Post-Translational Modification-Dependent Activity of Matrix Metalloproteinases


Department of Biotechnology and Biomedicine, Technical University of Denmark, DK-2800 Kongens Lyngby, Denmark
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(12), 3077; https://doi.org/10.3390/ijms20123077
Submission received: 27 May 2019
/
Revised: 16 June 2019
/
Accepted: 18 June 2019
/
Published: 24 June 2019
(This article belongs to the Special Issue Matrix Metalloproteinase)
Abstract
:Due to their capacity to process different proteins of the extracellular matrix (ECM), matrix metalloproteinases (MMPs) were initially described as a family of secreted proteases, functioning as main ECM regulators. However, through proteolytic processing of various biomolecules, MMPs also modulate intra- and extracellular pathways and networks. Thereby, they are functionally implicated in the regulation of multiple physiological and pathological processes. Consequently, MMP activity is tightly regulated through a combination of epigenetic, transcriptional, and post-transcriptional control of gene expression, proteolytic activation, post-translational modifications (PTMs), and extracellular inhibition. In addition, MMPs, their substrates and ECM binding partners are frequently modified by PTMs, which suggests an important role of PTMs in modulating the pleiotropic activities of these proteases. This review summarizes the recent progress towards understanding the role of PTMs (glycosylation, phosphorylation, glycosaminoglycans) on the activity of several members of the MMP family.
1. MMP Domain Structure and Classification
Matrix metalloproteinases (MMPs) comprise a family of 23 distinct secreted or membrane-anchored endopeptidases in humans that belong to the metzincin superfamily of metalloproteases. MMPs were initially described as regulators of the extracellular matrix due to their capacity to degrade ECM proteins like collagen, gelatin, laminin, aggrecan, fibronectin, elastin, and proteoglycans. Based on structure and substrate specificity, the MMPs are divided into collagenases (MMP1, MMP8, MMP13), gelatinases (MMP2, MMP9), stromelysins (MMP3, MMP10, MMP11), matrilysins (MMP7, MMP26), membrane-type MMPs (MMP14, MMP15, MMP16, MMP17, MMP24, MMP25), and other non-classified family members (MMP12, MMP19, MMP20, MMP21, MMP23, MMP27, MMP28). MMPs share a multidomain structure consisting of signal peptide, zymogenic pro-peptide domain, a catalytic domain, a variable linker ‘hinge’ region and a hemopexin domain. Some MMPs show variation in the domain arrangements, e.g., MMP7 and MMP26 are lacking the linker and hemopexin domain, MMP23 has a unique cysteine-rich and immunoglobulin-like instead of a hemopexin domain, and the two gelatinases MMP2 and MMP9 have additional fibronectin type-II-related domains. Membrane-type MMPs are anchored to the membrane either via glycosylphosphatidylinositol (GPI) or with help of a transmembrane domain (Figure 1). Additionally, MMPs are distinguished by the highly conserved HExGHxxGxxH motif in the catalytic domain, which contains three histidines that coordinate the zinc molecule in the active site, and the PRCGxPD motif identified in the pro-domain whose cysteine residue coordinates with the active zinc molecule to inhibit proteolysis. Each of these domains is associated with a specific function and very frequently post-translationally modified. This suggests an important role of PTMs in modulating the pleiotropic activities of MMPs [1,2].
2. MMP Substrates and Function
MMPs are primarily extracellular proteases, supporting the initial concept that they are generally associated with degradation and regulation of the ECM, thereby influencing many fundamental cellular events involving ECM remodeling [2,3,4,5]. More recently, it was observed that they also can irreversibly process growth factors, cell-surface receptors, cytokines, and chemokines, as well as other MMPs, other proteases and protease inhibitors and even act inside the cell [6]. The activation or inactivation of bioactive molecules by MMP proteolytic processing unraveled unexpected roles of these proteases in the regulation of extra- and intracellular signaling pathways [4,7,8,9]. Hence, the field of MMP research progressed from perceiving these enzymes solely as regulators of ECM to the conception that MMPs are functionally implicated in the regulation of multiple physiological and pathological signaling processes [8,10]. Accordingly, the physiological relevance of their function is emphasized by a direct association between altered expression and/or dysregulation of MMPs and development of pathological conditions, such as chronic inflammatory diseases, vascular diseases, neurological disorders, and cancer [10,11].
3. Multilayered Regulation of MMP Activity
Since MMPs regulate major physiological processes, a strict spatiotemporal control of their activity is essential to avoid possible detrimental activities of these proteases. Primarily, expression of MMPs is regulated at the transcriptional level, keeping these enzymes at very low levels in normal tissue homeostasis. MMPs share cis-regulatory elements in their promoter sequences, which allow induction of their expression by stimuli—e.g., in the form of growth factors, cytokines, or hormones. The cooperation between these cis-regulatory elements, coupled with the integration of multiple signaling pathways, provides a wide range of potential interactions between transcriptional regulators, ensuring tissue-specific expression of diverse MMP family members and facilitating a strict control of MMP transcriptional activity. In addition, transcription of MMP encoding genes is regulated by epigenetic mechanisms, such as DNA methylation or histone acetylation [4,5,11,12]. At the post-transcriptional level, regulation of MMP expression is mediated by modulation of mRNA stability and miRNA-based mechanisms that interfere with MMP expression by either transcriptional inhibition or mRNA degradation [13,14,15,16]. An important level of MMP regulation is achieved at the post-translational level, since most MMPs are secreted as inactive pro-enzymes. The ‘cysteine-switch’ region in the pro-peptide domain shields the active site, thereby preventing substrate access and maintaining the enzyme in an inactive state. The activation of proMMPs occurs upon direct proteolytic cleavage of the pro-domain (within the secretory pathway or extracellularly), or induction of conformational changes that disrupt the chelating cysteine residue and enable auto-proteolysis for removal of the pro-domain region [9,12]. Moreover, allosteric activation of proMMPs can be triggered by interaction with ECM components and cell surface molecules and by low-density lipoprotein receptor-related protein (LRP1)-mediated endocytosis [12,17,18,19,20]. Upon activation, mature MMPs are exclusively controlled by endogenous inhibitors, such as tissue inhibitors of metalloproteases (TIMPs) and α2-macroglobulin [21,22].
4. PTMs—An Additional Level of Protein Regulation
The enormous functional complexity of the cellular proteome is regulated by diverse mechanisms, including transcription, alternative splicing, translation, and PTMs [23]. Among these regulatory mechanisms, PTMs provide a significant genome-independent expansion and diversification of the proteome, thereby creating a continuously fine-tuned regulatory network implicated in many cellular processes [24,25,26]. Based on the type of modification, PTMs can be assigned to several categories: chemical modifications, including methylation, phosphorylation, acetylation, and oxidation; polypeptide modifications, including ubiquitination, SUMOylation and other ubiquitin-like protein conjugation; modifications by complex molecules, including glycosylation, lipids (e.g., acylation, prenylation) and extended structures (e.g., glycosaminoglycans (GAGs)); and modifications of the amino acids or of the polypeptide backbone, including deamidation, eliminylation, and protein cleavage through proteolysis [27,28,29,30]. By reversible or irreversible addition of these functional groups PTMs can modify protein function by altering protein structure, subcellular localization, protein–protein interactions, and degradation, thereby influencing many cellular processes in health and disease [31,32,33,34,35,36].
Like many enzymes, MMPs as well as their substrates are modified by a variety of PTMs. Different domains of the MMP structure are associated with specific functions and undergo different, highly specific PTM modifications, which suggests an important role of PTMs in modulating the pleiotropic activities of MMPs (Figure 2) [1]. Understanding the influence of specific PTMs on the activity of MMPs is necessary to fully understand MMP regulation. In this review, we provide an overview of different PTMs with a focus on glycosylation, phosphorylation, and interaction with extracellular GAGs, and describe their effects on activity of various MMP family members.
4.1. Glycosylation of MMP
Glycosylation describes the enzymatic reaction that links saccharides to lipids, other saccharides or proteins and represents the most abundant and diverse PTM. The most common forms of protein glycosylation are the N- and O-linked glycosylation [37,38]. N-linked glycosylation is a glycosidic linkage of glycans to the side chains of asparagine (Asn) residues. It is initiated by a single oligosaccharyltransferase complex with a transfer of N-acetylglucosamine (GlcNAc) phosphate at the consensus Asn-Xaa-Ser/Thr sequence (X, indicating any amino acid excluding proline) on the cytosolic side of the endoplasmic reticulum (ER) and extensively modified further in the lumen of the ER and Golgi prior to sorting to secretory vesicles [37,39,40,41]. O-linked glycosylation is a glycosidic linkage of glycans to the side chains of serine/threonine (Ser/Thr) or tyrosine (Tyr) residues. O-linked glycosylation shows a higher diversity than N-glycans, resulting from the complex synthesis of N-acetylgalactosamine (GalNAc)-type O-linked glycans in the Golgi, which can be initiated by up to 20 different polypeptide GalNAc-transferase isoforms that may compete for the same glycosylation site and the same glycan. Upon transfer of a GalNAc moiety from a donor substrate uridine diphosphate (UDP)-GalNAc to Ser/Thr residues, the O-linked glycan is branched out by different core-forming enzymes and further extended by fucosylation and sialylation to the final O-linked glycan structure [41,42,43,44,45]. Protein glycosylation regulates essential biological processes, such as protein folding, secretion, cell adhesion, and inter- and intra-cellular trafficking [46,47,48,49,50]. Consequently, alterations in glycosylation patterns are often associated with different pathological conditions like neurodegenerative diseases, diabetes, inflammatory conditions, and cancer [51,52]. The majority of MMPs are glycosylated, and N- and O-linked glycosylation are present across the MMP family [39]. The following section summarizes the current structural and functional information about glycosylation of different MMPs.
