Updates on Aptamer Research


1
Center for Aging and Associated Diseases, Zewail City of Science and Technology, Giza 12578, Egypt
2
Department of Biochemistry and Cell Biology, Stony Brook University, Stony Brook, NY 11794-5215, USA
3
Department of Pharmacology and Toxicology, Ahram Canadian University, 6th of October City, Giza 12566, Egypt
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(10), 2511; https://doi.org/10.3390/ijms20102511
Submission received: 27 March 2019
/
Revised: 26 April 2019
/
Accepted: 30 April 2019
/
Published: 21 May 2019
(This article belongs to the Section Molecular Biology)
Abstract
:For many years, different probing techniques have mainly relied on antibodies for molecular recognition. However, with the discovery of aptamers, this has changed. The science community is currently considering using aptamers in molecular targeting studies because of the many potential advantages they have over traditional antibodies. Some of these possible advantages are their specificity, higher binding affinity, better target discrimination, minimized batch-to-batch variation, and reduced side effects. Overall, these characteristics of aptamers have attracted scholars to use them as molecular probes in place of antibodies, with some aptamer-based targeting products being now available in the market. The present review is aimed at discussing the potential of aptamers as probes in molecular biology and in super-resolution microscopy.
1. Introduction
Nucleic acids (NAs) were for a long time considered compounds whose major functions were related to the storage of inherited information (DNA) and its transfer from gene to protein through RNA [1,2]. However, studies have over time confirmed that NAs perform other functions such as enzymatic catalysis and the regulation of transcription. The discovery of these additional functions of NAs has compelled the scholarly community to reconsider its original position concerning the functions of NAs [1]. Consequently, this change has led the scientific community to propose the “RNA world theory” [3,4,5]. In particular, this theory posits that NAs possess the ability to perform multiple functions and have presumably facilitated all the catalytic reactions that started life on earth [4,6]. The discovery of aptamers is indeed a valuable contribution towards the empirical determination of the multifunctional nature of NAs [1,7]. Essentially, aptamers are small, often ranging between 20 to 60 nucleotides, single-stranded DNA or RNA oligonucleotides that are capable of binding target molecules not only with high specificity but also with high affinity [8,9,10,11,12].
Aptamers, the output of systematic evolution of ligands by exponential enrichment (SELEX) process, have been available for about three decades. In particular, the development of aptamers is attributed to the work of Craig Tuerk and Larry Gold in 1990 [13]. Gold and Tuerk focused on investigating the nature of the “translational operator” within the bacteriophage T4 gene 43 mRNA. This research led to performing the first SELEX experiment that produced two winning hairpins, the wild-type T4 sequence as well as another containing quadruple mutations, both bound to the gene 43 protein with the same affinities [13,14]. Although, Tuerk’s experiments pioneered the SELEX process, the process was not named SELEX until Szostak and Ellington developed the same method and coined the term SELEX [15]. The term refers to a process that entails progressive separation of single-stranded DNAs or RNAs (ssDNA/RNA) combinatorial single-stranded oligonucleotide library via repeated rounds of binding, partitioning, and amplification [16,17,18]. It is noted that following the development of this novel technology, different modified SELEX techniques have emerged over the past three decades. Table 1 summarizes the different SELEX methods, as described in their original context. Some of these developments tackled the ssDNA/RNA library improvement, while many others addressed the development of the SELEX process. In the contemporary research environment, cell-SELEX (illustrated in Figure 1) has been extensively used to identify and select aptamers that can help in the diagnosis as well as the development of treatments for different diseases, particularly for cancer [18,19,20]. Thus, cell-SELEX technology has become increasingly important in medical research.
Currently, a significant number of generated aptamers are indeed capable of binding different targets, including large protein complexes, simple inorganic molecules, and whole cells [10]. From this prospective, aptamers are simply considered nucleotide analogues of antibodies [11,106,107]. However, compared to antibodies, the generation of aptamers is significantly easier and cheaper. Thus if appropriately developed, these characteristics make aptamers ideal candidates for a wide variety of applications such as disease diagnosis, biosensor design, nanotherapy, molecular imaging, and the purification of particular target molecules from complex mixtures [9,10,108].
