Expression of Longevity Genes Induced by a Low-Dose Fluvastatin and Valsartan Combination with the Potential to Prevent/Treat “Aging-Related Disorders”


{kind=link}
Abstract
:1. Introduction
2. Results
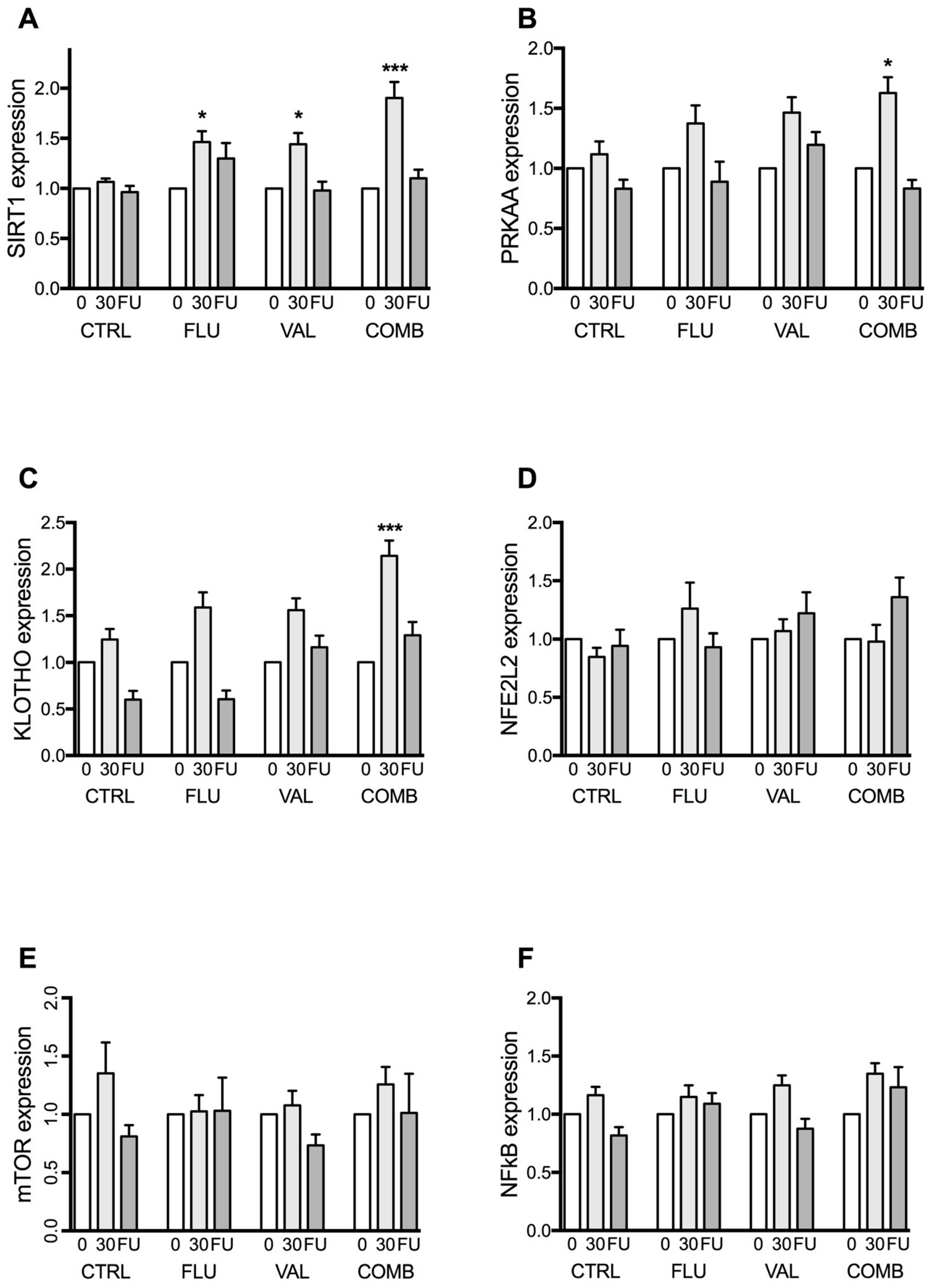
2.1. Expression of Longevity Genes
2.1.1. Sirtuin 1 (SIRT1) Gene Expression
2.1.2. 5’-AMP-Activated Protein Kinase Catalytic Subunit α-2 (PRKAA) Gene Expression
2.1.3. KLOTHO Gene Expression
2.1.4. Nuclear Factor (Erythroid-Derived 2)-Like 2 Gene Expression (NFE2L2)
2.1.5. Mechanistic Target of Rapamycin (mTOR) Gene Expression
2.1.6. Nuclear Factor κB (NF-κB) Gene Expression
2.2. Correlations between the Expression of Longevity Genes, Telomerase Activity and Arterial Wall Properties
3. Discussion
4. Materials and Methods
4.1. Participants and Study Design
4.2. Blood Sampling
4.3. RNA Isolation
4.4. Quantitative Real-Time PCR (qPCR) for Human Telomerase Reverse Transcriptase (hTERT) Expression
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| ANOVA | Analysis of variance |
| COMB | Combination group |
| CTRL | Placebo group |
| FDA | Food and Drug Administration |
| FLU | Fluvastatin group |
| FMD | Flow mediated dilation |
| hTERT | human telomerase reverse transcriptase |
| MILES | Metformin in Longevity Study |
| mTOR | Mechanistic target of rapamycin |
| NAD | Nicotinamide adenine dinucleotide |
| NF-κB1 | Nuclear factor κB |
| NFE2L2 | Nuclear factor (erythroid-derived 2)-like 2 gene |
| PRKAA | 5′-AMP-activated protein kinase catalytic subunit α-2 gene |
| PWV | Pulse wave velocity |
| qPCR | Quantitative real-time polymerase chain reaction |
| SIRT1 | Sirtuin 1 gene |
| VAL | Valsartan group |
References
- Stevens, W.; Peneva, D.; Li, J.Z.; Liu, L.Z.; Liu, G.; Gao, R.; Lakdawalla, D.N. Estimating the future burden of cardiovascular disease and the value of lipid and blood pressure control therapies in China. BMC Health Serv. Res. 2016, 16, 175. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Cardiovascular Diseases Collaboration; Roth, G.A.; Johnson, C.O.; Abate, K.H.; Abd-Allah, F.; Ahmed, M.; Alam, K.; Alam, T.; Alvis-Guzman, N.; Ansari, H.; et al. The Burden of Cardiovascular Diseases Among US States, 1990–2016. JAMA Cardiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Van Raamsdonk, J.M. Mechanisms underlying longevity: A genetic switch model of aging. Exp. Gerontol. 2017. [Google Scholar] [CrossRef]
- Rizvi, S.; Raza, S.T.; Mahdi, F. Telomere length variations in aging and age-related diseases. Curr. Aging Sci. 2014, 7, 161–167. [Google Scholar] [CrossRef]
- Yeh, J.K.; Wang, C.Y. Telomeres and Telomerase in Cardiovascular Diseases. Genes 2016, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Janic, M.; Lunder, M.; Sabovic, M. A new anti-ageing strategy focused on prevention of arterial ageing in the middle-aged population. Med. Hypotheses 2013, 80, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Lunder, M.; Janic, M.; Habjan, S.; Sabovic, M. Subtherapeutic, low-dose fluvastatin improves functional and morphological arterial wall properties in apparently healthy, middle-aged males—A pilot study. Atherosclerosis 2011, 215, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Lunder, M.; Janic, M.; Sabovic, M. Reduction of age-associated arterial wall changes by low-dose valsartan. Eur. J. Prev. Cardiol. 2012, 19, 1243–1249. [Google Scholar] [CrossRef]