4.1.1. MMP9
MMP9 is the most extensively glycosylated MMP. This protease contains two N-linked glycosylation sites, Asn38 and Asn120 (asparagine residues 38 and 120), in the pro-domain and the catalytic domain, respectively [53]. These N-linked glycans have been described as core-fucosylated biantennary structures, partially sialylated with variable fucosylation branches [54]. Despite their similar composition, the individual functions of these N-linked glycosylations are distinct, primarily due to their location within the protease. Since N-linked glycosylation is generally required for protein secretion, initially a potential role of these two glycans was related to the secretion of MMP9 [55]. Indeed, abrogation of MMP9 glycosylation at Asn120 reduced the efficiency of its secretion by increasing the interaction between MMP9 and calreticulin (a protein that prevents misfolded proteins from entering the secretory pathway), thereby inducing MMP9 retention in the ER. The glycosylation at Asn38 does not influence the secretion of MMP9, but Asn38-glycosylation-deficient MMP9 shows strong amino-acid dependency towards interaction with calreticulin, likely affecting MMP9 secretion in an N-glycosylation-independent manner [56]. Additionally, N-glycosylation at Asn38 was commonly associated with the activation of proMMP9, yet the process of MMP9 activation occurs independently of the glycosylation at Asn38. Recent molecular dynamics simulations suggest that the glycosylation at Asn38 is indirectly involved in the activation of proMMP9 by inducing conformational changes within the pro-domain, enabling MMP3 to access the two cleavage sites for proteolytic activation [57]. Moreover, interactions between the glycosylation at Asn38 and galectin 3 decrease the proteolytic activation of MMP9, whereas interactions with galectin 8 enhance the MMP3-mediated processing, suggesting that the presence of N-glycosylation is important for a fine-tuned regulation of MMP9 activity [58,59]. Furthermore, MMP9 has a proline-rich linker sequence between the active site and the hemopexin domain, which contains 14 O-linked glycans (denominated OG domain) [60]. Based on their glycan composition, these O-linked glycans comprise a heterogenic mixture, varying from core-1 (Galβ1-3GalNAc) to core-2 (Gal-(GlcNAc-)GalNAc) structures and further elongated to larger glycans [61]. The presence of these O-linked glycans increases the domain flexibility of MMP9, allowing the protease to adopt multiple enzyme conformations and facilitating individual movements of the catalytic and the hemopexin domain. This influences the recognition, binding, and processing of substrates, cell receptors, and endogenous inhibitors [62,63]. A study by den Steen et al. described that the OG domain is indispensable for correct orientation of the hemopexin domain for MMP9 internalization and degradation by LRP-1 and LRP-2, as well as inhibition by endogenous TIMP1 [60]. Consequently, deletion of this linker region significantly reduced the affinity towards TIMP1 and disrupted the interactions with LRP-1 and LRP-2 [60]. However, since deletion of the OG domain did not affect the activity of MMP9, the authors reported that the OG domain functions as a regulator of extracellular bioavailability of the protease, rather than as a regulator of its activity [60]. An independent analysis of MMP9 lacking the OG domain performed by Vandooren et al. showed a reduced gelatinolytic activity of the truncated protease [64]. Furthermore, a study by Dufour et al. reported reduced MMP9-induced cell migration in endothelial cells expressing MMP9 without the OG domain, which was independent of the proteolytic activity of the enzyme [26].
4.1.2. MMP14
Similar to MMP9, the transmembrane collagenase MMP14 (MT1-MMP) contains O-linked glycosylation sites (Thr291, Thr299, Thr300, and Ser301) identified in the linker region. The presence of these O-linked glycans does not affect zymogen activation, collagenase activity or the autolytic processing of MMP14. However, the O-linked glycosylation is important for the formation of a stable complex between MMP14, TIMP2, and proMMP2, which is required for subsequent cell-surface activation of MMP2. Being unable to recruit TIMP2, which connects the catalytic domain of MMP14 and the hemopexin domain of proMMP2, the glycan-deficient MMP14 cannot present a stable trimeric complex on the cell surface and thereby activate MMP2 [65,66,67]. Interestingly, although it has not appeared that glycosylation could directly affect MMP14 activity, recent reports describe increase of MMP14 activity upon alterations in the glycosylation pattern. Indeed, hyper-glycosylation of MMP14 led to higher proteolytic activity and promoted tumor growth [68].
4.1.3. MMP1
MMP1 has two potential glycosylation sites in the active site, but only N-glycosylation at Asn120 has been experimentally confirmed. Comparison between the glycosylated and non-glycosylated MMP1 showed no significant differences in activity, substrate specificity or inhibitory profiles of the two proteoforms. However, the presence of specific glycan motifs (e.g., α1,3-fucosylated LacdiNAc) can initiate MMP1 selectin-mediated binding to the surface of activated cells through a selectin/glycan interface and therefore may have a profound effect on cell migration [39,69].
4.1.4. MMP2
MMP2 has two potential N-linked glycosylation sites, Asn573 and Asn642, in the hemopexin domain [70,71]. The function of these N-linked oligosaccharides remains unclear. However, there are strong implications regarding their involvement in the regulation of MMP2, since the hemopexin domain is involved in MMP2 activation/inhibition, localization of its catalytic activity, and induction of cell signaling upon interaction of MMP2 with cell-surface receptors [71,72].
4.1.5. MMP3
Based on the consensus sequence for N-linked glycosylation, MMP3 also has two potential N-linked glycosylation sites, Asn120 in the catalytic domain and Asn398 in the hemopexin domain. However, only a small portion of MMP3 (~20%) is glycosylated, and no correlation has been established between these glycans and the function of the protease [39,73].
4.1.6. MMP13
MMP13 was shown to be potentially glycosylated at two asparagine residues (Asn117 and Asn152) in the catalytic domain. The N-linked glycosylation site at Asn117 has been experimentally verified, but the function of this N-linked glycan has not been determined since no differences were observed between glycosylated and non-glycosylated recombinant MMP13 [74,75].
4.1.7. MMP17
MMP17 (MT4-MMP), a glycosylphosphatidylinositol-anchored matrix metalloproteinase, has two N-linked glycans at Asn137 and Asn318 in the catalytic site and the linker region, respectively. The presence of these N-linked oligosaccharides stabilizes the dimeric form of MMP17 by promoting non-covalent interactions or facilitating folding and formation of disulfides [76,77,78].
4.2. Phosphorylation of MMPs
Protein phosphorylation is one of the most important PTMs for regulation of biological processes [79]. This dynamic PTM is characterized by the reversible enzymatic addition of a phosphate group to amino acid side chains of serine (Ser), threonine (Thr), or tyrosine (Tyr), resulting in alterations of protein structure, stability, and dynamics [79]. Hence, phosphorylation can initiate different conformational changes associated with differential activity and binding specificity, leading to an activation or deactivation of proteins [80,81]. Furthermore, the dynamic nature of phosphorylation resulting from the opposing activities of kinases (which add phosphate groups) and phosphatases (which remove phosphate groups) is functionally compatible with the modulation of intramolecular interactions, implicated in essential cellular processes, such as cell division, cellular proliferation and differentiation, apoptosis, and signal transduction [79]. Consequently, dysregulation of phosphorylation dynamics is often related with pathologies, e.g., tumor formation, chronic inflammatory diseases, autoimmune diseases, and neurodegenerative disorders [81,82].
As many secreted and extracellular proteins, the MMPs are phosphorylated. Ser-, Thr-, and Tyr phosphorylation sites are identified across the different structural domains of the MMPs. Yet, the biological significance of these phosphate groups is overall poorly characterized. Among the MMPs, MMP2 and membrane-type MMP14 metalloproteinase are the ones with most extensively studied phosphorylation-dependent effects.
4.2.1. MMP2
MMP2 contains 29 potential phosphorylation sites distributed across the pro-peptide domain, collagen-binding domain, collagenase-like domain-1 and -2, and hemopexin domain. However, only five of the predicted phosphorylation sites (S32, S160, Y271, T250, and S365) have been confirmed by mass spectrometry (MS). The phosphorylation of MMP2 noticeably diminishes its proteolytic activity, while dephosphorylation increases MMP2 activity. This is possibly due to the conformational changes observed in the secondary structure of dephosphorylated MMP2, with α-helices 50% longer and β-strands 17% shorter than phosphorylated MMP2. However, it is still unclear if protease activity is directly influenced by these conformational changes [83,84].
4.2.2. MMP14
MMP-14 phosphorylation is known to play an important role in fine-tuning the activity of MMP14 at the cell surface and in the activation of intracellular signaling, and it is critical for regulating the pro-metastatic function of this metalloproteinase. This membrane-type metalloprotease has nine potential phosphorylation sites in its multidomain structure (PhosphoSitePlus [85], UniProt [86]). The cytoplasmic domain of MMP14 regulates its internalization and trafficking, thereby modulating enzymatic activity at the cell surface. A study by García-Pardo et al. reported that this domain was essential for MMP14 mediated cellular invasion and migration [72]. The cytoplasmic domain of MMP14 contains two phosphorylation sites, at Thr567 and Tyr573, which have a significant influence on MMP14-induced cellular invasion and migration. A study by Williams et al. in fibrosarcoma cells showed that substitution of the residue Thr567 by alanine increased retention at the cell surface and reduced internalization of the protease significantly compromising invasion and migration. Contrarily, mimicking protease phosphorylation by substitution of Thr567 by glutamic acid reduced retention of MMP14 at the cell surface, increased efficiency of internalization, and was correlated with an increase in migration and invasion [87]. A study by Moss et al. showed that phospho-mimetic Thr567 mutants exhibit higher collagenolytic activity and three-dimensional growth within a collagen matrix, thereby promoting enhanced matrix invasion in ovarian cancer cells [88]. In addition, phosphorylation of Thr567 impacted the integrity of cell monolayer, cell motility and multicellular aggregate dynamics in ovarian cancer cells, promoting metastasis-associated behaviors [89]. Furthermore, a study by Nyalendo et al. reported that phosphorylation at Tyr573 influenced cell migration, suggested by the ability of a phospho-defective mutant to inhibit migration of cells endogenously expressing MMP14 [90]. Additionally, epidermal growth factor (EGF)-induced phosphorylation of Tyr573 prompted internalization of MMP14 together with pericellular collagen, establishing an environment for expansive growth in three–dimensional collagen matrix, whereas lack of responding to EGFR signaling triggered invasive growth. Therefore, the phosphorylation of Tyr573 modulates cell surface dynamics of MMP14, thereby regulating the transition between invasive and expansive growth [91].
4.2.3. Extracellular Phosphorylation of MMPs
A subset of MMPs (MMP1, 12, 13, 14, 16, 24, and 27) are tyrosine phosphorylated by the extracellular vertebrate lonesome kinase (VLK). These phosphotyrosines are found exclusively within the hemopexin domains of the MMPs. Interestingly, proteins distantly related to MMPs, which contain hemopexin-like domains, were found to be tyrosine phosphorylated at identical positions by VLK. The structural conservation of these phosphorylation sites suggests that they play a role in regulating MMP activity, a potential function, which still remains largely undiscovered [92,93]. Additionally, a study by Bordoli et al. showed that co-expression of VLK with MMPs supported their tyrosine phosphorylations that have been extensively observed in vivo. Bordoli also showed that by introducing a mutation in the ATP binding site of VLK or deletion of the proline glycine-rich domain (a conserved domain close to the kinase domain), the phosphorylation of the MMPs and other co-expressed substrates in the extracellular environment was eliminated. Similarly, shRNA-mediated downregulation of VLK expression and substitution of Tyr360 with phenylalanine in MMP1 reduced MMP13 and MMP1 tyrosine phosphorylation, respectively [93]. While VLK generates the majority of extracellular phosphotyrosines, additional secreted kinases have been identified, which may also be responsible for the extracellular protein phosphorylation events observed in vivo. This indicates a major impact of the secreted kinome on tissue homeostasis and disease pathogenesis. Discovery of novel extracellular kinases, identification of their substrates, and dissection of the regulatory mechanisms involved will provide us with a better understanding of their functions [94,95,96].