The science community has in the recent years developed super-resolution imaging techniques that offer three-dimensional imaging capabilities that overcome the diffraction restrictions of conventional light microscopy especially in viewing lateral dimensions [109,110]. For example, techniques such as single-molecule localization or stimulated emission depletion microscopy (STED) and secondary ion mass spectrometry microscopy have enabled scientists to visualize whether objects are in fact localized [111,112]. However, the advancement of these techniques requires additional improvements in the fluorescent labels to harness the power of super resolution as the image resolution is also restricted by the effectiveness of the labelling probes’ size, specificity, stability, and density [113]. Indeed, the development of these novel techniques has encouraged researchers to reconsider labelling in super resolution. Aptamers’ favourable characteristics have attracted scientists to consider using them in place of antibodies in super resolution. The focus of this review is to demonstrate the potential of aptamers as probing tools that can be used in super resolution.
2. Antibodies and Aptamers
Antibodies, as pointed out in the preceding section, are a popular class of compounds that have long been used in research to provide molecular identification for a broad array of applications, including disease diagnosis and therapy. Consequently, antibodies have made significant contributions towards the improvement of diagnostic assays [114,115]. In fact, it can be argued that antibodies have become increasingly indispensable in most of the clinical diagnostic tests in the modern medicine. However, the emergence of SELEX (outlined in Table 1) has made it possible for researchers to progressively isolate oligonucleotide sequences capable of recognizing nearly any class of target molecules not only with high affinity but with high specificity as well [115]. Although completely distinct from antibodies, aptamers are thought to emerge as a class of compounds that rival antibodies in terms of both diagnostic and therapeutic applications [116]. Indeed, the available body of knowledge has confirmed that this class of molecules mimics the properties of antibodies in several formats. Mairal et al. contended that the high demand for diagnostic assays that can effectively help in the management of present and emerging diseases has soared in recent years and it is suggested that aptamers could potentially satisfy various molecular recognition needs in these diagnostic assays [114]. Although the research on aptamers is still in its infancy, there is compelling evidence that this field of research is progressing at a fast pace [117].
The discovery of aptamers has already helped circumvent some of the key limitations associated with antibodies [118]. A critical evaluation of the present body of knowledge on antibodies and aptamers shows that these classes both have advantages and disadvantages (summarized in Table 2) over each other with multiple pieces of empirical evidence reveal that aptamers provide a broad array of clear-cut merits over conventional antibodies [115,119,120,121].
Some of the key advantages of aptamers include their inertness towards the surrounding cells [114,122,123]. Specific aptamers are capable of binding to a target molecule with high precision and, thus, it is thought that this can facilitate a broad array of diagnostic and therapeutic applications without the fear of potential non-specific binding [114]. Another advantage is that they often have a higher binding affinity [123,124]. It is indeed acknowledged that in cases where several ligands can bind to the same receptor site, aptamers have been found to offer a relatively higher binding affinity. It is worth noting that, in practice, the higher the affinity level of a biological molecule used in molecular recognition, the less quantity is needed in molecular identification, which may reduce the costs of performing molecular identification studies [122,125,126].
Aptamers also often exhibit excellent target molecule specificity compared to antibodies [123]. According to Kedzierski et al., there are aptamers that exhibit >10,000-fold binding affinity for theophylline over caffeine [123]. There is no doubt that this excellent target specificity makes aptamers ideal bio-markers as they increase the accuracy of molecular recognition. Another advantage is that aptamers facilitate the discovery of unknown biomarkers [123,127]. Kedzierski et al. explained that different SELEX techniques are capable of identifying unknown biomarkers and consequently, this has in the recent years hastened the discovery of diseases and therapeutics [123]. Some other advantages of aptamers over the conventional antibodies include discovery time savings, minimized batch-to-batch variation, in vitro vs. in vivo testing advantages, increased stability, and reduced side effects [123].