- Lunder, M.; Janic, M.; Jug, B.; Sabovic, M. The effects of low-dose fluvastatin and valsartan combination on arterial function: A randomized clinical trial. Eur. J. Int. Med. 2012, 23, 261–266. [Google Scholar] [CrossRef]
- Janic, M.; Lunder, M.; Cerkovnik, P.; Prosenc Zmrzljak, U.; Novakovic, S.; Sabovic, M. Low-Dose Fluvastatin and Valsartan Rejuvenate the Arterial Wall Through Telomerase Activity Increase in Middle-Aged Men. Rejuvenation Res. 2016, 19, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Janic, M.; Lunder, M.; Prezelj, M.; Sabovic, M. A combination of low-dose fluvastatin and valsartan decreases inflammation and oxidative stress in apparently healthy middle-aged males. J. Cardiopulm. Rehabil. Prev. 2014, 34, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Mori, R.; Shimokawa, I. Do sirtuins promote mammalian longevity? A critical review on its relevance to the longevity effect induced by calorie restriction. Mol. Cells 2013, 35, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Jayasena, T.; Poljak, A.; Sachdev, P.S. Sirtuins in cognitive ageing and Alzheimer’s disease. Curr. Opin. Psychiatry 2012, 25, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Song, Y.; Zhang, T.; Ma, L.; Bian, N.; Chen, X.; Feng, J.; Chang, Q.; Li, Z. Simvastatin attenuates TNFα-induced apoptosis in endothelial progenitor cells via the upregulation of SIRT1. Int. J. Mol. Med. 2014, 34, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Kilic, U.; Gok, O.; Elibol-Can, B.; Uysal, O.; Bacaksiz, A. Efficacy of statins on sirtuin 1 and endothelial nitric oxide synthase expression: The role of sirtuin 1 gene variants in human coronary atherosclerosis. Clin. Exp. Pharmacol. Physiol. 2015, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Qian, J.; Castillo, A.C.; Perez-Polo, J.R.; Birnbaum, Y. Aliskiren and Valsartan reduce myocardial AT1 receptor expression and limit myocardial infarct size in diabetic mice. Cardiovasc. Drugs Ther. 2011, 25, 505–515. [Google Scholar] [CrossRef]
- Shiota, A.; Shimabukuro, M.; Fukuda, D.; Soeki, T.; Sato, H.; Uematsu, E.; Hirata, Y.; Kurobe, H.; Maeda, N.; Sakaue, H.; et al. Telmisartan ameliorates insulin sensitivity by activating the AMPK/SIRT1 pathway in skeletal muscle of obese db/db mice. Cardiovasc. Diabetol. 2012, 11, 139. [Google Scholar] [CrossRef] [PubMed]