4.3. Glycosaminoglycans
An additional layer of regulation of extracellular proteolysis is mediated by interactions of MMPs with glycans in the extracellular space rather than direct glycosylation or phosphorylation of the protease or substrate. Glycosaminoglycans (GAGs) are extracellular glycans, which are composed of linear, unbranched repeats of disaccharide units. Heparan sulfate (HS) is a GAG-family member expressed by virtually every cell of a multicellular organism composed of repeats of glucoronic acid (GlcA) and N-acetylglucosamine (GlcNAc) dimers, which in most cases are attached to a cell-membrane associated core protein. During GAG-chain synthesis HS undergoes several sulfotransferase-mediated N- and O-sulfations. These modifications do not undergo completion, which results in strongly negatively charged sugar chains with variable length and degree of sulfation interrupted by stretches of unmodified, neutral regions. The sulfated regions bind basic peptide motifs of many signaling molecules, including morphogens (Hedgehogs, Wnts, TGFs), cytokines, and most chemokines [97,98,99]. This electrostatic interaction leads to partial neutralization of the protein surface and can have different effects on the biological functions of the interacting protein by facilitating, enhancing, or inhibiting the interaction with other proteins. It is important to note that these interactions, even though they are mainly based on electrostatics, are in most cases very specific and do not occur randomly. This is shown by the fact that many proteins contain specific GAG-binding motifs, with which they exclusively bind HS, even though they contain many lysine and arginine residues (for an in-depth review of the molecular basis of HS-protein interactions see Xu & Esko, 2014 [100]). Examples of HS-binding proteins are members of the fibroblast growth factor (FGF) family. Here, HS is part of the trimeric FGF/FGF-receptor/HS co-receptor complexes and activates FGF signaling in a sulfation pattern-dependent manner [101,102,103,104]. HS-induced oligomerization has been shown in many other processes including dimerization of amyloid precursor protein (APP) complexes or nearly all chemokines [105,106,107,108]. Interestingly, HS does not only mediate protein complex formation, but many proteins binding to HS, including the aforementioned candidates, are also known to undergo proteolytic processing on or near the cell surface. This indicates a crucial role of HS in regulating extracellular cleavage processes.
4.3.1. GAG-regulated Substrate Proteolysis
Signaling processes in the extracellular space usually involve at least one form of substrate proteolysis. Therefore, co-factors have to act as decision makers to ensure regulated processing of signaling proteins. Multiple cases have been reported, where this is mediated by substrate binding to GAGs. For example, similar to N-glycans, GAGs can modulate the stability of substrates. FGF2 binding to heparin, an extremely sulfated HS variant expressed by connective-tissue type macrophages, stabilizes FGF2 and protects it from degradation by proteolysis [109,110]. The same stabilizing effect has been shown for the degradation of stromal cell-derived factor 1 (SDF1)/CXCL12 and C-terminal processing of interferon-γ [111,112]. Despite this stabilizing effect GAGs have been demonstrated to play additional important roles in extracellular proteolysis. In the case of shedding of morphogens from the Hedgehog (Hh) family, HS acts as an assembly platform for protease-release complexes [113]. Here, HS recruits Hh co-factors that co-localize proteases and allow for cleavage of the substrate (Figure 3) [114]. Furthermore, direct HS binding of the N-terminal cleavage site of Hh also has a stabilizing effect by inhibiting N-terminal Hh processing and reduces its release in vitro and in Drosophila in vivo [114,115]. An example of how GAG binding can positively regulate substrate cleavage (Figure 3) was shown for the cleavage of viral capsid proteins. Binding of the human papillomavirus viral capsid protein L1 to highly sulfated HS is essential to induce a conformational change and leads to cleavage of L1 by the human trypsin-like serine proteinase kallikrein 8, a process which is required for virus internalization [116].
4.3.2. GAG-regulated MMP Activity
GAGs also regulate extracellular proteolysis at the protease level by modulating activation, localization, and protease–substrate interactions (Figure 3 middle). Many secreted vertebrate MMPs associate to the cell surface via binding to HS, including MMP9, which is specific, since excess soluble heparin extracts and solubilizes MMP2, MMP7, MMP9, and MMP13 [117,118]. Moreover, heparin, a highly sulfated form of HS, affects protease activity by increasing TIMP3 affinity to MMP2, MMP7, and MMP9, and HS affects MMP9 expression and plasma levels, depending on its sulfation and expressing cell type [119,120,121,122,123]. Finally, in vivo studies showed that MMPs in secretory granules of mast cells interact with heparin and that loss of heparin sulfation reduces MMP stability [124,125]. Like all MMPs, the wound repair and immune reaction associated MMP7 has to be converted into its proteolytic active form by removal of its inhibitory pro-sequence. MMP7 is able to activate itself by intermolecular autolytic processing. This is amplified by binding of MMP7 to highly sulfated GAGs (e.g., heparin or chondroitin sulfate), which significantly increases MMP7 auto-processing and also increases cleavage of specific physiological substrates [19].
5. Conclusions
MMP biology has been revolutionized with the recognition of extracellular proteolysis not as a simple mechanism of ECM degradation but as a regulatory mechanism for precise cellular control of biological processes. The paradigm shift for MMP functions from ECM degradative proteases to important regulators of essential cellular processes has highlighted the physiological relevance of these proteases, directly implicated by the relationship between MMP expression and disease development [4,8]. Hence, the multilayered regulation of MMPs emphasizes the tremendous importance of the balance between synthesis of active enzymes and their inhibition, which is pivotal to avoid the destructive activity of these proteases. Characterization of these regulatory mechanisms will aid the development of new therapeutics for various pathologies. MMP activity is regulated at the transcriptional level, post-translationally by pro-enzyme activation, by PTMs, and through extracellular inhibition by TIMPs and by non-specific proteinase inhibitors. Among these regulatory mechanisms, PTMs have recently obtained wide attention in the MMP community. MMPs are modified by PTMs at multiple sites (Table 1), which affects the activity of individual MMPs to a different extent, but the biological relevance of many of them is still unknown. However, many of the studies referenced in this review have been performed prior to the rapid progression of proteomics method development within the last decade. Many of the challenges that still limit our understanding of PTM function in MMP biology can now be approached by modern technologies of state-of-the-art proteomics. Advances in MS-based methods including multiplexed chemical labeling, novel label-free quantification strategies, improvement in PTM enrichment, more robust PTM analysis workflows and streamlined bioinformatics strategies will contribute to reliable identification and quantification of high numbers of PTMs [126,127]. The remaining challenge lies in defining the functional role of the physiologically relevant PTMs to understand PTM-dependent activity of MMPs in complex biological systems [128].
Funding
Ulrich auf dem Keller acknowledges support by a Novo Nordisk Foundation Young Investigator Award (NNF16OC0020670). Philipp Kastl is supported by a research fellowship from the German Research Foundation (project no. 415888450), and Fabio Sabino is recipient of a H.C. Ørsted COFUND Postdoc Fellowship (COFUNDfellowsDTU, grant agreement no. 713683).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| Asn | Asparagine amino acid |
| CA | Cysteine array |
| Cy | Cytosolic domain |
| ECM | Extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| ER | Endoplasmic reticulum |
| Fu | Furin-recognition site |
| GalNAc | N-acetyl galactosamine |
| GlcNAc | N-acetyl glucosamine |
| GPI | Glycosylphosphatidylinositol |
| LRP | Low-density lipoprotein receptor-related protein |
| MMP | Matrix metalloproteinase |
| Pro | Pro-domain |
| PTMs | Post-translational modifications |
| SA | Signal anchor |
| Ser | Serine amino acid |
| SH | Thiol group |
| SP | Signal peptide |
| Thr | Threonine amino acid |
| TIMPs | Tissue inhibitors of metalloproteinases |
| TM | Transmembrane domain |
| Tyr | Tyrosine amino acid |
| UDP | Uridine diphosphate |
References