It is worth noting that nanobodies, antibody-mimicking binders that are a single variable domain of an antibody, are also capable of specific binding and are also used in molecular targeting [128]. Previous studies have suggested that the use of affinity probes such as aptamers and nanobodies has many advantages in light microscopy, particularly in the field of super resolution [129,130]. De Castro et al. explained that steric hindrance causes less impairment on these small molecular probes and, thus, facilitates their penetration into biological samples and their binding to epitopes that are traditionally inaccessible to larger antibodies [129]. Indeed, traditional light microscopy suffers from limited resolution capabilities due to light diffraction [113,131]. Many methodologies that are capable of overcoming this limitation were developed in the recent years [113]. In fact, it has been reported that diffraction-unlimited microscopes are improving rapidly and, as a result, it is now practical to attain excellent resolutions of small elements of less than 10 nm [132]. Nevertheless, the enhancement of sample preparation and methodologies used in staining is still lagging behind [39]. The conventional immunostaining technique largely depends on affinity tools that at times are larger than the protein of interest making it impossible to fully exploit the potential of contemporary imaging techniques [129,133]. However, it is anticipated that small probes such as nanobodies and aptamers would have a significant impact on the precision of staining biological samples [129]. Recent studies have in fact suggested that when compared to traditional antibody techniques, nanobodies are capable of positioning the fluorescent molecules closer to the intended target, and thereby resulting in enhanced localization precisions using super-resolution microscopy [134,135]. Likewise, in a comparative study of aptamers vs. antibodies that involved STED, it was concluded that in super-resolution microscopy, aptamers offer superior staining of different cellular receptors. Since aptamers are cheap to select and nanobodies usually have higher affinities, it is thought that aptamers and nanobodies complement each other and together they should help advance the effectiveness of super-resolution imaging [136].
Although the use of aptamers as molecular identification compounds has emerged as a viable approach for diagnostics, therapeutics, and biosensing [124], aptamers do present a set of challenges that have inadvertently hampered both their research and commercialization. Firstly, relatively few aptamers bind small molecules [124]. It is argued that small molecules are crucial targets for research because of their clinical and commercial uses as well as diverse biological functions. Therefore, the availability of relatively few aptamers binding to small molecules restricts research on their biological functions, clinical use, and commercial use. Secondly, aptamers are likely to encounter rapid clearance rate from the circulation because of their small size as well as degradation by nucleases especially for unmodified aptamers [123]. Further, there are examples where developing a selective aptamer still represents a challenge. For example, aptamers that were selected against monomeric/oligomeric forms of amyloid peptide had very low affinity to these forms, although they exhibited a strong non-specific affinity to amyloid high-molecular weight fibrils [137,138]. It is thought, however, that aptamers still hold promise for the treatment of Alzheimer’s disease because current amyloid inhibitors and antibodies are not effective at the membrane surface, where inhibition of amyloid aggregate and channel formation is needed to stop disease progression [139]. Janas et al. 2019 shows that functional exosomes containing the selected pool of aptamers can inhibit the formation of amyloid aggregates and channel activity at the membrane surface [139].
Nevertheless, aptamer microarray technology has recently been developed in an effort to overcome the drawbacks and limitations of antibody microarrays, especially when targeting small molecules with one epitope [140]. Indeed, aptamers that target small organics, peptides, proteins, viruses, cells, and bacteria are now commercially available from a number of emerging companies such as Aptagen and AMSBIO. Other companies have been established focusing on aptamers-based drug discovery such as OSI Pharmaceuticals, Nascacell Technologies in collaboration with Discovery Partners International, and Aptanomics. In addition, aptamer-based clinical trials are currently being run by different companies such as Baxter Biosciences, Corgentech, Bristol-Myers Squibb, Gilead Sciences, NOXXON Pharma, Oxford Cancer Research, Pfizer, and Regado Biosciences (reviewed in [141,142,143]), while others are developing aptamer-based diagnostics or therapeutics including Aptamer Group, Base Pair Biotechnologies, and Ophthotech Corporation [144].
3. Super-Resolution Imaging
For many years, light microscopy has significantly improved the scientific understanding of how cells function [113]. In fact, the different spheres of biology are believed to have developed from images that have been acquired under light microscopes. Nonetheless, studies that utilize conventional light microscopy techniques have been limited to the resolution of about 200 nm [113]. The science community has for many years focused on developing several imaging techniques to address the diffraction of light [145]. Some of these imaging techniques include multiphoton fluorescence microscopy and confocal microscopy. Fundamentally, these novel imaging techniques have not only significantly enhanced the resolution of cellular images, but have also reduced the out-of-focus fluorescence background [113]. Consequently, this has facilitated optical sectioning and three-dimensional imaging. Although these techniques have significantly improved resolution, the approaches are still limited by light diffraction [146]. In practice, these advanced techniques have often achieved resolutions of ~100 nm in all three dimensions [113,146].
