Suppression of STAT3 Phosphorylation and RelA/p65 Acetylation Mediated by MicroRNA134 Plays a Pivotal Role in the Apoptotic Effect of Lambertianic Acid


 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
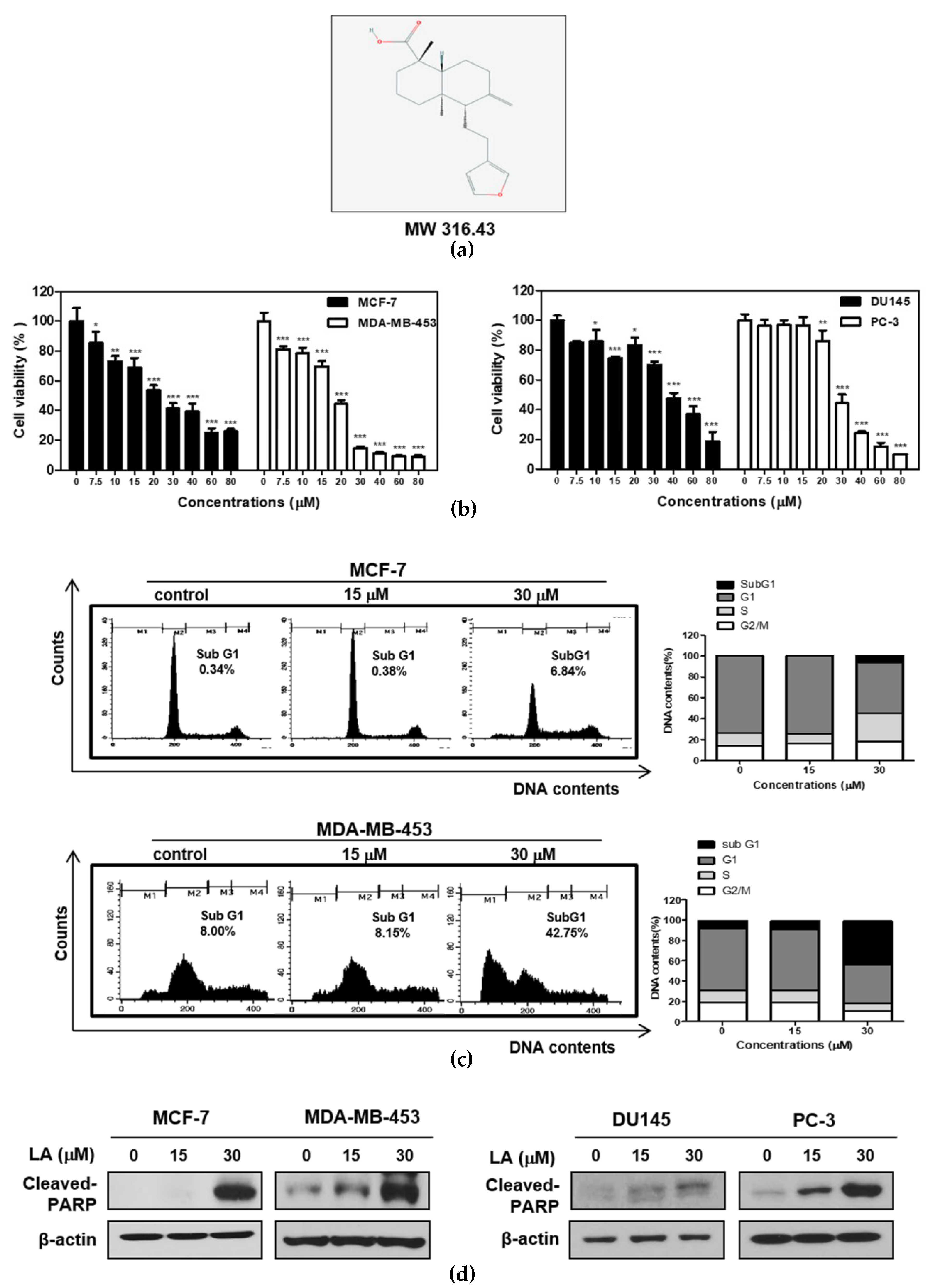
2.1. LA Induced Cytotoxicity and Sub-G1 Accumulation Increased the Cleavage of Poly (ADP-Ribose) Polymerase (PARP) in STAT3-Dependent or STAT3-Independent Cancer Cells
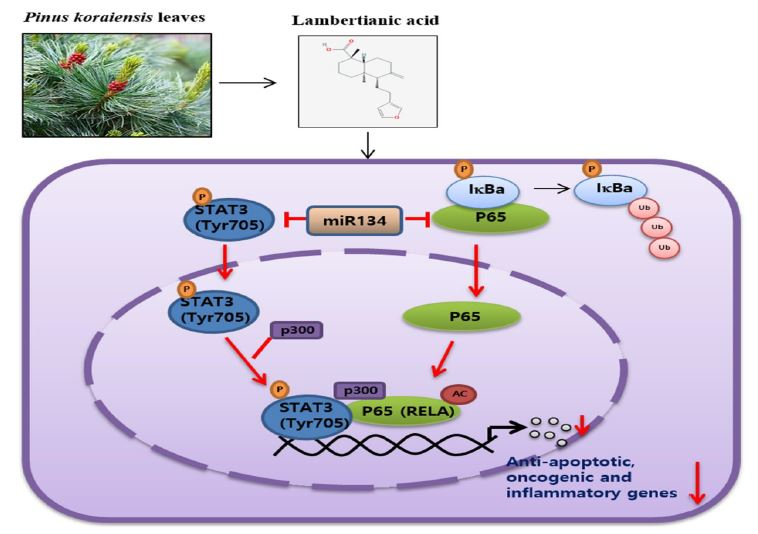
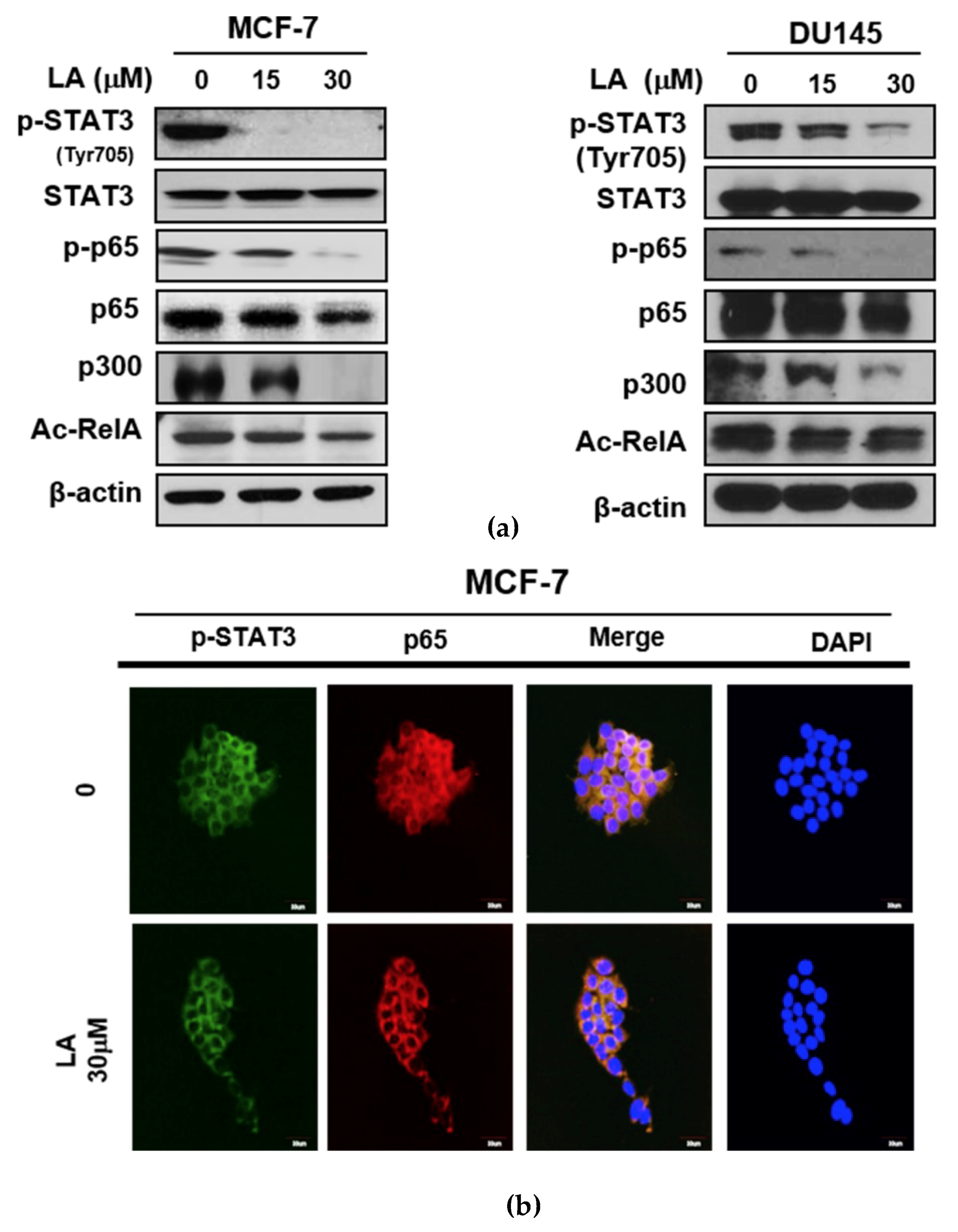
2.2. LA Suppressed the Phosphorylation of STAT3 and NF-κB, and the Expression of p300 and RelA Acetylation in MCF-7 and DU145 Cells
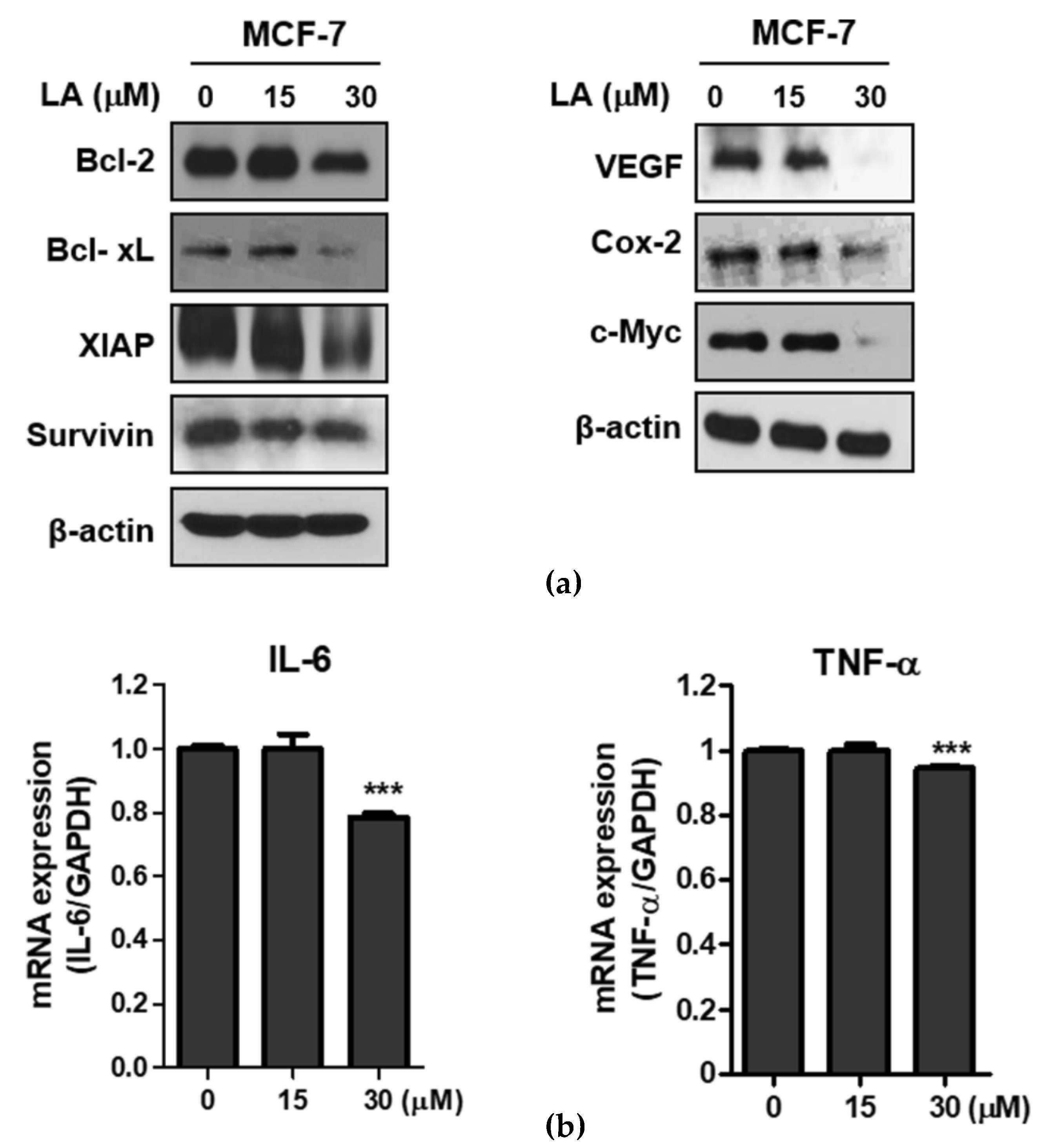
2.3. LA Attenuated the Expression of NF-κB Regulated Genes in MCF-7 Cells
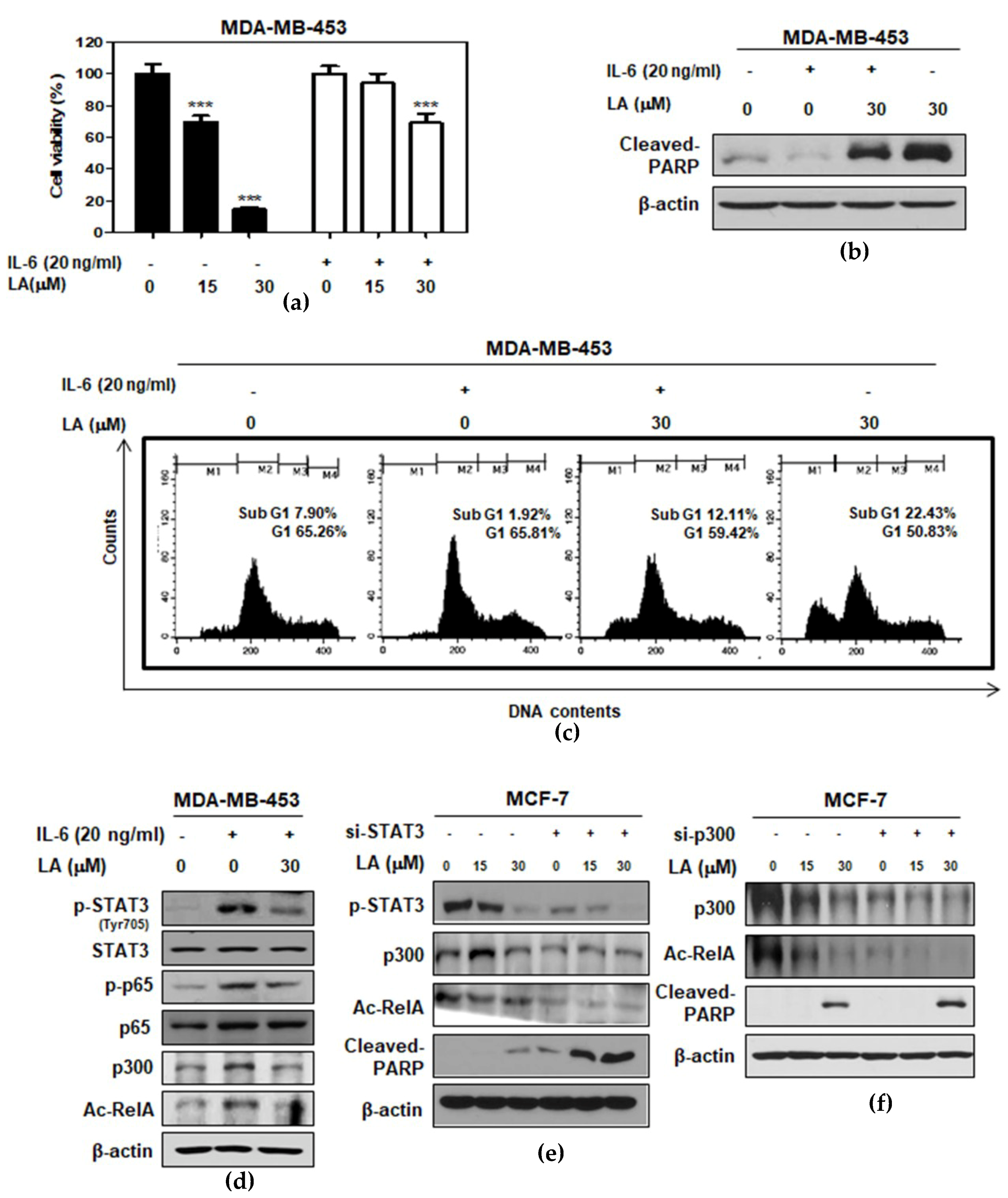
2.4. STAT3 Inducer IL-6 Suppressed Cytotoxic and Apoptotic Effects of LA in MDA-MB-453 Cells
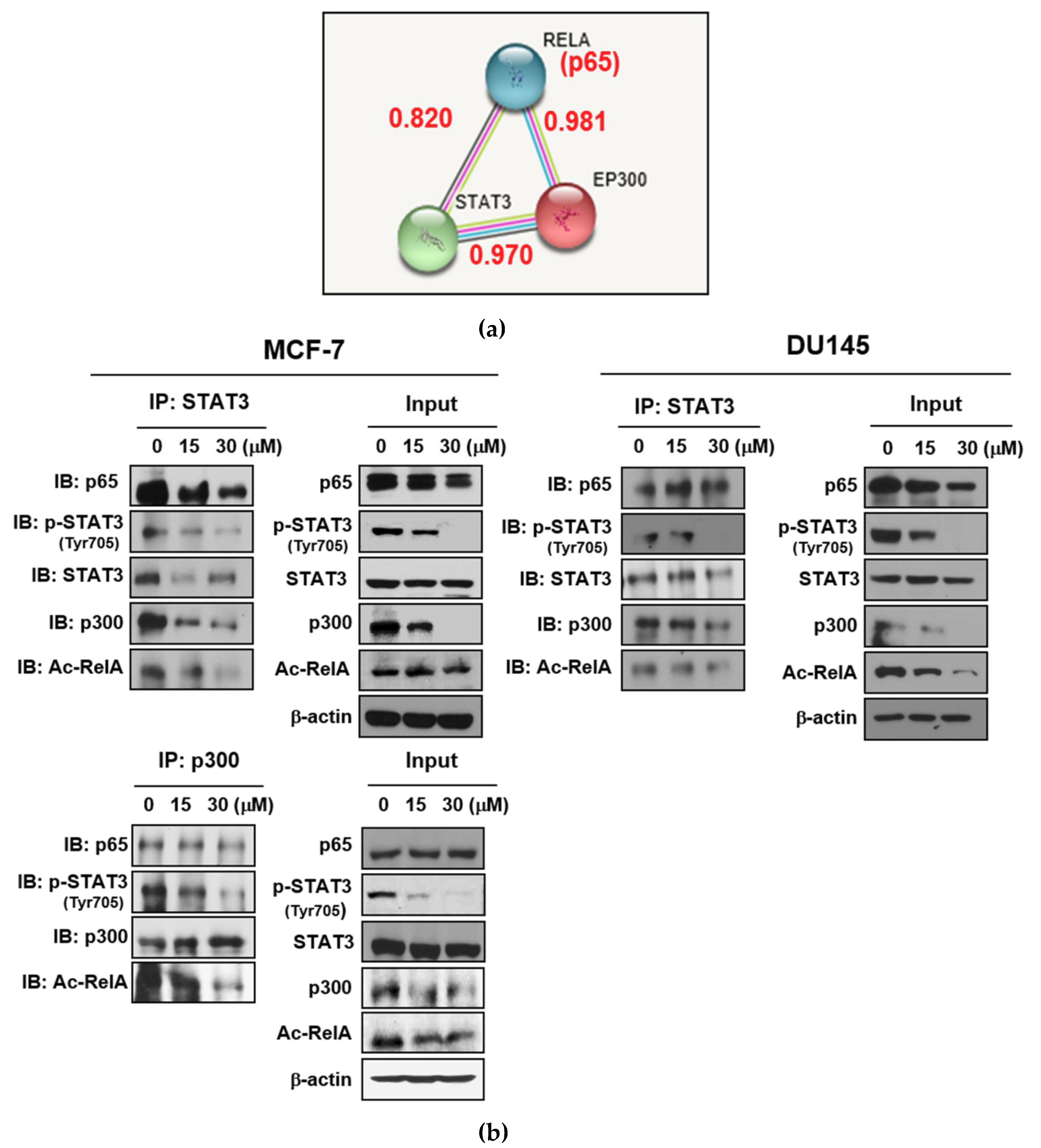
2.5. LA Inhibited the Expression of p300 and RelA/65 Acetylation, and Disrupted the Interaction between p-STAT3, p300, and NF-κB in MCF-7 and DU145 Cells
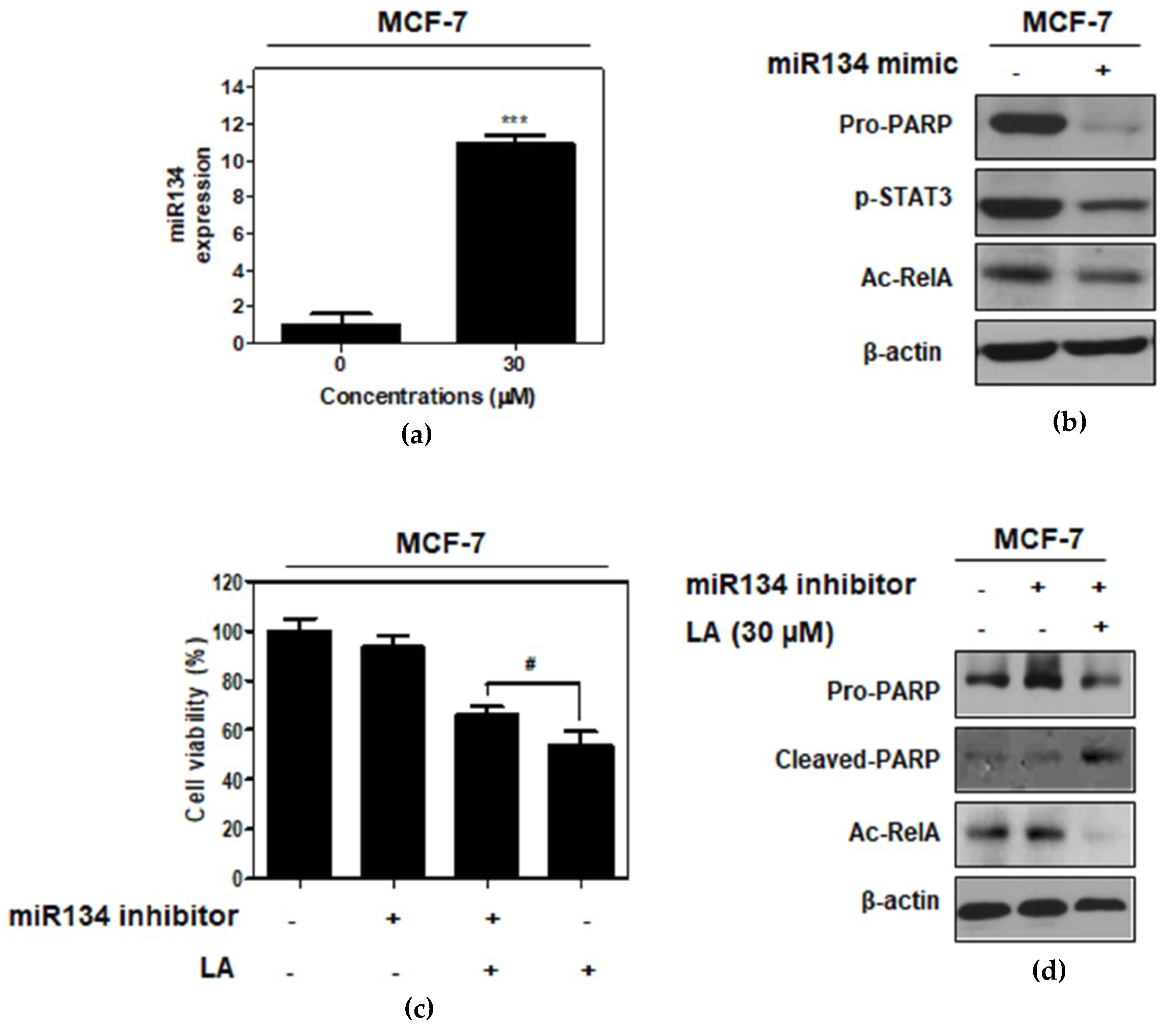
2.6. miR134 Plays a Pivotal Role in the LA-Induced Apoptotic Effect in MCF-7 Cells
3. Discussion
4. Materials and Methods
4.1. Isolation of Lambertianic Acid (LA)
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Cycle Analysis
4.5. Western Blotting
4.6. Immunofluorescence
4.7. Co-Immunoprecipitation
4.8. RNA Interference
4.9. RT-qPCR Analysis
4.10. MicroRNA Transfection Assay
4.11. Statistical Analysis