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzyme Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Loffek, S.; Schilling, O.; Franzke, C.-W.; Claustrat, B.; Brun, J.; Chazot, G. Biological role of matrix metalloproteinases: a critical balance. Eur. Respir. J. 2011, 38, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I. Extracellular matrix remodelling: The role of matrix metalloproteinases. J. Pathol. 2003, 200, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Marchant, D.J.; Bellac, C.L.; Moraes, T.J.; Wadsworth, S.J.; Dufour, A.; Butler, G.S.; Bilawchuk, L.M.; Hendry, R.G.; Robertson, A.G.; Cheung, C.T.; et al. A new transcriptional role for matrix metalloproteinase-12 in antiviral immunity. Nat. Med. 2014, 20, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Cauwe, B.; Opdenakker, G. Intracellular substrate cleavage: A novel dimension in the biochemistry, biology and pathology of matrix metalloproteinases. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 351–423. [Google Scholar] [CrossRef]
- Tallant, C.; Marrero, A.; Gomis-Rüth, F.X. Matrix metalloproteinases: Fold and function of their catalytic domains. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 20–28. [Google Scholar] [CrossRef]
- Rodríguez, D.; Morrison, C.J.; Overall, C.M. Matrix metalloproteinases: What do they not do? New substrates and biological roles identified by murine models and proteomics. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Fanjul-Fernández, M.; Folgueras, A.R.; Cabrera, S.; López-Otín, C. Matrix metalloproteinases: Evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Murphy, G.; Troeberg, L. Extracellular regulation of metalloproteinases. Matrix Biol. 2015, 44–46, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Huwiler, A.; Akool, E.S.; Aschrafi, A.; Hamada, F.M.A.; Pfeilschifter, J.; Eberhardt, W. ATP Potentiates Interleukin-1??-induced MMP-9 Expression in Mesangial Cells via Recruitment of the ELAV Protein HuR. J. Biol. Chem. 2003, 278, 51758–51769. [Google Scholar] [CrossRef] [PubMed]
- Fähling, M.; Steege, A.; Perlewitz, A.; Nafz, B.; Mrowka, R.; Persson, P.B.; Thiele, B.J. Role of nucleolin in posttranscriptional control of MMP-9 expression. Biochim. Biophys. Acta Gene Struct. Expr. 2005, 1731, 32–40. [Google Scholar] [CrossRef]
- Clark, I.M.; Swingler, T.E.; Sampieri, C.L.; Edwards, D.R. The regulation of matrix metalloproteinases and their inhibitors. Int. J. Biochem. Cell Biol. 2008, 40, 1362–1378. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Zhang, Z.; Chen, W.; Huang, G.; He, A.; Hou, C.; Long, Y.; Yang, Z.; Zhang, Z.; Liao, W. MicroRNA-320 regulates matrix metalloproteinase-13 expression in chondrogenesis and interleukin-1β-induced chondrocyte responses. Osteoarthr. Cartil. 2016, 24, 932–941. [Google Scholar] [CrossRef]
- Bannikov, G.A.; Karelina, T.V.; Collier, I.E.; Marmer, B.L.; Goldberg, G.I. Substrate binding of gelatinase B induces its enzymatic activity in the presence of intact propeptide. J. Biol. Chem. 2002, 277, 16022–16027. [Google Scholar] [CrossRef]
- Emonard, H.; Hornebeck, W. Binding of 92 kDa and 72 kDa progelatinases to insoluble elastin modulates their proteolytic activation. Biol. Chem. 1997, 378, 265–271. [Google Scholar] [CrossRef]
- Ra, H.J.; Harju-Baker, S.; Zhang, F.; Linhardt, R.J.; Wilson, C.L.; Parks, W.C. Control of promatrilysin (MMP7) activation and substrate-specific activity by sulfated glycosaminoglycans. J. Biol. Chem. 2009, 284, 27924–27932. [Google Scholar] [CrossRef]
- Geurts, N.; Martens, E.; Van Aelst, I.; Proost, P.; Opdenakker, G.; Van Den Steen, P.E. β-hematin interaction with the hemopexin domain of gelatinase B/MMP-9 provokes autocatalytic processing of the propeptide, thereby priming activation by MMP-3. Biochemistry 2008, 47, 2689–2699. [Google Scholar] [CrossRef]
- Murphy, G.; Nagase, H. Progress in matrix metalloproteinase research. Mol. Aspects Med. 2009, 29, 290–308. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.H. Metalloproteinase inhibitors: biological actions and therapeutic opportunities. J. Cell Sci. 2002, 115, 3719–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, J.W.; Bennett, E.J. Proteome complexity and the forces that drive proteome imbalance. Nature 2016, 537, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.L.; Park, K.M.; Murray, J.; Kim, K.; Ryu, S.H. Direct Profiling the Post-Translational Modification Codes of a Single Protein Immobilized on a Surface Using Cu-free Click Chemistry. ACS Cent. Sci. 2018, 4, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Deribe, Y.L.; Pawson, T.; Dikic, I. Post-translational modifications in signal integration. Nat. Struct. Mol. Biol. 2010, 17, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Peterson, S.E.; Loring, J.F. Protein post-translational modifications and regulation of pluripotency in human stem cells. Cell Res. 2014, 24, 143–160. [Google Scholar] [CrossRef]
- Ricard-Blum, S. Protein–Glycosaminoglycan interaction networks: Focus on heparan sulfate. Perspect. Sci. 2017. [Google Scholar] [CrossRef]
- Audagnotto, M.; Dal Peraro, M. Protein post-translational modifications: In silico prediction tools and molecular modeling. Comput. Struct. Biotechnol. J. 2017, 15, 307–319. [Google Scholar] [CrossRef]
- Spoel, S.H. Orchestrating the proteome with post-translational modifications. J. Exp. Bot. 2018, 69, 4499–4503. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.D.; Overall, C.M. Proteolytic Post-translational Modification of Proteins: Proteomic Tools and Methodology. Mol. Cell. Proteomics 2013, 12, 3532–3542. [Google Scholar] [CrossRef] [Green Version]
- Beltrao, P.; Bork, P.; Krogan, N.J.; van Noort, V.; Abu-Qarn, M.; Eichler, J.; Sharon, N.; Alexander, J.; Lim, D.; Joughin, B.; et al. Evolution and functional cross-talk of protein post-translational modifications. Mol. Syst. Biol. 2013, 9, 714. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Lee, K.J. Post-translational modifications and their biological functions: proteomic analysis and systematic approaches. J. Biochem. Mol. Biol. 2004, 37, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Vanheule, V.; Metzemaekers, M.; Janssens, R.; Struyf, S.; Proost, P. How post-translational modifications influence the biological activity of chemokines. Cytokine 2018, 109, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Yang, F. Post-translational modification control of HBV biological processes. Front. Microbiol. 2018, 9, 2661. [Google Scholar] [CrossRef] [PubMed]
- Arbez, N.; Ratovitski, T.; Roby, E.; Chighladze, E.; Stewart, J.C.; Ren, M.; Wang, X.; Lavery, D.J.; Ross, C.A. Post-translational modifications clustering within proteolytic domains decrease mutant huntingtin toxicity. J. Biol. Chem. 2017, 292, 19238–19249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, B.J.; Nissim, A.; Winyard, P.G. Oxidative post-translational modifications and their involvement in the pathogenesis of autoimmune diseases. Redox Biol. 2014, 2, 715–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeken, J. Glycosylation and its Disorders: General Overview. Ref. Modul. Biomed. Sci. 2016, 1–8. [Google Scholar] [CrossRef]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef]
- Boon, L.; Ugarte-Berzal, E.; Vandooren, J.; Opdenakker, G. Glycosylation of matrix metalloproteases and tissue inhibitors: present state, challenges and opportunities. Biochem. J. 2016, 473, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, F.; Aebi, M. Mechanisms and principles of N-linked protein glycosylation. Curr. Opin. Struct. Biol. 2011, 21, 576–582. [Google Scholar] [CrossRef]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.B.G.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiro RG Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [CrossRef] [PubMed]
- Tian, E.; Ten Hagen, K.G. Recent insights into the biological roles of mucin-type O-glycosylation. Glycoconj. J. 2009, 26, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.J.; Clausen, H.; Bard, F. Location, location, location: New insights into O-GalNAc protein glycosylation. Trends Cell Biol. 2011, 21, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Guerrero, R. Recent structural and mechanistic insights into protein O-GalNAc glycosylation. Biochem. Soc. Trans. 2016, 44, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Helenius, A.; Aebi, M. Intracellular functions of N-linked glycans. Science 2001, 291, 2364–2369. [Google Scholar] [CrossRef]
- Lee, H.S.; Qi, Y.; Im, W. Effects of N-glycosylation on protein conformation and dynamics: Protein Data Bank analysis and molecular dynamics simulation study. Sci. Rep. 2015, 5, 8926. [Google Scholar] [CrossRef] [PubMed]
- Goettig, P. Effects of glycosylation on the enzymatic activity and mechanisms of proteases. Int. J. Mol. Sci. 2016, 17, 1969. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.; Fu, J.; Xia, L. Biological Functions of C1GalT1 and Mucin-Type O-Glycans. In Glycoscience: Biology and Medicine; Springer: Tokyo, Japan, 2015; pp. 1073–1080. ISBN 9784431548416. [Google Scholar]
- Tran, D.T.; Ten Hagen, K.G. Mucin-type o-glycosylation during development. J. Biol. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Hincapie, M.; Packer, N.; Baker, M.S.; Hancock, W.S.; Fanayan, S. An optimized approach for enrichment of glycoproteins from cell culture lysates using native multi-lectin affinity chromatography. J. Sep. Sci. 2012, 35, 2445–2452. [Google Scholar] [CrossRef] [PubMed]
- Vandooren, J.; Van Den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef] [PubMed]
- Kotra, L.P.; Zhang, L.; Fridman, R.; Orlando, R.; Mobashery, S. N-glycosylation pattern of the zymogenic form of human matrix metalloproteinase-9. Bioorg. Chem. 2002, 30, 356–370. [Google Scholar] [CrossRef]
- Roth, J. Protein N-glycosylation along the Secretory Pathway: Relationship to organelle topography and function, protein quality control, and cell interactions. Chem. Rev. 2002, 102, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Duellman, T.; Burnett, J.; Yang, J. Functional Roles of N-Linked Glycosylation of Human Matrix Metalloproteinase 9. Traffic 2015, 16, 1108–1126. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Cieplak, P. Role of N-glycosylation in activation of proMMP-9. A molecular dynamics simulations study. PLoS ONE 2018, 13, e0191157. [Google Scholar] [CrossRef] [PubMed]
- Nishi, N.; Shoji, H.; Seki, M.; Itoh, A.; Miyanaka, H.; Yuube, K.; Hirashima, M.; Nakamura, T. Galectin-8 modulates neutrophil function via interaction with integrin αM. Glycobiology 2003, 13, 755–763. [Google Scholar] [CrossRef]
- Boon, L.; Ugarte-Berzal, E.; Martens, E.; Vandooren, J.; Rybakin, V.; Colau, D.; Gordon-Alonso, M.; van der Bruggen, P.; Stöcker, W.; Becker-Pauly, C.; et al. Propeptide glycosylation and galectin-3 binding decrease proteolytic activation of human proMMP-9/progelatinase B. FEBS J. 2019, 285, 930–945. [Google Scholar] [CrossRef]
- Van Den Steen, P.E.; Van Aelst, I.; Hvidberg, V.; Piccard, H.; Fiten, P.; Jacobsen, C.; Moestrup, S.K.; Fry, S.; Royle, L.; Wormald, M.R.; et al. The hemopexin and O-glycosylated domains tune gelatinase B/MMP-9 bioavailability via inhibition and binding to cargo receptors. J. Biol. Chem. 2006, 281, 18626–18637. [Google Scholar] [CrossRef]
- Mattu, T.S.; Royle, L.; Langridge, J.; Wormald, M.R.; Van den Steen, P.E.; Van Damme, J.; Opdenakker, G.; Harvey, D.J.; Dwek, R.A.; Rudd, P.M. O-glycan analysis of natural human neutrophil gelatinase B using a combination of normal phase- HPLC and online tandem mass spectrometry: Implications for the domain organization of the enzyme. Biochemistry 2000, 39, 15695–15704. [Google Scholar] [CrossRef]
- Rosenblum, G.; Van den Steen, P.E.; Cohen, S.R.; Grossmann, J.G.; Frenkel, J.; Sertchook, R.; Slack, N.; Strange, R.W.; Opdenakker, G.; Sagi, I. Insights into the Structure and Domain Flexibility of Full-Length Pro-Matrix Metalloproteinase-9/Gelatinase B. Structure 2007, 15, 1227–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandooren, J.; Knoops, S.; Buzzo, J.L.A.; Boon, L.; Martens, E.; Opdenakker, G.; Kolaczkowska, E. Differential inhibition of activity, activation and gene expression of MMP-9 in THP-1 cells by azithromycin and minocycline versus bortezomib: A comparative study. PLoS ONE 2017, 12, e0174853. [Google Scholar] [CrossRef] [PubMed]
- Vandooren, J. Gelatin degradation assay reveals MMP-9 inhibitors and function of O-glycosylated domain. World J. Biol. Chem. 2011, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Zucker, S.; Drews, M.; Conner, C.; Foda, H.D.; DeClerck, Y.A.; Langley, K.E.; Bahou, W.F.; Docherty, A.J.P.; Cao, J. Tissue inhibitor of metalloproteinase-2 (TIMP-2) binds to the catalytic domain of the cell surface receptor, membrane type 1-matrix metalloproteinase 1 (MT1-MMP). J. Biol. Chem. 1998, 273, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Sato, H.; Okada, A.; Ohuchi, E.; Imai, K.; Okada, Y.; Seiki, M. TIMP-2 promotes activation of progelatinase A by membrane-type 1 matrix metalloproteinase immobilized on agarose beads. J. Biol. Chem. 1998, 273, 16098–16103. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.I.; Munshi, H.G.; Sen, R.; Snipas, S.J.; Salvesen, G.S.; Fridman, R.; Stack, M.S. Glycosylation Broadens the Substrate Profile of Membrane Type 1 Matrix Metalloproteinase. J. Biol. Chem. 2004, 279, 8278–8289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Growth, T.; Nguyen, A.T.; Chia, J.; Ros, M.; Hui, K.M.; Saltel, F.; Bard, F. Organelle Specific O-Glycosylation Drives MMP14 Activation, Tumor Growth, and Metastasis. Cancer Cell 2017, 32, 639–653.e6. [Google Scholar] [CrossRef] [Green Version]
- Saarinen, J.; Welgus, H.G.; Flizar, C.A.; Kalkkinen, N.; Helin, J. N-Glycan structures of matrix metalloproteinase-1 derived from human fibroblasts and from HT-1080 fibrosarcoma cells. Eur. J. Biochem. 1999, 259, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Piccard, H.; Van den Steen, P.E.; Opdenakker, G. Hemopexin domains as multifunctional liganding modules in matrix metalloproteinases and other proteins. J. Leukoc. Biol. 2007, 81, 870–892. [Google Scholar] [CrossRef]
- Dufour, A.; Sampson, N.S.; Zucker, S.; Cao, J. Role of the hemopexin domain of matrix metalloproteinases in cell migration. J. Cell. Physiol. 2008, 217, 643–651. [Google Scholar] [CrossRef] [Green Version]
- García-Pardo, A.; Opdenakker, G. Nonproteolytic functions of matrix metalloproteinases in pathology and insights for the development of novel therapeutic inhibitors. Met. Med. 2015, 2, 19–28. [Google Scholar] [CrossRef]
- Nagase, H. Matrix Metalloproteinase 3/Stromelysin 1. In Handbook of Proteolytic Enzymes, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 1, pp. 763–774. [Google Scholar] [CrossRef]
- Henriet, P.; Eeckhout, Y. Eeckhout, Y. Matrix Metallopeptidase-13/Collagenase 3. In Handbook of Proteolytic Enzymes, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 1, pp. 734–744. [Google Scholar] [CrossRef]
- Knäuper, V.; López-Otin, C.; Smith, B.; Knight, G.; Murphy, G. Biochemical characterization of human collagenase-3. J. Biol. Chem. 1996, 271, 1544–1550. [Google Scholar] [CrossRef] [PubMed]
- Hieronimus, B.; Pfohl, J.; Busch, C.; Graeve, L. Expression and characterization of membrane-type 4 matrix metalloproteinase (MT4-MMP) and its different forms in melanoma. Cell. Physiol. Biochem. 2017, 42, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Kajita, M.; Kinoh, H.; Mori, H.; Okada, A.; Seiki, M. Membrane type 4 matrix metalloproteinase (MT4-MMP, MMP-17) is a glycosylphosphatidylinositol-anchored proteinase. J. Biol. Chem. 1999, 274, 34260–34266. [Google Scholar] [CrossRef] [PubMed]
- Sohail, A.; Marco, M.; Zhao, H.; Shi, Q.; Merriman, S.; Mobashery, S.; Fridman, R. Characterization of the dimerization interface of membrane type 4 (MT4)-matrix metalloproteinase. J. Biol. Chem. 2011, 286, 33178–33189. [Google Scholar] [CrossRef] [PubMed]
- Ardito, F.; Giuliani, M.; Perrone, D.; Troiano, G.; Muzio, L. Lo The crucial role of protein phosphorylation in cell signalingand its use as targeted therapy (Review). Int. J. Mol. Med. 2017, 40, 271–280. [Google Scholar] [CrossRef]
- Nishi, H.; Shaytan, A.; Panchenko, A.R. Physicochemical mechanisms of protein regulation by phosphorylation. Front. Genet. 2014, 5, 270. [Google Scholar] [CrossRef] [Green Version]
- Cutillas, P.R. Targeted In-Depth Quantification of Signaling Using Label-Free Mass Spectrometry. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 585, pp. 245–268. ISBN 9780128097427. [Google Scholar]
- Cohen, P. The role of protein phosphorylation in human health and disease. Eur. J. Biochem. 2001, 268, 5001–5010. [Google Scholar] [CrossRef]
- Sariahmetoglu, M.; Crawford, B.D.; Leon, H.; Sawicka, J.; Li, L.; Ballermann, B.J.; Holmes, C.; Berthiaume, L.G.; Holt, A.; Sawicki, G.; et al. Regulation of matrix metalloproteinase-2 (MMP-2) activity by phosphorylation. FASEB J. 2007, 21, 2486–2495. [Google Scholar] [CrossRef]
- Jacob-Ferreira, A.L.; Kondo, M.Y.; Baral, P.K.; James, M.N.G.; Holt, A.; Fan, X.; Schulz, R. Phosphorylation Status of 72 kDa MMP-2 Determines Its Structure and Activity in Response to Peroxynitrite. PLoS ONE 2013, 8, e71794. [Google Scholar] [CrossRef]
- 3rd Millenium, PhosphoSitePlus. Cell Signalling Technology Inc. 2011. Available online: http://www.phosphosite.org/proteinAction.do?id=662&showAllSites=true (accessed on 22 May 2019).
- UniProt Consortium, T. UniProt: the universal protein knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.C.; Coppolino, M.G. Phosphorylation of membrane type 1-matrix metalloproteinase (MT1-MMP) and its vesicle-associated membrane protein 7 (VAMP7)-dependent trafficking facilitate cell invasion and migration. J. Biol. Chem. 2011, 286, 43405–43416. [Google Scholar] [CrossRef] [PubMed]
- Moss, N.M.; Wu, Y.I.; Liu, Y.; Munshi, H.G.; Stack, M.S. Modulation of the membrane type 1 matrix metalloproteinase cytoplasmic tail enhances tumor cell invasion and proliferation in three-dimensional collagen matrices. J. Biol. Chem. 2009, 284, 19791–19799. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kasberg, W.C.; Celo, A.; Liang, Z.; Quispe, K.; Sharon Stack, M. Post-translational modification of the membrane type 1 matrix metalloproteinase (MT1-MMP) cytoplasmic tail impacts ovarian cancer multicellular aggregate dynamics. J. Biol. Chem. 2017, 292, 13111–13121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyalendo, C.; Michaud, M.; Beaulieu, E.; Roghi, C.; Murphy, G.; Gingras, D.; Béliveau, R. Src-dependent Phosphorylation of Membrane Type I Matrix Metalloproteinase on Cytoplasmic Tyrosine 573. J. Biol. Chem. 2007, 282, 15690–15699. [Google Scholar] [CrossRef] [Green Version]
- Moss, N.M.; Liu, Y.; Johnson, J.J.; Debiase, P.; Jones, J.; Hudson, L.G.; Munshi, H.G.; Stack, M.S. Epidermal Growth Factor Receptor-Mediated Membrane Type 1 Matrix Metalloproteinase Endocytosis Regulates the Transition between Invasive versus Expansive Growth of Ovarian Carcinoma Cells in Three-Dimensional Collagen. Mol. Cancer Res. 2009, 7, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Rinschen, M.M.; Yu, M.-J.; Wang, G.; Boja, E.S.; Hoffert, J.D.; Pisitkun, T.; Knepper, M.A. Quantitative phosphoproteomic analysis reveals vasopressin V2-receptor–dependent signaling pathways in renal collecting duct cells. Proc. Natl. Acad. Sci. USA 2010, 107, 3882–3887. [Google Scholar] [CrossRef]
- Bordoli, M.R.; Yum, J.; Breitkopf, S.B.; Thon, J.N.; Italiano, J.E.; Xiao, J.; Worby, C.; Wong, S.K.; Lin, G.; Edenius, M.; et al. A secreted tyrosine kinase acts in the extracellular environment. Cell 2014, 158, 1033–1044. [Google Scholar] [CrossRef]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Sreelatha, A.; Kinch, L.N.; Tagliabracci, V.S. The secretory pathway kinases. Biochim. Biophys. Acta Proteins Proteomics 2015, 1854, 1687–1693. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.O.; Takeuchi, H.; Haltiwanger, R.S.; Irvine, K.D. Four-jointed is a Golgi kinase that phosphorylates a subset of cadherin domains. Science 2008, 321, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Lin, X. Functions of heparan sulfate proteoglycans in cell signaling during development. Development 2004, 131, 6009–6021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proudfoot, E.A.; Johnson, Z.; Bonvin, P.; Handel, M.T. Glycosaminoglycan Interactions with Chemokines Add Complexity to a Complex System. Pharmaceuticals 2017, 10, 70. [Google Scholar] [CrossRef]
- Raman, R.; Sasisekharan, V.; Sasisekharan, R. Structural Insights into Biological Roles of Protein-Glycosaminoglycan Interactions. Chem. Biol. 2005, 12, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Esko, J.D. Demystifying Heparan Sulfate–Protein Interactions. Annu. Rev. Biochem. 2014, 83, 129–157. [Google Scholar] [CrossRef]
- Lin, X.; Buff, E.M.; Perrimon, N.; Michelson, A.M. Heparan sulfate proteoglycans are essential for FGF receptor signaling during Drosophila embryonic development. Development 1999, 126, 3715–3723. [Google Scholar] [PubMed]
- Guimond, S.; Maccarana, M.; Olwin, B.B.; Lindahl, U.; Rapraeger, A.C. Activating and inhibitory heparin sequences for FGF-2 (basic FGF). Distinct requirements for FGF-1, FGF-2, and FGF-4. J. Biol. Chem. 1993, 268, 23906–23914. [Google Scholar]
- Pye, D.A.; Vives, R.R.; Turnbull, J.E.; Hyde, P.; Gallagher, J.T. Heparan Sulfate Oligosaccharides Require 6-O-Sulfation for Promotion of Basic Fibroblast Growth Factor Mitogenic Activity. J. Biol. Chem. 1998, 273, 22936–22942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapraeger, A.C. In the clutches of proteoglycans: how does heparan sulfate regulate FGF binding? Chem. Biol. 1995, 2, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Lee, S.; Wang, Y.; Ha, Y. Crystal Structure of the E2 Domain of Amyloid Precursor Protein-like Protein 1 in Complex with Sucrose Octasulfate. J. Biol. Chem. 2011, 286, 29748–29757. [Google Scholar] [CrossRef] [Green Version]
- Gralle, M.; Botelho, M.G.; Wouters, F.S. Neuroprotective Secreted Amyloid Precursor Protein Acts by Disrupting Amyloid Precursor Protein Dimers. J. Biol. Chem. 2009, 284, 15016–15025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogewerf, A.J.; Kuschert, G.S.V.; Proudfoot, A.E.I.; Borlat, F.; Clark-Lewis, I.; Power, C.A.; Wells, T.N.C. Glycosaminoglycans Mediate Cell Surface Oligomerization of Chemokines. Biochemistry 1997, 36, 13570–13578. [Google Scholar] [CrossRef] [PubMed]
- Salanga, C.L.; Handel, T.M. Chemokine oligomerization and interactions with receptors and glycosaminoglycans: The role of structural dynamics in function. Exp. Cell Res. 2011, 317, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Gospodarowicz, D.; Cheng, J. Heparin protects basic and acidic FGF from inactivation. J. Cell. Physiol. 1986, 128, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Saksela, O.; Moscatelli, D.; Sommer, A.; Rifkin, D.B. Endothelial cell-derived heparan sulfate binds basic fibroblast growth factor and protects it from proteolytic degradation. J. Cell Biol. 1988, 107, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Sadir, R.; Imberty, A.; Baleux, F.; Lortat-Jacob, H. Heparan Sulfate/Heparin Oligosaccharides Protect Stromal Cell-derived Factor-1 (SDF-1)/CXCL12 against Proteolysis Induced by CD26/Dipeptidyl Peptidase IV. J. Biol. Chem. 2004, 279, 43854–43860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lortat-Jacob, H.; Baltzer, F.; Grimaud, J.-A. Heparin Decreases the Blood Clearance of Interferon-$γ$ and Increases Its Activity by Limiting the Processing of Its Carboxyl-terminal Sequence. J. Biol. Chem. 1996, 271, 16139–16143. [Google Scholar] [CrossRef]
- Jakobs, P.; Schulz, P.; Ortmann, C.; Schürmann, S.; Exner, S.; Rebollido-Rios, R.; Dreier, R.; Seidler, D.G.; Grobe, K. Bridging the gap: heparan sulfate and Scube2 assemble Sonic hedgehog release complexes at the surface of producing cells. Sci. Rep. 2016, 6, 26435. [Google Scholar] [CrossRef]
- Jakobs, P.; Schulz, P.; Schürmann, S.; Niland, S.; Exner, S.; Rebollido-Rios, R.; Manikowski, D.; Hoffmann, D.; Seidler, D.G.; Grobe, K. Ca2+ coordination controls sonic hedgehog structure and its Scube2-regulated release. J. Cell Sci. 2017, 130, 3261–3271. [Google Scholar] [CrossRef]
- Kastl, P.; Manikowski, D.; Steffes, G.; Schürmann, S.; Bandari, S.; Klämbt, C.; Grobe, K. Disrupting Hedgehog Cardin-Weintraub sequence and positioning changes cellular differentiation and compartmentalization in vivo. Development 2018, 145, dev167221. [Google Scholar] [CrossRef]
- Cerqueira, C.; Samperio Ventayol, P.; Vogeley, C.; Schelhaas, M. Kallikrein-8 Proteolytically Processes Human Papillomaviruses in the Extracellular Space To Facilitate Entry into Host Cells. J. Virol. 2015, 89, 7038–7052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Naruo, H.; Yoshitomi, Y.; Munesue, S.; Kiyono, S.; Kusano, Y.; Hashimoto, K.; Yokoi, T.; Nakanishi, H.; Shimizu, S.; et al. Matrix Metalloproteinase-9 Associated with Heparan Sulphate Chains of GPI-Anchored Cell Surface Proteoglycans Mediates Motility of Murine Colon Adenocarcinoma Cells. J. Biochem. 2008, 143, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.-H.; Woessner, J.F. Heparan Sulfate Proteoglycans as Extracellular Docking Molecules for Matrilysin (Matrix Metalloproteinase 7). J. Biol. Chem. 2000, 275, 4183–4191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, G.S.; Apte, S.S.; Willenbrock, F.; Murphy, G. Human Tissue Inhibitor of Metalloproteinases 3 Interacts with Both the N- and C-terminal Domains of Gelatinases A and B: REGULATION BY POLYANIONS. J. Biol. Chem. 1999, 274, 10846–10851. [Google Scholar] [CrossRef] [PubMed]
- Mannello, F.; Jung, K.; Tonti, G.A.; Canestrari, F. Heparin affects matrix metalloproteinases and tissue inhibitors of metalloproteinases circulating in peripheral blood. Clin. Biochem. 2008, 41, 1466–1473. [Google Scholar] [CrossRef] [PubMed]
- Di Simone, N.; Di Nicuolo, F.; Sanguinetti, M.; Ferrazzani, S.; D’Alessio, M.C.; Castellani, R.; Bompiani, A.; Caruso, A. Low-molecular Weight Heparin Induces In Vitro Trophoblast Invasiveness: Role of Matrix Metalloproteinases and Tissue Inhibitors. Placenta 2007, 28, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Isnard, N.; Robert, L.; Renard, G. Effect of Sulfated GAGs on the Expression and Activation of MMP-2 and MMP-9 in Corneal and Dermal Explant Cultures. Cell Biol. Int. 2003, 27, 779–784. [Google Scholar] [CrossRef]
- Rababah, M.; Worthmann, H.; Deb, M.; Tryc, A.B.; Ma, Y.T.; El Bendary, O.M.; Hecker, H.; Goldbecker, A.; Heeren, M.; Brand, K.; et al. Anticoagulants affect matrix metalloproteinase 9 levels in blood samples of stroke patients and healthy controls. Clin. Biochem. 2012, 45, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, E.; Pejler, G.; Ringvall, M.; Lunderius, C.; Tomasini-Johansson, B.; Kusche-Gullberg, M.; Eriksson, I.; Ledin, J.; Hellman, L.; Kjellén, L. Abnormal mast cells in mice deficient in a heparin-synthesizing enzyme. Nature 1999, 400, 773. [Google Scholar] [CrossRef] [PubMed]
- Humphries, D.E.; Wong, G.W.; Friend, D.S.; Gurish, M.F.; Qiu, W.-T.; Huang, C.; Sharpe, A.H.; Stevens, R.L. Heparin is essential for the storage of specific granule proteases in mast cells. Nature 1999, 400, 769. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Mann, M. Status of Large-scale Analysis of Post-translational Modifications by Mass Spectrometry. Mol. Cell. Proteomics 2013, 12, 3444–3452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryšlavá, H.; Doubnerová, V.; Kavan, D.; Vaněk, O. Effect of posttranslational modifications on enzyme function and assembly. J. Proteomics 2013, 92, 80–109. [Google Scholar] [CrossRef] [PubMed]
- Pascovici, D.; Wu, J.X.; McKay, M.J.; Joseph, C.; Noor, Z.; Kamath, K.; Wu, Y.; Ranganathan, S.; Gupta, V.; Mirzaei, M. Clinically Relevant Post-Translational Modification Analyses-Maturing Workflows and Bioinformatics Tools. Int. J. Mol. Sci. 2018, 20, 16. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Eisen, A.Z.; Teter, M.; Clark, S.D.; Kronberger, A.; Goldberg, G. Human fibroblast collagenase: glycosylation and tissue-specific levels of enzyme synthesis. Proc. Natl. Acad. Sci. USA 2006, 83, 3756–3760. [Google Scholar] [CrossRef] [PubMed]
- Huanna, T.; Tao, Z.; Xiangfei, W.; Longfei, A.; Yuanyuan, X.; Jianhua, W.; Cuifang, Z.; Manjing, J.; Wenjing, C.; Shaochuan, Q.; et al. GALNT14 mediates tumor invasion and migration in breast cancer cell MCF-7. Mol. Carcinog. 2015, 54, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
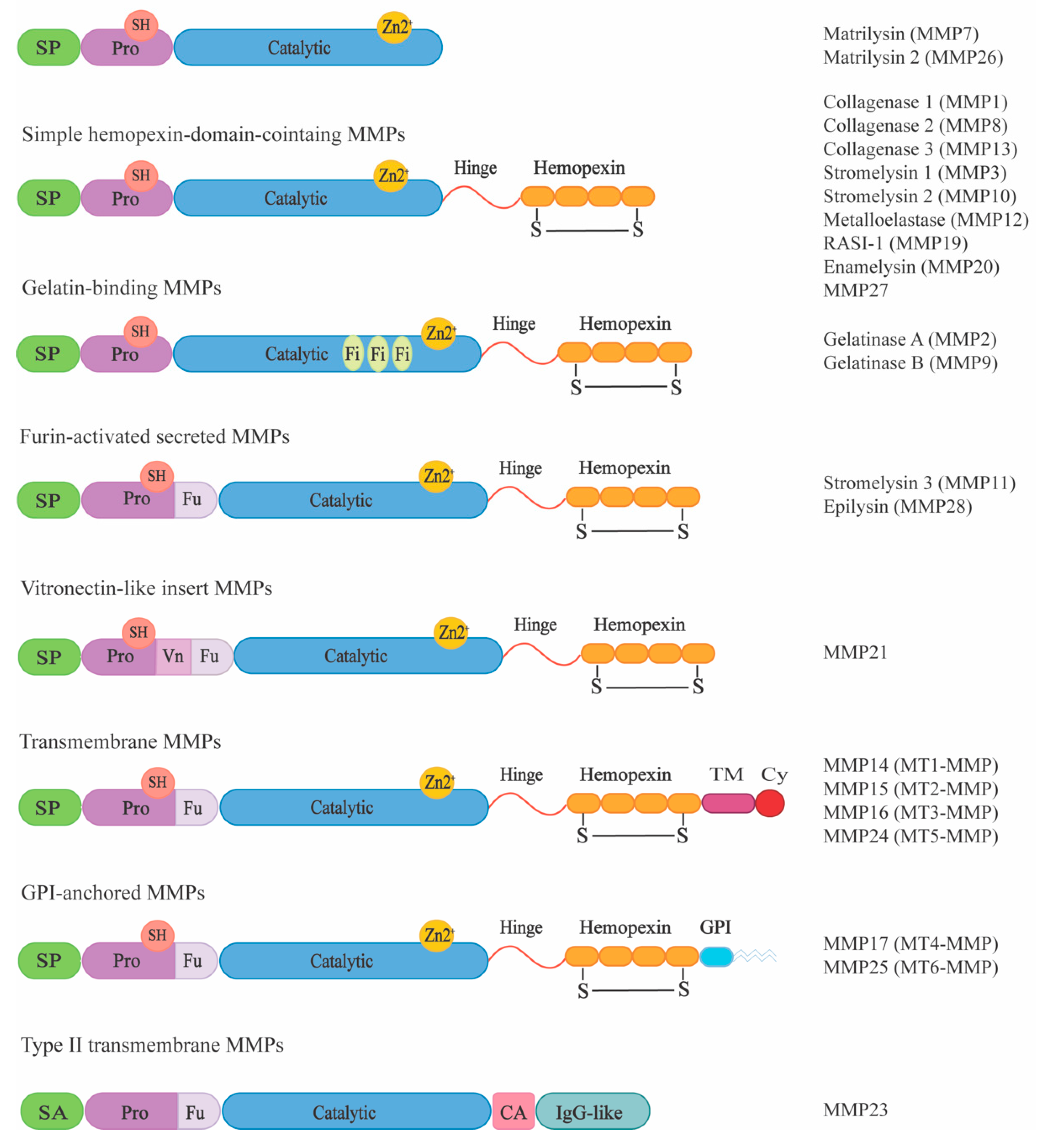
Figure 1.
MMP multidomain organization. Based on their domain structure, MMPs can be subdivided into eight different groups. All MMPs share a common structure comprising a signal peptide (SP), a pro-domain (Pro), containing a thiol group (SH), a catalytic domain (Catalytic) with a zinc (Zn) binding site, a linker region (Hinge) and a hemopexin domain (Hemopexin), which has a disulfide bond (S-S). Exceptions to this are the two gelatinases, which contain three fibronectin repeats (Fi) within their catalytic domain and furin-activated MMPs which have a furin-recognition site (Fu) within their pro-domain and MMP21 with an additional vitronectin-like insert (Vn). Some membrane-type MMPs are anchored to the membrane via glycosylphosphatidylinositol (GPI), whereas some MT-MMPs have transmembrane (TM) and cytosolic domains (Cy). In type II MT-MMPs, an N-terminal signal anchor (SA), a cysteine array (CA) domain, and an immunoglobulin-like (Ig-like) domain are present. Adapted by permission from “Springer Nature: New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. Egeblad & Werb, Copyright (2002)”.
Figure 1.
MMP multidomain organization. Based on their domain structure, MMPs can be subdivided into eight different groups. All MMPs share a common structure comprising a signal peptide (SP), a pro-domain (Pro), containing a thiol group (SH), a catalytic domain (Catalytic) with a zinc (Zn) binding site, a linker region (Hinge) and a hemopexin domain (Hemopexin), which has a disulfide bond (S-S). Exceptions to this are the two gelatinases, which contain three fibronectin repeats (Fi) within their catalytic domain and furin-activated MMPs which have a furin-recognition site (Fu) within their pro-domain and MMP21 with an additional vitronectin-like insert (Vn). Some membrane-type MMPs are anchored to the membrane via glycosylphosphatidylinositol (GPI), whereas some MT-MMPs have transmembrane (TM) and cytosolic domains (Cy). In type II MT-MMPs, an N-terminal signal anchor (SA), a cysteine array (CA) domain, and an immunoglobulin-like (Ig-like) domain are present. Adapted by permission from “Springer Nature: New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. Egeblad & Werb, Copyright (2002)”.

Figure 2.
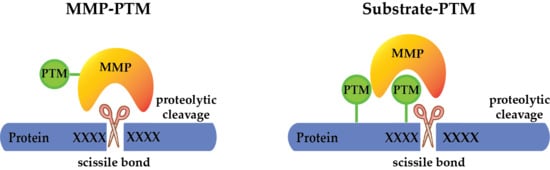
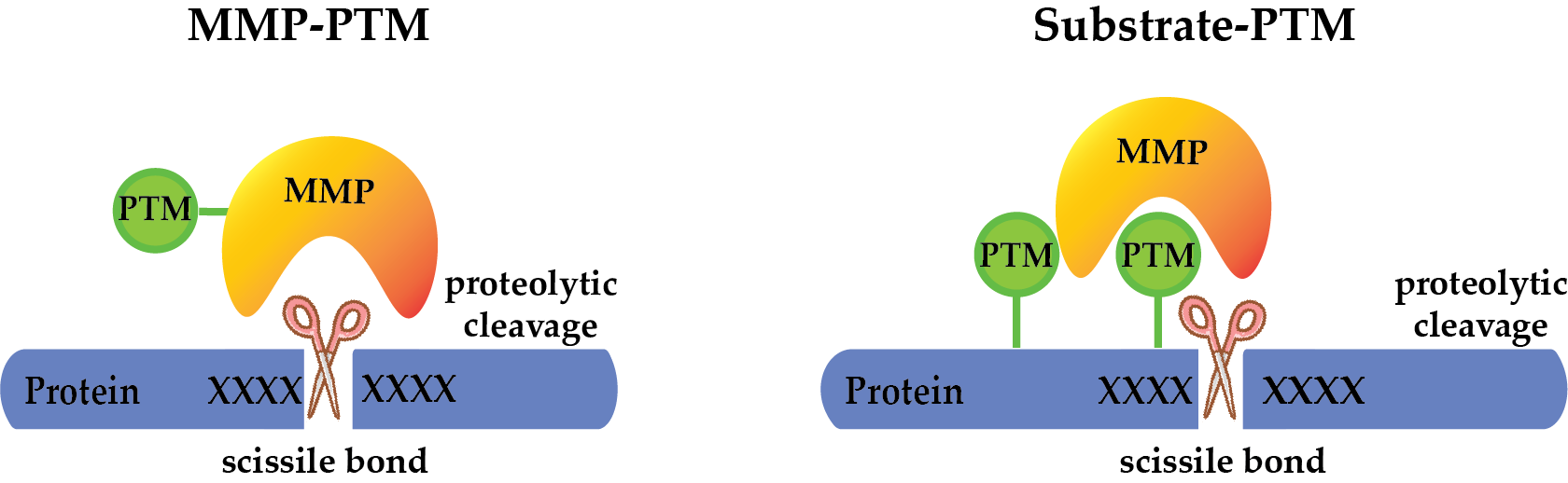
Possibilities of PTM crosstalk with MMPs. PTMs can regulate the activity of MMPs on the protease level by (a) activating the protease to allow cleavage of the target protein, (b) inactivating the protease, or (c) modulating its substrate specificity. Additionally, PTMs may regulate substrate cleavage by MMPs, through (d) directly modifying the cleavage motif (catalytic interaction), or substrate domains outside the cleavage site (non-catalytic interaction), guiding MMPs to cleave the same (cis) (e) or another (trans) protein (f).
Figure 2.
Possibilities of PTM crosstalk with MMPs. PTMs can regulate the activity of MMPs on the protease level by (a) activating the protease to allow cleavage of the target protein, (b) inactivating the protease, or (c) modulating its substrate specificity. Additionally, PTMs may regulate substrate cleavage by MMPs, through (d) directly modifying the cleavage motif (catalytic interaction), or substrate domains outside the cleavage site (non-catalytic interaction), guiding MMPs to cleave the same (cis) (e) or another (trans) protein (f).
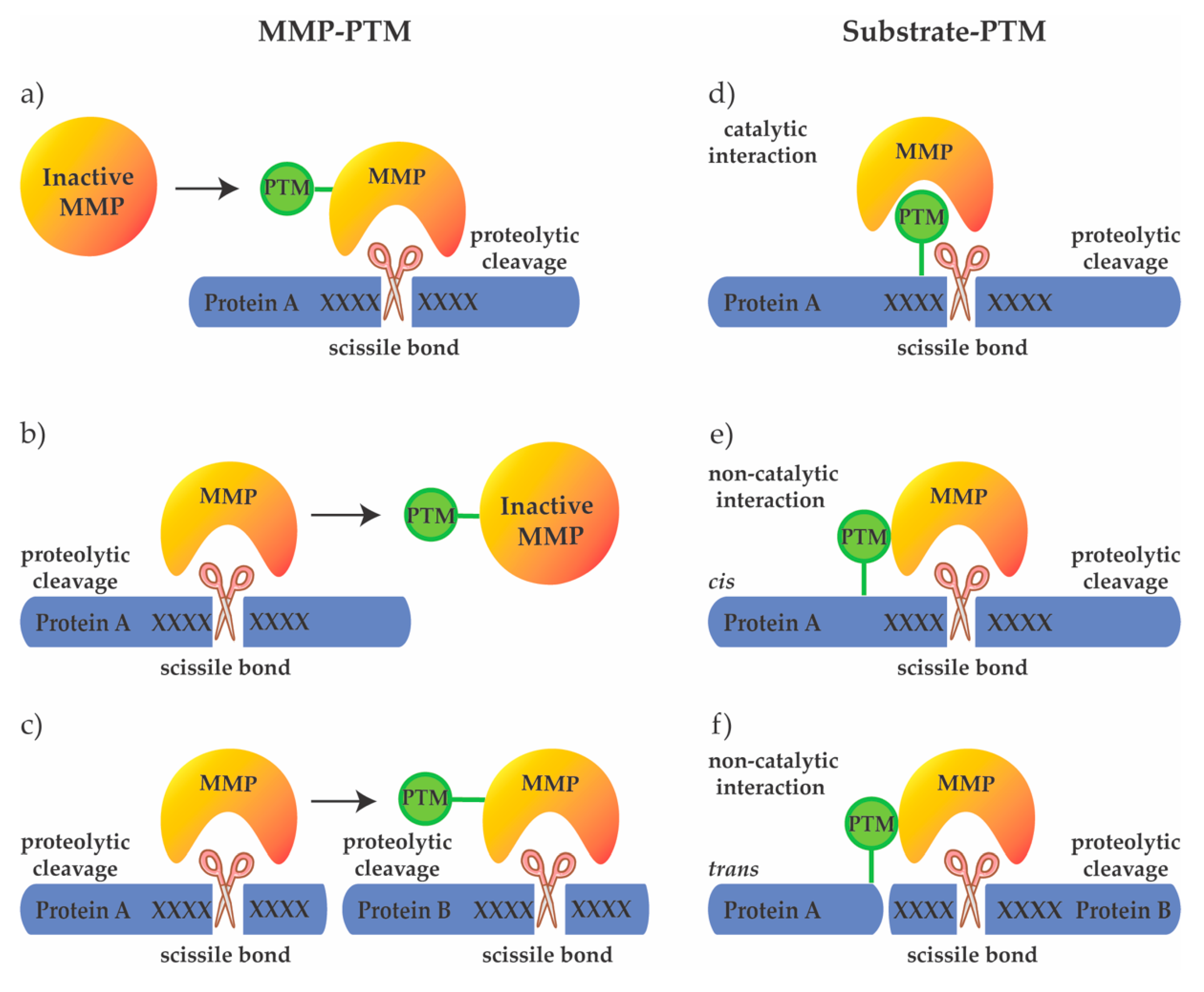
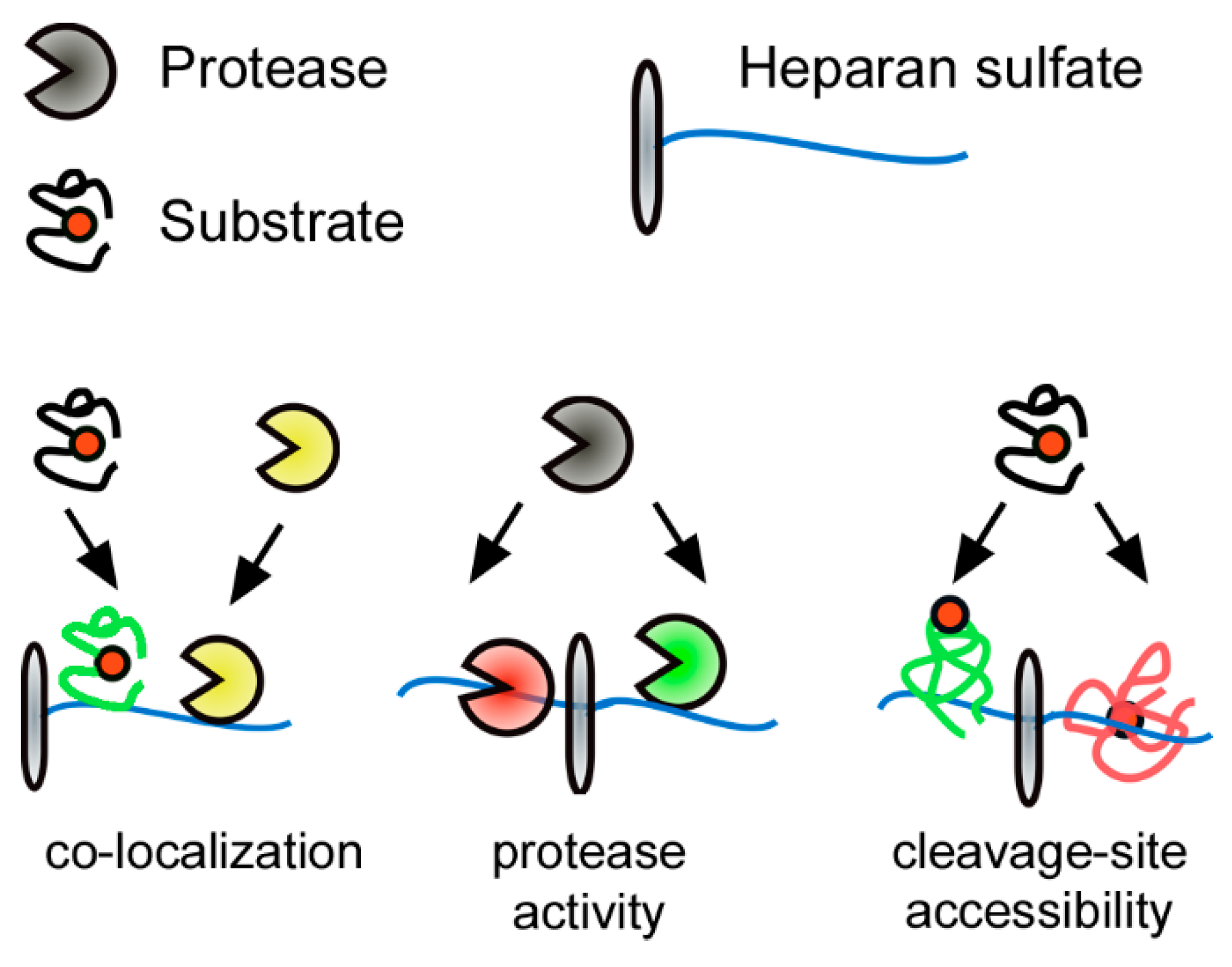
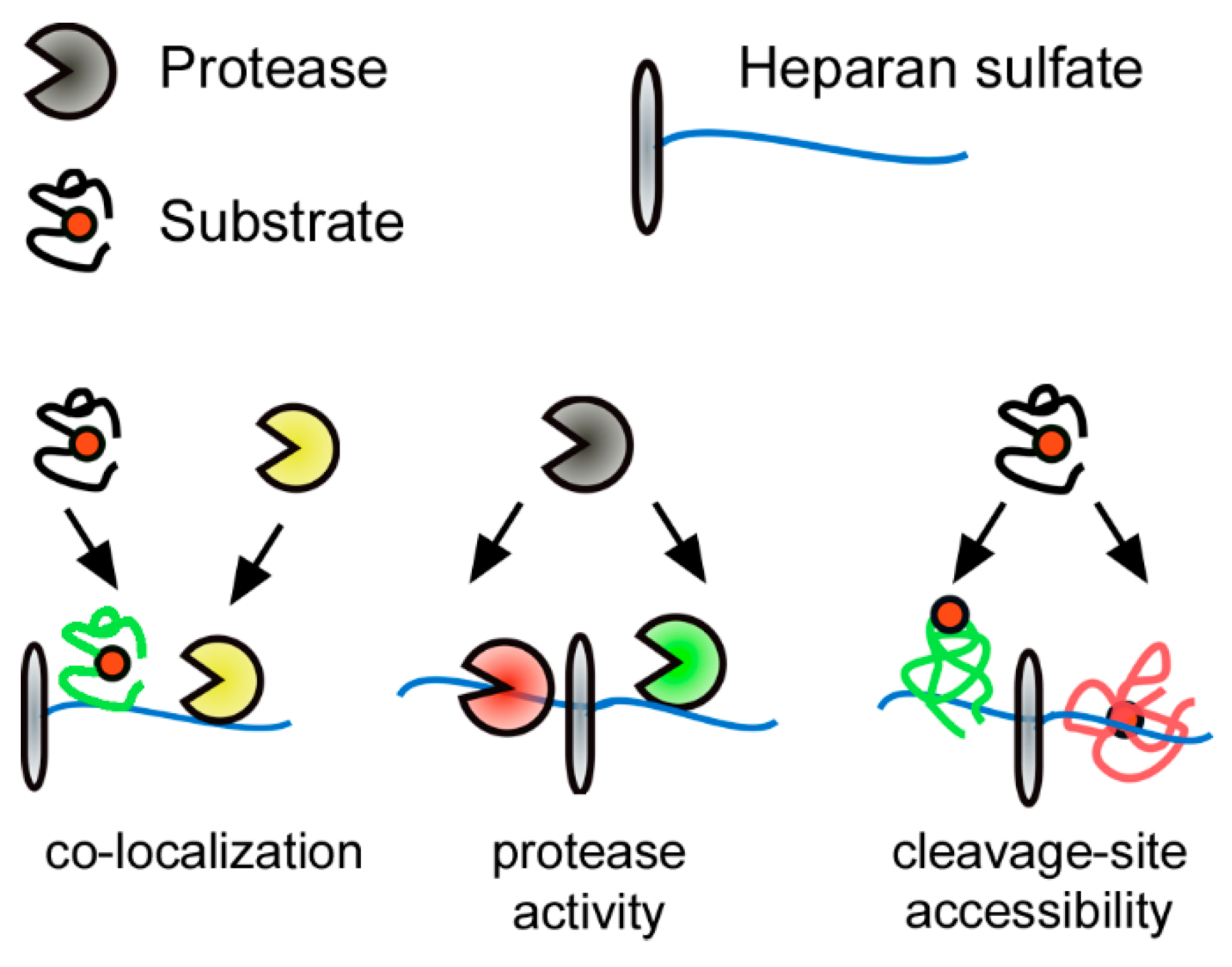
Figure 3.
Heparan sulfate (HS)-protein interaction as a post translational regulator of extracellular proteolysis. Electrostatic interactions of proteins with negatively charged HS affect extracellular proteolysis at three different levels: (1) by mediating co-localization of reactants; (2) on the protease level by affecting protease activity/accessibility; or (3) on the substrate level by modulating accessibility to cleavage sites. The substrate cleavage site is indicated by a red circle, green substrate/protease color denotes activity; red substrate/protease color denotes inactivity/inaccessibility.
Figure 3.
