |
| Bmax | Total density (concentration) of the receptor |
| BPND | Binding Potential (Non-displaceable) |
| BSA | Bovine Serum Albumin |
| DMT | N,N-dimethyltryptamine |
| DOI | 2,5-dimethoxy-4-iodoamphetamine |
| GPCR | G-protein coupled receptor |
| KD | Equilibrium dissociation Constant |
| LSD | Lysergic acid diethylamide |
| MAP2 | Microtubule-associated protein 2 |
| mTOR | Mammalian Target of Rapamycin |
| NSB | Non-specific Binding |
| PET | Positron Emission Tomography |
| pERK | Endoplasmic Reticulum Protein Kinase |
| PFC | Prefrontal Cortex |
| RCP | Radiochemical purity |
| R-TLC | Radio-Thin Layer Chromatography |
| SV2A | Synaptic Vesicular Protein 2A |
| TB | Total Binding |
| TE | Tissue Equivalent |
| TrkB | Tropomyosin Receptor Kinase B |
| VGLUT1 | Vesicular Glutamate Transporter 1 |
| Mean |
Appendix A
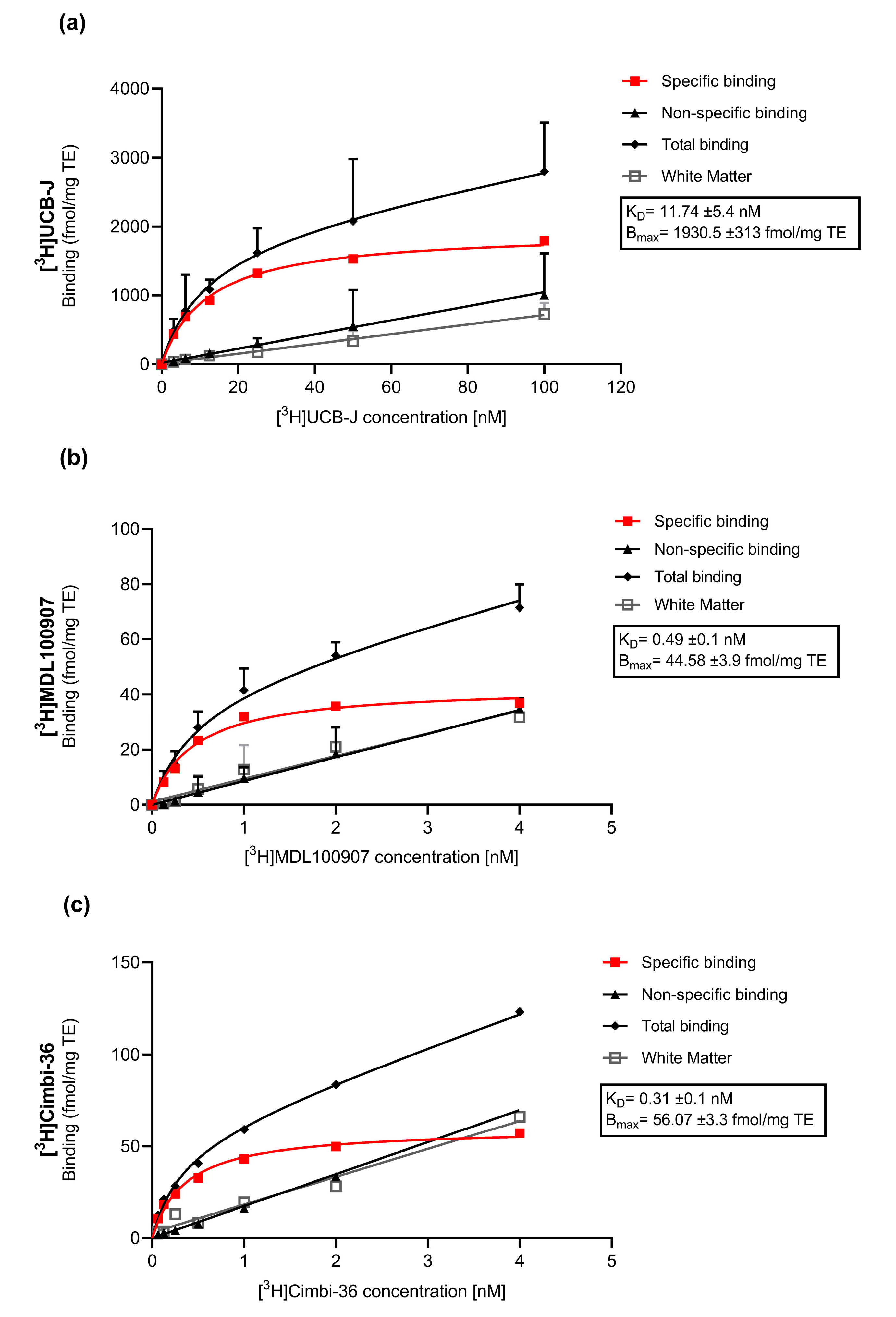
Appendix A.1. Saturation Assays
Appendix A.1.1. Methods
Appendix A.1.2. Results

References
- Nichols, D.E. Hallucinogens. Pharmacol. Ther. 2004, 101, 131–181. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, F.X.; Kometer, M. The neurobiology of psychedelic drugs: Implications for the treatment of mood disorders. Nat. Rev. Neurosci. 2010, 11, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, F.X.; Preller, K.H. Psychedelic drugs: Neurobiology and potential for treatment of psychiatric disorders. Nat. Rev. Neurosci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rickli, A.; Moning, O.D.; Hoener, M.C.; Liechti, M.E. Receptor interaction profiles of novel psychoactive tryptamines compared with classic hallucinogens. Eur. Neuropsychopharmacol. 2016, 26, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, J.B.; Kurrasch-Orbaugh, D.; Marona-Lewicka, D.; Gumbay, M.G.; Watts, V.J.; Barker, E.L.; Nichols, D.E. Effect of ring fluorination on the pharmacology of hallucinogenic tryptamines. J. Med. Chem. 2000, 43, 4701–4710. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Bolstridge, M.; Rucker, J.; Day, C.M.J.J.; Erritzoe, D.; Kaelen, M.; Bloomfield, M.; Rickard, J.A.; Forbes, B.; Feilding, A.; et al. Psilocybin with psychological support for treatment-resistant depression: An open-label feasibility study. Lancet Psychiatry 2016, 3, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.R.; Johnson, M.W.; Carducci, M.A.; Umbricht, A.; Richards, W.A.; Richards, B.D.; Cosimano, M.P.; Klinedinst, M.A. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: A randomized double-blind trial. J. Psychopharmacol. 2016, 30, 1181–1197. [Google Scholar] [CrossRef]