- Pantazi, E.; Bejaoui, M.; Zaouali, M.A.; Folch-Puy, E.; Pinto Rolo, A.; Panisello, A.; Palmeira, C.M.; Rosello-Catafau, J. Losartan activates sirtuin 1 in rat reduced-size orthotopic liver transplantation. World J. Gastroenterol. 2015, 21, 8021–8031. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, Y.; Vanhoutte, P.M. SIRT1 and AMPK in regulating mammalian senescence: A critical review and a working model. FEBS Lett. 2011, 585, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Wu, C.; Chen, Z.; Lu, G. Atorvastatin inhibits homocysteine-induced endoplasmic reticulum stress through activation of AMP-activated protein kinase. Cardiovasc. Ther. 2012, 30, 317–325. [Google Scholar] [CrossRef]
- Sun, W.; Lee, T.S.; Zhu, M.; Gu, C.; Wang, Y.; Zhu, Y.; Shyy, J.Y. Statins activate AMP-activated protein kinase in vitro and in vivo. Circulation 2006, 114, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Hermida, N.; Markl, A.; Hamelet, J.; Van Assche, T.; Vanderper, A.; Herijgers, P.; van Bilsen, M.; Hilfiker-Kleiner, D.; Noppe, G.; Beauloye, C.; et al. HMGCoA reductase inhibition reverses myocardial fibrosis and diastolic dysfunction through AMP-activated protein kinase activation in a mouse model of metabolic syndrome. Cardiovasc. Res. 2013, 99, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.M.; Park, E.J.; Kang, Y.J.; Park, S.W.; Kim, H.J.; Chang, K.C. Valsartan independent of AT(1) receptor inhibits tissue factor, TLR-2 and -4 expression by regulation of Egr-1 through activation of AMPK in diabetic conditions. J. Cell. Mol. Med. 2014, 18, 2031–2043. [Google Scholar] [CrossRef]
- Kurokawa, H.; Sugiyama, S.; Nozaki, T.; Sugamura, K.; Toyama, K.; Matsubara, J.; Fujisue, K.; Ohba, K.; Maeda, H.; Konishi, M.; et al. Telmisartan enhances mitochondrial activity and alters cellular functions in human coronary artery endothelial cells via AMP-activated protein kinase pathway. Atherosclerosis 2015, 239, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Donate-Correa, J.; Martin-Nunez, E.; Mora-Fernandez, C.; Muros-de-Fuentes, M.; Perez-Delgado, N.; Navarro-Gonzalez, J.F. Klotho in cardiovascular disease: Current and future perspectives. World J. Biol. Chem. 2015, 6, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Karalliedde, J.; Maltese, G.; Hill, B.; Viberti, G.; Gnudi, L. Effect of renin-angiotensin system blockade on soluble Klotho in patients with type 2 diabetes, systolic hypertension, and albuminuria. Clin. J. Am. Soc. Nephrol. 2013, 8, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.E.; Lim, S.W.; Piao, S.G.; Song, J.H.; Kim, J.; Yang, C.W. Statin upregulates the expression of KLOTHO, an anti-aging gene, in experimental cyclosporine nephropathy. Nephron. Exp. Nephrol. 2012, 120, e123–e133. [Google Scholar] [CrossRef] [PubMed]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Yvonne Alexander, M.; White, S.J. The Role of Nrf2 in Cardiovascular Function and Disease. Oxid. Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef]
- Hwang, A.R.; Han, J.H.; Lim, J.H.; Kang, Y.J.; Woo, C.H. Fluvastatin inhibits AGE-induced cell proliferation and migration via an ERK5-dependent Nrf2 pathway in vascular smooth muscle cells. PLoS ONE 2017, 12, e0178278. [Google Scholar] [CrossRef]
- Makabe, S.; Takahashi, Y.; Watanabe, H.; Murakami, M.; Ohba, T.; Ito, H. Fluvastatin protects vascular smooth muscle cells against oxidative stress through the Nrf2-dependent antioxidant pathway. Atherosclerosis 2010, 213, 377–384. [Google Scholar] [CrossRef]
- Sciarretta, S.; Volpe, M.; Sadoshima, J. Mammalian target of rapamycin signaling in cardiac physiology and disease. Circ. Res. 2014, 114, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Long, C.; Zhu, J.; Tian, J. Protective Effects of Fluvastatin on Reproductive Function in Obese Male Rats Induced by High-Fat Diet through Enhanced Signaling of mTOR. Cell. Physiol. Biochem. 2017, 41, 598–608. [Google Scholar] [CrossRef]
- Wagner, R.J.; Martin, K.A.; Powell, R.J.; Rzucidlo, E.M. Lovastatin induces VSMC differentiation through inhibition of Rheb and mTOR. Am. J. Physiol. Cell Physiol. 2010, 299, C119–C127. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; He, L.; Cai, Y.; Zhang, G.; He, Y.; Zhang, Z.; He, X.; He, Y.; Zhang, G.; Luo, J. Induction of autophagy contributes to the myocardial protection of valsartan against ischemiareperfusion injury. Mol. Med. Rep. 2013, 8, 1824–1830. [Google Scholar] [CrossRef] [PubMed]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, metabolism and cardiovascular disease. J. Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Lenglet, S.; Quercioli, A.; Fabre, M.; Galan, K.; Pelli, G.; Nencioni, A.; Bauer, I.; Pende, A.; Python, M.; Bertolotto, M.; et al. Statin treatment is associated with reduction in serum levels of receptor activator of NF-κB ligand and neutrophil activation in patients with severe carotid stenosis. Mediators Inflamm. 2014, 2014, 720987. [Google Scholar] [CrossRef]
- Wu, B.; Lin, R.; Dai, R.; Chen, C.; Wu, H.; Hong, M. Valsartan attenuates oxidative stress and NF-κB activation and reduces myocardial apoptosis after ischemia and reperfusion. Eur. J. Pharmacol. 2013, 705, 140–147. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janić, M.; Lunder, M.; Novaković, S.; Škerl, P.; Šabovič, M. Expression of Longevity Genes Induced by a Low-Dose Fluvastatin and Valsartan Combination with the Potential to Prevent/Treat “Aging-Related Disorders”. Int. J. Mol. Sci. 2019, 20, 1844. https://doi.org/10.3390/ijms20081844
Janić M, Lunder M, Novaković S, Škerl P, Šabovič M. Expression of Longevity Genes Induced by a Low-Dose Fluvastatin and Valsartan Combination with the Potential to Prevent/Treat “Aging-Related Disorders”. International Journal of Molecular Sciences. 2019; 20(8):1844. https://doi.org/10.3390/ijms20081844
Chicago/Turabian StyleJanić, Miodrag, Mojca Lunder, Srdjan Novaković, Petra Škerl, and Mišo Šabovič. 2019. "Expression of Longevity Genes Induced by a Low-Dose Fluvastatin and Valsartan Combination with the Potential to Prevent/Treat “Aging-Related Disorders”" International Journal of Molecular Sciences 20, no. 8: 1844. https://doi.org/10.3390/ijms20081844