Recently, this limitation in light microscopy has been overcome by many improved microscopy techniques, with some of the techniques achieving resolutions of ~10 nm [130]. The capability of these novel advanced microscopy techniques to circumvent light diffraction has significantly improved imaging precision [113]. Consequently, this has enabled researchers to identify important cellular details that could not be isolated using the traditional diffraction-limited instruments. STED, single-molecule localization techniques (SMLM), positron-emission tomography (PET), single-photon emission computed tomography (SPECT), and cryo-electron microscopy (cryo-EM) are some of the common diffraction-unlimited techniques that have made a significant impact in biological sciences [147,148,149,150,151]. For example, stochastic optical reconstruction microscopy (STORM), an SMLM technique, offers a high spatial resolution for the position of the fluorophore [152]. It is worth noting that large labels, for example, antibodies, can provide a misleading position of the fluorophore from the target molecule [153]. However, the discovery and subsequent use of different SMLM techniques made it possible to accurately pinpoint the position of the fluorophore from the target molecule.
In principle, STED utilizes pairs of synchronized laser pulses, which play a critical role in overcoming the challenge of light diffraction [154]. The basic functioning mechanism of this super-resolution microscopy technique is that it creates images through selective deactivation of fluorophores [155]. This deactivation reduces the illumination area at the focal point and thus improves the achievable resolution for a specific system.
PET has also remarkably revolutionized the field of super resolution, particularly in biomedical research [91,93,94]. This advanced imaging technique utilizes small amounts of radiotracers, a unique camera, and a computer to help in the evaluation of tissue and organ functions in patients [156]. It is actually possible to detect early onset of diseases by identifying changes happening at the cellular level using PET [130,157,158,159].
Finally, cryo-EM is another super-resolution technique that is used to image frozen hydrated specimens at cryogenic temperatures [160,161]. This imaging technique allows the specimens used in super-resolution studies to remain in their native state [161]. Thus, this enables the study of fine cellular structures, protein complexes, and viruses at molecular resolution [162].
4. Aptamers for Super-Resolution Imaging
The enhanced imaging precision that resulted from the development of the previously discussed imaging techniques has also revealed that traditional staining using large affinity tags, for example, antibodies, are not sufficiently accurate [130]. It is argued that since aptamers are significantly smaller than antibodies, these molecules might provide a practical advantage in super-resolution imaging [129,130,163].
In their study, de Castro et al. particularly compared the live staining of transferrin receptors acquired with varied fluorescently labelled affinity probes, including the natural receptor ligand transferrin, specific monoclonal antibodies, or aptamers [130]. Analysis of the collected data showed that there were insignificant variations between the three mentioned staining strategies when imaging is carried out with traditional laser scanning confocal microscopy. Nevertheless, the aptamer tag showed clear superiority over antibodies in the super-resolved images acquired with STED microscopy. Thus it was concluded that compared to the natural receptor ligand transferrin and specific monoclonal antibodies, aptamer staining is ideal in super-resolution microscopy [130]. It is worth noting that similar observations were reported when aptamers were compared to antibodies targeting different epitopes including prostate-specific membrane antigen and EGFR [18,129,136,164].
In another study that targeted three membrane receptors that are relevant to human health and cycle between the intracellular space and the plasma membrane, de Castro et al. observed that, compared to the majority of antibodies, aptamers were capable of revealing more epitopes and thus, offered denser labelling of stained structures, which enhanced the quality of the information obtained from the images [129]. In this study aptamer labelling was advantageous under both super-resolution imaging and light microscopy imaging.
EGFR aptamer labelling also achieved a better quality in dSTORM imaging. In a recent study, aptamer labelling achieved detailed and precise structural analysis of the active EGFR that forms large clusters compared to EGFR at rest. This active-to-rest EGFR structure difference has not been detected using traditional antibodies [163]. The ability of aptamers to detect small structural changes makes them promising candidates for studying the morphological changes occur for the membrane proteins during different biological activities [163].
Additionally, in vivo PET scanning of BT474 mice showed higher uptake of 18F-labelled aptamers that specifically bind to breast cancer expressing HER2 [165]. The confocal images also confirmed the specificity of 18F-labelled aptamers to HER2-positive cells [165]. For strategies employed in the development of aptamer-based SPECT and PET imaging we refer the readers to an informative review by Hassanzadeh et al. [166].