- Ross, S.; Bossis, A.; Guss, J.; Agin-Liebes, G.; Malone, T.; Cohen, B.; Mennenga, S.E.; Belser, A.; Kalliontzi, K.; Babb, J.; et al. Rapid and sustained symptom reduction following psilocybin treatment for anxiety and depression in patients with life-threatening cancer: A randomized controlled trial. J. Psychopharmacol. 2016, 30, 1165–1180. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.K.; Barrett, F.S.; May, D.G.; Cosimano, M.P.; Sepeda, N.D.; Johnson, M.W.; Finan, P.H.; Griffiths, R.R. Effects of Psilocybin-Assisted Therapy on Major Depressive Disorder: A Randomized Clinical Trial. JAMA Psychiatry 2020. [Google Scholar] [CrossRef]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Eden, V.; Wilson, P.C.; Burbach, K.F.; Zarandi, S.S.; Paddy, M.R.; Duim, W.C.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef]
- Morales-García, J.A.; De La Fuente Revenga, M.; Alonso-Gil, S.; Rodríguez-Franco, M.I.; Feilding, A.; Perez-Castillo, A.; Riba, J. The alkaloids of Banisteriopsis caapi, the plant source of the Amazonian hallucinogen Ayahuasca, stimulate adult neurogenesis in vitro. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Fries, G.R.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Chronic administration of harmine elicits antidepressant-like effects and increases BDNF levels in rat hippocampus. J. Neural Transm. 2010, 117, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Morales-Garcia, J.A.; Calleja-Conde, J.; Lopez-Moreno, J.A.; Alonso-Gil, S.; Sanz-SanCristobal, M.; Riba, J.; Perez-Castillo, A. N,N-dimethyltryptamine compound found in the hallucinogenic tea ayahuasca, regulates adult neurogenesis in vitro and in vivo. Transl. Psychiatry 2020, 10, 331. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.A.; Srivastava, D.P.; Allen, J.A.; Strachan, R.T.; Roth, B.L.; Penzes, P. Rapid modulation of spine morphology by the 5-HT2A serotonin receptor through kalirin-7 signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 19575–19580. [Google Scholar] [CrossRef] [Green Version]
- Duric, V.; Banasr, M.; Stockmeier, C.A.; Simen, A.A.; Newton, S.S.; Overholser, J.C.; Jurjus, G.J.; Dieter, L.; Duman, R.S. Altered expression of synapse and glutamate related genes in post-mortem hippocampus of depressed subjects. Int. J. Neuropsychopharmacol. 2013, 16, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Voleti, B.; Hajszan, T.; Rajkowska, G.; Stockmeier, C.A.; Licznerski, P.; Lepack, A.; Majik, M.S.; Jeong, L.S.; Banasr, M.; et al. Decreased expression of synapse-related genes and loss of synapses in major depressive disorder. Nat. Med. 2012, 18, 1413–1417. [Google Scholar] [CrossRef]
- Holmes, S.E.; Scheinost, D.; Finnema, S.J.; Naganawa, M.; Davis, M.T.; DellaGioia, N.; Nabulsi, N.; Matuskey, D.; Angarita, G.A.; Pietrzak, R.H.; et al. Lower synaptic density is associated with depression severity and network alterations. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Bartholome, O.; Van den Ackerveken, P.; Sánchez Gil, J.; de la Brassinne Bonardeaux, O.; Leprince, P.; Franzen, R.; Rogister, B.; Surguchov, A.; Conti, F.; Haucke, V.; et al. Puzzling Out Synaptic Vesicle 2 Family Members Functions. Front. Mol. Neurosci 2017, 10, 148. [Google Scholar] [CrossRef]
- Heurling, K.; Ashton, N.J.; Leuzy, A.; Zimmer, E.R.; Blennow, K.; Zetterberg, H.; Eriksson, J.; Lubberink, M.; Schöll, M. Synaptic vesicle protein 2A as a potential biomarker in synaptopathies. Mol. Cell. Neurosci. 2019, 97, 34–42. [Google Scholar] [CrossRef]
- Nowack, A.; Yao, J.; Custer, K.L.; Bajjalieh, S.M. SV2 regulates neurotransmitter release via multiple mechanisms. Am. J. Physiol. Cell Physiol. 2010, 299, 960–967. [Google Scholar] [CrossRef] [Green Version]
- Finnema, S.J.; Nabulsi, N.B.; Eid, T.; Detyniecki, K.; Lin, S.F.; Chen, M.K.; Dhaher, R.; Matuskey, D.; Baum, E.; Holden, D.; et al. Imaging synaptic density in the living human brain. Sci. Transl. Med. 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varnäs, K.; Stepanov, V.; Halldin, C. Autoradiographic mapping of synaptic vesicle glycoprotein 2A in non-human primate and human brain. Synapse 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Giménez, J.F.; Villazón, M.; Brea, J.; Loza, M.I.; Palacios, J.M.; Mengod, G.; Vilaró, M.T. Multiple conformations of native and recombinant human 5-hydroxytryptamine2A receptors are labeled by agonists and discriminated by antagonists. Mol. Pharmacol. 2001, 60, 690–699. [Google Scholar] [PubMed]
- Song, J.; Hanniford, D.; Doucette, C.; Graham, E.; Poole, M.F.; Ting, A.; Sherf, B.; Harrington, J.; Brunden, K.; Stricker-Krongrad, A. Development of homogenous high-affinity agonist binding assays for 5-HT2 receptor subtypes. Assay Drug Dev. Technol. 2005, 3, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Shalgunov, V.; van Waarde, A.; Booij, J.; Michel, M.C.; Dierckx, R.A.J.O.; Elsinga, P.H. Hunting for the high-affinity state of G-protein-coupled receptors with agonist tracers: Theoretical and practical considerations for positron emission tomography imaging. Med. Res. Rev. 2019, 39, 1014–1052. [Google Scholar] [CrossRef]
- Kent, R.S.; De Lean, A.; Lefkowitz, R.J. A quantitative analysis of beta-adrenergic receptor interactions: Resolution of high and low affinity states of the receptor by computer modeling of ligand binding data. Mol. Pharmacol. 1980, 17, 14–23. [Google Scholar]
- López-Giménez, J.F.; Vilaró, M.T.; Palacios, J.M.; Mengod, G. Multiple conformations of 5-HT2A and 5-HT2C receptors in rat brain: An autoradiographic study with [125I](±)DOI. Exp. Brain Res. 2013, 230, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Berry, S.A.; Shah, M.C.; Khan, N.; Roth, B.L. Rapid agonist-induced internalization of the 5-hydroxytryptamine(2A) receptor occurs via the endosome pathway in vitro. Mol. Pharmacol. 1996, 50, 306–313. [Google Scholar]
- Gray, J.A.; Roth, B.L. Paradoxical trafficking and regulation of 5-HT2A receptors by agonists and antagonists. Brain Res. Bull. 2001, 56, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Donovan, L.L.; Johansen, J.V.; Ros, N.F.; Jaberi, E.; Johansen, S.S.; Ozenne, B.; Issazadeh-Navikas, S.; Hansen, H.D.; Knudsen, G.M. Effects of a single dose of psilocybin on behaviour, brain 5-HT2A receptor occupancy and gene expression in the pig. Eur. Neuropsychopharmacol. 2021, 42, 1–11. [Google Scholar] [CrossRef]
- Zhang, M.; Radford, K.D.; Driscoll, M.; Purnomo, S.; Kim, J.; Choi, K.H. Effects of subanesthetic intravenous ketamine infusion on neuroplasticity-related proteins in the prefrontal cortex, amygdala, and hippocampus of Sprague-Dawley rats. IBRO Rep. 2019, 6, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hutten, N.R.P.W.; Mason, N.L.; Dolder, P.C.; Theunissen, E.L.; Holze, F.; Liechti, M.E.; Varghese, N.; Eckert, A.; Feilding, A.; Ramaekers, J.G.; et al. Low Doses of LSD Acutely Increase BDNF Blood Plasma Levels in Healthy Volunteers. ACS Pharmacol. Trans. Sci. 2020, acsptsci.0c00099. [Google Scholar] [CrossRef]
- Beliveau, V.; Ganz, M.; Feng, L.; Ozenne, B.; Højgaard, L.; Fisher, P.M.; Svarer, C.; Greve, D.N.; Knudsen, G.M. A high-resolution in vivo atlas of the human brain’s serotonin system. J. Neurosci. 2017, 37, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Anji, A.; Kumari, M.; Sullivan Hanley, N.R.; Bryan, G.L.; Hensler, J.G. Regulation of 5-HT2A receptor mRNA levels and binding sites in rat frontal cortex by the agonist DOI and the antagonist mianserin. Neuropharmacology 2000, 39, 1996–2005. [Google Scholar] [CrossRef]
- Pompeiano, M.; Palacios, J.M.; Mengod, G. Distribution of the serotonin 5-HT2 receptor family mRNAs: Comparison between 5-HT2A and 5-HT2C receptors. Mol. Brain Res. 1994, 23, 163–178. [Google Scholar] [CrossRef]
- Marazziti, D.; Rossi, A.; Giannaccini, G.; Zavaglia, K.M.; Dell’Osso, L.; Lucacchini, A.; Cassano, G.B. Distribution and characterization of [ 3 H]mesulergine binding in human brain postmortem. Eur. Neuropsychopharmacol. 1999, 10, 21–26. [Google Scholar] [CrossRef]
- Ettrup, A.; Holm, S.; Hansen, M.; Wasim, M.; Santini, M.A.; Palner, M.; Madsen, J.; Svarer, C.; Kristensen, J.L.; Knudsen, G.M. Preclinical safety assessment of the 5-HT2A receptor agonist PET radioligand [11C]cimbi-36. Mol. Imaging Biol. 2013, 15, 376–383. [Google Scholar] [CrossRef]
- Finnema, S.J.; Stepanov, V.; Ettrup, A.; Nakao, R.; Amini, N.; Svedberg, M.; Lehmann, C.; Hansen, M.; Knudsen, G.M.; Halldin, C. Characterization of [11C]Cimbi-36 as an agonist PET radioligand for the 5-HT2A and 5-HT2C receptors in the nonhuman primate brain. Neuroimage 2014, 84, 342–353. [Google Scholar] [CrossRef]
- Ettrup, A.; Svarer, C.; McMahon, B.; da Cunha-Bang, S.; Lehel, S.; Møller, K.; Dyssegaard, A.; Ganz, M.; Beliveau, V.; Jørgensen, L.M.; et al. Serotonin 2A receptor agonist binding in the human brain with [11C]Cimbi-36: Test–retest reproducibility and head-to-head comparison with the antagonist [18 F]altanserin. Neuroimage 2016, 130, 167–174. [Google Scholar] [CrossRef]
- García-Bea, A.; Miranda-Azpiazu, P.; Muguruza, C.; Marmolejo-Martinez-Artesero, S.; Diez-Alarcia, R.; Gabilondo, A.M.; Callado, L.F.; Morentin, B.; González-Maeso, J.; Meana, J.J. Serotonin 5-HT2A receptor expression and functionality in postmortem frontal cortex of subjects with schizophrenia: Selective biased agonism via Gαi1-proteins. Eur. Neuropsychopharmacol. 2019, 29, 1453–1463. [Google Scholar] [CrossRef]
- Miranda-Azpiazu, P.; Díez-Alarcia, R.; García-Bea, A.; González-Maeso, J.; Morentín, B.; Meana, J.J. P.1.g.022 Hallucinogenic and non-hallucinogenic 5-HT2A receptor agonists induce distinct patterns of G protein coupling in postmortem human brain. Eur. Neuropsychopharmacol. 2013, 23, S201. [Google Scholar] [CrossRef]
- Buckholtz, N.S.; Zhou, D.; Freedman, D.X. Serotonin2 agonist administration down-regulates rat brain serotonin2 receptors. Life Sci. 1988, 42, 2439–2445. [Google Scholar] [CrossRef]
- Madsen, M.K.; Fisher, P.M.; Burmester, D.; Dyssegaard, A.; Stenbæk, D.S.; Kristiansen, S.; Johansen, S.S.; Lehel, S.; Linnet, K.; Svarer, C.; et al. Psychedelic effects of psilocybin correlate with serotonin 2A receptor occupancy and plasma psilocin levels. Neuropsychopharmacology 2019, 44, 1328–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menneson, S.; Ménicot, S.; Ferret-Bernard, S.; Guérin, S.; Romé, V.; Le Normand, L.; Randuineau, G.; Gambarota, G.; Noirot, V.; Etienne, P.; et al. Validation of a psychosocial chronic stress model in the pig using a multidisciplinary approach at the gut-brain and behavior levels. Front. Behav. Neurosci. 2019, 13, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis HHS Public Access. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hippocampus | Prefrontal Cortex | |||
|---|---|---|---|---|
| 1 day | 7 Days | 1 Day | 7 Days | |
| [3H]UCB-J (psilocybin vs. saline) | +4.42% (p < 0.0001) | +9.24% (p = 0.024) | +0.25% (p = 1) | +6.10% (p < 0.0001) |
| [3H]MDL100907 (psilocybin vs. saline) | −29.60% (p < 0.0001) | −3.58% (p = 1) | −15.21% (p = 0.162) | +1.32% (p = 1) |
| [3H]Cimbi-36 (psilocybin vs. saline) | −43.39% (p = 0.013) | +3.31% (p = 1) | −50.19% (p < 0.0001) | +2.23% (p = 1) |
| [3H]MDL100907 vs. [3H]Cimbi-36 | NA | NA | −41.26% (p = 0.033) | +0.90% (p = 0.921) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raval, N.R.; Johansen, A.; Donovan, L.L.; Ros, N.F.; Ozenne, B.; Hansen, H.D.; Knudsen, G.M. A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT2A Receptor Density in the Pig Brain. Int. J. Mol. Sci. 2021, 22, 835. https://doi.org/10.3390/ijms22020835
Raval NR, Johansen A, Donovan LL, Ros NF, Ozenne B, Hansen HD, Knudsen GM. A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT2A Receptor Density in the Pig Brain. International Journal of Molecular Sciences. 2021; 22(2):835. https://doi.org/10.3390/ijms22020835
Chicago/Turabian StyleRaval, Nakul Ravi, Annette Johansen, Lene Lundgaard Donovan, Nídia Fernandez Ros, Brice Ozenne, Hanne Demant Hansen, and Gitte Moos Knudsen. 2021. "A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT2A Receptor Density in the Pig Brain" International Journal of Molecular Sciences 22, no. 2: 835. https://doi.org/10.3390/ijms22020835