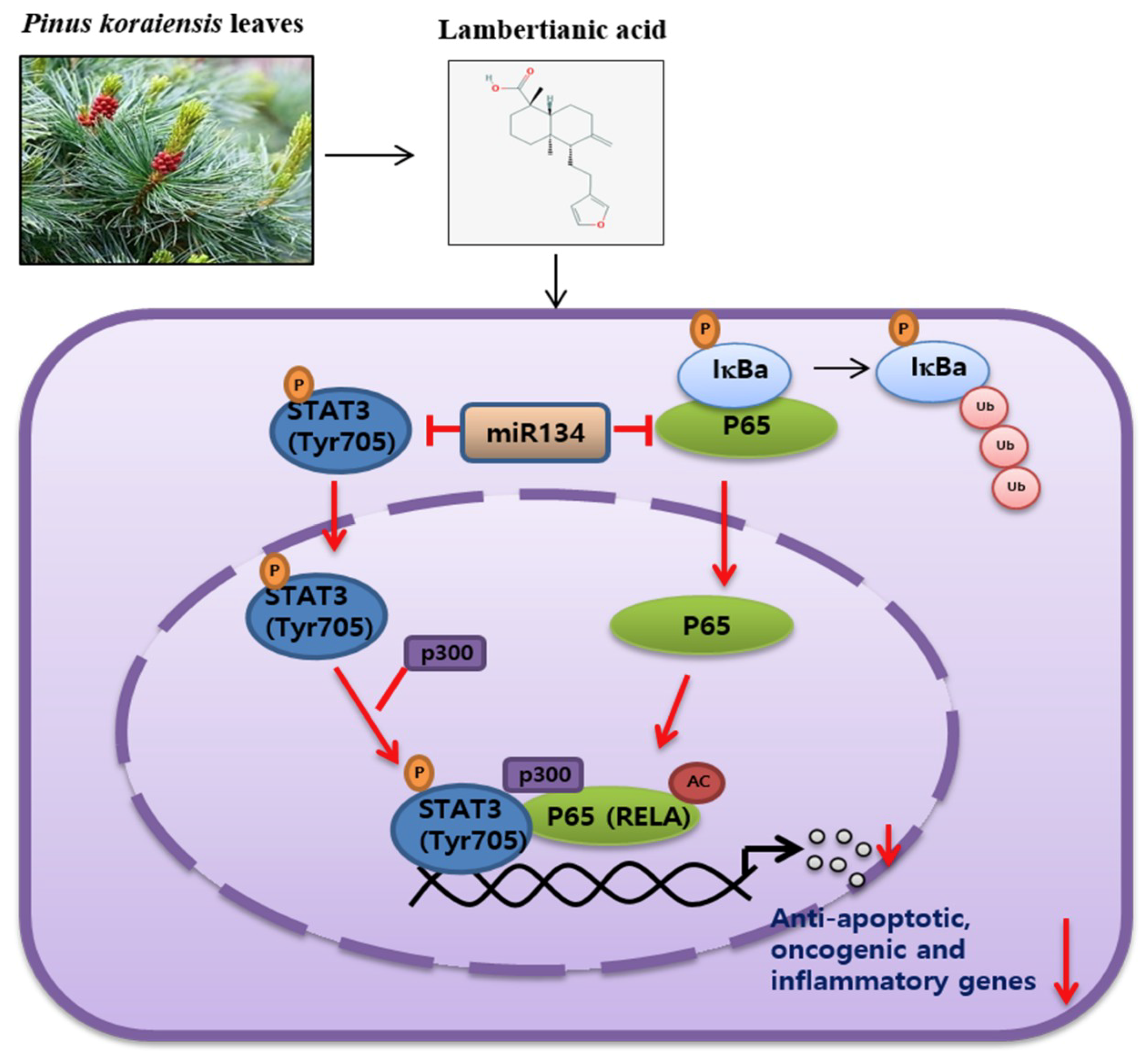
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| STAT3 | Signal transducers and activators of transcription 3 |
| PARP | Poly (ADP-ribose) polymerase |
| IL-6 | Interleukin 6 |
| PPI | Protein–protein interaction |
| Ac-RelA | RelA/p65 acetylation |
| HAT | Histone acetyltransferase |
References
- Wang, Y.; van Boxel-Dezaire, A.H.; Cheon, H.; Yang, J.; Stark, G.R. STAT3 activation in response to IL-6 is prolonged by the binding of IL-6 receptor to EGF receptor. Proc. Natl. Acad. Sci. USA 2013, 110, 16975–16980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuno, Y.; Kiwamoto, T.; Morishima, Y.; Ishii, Y.; Hizawa, N.; Hogaboam, C.M. Notch signaling regulates cell density-dependent apoptosis of NIH 3T3 through an IL-6/STAT3 dependent mechanism. Eur. J. Cell Biol. 2018, 97, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Tapia, M.; Hernandez, C.J.; Ascui, G.; Kramm, K.; Morales, M.; Ga Rate, V.; Zuniga, R.; Bustamante, M.; Aguillon, J.C.; Catala, N.D.; et al. STAT3 inhibition by STA21 increases cell surface expression of MICB and the release of soluble MICB by gastric adenocarcinoma cells. Immunobiology 2017, 222, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Zhu, L.; Wang, Y.; Shi, Y.; Gong, A.; Wu, C. Stattic enhances radiosensitivity and reduces radio-induced migration and invasion in HCC cell lines through an apoptosis pathway. BioMed Res. Int. 2017, 2017, 1832494. [Google Scholar] [CrossRef] [PubMed]