Aptamers are particularly appropriate structure-changing molecules that are capable of combining highly selective biorecognition as well as signal transduction [167]. It is possible to select aptamers towards nearly any ligand molecules through SELEX and can readily be incorporated into the functional devices that boost new applications that range from DNA-machines in sensors to the bioseparations. It has been reported that binding of ligands often affects the stability of the nucleotides that the ligand directly contacts [168]. Ligand binding also causes distal rearrangements in the structure of aptamers. These structural changes can easily be observed by evaluating the cleavage patterns using cryo-EM. Aptamer-based labelling need to be done before the freezing step [162]. In their study, Stanlis et al. showed that ssDNA aptamers could be a useful tool for achieving accurate localization of specific proteins, under freeze-substitution fixation conditions, during studying the cellular fine structures using cryo-EM [169]. They indicated that aptamers are sufficiently soluble probes to be used along with the organic solvents used in freeze-substitution conditions; however, it failed to achieve high-affinity binding [169].
Apart from possessing unique physical properties that make them superior to the conventional antibodies in super-resolution imaging, there are several particular modifications that allow aptamers to achieve the highest possible resolution. Choosing an acceptable photostable probe is one of the key features dictating the super-resolution imaging quality [170,171,172]. Using stable tagged-aptamer-based probes in super-resolution microscopy allows obtaining favourable signal-to-noise ratio and also localizing the probes to specific targets without being degraded by the cellular machinery. One particular approach to achieve acceptable aptamer stability is the use of Spiegelmer technology. For example, instead of endonuclease-sensitive RNA aptamers (D-form), researchers select endonuclease-resistant RNA aptamers (L-form) that result from binding to the mirror-image of the intended target molecule [173,174,175,176]. The mirror-image aptamer (L-form) subsequently is expected to bind the natural target molecule [173,177]. Additionally, the substitution of the natural D-ribose with L-ribose renders the mirror-image aptamer completely stable. Another strategy that can be used to enable aptamers to achieve better resolution is to optimize the density of the target molecule and similarly the concentration of the library, that would increase the signal (target-aptamer interaction)-to-noise (interfering species) ratio [174,178,179].
The discovery of aptamers presents an opportunity for biomedical researchers to develop novel molecular targeting probes and effective diagnostic/treatment tools for different diseases. Undoubtedly as well, these molecules are promising tools for super-resolution microscopy and have already been shown superior to conventional antibodies in some applications.
Funding
This research was funded by Science & Technology Development Fund (STDF), Project ID 15193, Ministry of Scientific Research, Egypt.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| NAs | nucleic acids |
| SELEX | systematic evolution of ligands by exponential enrichment |
| STED | stimulated emission depletion microscopy |
| SMLM | single-molecule localization techniques |
| PET | positron-emission tomography |
| Cryo-EM | cryo-electron microscopy |
| STORM | stochastic optical reconstruction microscopy |
References
- Lakhin, A.; Tarantul, V.; Gening, L. Aptamers: Problems, solutions and prospects. Acta Nat. 2013, 5, 34–43. [Google Scholar]
- Saenger, W. Principles of Nucleic acid Structure; Springer Science & Business Media: Des Moines, IA, USA, 2013; pp. 1–8. [Google Scholar]
- Yarus, M. Getting past the rna world: The initial darwinian ancestor. Cold Spring Harb. Perspect. Biol. 2011, 3, a003590. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.P.; Joyce, G.F. The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef]
- Higgs, P.G.; Lehman, N. The RNA world: Molecular cooperation at the origins of life. Nat. Rev. Genet. 2015, 16, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Blumenstiel, J.P. Evolutionary dynamics of transposable elements in a small RNA world. Trends Genet. 2011, 27, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Wrenger, C. Disease-specific biomarker discovery by aptamers. Cytom. Part A 2009, 75, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Tucker, W.O. Towards Specific DNA Aptamers which Bind and Inhibit wwp1 Hect Ubiquitin Ligase in the Osteoblast. Ph.D. Thesis, The University of Hong Kong, Hong Kong, China, June 2013. [Google Scholar]
- Klussmann, S. The Aptamer Handbook: Functional Oligonucleotides and Their Applications; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Veedu, R.N. Aptamers: Tools for Nanotherapy and Molecular Imaging; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kaur, H.; Bruno, J.G.; Kumar, A.; Sharma, T.K. Aptamers in the therapeutics and diagnostics pipelines. Theranostics 2018, 8, 4016–4032. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Yung, L.Y. Probing high affinity sequences of DNA aptamer against vegf165. PLoS ONE 2012, 7, e31196. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage t4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Janjic, N.; Jarvis, T.; Schneider, D.; Walker, J.J.; Wilcox, S.K.; Zichi, D. Aptamers and the RNA world, past and present. Cold Spring Harb. Perspect. Biol. 2012, 4, a003582. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818. [Google Scholar] [CrossRef]
- Guo, K.-T.; Ziemer, G.; Paul, A.; Wendel, H.P. Cell-selex: Novel perspectives of aptamer-based therapeutics. Int. J. Mol. Sci. 2008, 9, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Qian, J.; Yang, Z.; Pang, Z.; Xi, Z.; Cao, S.; Wang, Y.; Pan, S.; Zhang, S.; Wang, W. Whole-cell selex aptamer-functionalised poly (ethyleneglycol)-poly (ε-caprolactone) nanoparticles for enhanced targeted glioblastoma therapy. Biomaterials 2012, 33, 6264–6272. [Google Scholar] [CrossRef]
- Chen, J.; Tong, T.; Wang, H. Super-resolution imaging in glycoscience: New developments and challenges. J. Innov. Opt. Health Sci. 2016, 09, 1630007. [Google Scholar] [CrossRef] [Green Version]
- Sefah, K.; Shangguan, D.; Xiong, X.; O’donoghue, M.B.; Tan, W. Development of DNA aptamers using cell-selex. Nat. Protoc. 2010, 5, 1169–1185. [Google Scholar] [CrossRef]
- Kaur, H. Recent developments in cell-selex technology for aptamer selection. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. Selection in vitro of single-stranded DNA molecules that fold into specific ligand-binding structures. Nature 1992, 355, 850. [Google Scholar] [CrossRef] [PubMed]
- Berkhout, B.; Klaver, B. In vivo selection of randomly mutated retroviral genomes. Nucleic Acids Res. 1993, 21, 5020–5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulter, L.R.; Landree, M.A.; Cooper, T.A. Identification of a new class of exonic splicing enhancers by in vivo selection. Mol. Cell. Biol. 1997, 17, 2143–2150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenison, R.D.; Gill, S.C.; Pardi, A.; Polisky, B. High-resolution molecular discrimination by RNA. Science 1994, 263, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Dobbelstein, M.; Shenk, T. In vitro selection of RNA ligands for the ribosomal l22 protein associated with epstein-barr virus-expressed RNA by using randomized and cdna-derived RNA libraries. J. Virol. 1995, 69, 8027–8034. [Google Scholar]
- Smith, D.; Kirschenheuter, G.P.; Charlton, J.; Guidot, D.M.; Repine, J.E. In vitro selection of RNA-based irreversible inhibitors of human neutrophil elastase. Chem. Biol. 1995, 2, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.B.; Atkinson, B.L.; Willis, M.C.; Koch, T.H.; Gold, L. Using in vitro selection to direct the covalent attachment of human immunodeficiency virus type 1 rev protein to high-affinity RNA ligands. Proc. Natl. Acad. Sci. USA 1995, 92, 12220–12224. [Google Scholar] [CrossRef]
- Golden, M.C.; Collins, B.D.; Willis, M.C.; Koch, T.H. Diagnostic potential of photoselex-evolved ssdna aptamers. J. Biotechnol. 2000, 81, 167–178. [Google Scholar] [CrossRef]
- Klußmann, S.; Nolte, A.; Bald, R.; Erdmann, V.A.; Fürste, J.P. Mirror-image RNA that binds d-adenosine. Nat. Biotechnol. 1996, 14, 1112. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.G. In vitroselection of DNA to chloroaromatics using magnetic microbead-based affinity separation and fluorescence detection. Biochem. Biophys. Res. Commun. 1997, 234, 117–120. [Google Scholar] [CrossRef]
- Tsai, R.Y.; Reed, R.R. Identification of DNA recognition sequences and protein interaction domains of the multiple-zn-finger protein roaz. Mol. Cell. Biol. 1998, 18, 6447–6456. [Google Scholar] [CrossRef]
- Vant-Hull, B.; Payano-Baez, A.; Davis, R.H.; Gold, L. The mathematics of selex against complex targets1. J. Mol. Biol. 1998, 278, 579–597. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.N.; Jensen, K.B.; Julin, C.M.; Weil, M.; Gold, L. High affinity ligands from in vitro selection: Complex targets. Proc. Natl. Acad. Sci. USA 1998, 95, 2902–2907. [Google Scholar] [CrossRef] [PubMed]
- Homann, M.; Göringer, H.U. Combinatorial selection of high affinity RNA ligands to live african trypanosomes. Nucleic Acids Res. 1999, 27, 2006–2014. [Google Scholar] [CrossRef]
- Burke, D.H.; Willis, J.H. Recombination, RNA evolution, and bifunctional RNA molecules isolated through chimeric selex. RNA 1998, 4, 1165–1175. [Google Scholar] [CrossRef]
- Wu, L.; Curran, J.F. An allosteric synthetic DNA. Nucleic Acids Res. 1999, 27, 1512–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, J.; Imanaka, H.; Yokota, Y.; Sugimoto, N. In vitro selection of aptamers that act with Zn2+. J. Inorg. Biochem. 2000, 82, 197–206. [Google Scholar] [CrossRef]
- White, R.; Rusconi, C.; Scardino, E.; Wolberg, A.; Lawson, J.; Hoffman, M.; Sullenger, B. Generation of species cross-reactive aptamers using “toggle” selex. Mol. Ther. 2001, 4, 567–573. [Google Scholar] [CrossRef]
- Pagratis, N.; Gold, L.; Shtatland, T.; Javornik, B. Truncation selex method. U.S. Patent 6,261,774, 17 July 2001. [Google Scholar]
- Wen, J.-D.; Gray, D.M. Selection of genomic sequences that bind tightly to ff gene 5 protein: Primer-free genomic selex. Nucleic Acids Res. 2004, 32, e182. [Google Scholar] [CrossRef]
- Pan, W.; Xin, P.; Clawson, G.A. Minimal primer and primer-free selex protocols for selection of aptamers from random DNA libraries. BioTechniques 2008, 44, 351–362. [Google Scholar] [CrossRef]
- Pan, W.; Clawson, G.A. Primer-free aptamer selection using a random DNA library. In RNA Therapeutics; Springer: Berlin/Heidelberg, Germany, 2010; pp. 367–383. [Google Scholar]
- Martell, R.E.; Nevins, J.R.; Sullenger, B.A. Optimizing aptamer activity for gene therapy applications using expression cassette selex. Mol. Ther. 2002, 6, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.; Krylov, S.N. Nonequilibrium capillary electrophoresis of equilibrium mixtures− a single experiment reveals equilibrium and kinetic parameters of protein− DNA interactions. J. Am. Chem. Soc. 2002, 124, 13674–13675. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Yang, G.; Zhang, D.; Ding, H.; Wang, H.; Fan, M.; Shen, B.; Shao, N. Single-stranded DNA aptamers that bind differentiated but not parental cells: Subtractive systematic evolution of ligands by exponential enrichment. J. Biotechnol. 2003, 102, 15–22. [Google Scholar] [CrossRef]
- Vater, A.; Jarosch, F.; Buchner, K.; Klussmann, S. Short bioactive spiegelmers to migraine-associated calcitonin gene-related peptide rapidly identified by a novel approach: Tailored-selex. Nucleic Acids Res. 2003, 31, e130. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, S.D.; Bowser, M.T. In vitro evolution of functional DNA using capillary electrophoresis. J. Am. Chem. Soc. 2004, 126, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Okhonin, V.; Berezovski, M.; Krylov, S.N. Sweeping capillary electrophoresis: A non-stopped-flow method for measuring bimolecular rate constant of complex formation between protein and DNA. J. Am. Chem. Soc. 2004, 126, 7166–7167. [Google Scholar] [CrossRef] [PubMed]
- Asai, R.; Nishimura, S.I.; Aita, T.; Takahashi, K. In vitro selection of DNA aptamers on chips using a method for generating point mutations. Anal. Lett. 2004, 37, 645–656. [Google Scholar] [CrossRef]
- Drabovich, A.; Berezovski, M.; Krylov, S.N. Selection of smart aptamers by equilibrium capillary electrophoresis of equilibrium mixtures (eceem). J. Am. Chem. Soc. 2005, 127, 11224–11225. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. Flumag-selex as an advantageous method for DNA aptamer selection. Anal. Bioanal. Chem. 2005, 383, 83–91. [Google Scholar] [CrossRef]
- Berezovski, M.V.; Musheev, M.U.; Drabovich, A.P.; Jitkova, J.V.; Krylov, S.N. Non-selex: Selection of aptamers without intermediate amplification of candidate oligonucleotides. Nat. Protoc. 2006, 1, 1359. [Google Scholar] [CrossRef] [PubMed]
- Win, M.N.; Klein, J.S.; Smolke, C.D. Codeine-binding RNA aptamers and rapid determination of their binding constants using a direct coupling surface plasmon resonance assay. Nucleic Acids Res 2006, 34, 5670–5682. [Google Scholar] [CrossRef]
- Khati, M.; Schüman, M.; Ibrahim, J.; Sattentau, Q.; Gordon, S.; James, W. Neutralization of infectivity of diverse r5 clinical isolates of human immunodeficiency virus type 1 by gp120-binding 2′ f-RNA aptamers. J. Virol. 2003, 77, 12692–12698. [Google Scholar] [CrossRef] [PubMed]
- Ohuchi, S.P.; Ohtsu, T.; Nakamura, Y. Selection of RNA aptamers against recombinant transforming growth factor-β type iii receptor displayed on cell surface. Biochimie 2006, 88, 897–904. [Google Scholar] [CrossRef]
- Lorenz, C.; Gesell, T.; Zimmermann, B.; Schoeberl, U.; Bilusic, I.; Rajkowitsch, L.; Waldsich, C.; Von Haeseler, A.; Schroeder, R. Genomic selex for hfq-binding RNAs identifies genomic aptamers predominantly in antisense transcripts. Nucleic Acids Res. 2010, 38, 3794–3808. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Nakamura, Y.; Ohuchi, S. Hexim1-binding elements on mRNAs identified through transcriptomic selex and computational screening. Biochimie 2012, 94, 1900–1909. [Google Scholar] [CrossRef] [PubMed]
- Layzer, J.M.; Sullenger, B.A. Simultaneous generation of aptamers to multiple gamma-carboxyglutamic acid proteins from a focused aptamer library using deselex and convergent selection. Oligonucleotides 2007, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, A.; Kurth, A.; Dunkhorst, A.; Pänke, O.; Sielaff, H.; Junge, W.; Muth, D.; Scheller, F.; Stöcklein, W.; Dahmen, C. One-step selection of vaccinia virus-binding DNA aptamers by monolex. BMC Biotechnol. 2007, 7, 48. [Google Scholar] [CrossRef]
- Peng, L.; Stephens, B.J.; Bonin, K.; Cubicciotti, R.; Guthold, M. A combined atomic force/fluorescence microscopy technique to select aptamers in a single cycle from a small pool of random oligonucleotides. Microsc. Res. Tech. 2007, 70, 372–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, M.; McBurnett, S.R.; Andrews, C.J.; Allman, A.M.; Bruno, J.G.; Kiel, J.L. Aptamer selection express: A novel method for rapid single-step selection and sensing of aptamers. J. Biomol. Tech. JBT 2008, 19, 311. [Google Scholar]
- Raddatz, M.S.L.; Dolf, A.; Endl, E.; Knolle, P.; Famulok, M.; Mayer, G. Enrichment of cell-targeting and population-specific aptamers by fluorescence-activated cell sorting. Angew. Chem. 2008, 120, 5268–5271. [Google Scholar] [CrossRef]
- Mayer, G.; Ahmed, M.-S.L.; Dolf, A.; Endl, E.; Knolle, P.A.; Famulok, M. Fluorescence-activated cell sorting for aptamer selex with cell mixtures. Nat. Protoc. 2010, 5, 1993. [Google Scholar] [CrossRef]
- Tok, J.B.-H.; Fischer, N.O. Single microbead selex for efficient ssdna aptamer generation against botulinum neurotoxin. Chem. Commun. 2008, 1883–1885. [Google Scholar] [CrossRef]
- Knight, C.G.; Platt, M.; Rowe, W.; Wedge, D.C.; Khan, F.; Day, P.J.; McShea, A.; Knowles, J.; Kell, D.B. Array-based evolution of DNA aptamers allows modelling of an explicit sequence-fitness landscape. Nucleic Acids Res. 2008, 37, e6. [Google Scholar] [CrossRef]
- Chushak, Y.; Stone, M.O. In silico selection of RNA aptamers. Nucleic Acids Res. 2009, 37, e87. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Ding, H.; Huang, Y.; Cao, X.; Yang, G.; