Cardioprotection Conferred by Sitagliptin Is Associated with Reduced Cardiac Angiotensin II/Angiotensin-(1-7) Balance in Experimental Chronic Kidney Disease

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. DPPIV Inhibition Attenuates Renal Dysfunction and the Increase in Blood Pressure in CKD Rats
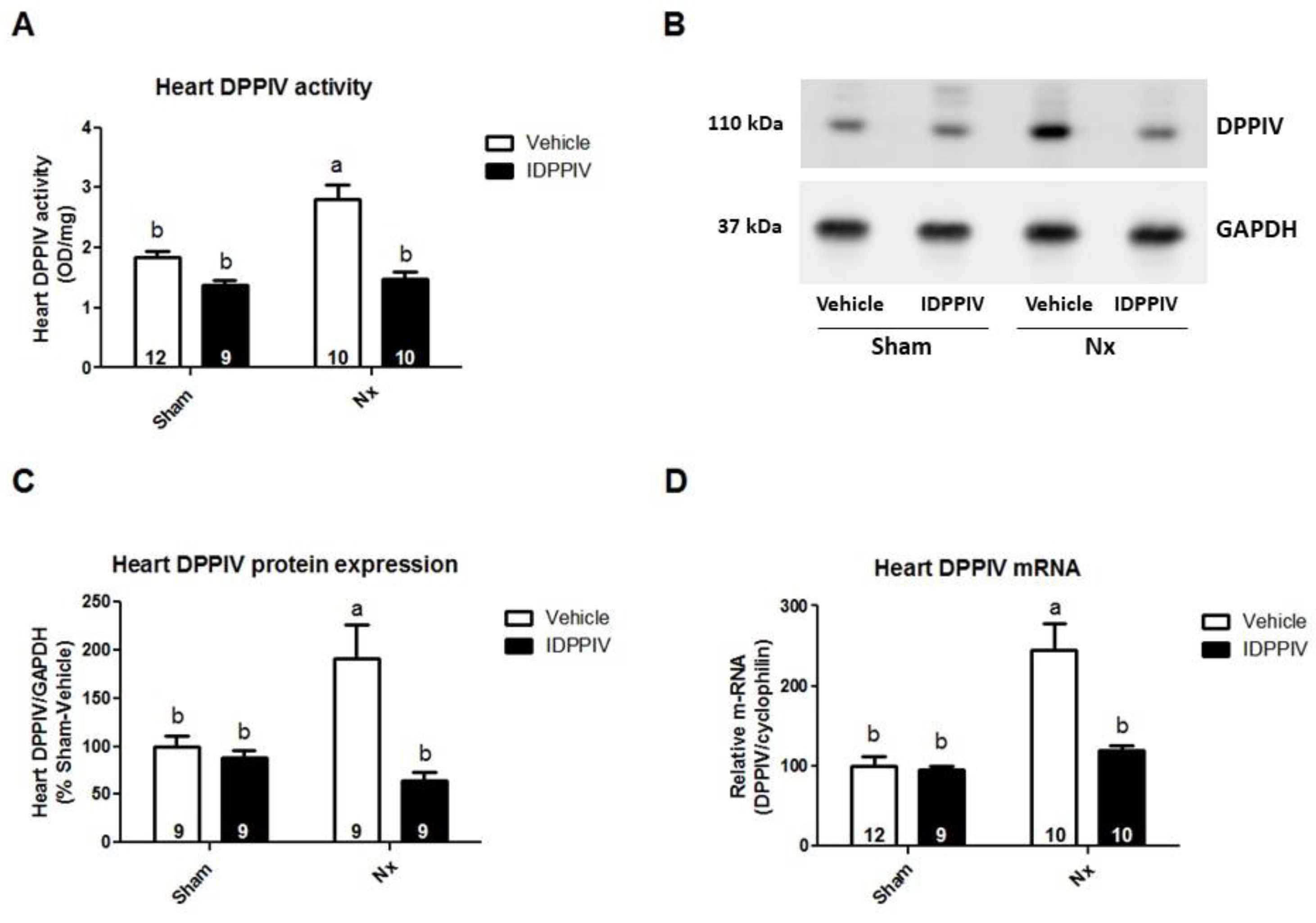
2.2. Cardiac DPPIV Activity and Expression Are Upregulated in CKD Rats
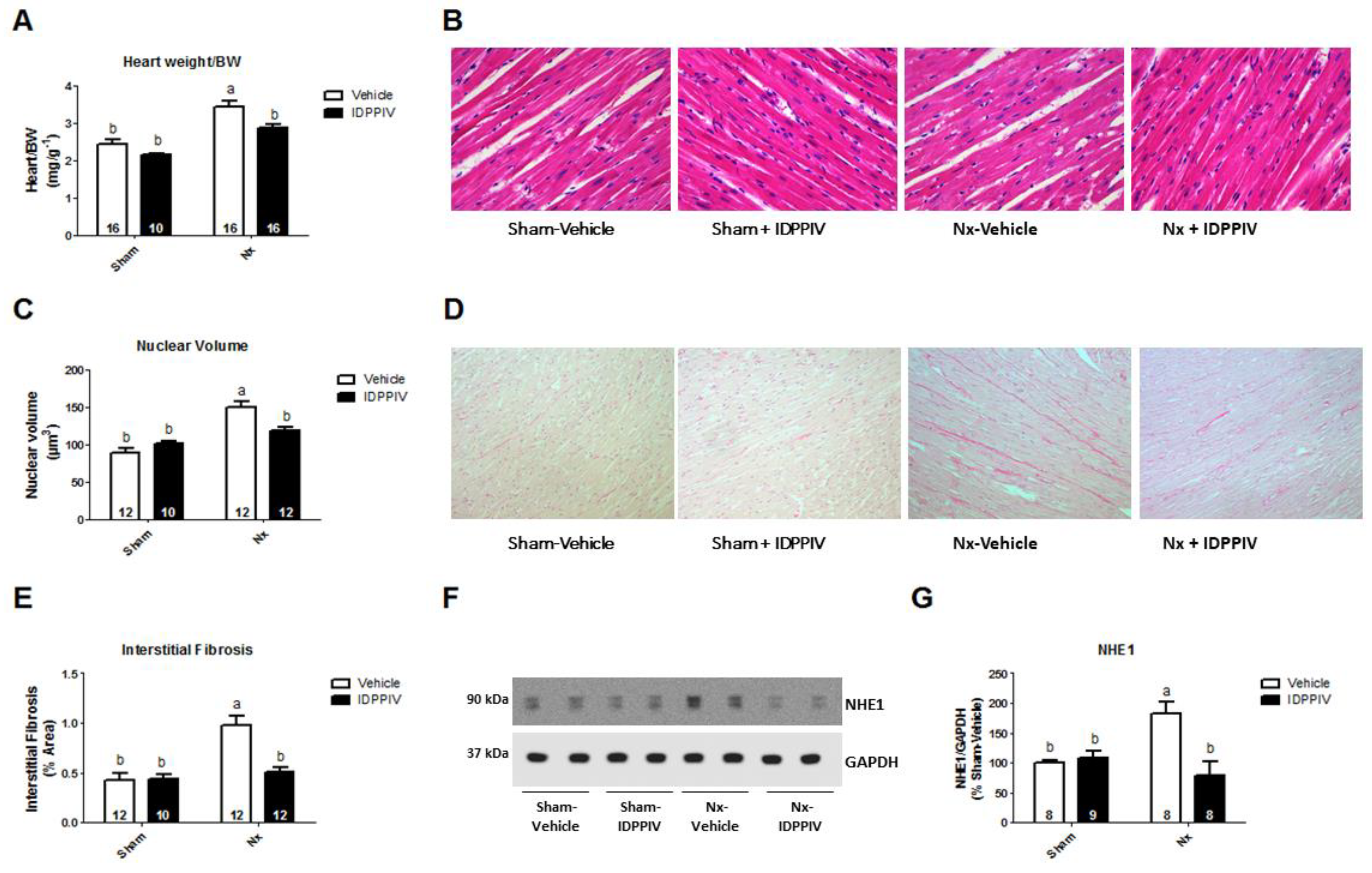
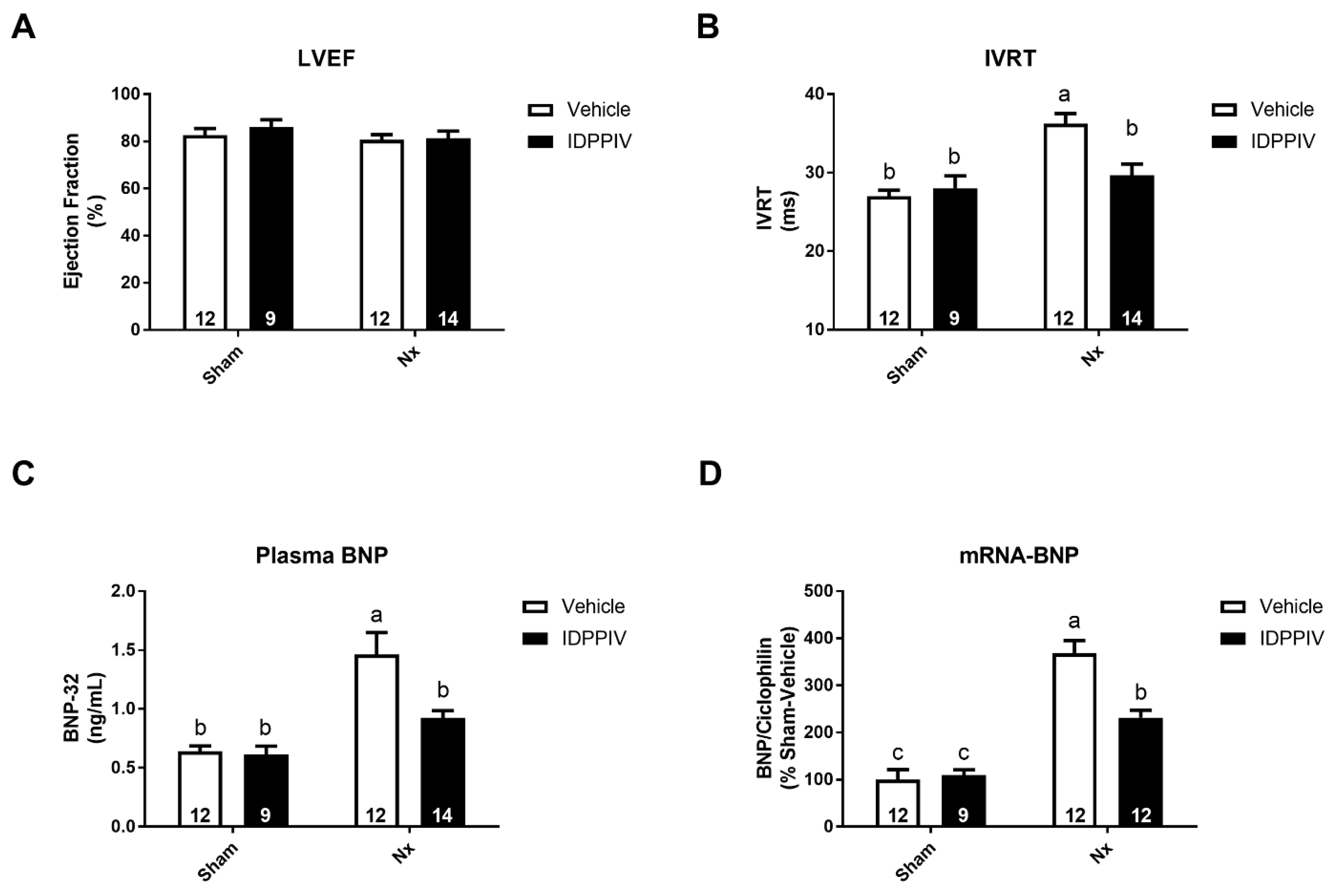
2.3. DPPIV Inhibition Mitigates Cardiac Remodeling and Diastolic Dysfunction in CKD Rats
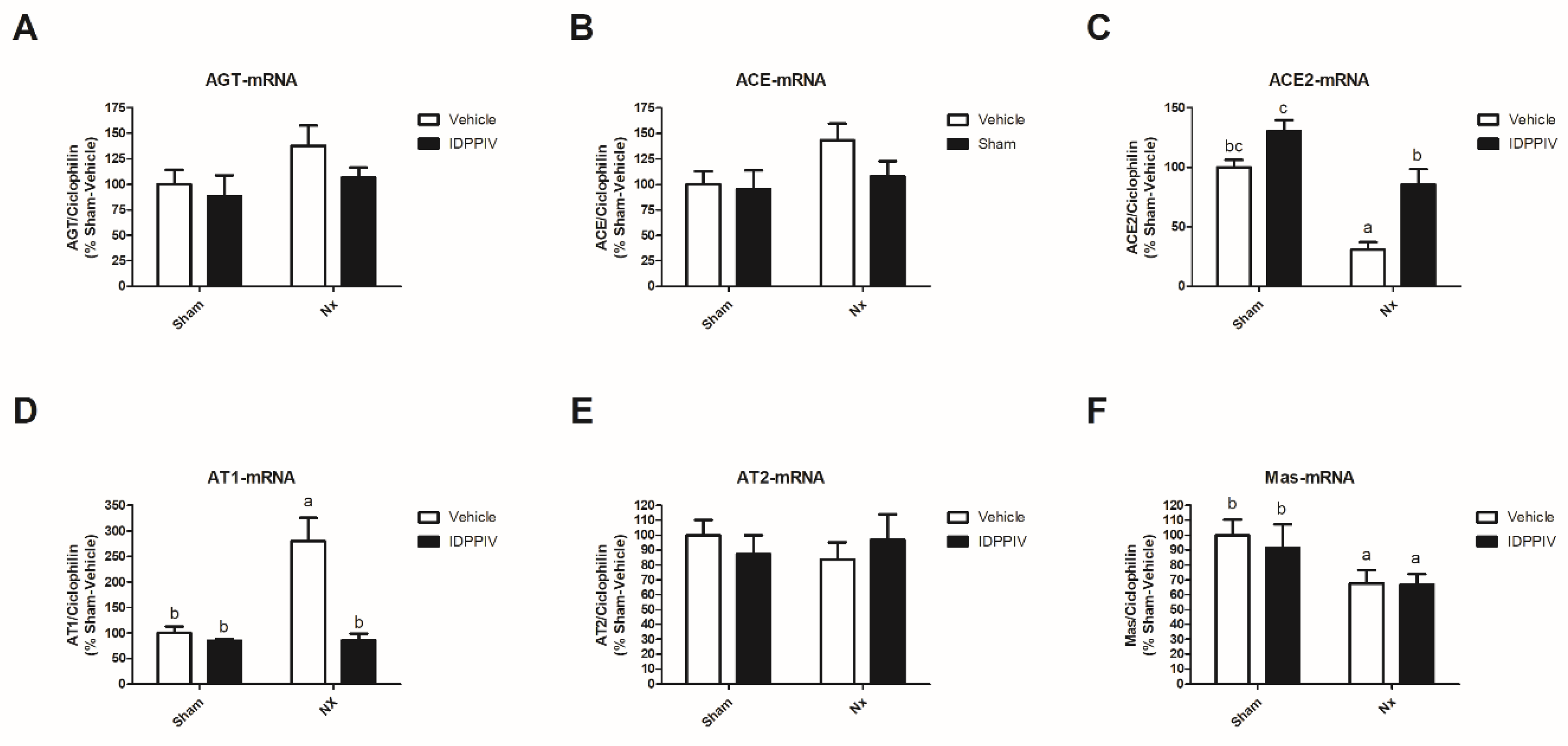
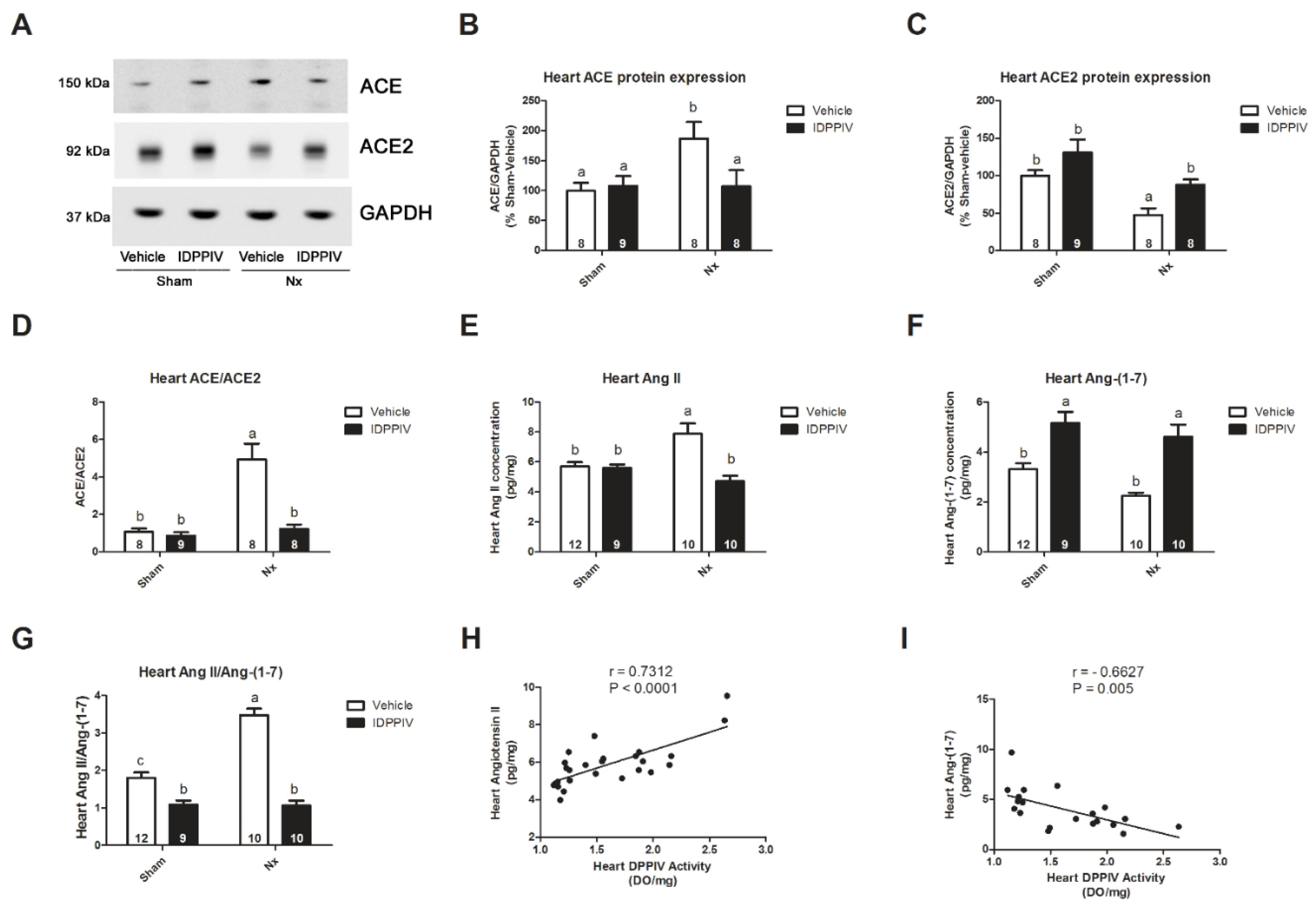
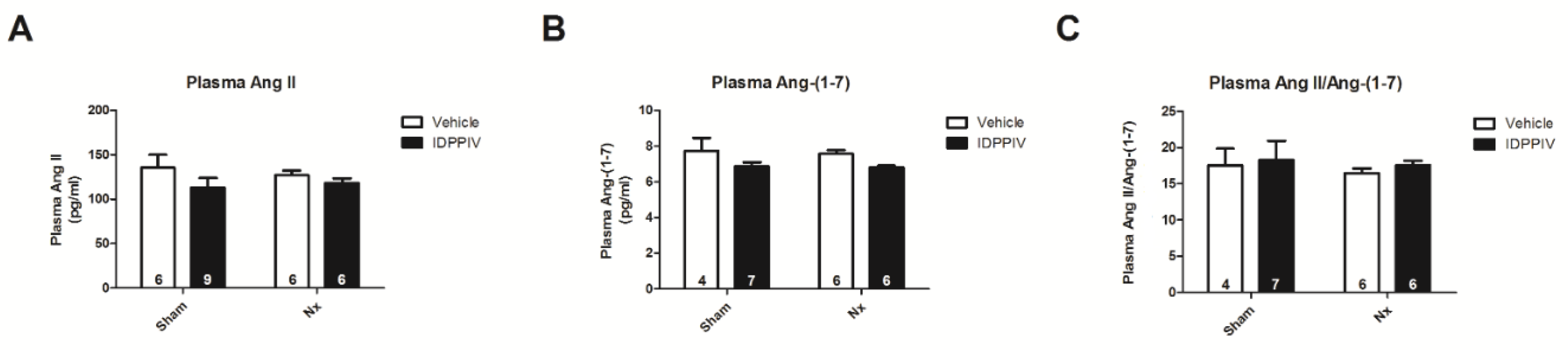
2.4. DPPIV Inhibition Influences the Cardiac Renin-Angiotensin System (RAS) in CKD Rats
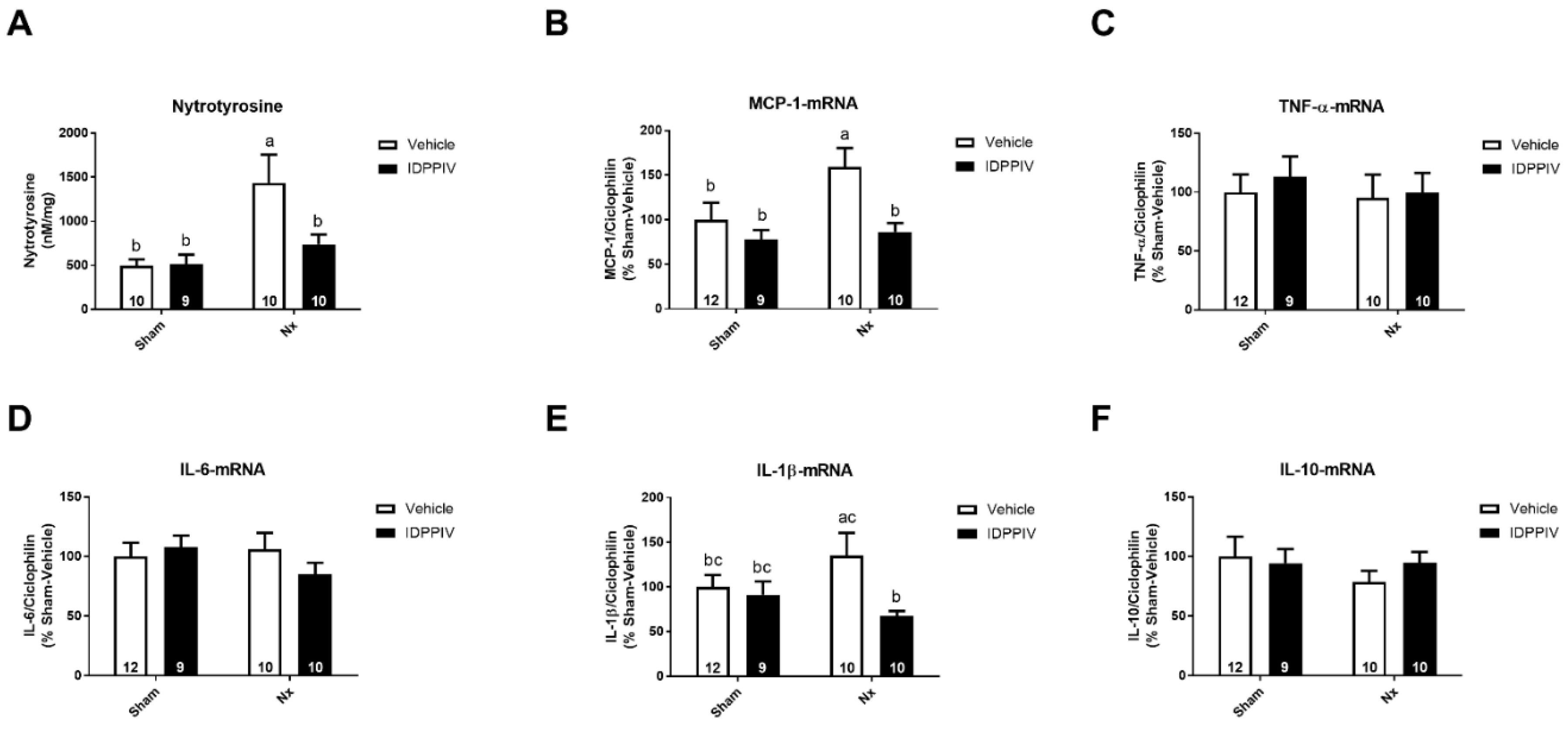
2.5. DPPIV Inhibition Exerts Antioxidant and Anti-Inflammatory Actions in the Heart of CKD Rats
3. Discussion
4. Materials and Methods
4.1. Animals and Surgical Procedures
4.2. Blood Pressure Measurement
4.3. Renal Function Assessment
4.4. Urine and Blood Analysis
4.5. Echocardiography
4.6. Biometric and Morphometric Analysis
4.7. Determination of DPPIV Activity and Abundance
4.8. SDS-PAGE and Immunoblotting
4.9. Quantitative RT-PCR
4.10. Assessment of RAS Components
4.11. Heart Nitrotyrosine Levels
4.12. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Scirica, B.M.; Bhatt, D.L.; Braunwald, E.; Steg, P.G.; Davidson, J.; Hirshberg, B.; Ohman, P.; Frederich, R.; Wiviott, S.D.; Hoffman, E.B.; et al. Saxagliptin and cardiovascular outcomes in patients with type 2 diabetes mellitus. N. Engl. J. Med. 2013, 369, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Udell, J.A.; Bhatt, D.L.; Braunwald, E.; Cavender, M.A.; Mosenzon, O.; Steg, P.G.; Davidson, J.A.; Nicolau, J.C.; Corbalan, R.; Hirshberg, B.; et al. Saxagliptin and cardiovascular outcomes in patients with type 2 diabetes and moderate or severe renal impairment: Observations from the SAVOR-TIMI 53 Trial. Diabetes Care 2015, 38, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Joo, K.W.; Kim, S.; Ahn, S.Y.; Chin, H.J.; Chae, D.W.; Lee, J.; Han, J.S.; Na, K.Y. Dipeptidyl peptidase IV inhibitor attenuates kidney injury in rat remnant kidney. BMC Nephrol. 2013, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Tsuprykov, O.; Ando, R.; Reichetzeder, C.; von Websky, K.; Antonenko, V.; Sharkovska, Y.; Chaykovska, L.; Rahnenführer, J.; Hasan, A.A.; Tammen, H.; et al. The dipeptidyl peptidase inhibitor linagliptin and the angiotensin II receptor blocker telmisartan show renal benefit by different pathways in rats with 5/6 nephrectomy. Kidney Int. 2016, 89, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Eun Lee, J.; Kim, J.E.; Lee, M.H.; Song, H.K.; Ghee, J.Y.; Kang, Y.S.; Min, H.S.; Kim, H.W.; Cha, J.J.; Han, J.Y.; et al. DA-1229, a dipeptidyl peptidase IV inhibitor, protects against renal injury by preventing podocyte damage in an animal model of progressive renal injury. Lab. Investig. 2016, 96, 547–560. [Google Scholar] [CrossRef] [Green Version]
- Chaykovska, L.; von Websky, K.; Rahnenführer, J.; Alter, M.; Heiden, S.; Fuchs, H.; Runge, F.; Klein, T.; Hocher, B. Effects of DPP-4 inhibitors on the heart in a rat model of uremic cardiomyopathy. PLoS ONE 2011, 6, e27861. [Google Scholar] [CrossRef]
- Dos Santos, L.; Salles, T.A.; Arruda-Junior, D.F.; Campos, L.C.G.; Pereira, A.C.; Barreto, A.L.T.; Antonio, E.L.; Mansur, A.J.; Tucci, P.J.F.; Krieger, J.E.; et al. Circulating dipeptidyl peptidase IV activity correlates with cardiac dysfunction in human and experimental heart failure. Circul. Heart Fail. 2013, 6, 1029–1038. [Google Scholar] [CrossRef]
- Arruda-Junior, D.F.; Martins, F.L.; Dariolli, R.; Jensen, L.; Antonio, E.L.; Dos Santos, L.; Tucci, P.J.; Girardi, A.C. Dipeptidyl Peptidase IV Inhibition Exerts Renoprotective Effects in Rats with Established Heart Failure. Front. Physiol. 2016, 7, 293. [Google Scholar] [CrossRef]
- Gomez, N.; Touihri, K.; Matheeussen, V.; Mendes Da Costa, A.; Mahmoudabady, M.; Mathieu, M.; Baerts, L.; Peace, A.; Lybaert, P.; Scharpé, S.; et al. Dipeptidyl peptidase IV inhibition improves cardiorenal function in overpacing-induced heart failure. Eur. J. Heart Fail. 2012, 14, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salles, T.A.; dos Santos, L.; Barauna, V.G.; Girardi, A.C.C. Potential role of dipeptidyl peptidase IV in the pathophysiology of heart failure. Int. J. Mol. Sci. 2015, 16, 4226–4249. [Google Scholar] [CrossRef]