- Affandi, I.S.M.; Lee, W.Q.; Feroz, S.R.; Mohamad, S.B.; Tayyab, S. Interaction of stattic, a STAT3 inhibitor with human serum albumin: Spectroscopic and computational study. J. Biomol. Struct. Dyn. 2017, 35, 3581–3590. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, A.; Dadmanesh, M.; Ghorban, K. Suppression of STAT3 by S31-201 to reduce the production of immunoinhibitory cytokines in a HIF1-alpha-dependent manner: A study on the MCF-7 cell line. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Resetca, D.; Haftchenary, S.; Gunning, P.T.; Wilson, D.J. Changes in signal transducer and activator of transcription 3 (STAT3) dynamics induced by complexation with pharmacological inhibitors of Src homology 2 (SH2) domain dimerization. J. Biol. Chem. 2014, 289, 32538–32547. [Google Scholar] [CrossRef] [PubMed]
- Johnston, P.A.; Grandis, J.R. STAT3 signaling: Anticancer strategies and challenges. Mol. Interv. 2011, 11, 18–26. [Google Scholar] [CrossRef]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim. Biophys. Acta 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.F.; Greene, W.C. Shaping the nuclear action of NF-kappaB. Nat. Rev. Mol. Cell Biol. 2004, 5, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.C.; Jung, M.G.; Lee, Y.H.; Yoon, J.C.; Kwon, S.H.; Kang, H.B.; Kim, M.J.; Cha, J.H.; Kim, Y.J.; Jun, W.J.; et al. Epigallocatechin-3-gallate, a histone acetyltransferase inhibitor, inhibits EBV-induced B lymphocyte transformation via suppression of RelA acetylation. Cancer Res. 2009, 69, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.C.; Lee, Y.H.; Jung, M.G.; Kwon, S.H.; Kim, M.J.; Jun, W.J.; Lee, J.; Lee, J.M.; Yoon, H.G. Gallic