Heparan sulfate (HS)-protein interaction as a post translational regulator of extracellular proteolysis. Electrostatic interactions of proteins with negatively charged HS affect extracellular proteolysis at three different levels: (1) by mediating co-localization of reactants; (2) on the protease level by affecting protease activity/accessibility; or (3) on the substrate level by modulating accessibility to cleavage sites. The substrate cleavage site is indicated by a red circle, green substrate/protease color denotes activity; red substrate/protease color denotes inactivity/inaccessibility.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of identified PTMs in MMPs.
| MMP | Modification | Biological Effect | Reference |
|---|---|---|---|
| MMP1 | N-linked glycosylation at Asn120 | Tumor cell invasion and angiogenesis | [39,69,129] |
| Phosphorylation at Tyr360 | Not reported | [93] | |
| MMP2 | O-linked glycosylation at Ser32, Thr96, 262, 458, 460 | Upregulation of MMP2 | [39,71,72] |
| N-linked glycosylation at Asn573 and Asn642 | Not reported | [130] | |
| Phosphorylation at Ser32, Ser160, Tyr271, Thr250 and Ser365 | Phosphorylation decreases, while dephosphorylation increases protease activity | [83,84] | |
| Heparan sulfate | Cell surface localization; affects protease activity by increasing TIMP3 affinity | [119,120,121,122] | |
| MMP3 | N-linked glycosylation at Asn120 and Asn398 | Not reported | [39,73] |
| Three potential O-linked glycosylation at Ser56, Ser269 and Thr277 | |||
| MMP7 | Heparan sulfate, Chondroitin sulfate | Cell surface localization; affects protease activity by increasing TIMP3 affinity; increases MMP7 auto-processing and activity | [19,120,121] |
| MMP9 | N-linked glycosylation at Asn38 and Asn120 | MMP9 secretion and activation | [53,54,55,56,57,58,59] |
| O-linked glycosylation in the linker region | Increases the domain flexibility; necessary for internalization and degradation; protects against proteolytic degradation; reduces gelatinolytic activity | [60,62,63,64] | |
| Heparan sulfate | Cell surface localization; affects protease activity by increasing TIMP3 affinity; affects MMP9 expression and plasma levels | [119,120,121,122,123] | |
| MMP12 | Phosphorylation at Tyr414 | Not reported | [93] |
| MMP13 | N-linked glycosylation at Asn117 and Asn152 | Not reported | [1,74,75] |
| O-linked glycosylation at Ser24 and Ser62 | Not reported | [1,74,75] | |
| Phosphorylation at Tyr366 | Not reported | [93] | |
| MMP14 | N-linked glycosylation at Asn229 and Asn311 | Not reported | [39] |
| O-linked glycosylation at Thr291, Thr299, Thr300, and Ser301 | Required for formation of a stable complex with proMMP2 and TIMP2; increases activity upon glycosylation perturbation | [65,66,67,68] | |
| Phosphorylation at Thr567, Tyr573 and Tyr353 | Regulates MMP14 induced cellular invasion and migration; cell surface dynamics and internalization; mimetic mutants exhibit higher collagenolytic activity and three-dimensional growth; promotes metastasis-associated behaviors | [72,87,88,89,90,91,93] | |
| MMP16 | Phosphorylation at Tyr377 and Tyr521 | Not reported | [93] |
| MMP17 | N-linked glycosylation at Asn137 and Asn318 | Stabilizes the dimeric form of MMP17 | [76,77,78] |
| MMP24 | Phosphorylation at Tyr534 | Not reported | [93] |
| MMP27 | Phosphorylation at Tyr360 | Not reported | [93] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Madzharova, E.; Kastl, P.; Sabino, F.; auf dem Keller, U. Post-Translational Modification-Dependent Activity of Matrix Metalloproteinases. Int. J. Mol. Sci. 2019, 20, 3077. https://doi.org/10.3390/ijms20123077
AMA Style
Madzharova E, Kastl P, Sabino F, auf dem Keller U. Post-Translational Modification-Dependent Activity of Matrix Metalloproteinases. International Journal of Molecular Sciences. 2019; 20(12):3077. https://doi.org/10.3390/ijms20123077
Chicago/Turabian StyleMadzharova, Elizabeta, Philipp Kastl, Fabio Sabino, and Ulrich auf dem Keller. 2019. "Post-Translational Modification-Dependent Activity of Matrix Metalloproteinases" International Journal of Molecular Sciences 20, no. 12: 3077. https://doi.org/10.3390/ijms20123077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.