- Takashima, S.; Fujita, H.; Fujishima, H.; Shimizu, T.; Sato, T.; Morii, T.; Tsukiyama, K.; Narita, T.; Takahashi, T.; Drucker, D.J.; et al. Stromal cell-derived factor-1 is upregulated by dipeptidyl peptidase-4 inhibition and has protective roles in progressive diabetic nephropathy. Kidney Int. 2016, 90, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Farah, L.X.; Valentini, V.; Pessoa, T.D.; Malnic, G.; McDonough, A.A.; Girardi, A.C. The physiological role of glucagon-like peptide-1 in the regulation of renal function. Am. J. Physiol. Renal Physiol. 2016, 310, F123–F127. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J.; Deacon, C.F. Glucagon-like peptide-1 mediates the therapeutic actions of DPP-IV inhibitors. Diabetologia 2005, 48, 612–615. [Google Scholar] [CrossRef]
- Siragy, H.M.; Carey, R.M. Role of the intrarenal renin-angiotensin-aldosterone system in chronic kidney disease. Am. J. Nephrol. 2010, 31, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Remuzzi, G.; Perico, N.; Macia, M.; Ruggenenti, P. The role of renin-angiotensin-aldosterone system in the progression of chronic kidney disease. Kidney Int. Suppl. 2005, S57–S65. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.K.; Kam, K.K.; Yan, B.P.; Lam, Y.Y. Renin-angiotensin-aldosterone system blockade for cardiovascular diseases: Current status. Br. J. Pharmacol. 2010, 160, 1273–1292. [Google Scholar] [CrossRef] [PubMed]
- Aroor, A.; Zuberek, M.; Duta, C.; Meuth, A.; Sowers, J.R.; Whaley-Connell, A.; Nistala, R. Angiotensin II Stimulation of DPP4 Activity Regulates Megalin in the Proximal Tubules. Int. J. Mol. Sci. 2016, 17, 780. [Google Scholar] [CrossRef]
- Kawase, H.; Bando, Y.K.; Nishimura, K.; Aoyama, M.; Monji, A.; Murohara, T. A dipeptidyl peptidase-4 inhibitor ameliorates hypertensive cardiac remodeling via angiotensin-II/sodium-proton pump exchanger-1 axis. J. Mol. Cell Cardiol. 2016, 98, 37–47. [Google Scholar] [CrossRef]
- Karmazyn, M.; Kilić, A.; Javadov, S. The role of NHE-1 in myocardial hypertrophy and remodelling. J. Mol. Cell Cardiol. 2008, 44, 647–653. [Google Scholar] [CrossRef]
- Filippatos, G.; Farmakis, D.; Parissis, J. Renal dysfunction and heart failure: Things are seldom what they seem. Eur. Heart J. 2014, 35, 416–418. [Google Scholar] [CrossRef]
- Segall, L.; Nistor, I.; Covic, A. Heart failure in patients with chronic kidney disease: A systematic integrative review. Biomed. Res. Int. 2014, 2014, 937398. [Google Scholar] [CrossRef] [PubMed]
- Sharkovska, Y.; Reichetzeder, C.; Alter, M.; Tsuprykov, O.; Bachmann, S.; Secher, T.; Klein, T.; Hocher, B. Blood pressure and glucose independent renoprotective effects of dipeptidyl peptidase-4 inhibition in a mouse model of type-2 diabetic nephropathy. J. Hypertens. 2014, 32, 2211–2223. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, B.P.M.; Crajoinas, R.O.; Couto, G.K.; Davel, A.P.C.; Lessa, L.M.; Rossoni, L.V.; Girardi, A.C.C. Dipeptidyl peptidase IV inhibition attenuates blood pressure rising in young spontaneously hypertensive rats. J. Hypertens. 2011, 29, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.W.; Jin, L.; Piao, S.G.; Chung, B.H.; Yang, C.W. Inhibition of dipeptidyl peptidase IV protects tacrolimus-induced kidney injury. Lab. Investig. 2015, 95, 1174–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeta, T.; Aoyama, M.; Bando, Y.K.; Monji, A.; Mitsui, T.; Takatsu, M.; Cheng, X.W.; Okumura, T.; Hirashiki, A.; Nagata, K.; et al. Dipeptidyl peptidase-4 modulates left ventricular dysfunction in chronic heart failure via angiogenesis-dependent and -independent actions. Circulation 2012, 126, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Lin, C.C. Sitagliptin attenuates inflammatory responses in lipopolysaccharide-stimulated cardiomyocytes via nuclear factor-κB pathway inhibition. Exp. Ther. Med. 2016, 11, 2609–2615. [Google Scholar] [CrossRef]
- Picatoste, B.; Ramírez, E.; Caro-Vadillo, A.; Iborra, C.; Ares-Carrasco, S.; Egido, J.; Tuñón, J.; Lorenzo, O. Sitagliptin reduces cardiac apoptosis, hypertrophy and fibrosis primarily by insulin-dependent mechanisms in experimental type-II diabetes. Potential roles of GLP-1 isoforms. PLoS ONE 2013, 8, e78330. [Google Scholar] [CrossRef]
- Zhang, H.; Xiong, Z.; Wang, J.; Zhang, S.; Lei, L.; Yang, L.; Zhang, Z. Glucagon-like peptide-1 protects cardiomyocytes from advanced oxidation protein product-induced apoptosis via the PI3K/Akt/Bad signaling pathway. Mol. Med. Rep. 2016, 13, 1593–1601. [Google Scholar] [CrossRef]
- Balteau, M.; Van Steenbergen, A.; Timmermans, A.D.; Dessy, C.; Behets-Wydemans, G.; Tajeddine, N.; Castanares-Zapatero, D.; Gilon, P.; Vanoverschelde, J.L.; Horman, S.; et al. AMPK activation by glucagon-like peptide-1 prevents NADPH oxidase activation induced by hyperglycemia in adult cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1120–H1133. [Google Scholar] [CrossRef]
- Kanasaki, K.; Shi, S.; Kanasaki, M.; He, J.; Nagai, T.; Nakamura, Y.; Ishigaki, Y.; Kitada, M.; Srivastava, S.P.; Koya, D. Linagliptin-mediated DPP-4 inhibition ameliorates kidney fibrosis in streptozotocin-induced diabetic mice by inhibiting endothelial-to-mesenchymal transition in a therapeutic regimen. Diabetes 2014, 63, 2120–2131. [Google Scholar] [CrossRef]
- Connelly, K.A.; Bowskill, B.B.; Advani, S.L.; Thai, K.; Chen, L.H.; Kabir, M.G.; Gilbert, R.E.; Advani, A. Dipeptidyl peptidase-4 inhibition improves left ventricular function in chronic kidney disease. Clin. Investig. Med. 2014, 37, E172. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Tanaka, A.; Kusunose, K.; Amano, R.; Matsuhisa, M.; Daida, H.; Ito, M.; Tsutsui, H.; Nanasato, M.; Kamiya, H.; et al. Effect of sitagliptin on the echocardiographic parameters of left ventricular diastolic function in patients with type 2 diabetes: A subgroup analysis of the PROLOGUE study. Cardiovasc. Diabetol. 2017, 16, 63. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M. Cardiac remodelling and RAS inhibition. Ther. Adv. Cardiovasc. Dis. 2016, 10, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.H.; Pang, X.F.; Bai, F.; Wang, N.P.; Shah, A.I.; McKallip, R.J.; Li, X.W.; Wang, X.; Zhao, Z.Q. Preservation of Glucagon-Like Peptide-1 Level Attenuates Angiotensin II-Induced Tissue Fibrosis by Altering AT1/AT 2 Receptor Expression and Angiotensin-Converting Enzyme 2 Activity in Rat Heart. Cardiovasc. Drugs Ther. 2015, 29, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Matsui, H.; Barry, W.H.; Livsey, C.; Spitzer, K.W. Angiotensin II stimulates sodium-hydrogen exchange in adult rabbit ventricular myocytes. Cardiovasc. Res. 1995, 29, 215–221. [Google Scholar] [CrossRef]
- Girardi, A.C.C.; Knauf, F.; Demuth, H.U.; Aronson, P.S. Role of dipeptidyl peptidase IV in regulating activity of Na +/H+ exchanger isoform NHE3 in proximal tubule cells. Am. J. Physiol. Cell Physiol. 2004, 287, C1238–C1245. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Nishida, K. Oxidative stress and left ventricular remodelling after myocardial infarction. Cardiovasc. Res. 2009, 81, 457–464. [Google Scholar] [CrossRef]
- Choi, S.Y.; Ryu, H.M.; Oh, E.J.; Choi, J.Y.; Cho, J.H.; Kim, C.D.; Kim, Y.L.; Park, S.H. Dipeptidyl peptidase-4 inhibitor gemigliptin protects against vascular calcification in an experimental chronic kidney disease and vascular smooth muscle cells. PLoS ONE 2017, 12, e0180393. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Park, S.; Oh, C.J.; Leem, J.; Park, K.G.; Lee, I.K. Dipeptidyl peptidase-4 inhibition by gemigliptin prevents abnormal vascular remodeling via NF-E2-related factor 2 activation. Vascul. Pharmacol. 2015, 73, 11–19. [Google Scholar] [CrossRef]
- Koibuchi, N.; Hasegawa, Y.; Katayama, T.; Toyama, K.; Uekawa, K.; Sueta, D.; Kusaka, H.; Ma, M.; Nakagawa, T.; Lin, B.; et al. DPP-4 inhibitor linagliptin ameliorates cardiovascular injury in salt-sensitive hypertensive rats independently of blood glucose and blood pressure. Cardiovasc. Diabetol. 2014, 13, 157. [Google Scholar] [CrossRef] [Green Version]
- Sauvé, M.; Ban, K.; Momen, M.A.; Zhou, Y.Q.; Henkelman, R.M.; Husain, M.; Drucker, D.J. Genetic deletion or pharmacological inhibition of dipeptidyl peptidase-4 improves cardiovascular outcomes after myocardial infarction in mice. Diabetes 2010, 59, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Suda, M.; Shimizu, I.; Yoshida, Y.; Hayashi, Y.; Ikegami, R.; Katsuumi, G.; Wakasugi, T.; Okuda, S.; Soga, T.; Minamino, T. Inhibition of dipeptidyl peptidase-4 ameliorates cardiac ischemia and systolic dysfunction by up-regulating the FGF-2/EGR-1 pathway. PLoS ONE 2017, 12, e0182422. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.H.; Coimbra, T.M.; de Araujo, M.; Menezes, L.F.; Seguro, A.C. N-acetylcysteine attenuates the progression of chronic renal failure. Kidney Int. 2005, 68, 2208–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, B.H.; Arruda-Junior, D.F.; Campos, L.C.G.; Barreto, A.L.T.; Rodrigues, M.V.; Krieger, J.E.; Girardi, A.C.C. Progression of microalbuminuria in SHR is associated with lower expression of critical components of the apical endocytic machinery in the renal proximal tubule. Am. J. Physiol. Renal Physiol. 2013, 305, F216–F226. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Ariyoshi, M.; Mizuno, M.; Morisue, Y.; Shimada, M.; Fujita, S.; Nasu, J.; Okada, H.; Shimomura, H.; Yamamoto, K.; Tsuji, T. Identification of a target antigen recognized by a mouse monoclonal antibody to the bile canalicular surface of rat hepatocytes with a random phage display library. Acta Med. Okayama 2002, 56, 187–191. [Google Scholar] [CrossRef]
- Rutherford, P.A.; Pizzonia, J.H.; Biemesderfer, D.; Abu-Alfa, A.; Reilly, R.; Aronson, P.S. Expression of Na(+)-H+ exchanger isoforms NHE1 and NHE3 in kidney and blood cells of rabbit and rat. Exp. Nephrol. 1997, 5, 490–497. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Parameter | Sham | Sham + IDPPIV | Nx | Nx + IDPPIV |
|---|---|---|---|---|
| Initial body weight, g | 193 ± 4 (16) | 192 ± 6 (10) | 198 ± 4 (16) | 202 ± 3 (16) |
| Final body weight, g | 323 ± 7 (9) | 331 ± 15 (7) | 311 ± 12 (10) | 337 ± 7 (10) |
| Body weight gain, % | 67 ± 3 (9) | 73 ± 5 (7) | 58 ± 6 (10) | 67 ± 3 (10) |
| Blood Glucose, mg/dL | 80 ± 3 (9) | 83 ± 2 (8) | 79 ± 1 (10) | 81 ± 2 (10) |
| Blood Creatinine, mg/dL | 0.45 ± 0.03 c (10) | 0.40 ± 0.03 c (7) | 0.99 ± 0.05 a (11) | 0.72 ± 0.03 b (12) |
| GFR, mL/min/kg | 8.2 ± 0.4 c (10) | 7.5 ± 0,9 c (7) | 2.2 ± 0.4 a (11) | 4.6 ± 0.3 b (12) |
| Blood Urea, mg/dL | 41 ± 2 c (10) | 37 ± 2 c (7) | 94 ± 4 a (11) | 70 ± 4 b (12) |
| Protein excretion, mg/24 h/kg | 60 ± 7 c (10) | 61 ± 5 c (7) | 602 ± 86 a (11) | 294 ± 43 b (12) |
| Albumin excretion, mg/24 h/kg | 9 ± 3 c (8) | 6 ± 3 c (7) | 162 ± 25 a (10) | 97 ± 11b (10) |
| Albumin/creatinine | 0.18 ± 0.05 c (8) | 0.13 ± 0.07 c (7) | 6.42 ± 0.73 a (10) | 2.91 ± 0.52 b (10) |
| Tail cuff BP, mmHg | 124 ± 5 c (11) | 126 ± 5 c (9) | 184 ± 5 a (12) | 166 ± 5 b (11) |
| Serum DPPIV activity, OD | 0.399 ± 0.017 b (16) | 0.096 ± 0.006 a (10) | 0.419± 0.012 b (16) | 0.091 ± 0.009 a (16) |
| Gene | Sense (S)/Antisense (AS) | Sequence |
|---|---|---|
| ACE | S | GACCAAAAGCTGCGAAGGAT |
| AS | TTGTTGGGGAAGCAGACCTT | |
| ACE-2 | S | TGTGGGGTAGGTTTTGGACA |
| AS | GGAAGGCCAACAGAAACGAA | |
| ACT-β | S | CTGTGACATCCGTAAGACC |
| AS | GCCACCAATCCACACAGA | |
| AGT | S | TGGATAAAGAACCCGCCTCC |
| AS | TTGAGAACCTCTCCCACTCG | |
| AGT R-1α | S | TCTGCCACATTCCCTGAGTTA |
| AS | CTTGGGGCAGTCATCTTGGA | |
| AGT R-1β | S | AGTGACAGAGACCAGACCAGA |
| AS | TTGGGGCAGTCATCTTGGATT | |
| AGT R-2 | S | TTTGCCATCCTCCTGGGATT |
| AS | GCCTTGGAGCCAAGTAATGG | |
| BNP | S | GATTCTGCTCCTGCTTTTCC |
| AS | TCTTTTGTAGGGCCTTGGTC | |
| CYCLO | S | AATGCTGGACCAAACACAAA |
| AS | CCTTCTTTCACCTTCCCAAA | |
| GAPDH | S | ATGGTGAAGGTCGGTGTG |
| AS | GAACTTGCCGTGGGTAGAG | |
| IL-1β | S | CCTGTGTGATGAAAGACGGC |
| AS | TATGTCCCGACCATTGCTGT | |
| IL-6 | S | CTGGTCTTCTGGAGTTCCGT |
| AS | GCCACTCCTTCTGTGACTCT | |
| IL-10 | S | TGGGAGAGAAGCTGAAGACC |
| AS | AGATGCCGGGTGGTTCAAT | |
| MCP-1 | S | TGCCCACTCACCTGCTGCT |
| AS | TGGGGTCAGCACAGATCTCTCTCT | |
| MAS1 | S | GACCAGTCATCCTGCCAGA |
| AS | CATGAGGAGTTCTTGTGCTGG | |
| TNF-α | S | ATCGGTCCCAACAAGGAGG |
| AS | GATAAGGTACAGCCCATCTGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beraldo, J.I.; Benetti, A.; Borges-Júnior, F.A.; Arruda-Junior, D.F.; Martins, F.L.; Jensen, L.; Dariolli, R.; Shimizu, M.H.; Seguro, A.C.; Luchi, W.M.; et al. Cardioprotection Conferred by Sitagliptin Is Associated with Reduced Cardiac Angiotensin II/Angiotensin-(1-7) Balance in Experimental Chronic Kidney Disease. Int. J. Mol. Sci. 2019, 20, 1940. https://doi.org/10.3390/ijms20081940
Beraldo JI, Benetti A, Borges-Júnior FA, Arruda-Junior DF, Martins FL, Jensen L, Dariolli R, Shimizu MH, Seguro AC, Luchi WM, et al. Cardioprotection Conferred by Sitagliptin Is Associated with Reduced Cardiac Angiotensin II/Angiotensin-(1-7) Balance in Experimental Chronic Kidney Disease. International Journal of Molecular Sciences. 2019; 20(8):1940. https://doi.org/10.3390/ijms20081940
Chicago/Turabian StyleBeraldo, Juliana Isa, Acaris Benetti, Flávio Araújo Borges-Júnior, Daniel F. Arruda-Junior, Flavia Letícia Martins, Leonardo Jensen, Rafael Dariolli, Maria Heloisa Shimizu, Antonio C. Seguro, Weverton M. Luchi, and et al. 2019. "Cardioprotection Conferred by Sitagliptin Is Associated with Reduced Cardiac Angiotensin II/Angiotensin-(1-7) Balance in Experimental Chronic Kidney Disease" International Journal of Molecular Sciences 20, no. 8: 1940. https://doi.org/10.3390/ijms20081940