acid suppresses lipopolysaccharide-induced nuclear factor-kappaB signaling by preventing RelA acetylation in A549 lung cancer cells. Mol. Cancer Res. 2009, 7, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.J.; Tsai, T.; Peng, P.C.; Li, P.T.; Chen, C.T. Histone acetyltransferase p300 is induced by p38MAPK after photodynamic therapy: The therapeutic response is increased by the p300HAT inhibitor anacardic acid. Free Radic. Biol. Med. 2015, 86, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Ryan, B.C.; Werner, T.S.; Howard, P.L.; Chow, R.L. ImiRP: A computational approach to microRNA target site mutation. BMC Bioinform. 2016, 17, 190. [Google Scholar] [CrossRef]
- Ye, Q.; Su, L.; Chen, D.; Zheng, W.; Liu, Y. Astragaloside IV induced miR-134 expression reduces EMT and increases chemotherapeutic sensitivity by suppressing CREB1 signaling in colorectal cancer cell line SW-480. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 1617–1626. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Qian, J.; Bao, M.; Meng, X.; Zhang, S.; Zhang, L.; Zhao, R.; Li, S.; Cao, Q.; et al. miR-134 functions as a tumor suppressor in cell proliferation and epithelial-to-mesenchymal Transition by targeting KRAS in renal cell carcinoma cells. DNA Cell Biol. 2015, 34, 429–436. [Google Scholar] [CrossRef]
- Lee, M.S.; Cho, S.M.; Lee, M.H.; Lee, E.O.; Kim, S.H.; Lee, H.J. Ethanol extract of Pinus koraiensis leaves containing lambertianic acid exerts anti-obesity and hypolipidemic effects by activating adenosine monophosphate-activated protein kinase (AMPK). BMC Complement. Altern. Med. 2016, 16, 51. [Google Scholar] [CrossRef]
- Chae, H.S.; Chin, Y.W. Anti-allergic effect of lambertianic acid from Thuja orientalis in mouse bone marrow-derived mast cells. Immunopharmacol. Immunotoxicol. 2012, 34, 250–255. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, H.J.; Sim, D.Y.; Jung, J.H.; Kim, K.R.; Kim, S.H. Apoptotic effect of lambertianic acid through AMPK/FOXM1 signaling in MDA-MB231 breast cancer cells. Phytother. Res. 2018, 32, 1755–1763. [Google Scholar] [CrossRef]
- Jeong, A.; Kim, J.H.; Lee, H.J.; Kim, S.H. Reactive oxygen species dependent phosphorylation of the liver kinase B1/AMP activated protein kinase/acetyl-CoA carboxylase signaling is critically involved in apoptotic effect of lambertianic acid in hepatocellular carcinoma cells. Oncotarget 2017, 8, 70116–70129. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Lee, S.O.; Kim, S.H.; Lee, E.O.; Lee, H.J. Anti-cancer effect of lambertianic acid by inhibiting the AR in LNCaP cells. Int. J. Mol. Sci. 2016, 17, 1066. [Google Scholar] [CrossRef] [PubMed]
- Lieblein, J.C.; Ball, S.; Hutzen, B.; Sasser, A.K.; Lin, H.J.; Huang, T.H.; Hall, B.M.; Lin, J. STAT3 can be activated through paracrine signaling in breast epithelial cells. BMC Cancer 2008, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Karin, M. NF-kappaB and STAT3-key players in liver inflammation and cancer. Cell Res. 2011, 21, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef]
- Kaliyaperumal, K.; Sharma, A.K.; McDonald, D.G.; Dhindsa, J.S.; Yount, C.; Singh, A.K.; Won, J.S.; Singh, I. S-Nitrosoglutathione-mediated STAT3 regulation in efficacy of radiotherapy and cisplatin therapy in head and neck squamous cell carcinoma. Redox Biol. 2015, 6, 41–50. [Google Scholar] [CrossRef]
- Gong, J.; Xie, J.; Bedolla, R.; Rivas, P.; Chakravarthy, D.; Freeman, J.W.; Reddick, R.; Kopetz, S.; Peterson, A.; Wang, H.; et al. Combined targeting of STAT3/NF-kappaB/COX-2/EP4 for effective management of pancreatic cancer. Clin. Cancer Res. 2014, 20, 1259–1273. [Google Scholar] [CrossRef]
- Kim, J.W.; Jang, S.M.; Kim, C.H.; An, J.H.; Kang, E.J.; Choi, K.H. New molecular bridge between RelA/p65 and NF-kappaB target genes via histone acetyltransferase TIP60 cofactor. J. Biol. Chem. 2012, 287, 7780–7791. [Google Scholar] [CrossRef]
- Lee, H.; Herrmann, A.; Deng, J.H.; Kujawski, M.; Niu, G.; Li, Z.; Forman, S.; Jove, R.; Pardoll, D.M.; Yu, H. Persistently activated Stat3 maintains constitutive NF-kappaB activity in tumors. Cancer Cell 2009, 15, 283–293. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-kappaB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Oh, J.H.; Cho, M.C.; Kim, J.H.; Lee, S.Y.; Kim, H.J.; Park, E.S.; Ban, J.O.; Kang, J.W.; Lee, D.H.; Shim, J.H.; et al. IL-32gamma inhibits cancer cell growth through inactivation of NF-kappaB and STAT3 signals. Oncogene 2011, 30, 3345–3359. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Zhang, G.; Hwa, Y.L.; Li, J.; Dowdy, S.C.; Jiang, S.W. Nonhistone protein acetylation as cancer therapy targets. Expert Rev. Anticancer Ther. 2010, 10, 935–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Martile, M.; Del Bufalo, D.; Trisciuoglio, D. The multifaceted role of lysine acetylation in cancer: Prognostic biomarker and therapeutic target. Oncotarget 2016, 7, 55789–55810. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, X.; Chen, S.; Price, B.D. Inhibition of histone acetyltransferase activity by anacardic acid sensitizes tumor cells to ionizing radiation. FEBS Lett. 2006, 580, 4353–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arif, M.; Pradhan, S.K.; Thanuja, G.R.; Vedamurthy, B.M.; Agrawal, S.; Dasgupta, D.; Kundu, T.K. Mechanism of p300 specific histone acetyltransferase inhibition by small molecules. J. Med. Chem. 2009, 52, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Marcu, M.G.; Jung, Y.J.; Lee, S.; Chung, E.J.; Lee, M.J.; Trepel, J.; Neckers, L. Curcumin is an inhibitor of p300 histone acetylatransferase. Med. Chem. 2006, 2, 169–174. [Google Scholar]
- Sung, B.; Pandey, M.K.; Ahn, K.S.; Yi, T.; Chaturvedi, M.M.; Liu, M.; Aggarwal, B.B. Anacardic acid (6-nonadecyl salicylic acid), an inhibitor of histone acetyltransferase, suppresses expression of nuclear factor-kappaB-regulated gene products involved in cell survival, proliferation, invasion, and inflammation through inhibition of the inhibitory subunit of nuclear factor-kappaBalpha kinase, leading to potentiation of apoptosis. Blood 2008, 111, 4880–4891. [Google Scholar]
- Sassen, S.; Miska, E.A.; Caldas, C. MicroRNA: Implications for cancer. Virchows Arch. Int. J. Pathol. 2008, 452, 1–10. [Google Scholar] [CrossRef]
- Wang, S.; Wu, G.; Han, Y.; Song, P.; Chen, J.; Wu, Y.; Yang, J.; Liang, P. miR-124 regulates STAT3-mediated cell proliferation, migration and apoptosis in bladder cancer. Oncol. Lett. 2018, 16, 5875–5881. [Google Scholar] [CrossRef]
- Yang, X.; Ding, Y.; Sun, Z.H.; Zhang, D.M. [Studies on chemical constituents of Pinus armandii]. Acta Pharm. Sin. 2005, 40, 435–437. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, D.Y.; Lee, H.-J.; Jung, J.H.; Im, E.; Hwang, J.; Kim, D.S.; Kim, S.-H. Suppression of STAT3 Phosphorylation and RelA/p65 Acetylation Mediated by MicroRNA134 Plays a Pivotal Role in the Apoptotic Effect of Lambertianic Acid. Int. J. Mol. Sci. 2019, 20, 2993. https://doi.org/10.3390/ijms20122993
Sim DY, Lee H-J, Jung JH, Im E, Hwang J, Kim DS, Kim S-H. Suppression of STAT3 Phosphorylation and RelA/p65 Acetylation Mediated by MicroRNA134 Plays a Pivotal Role in the Apoptotic Effect of Lambertianic Acid. International Journal of Molecular Sciences. 2019; 20(12):2993. https://doi.org/10.3390/ijms20122993
Chicago/Turabian StyleSim, Deok Yong, Hyo-Jung Lee, Ji Hoon Jung, Eunji Im, Jisung Hwang, Dong Sub Kim, and Sung-Hoon Kim. 2019. "Suppression of STAT3 Phosphorylation and RelA/p65 Acetylation Mediated by MicroRNA134 Plays a Pivotal Role in the Apoptotic Effect of Lambertianic Acid" International Journal of Molecular Sciences 20, no. 12: 2993. https://doi.org/10.3390/ijms20122993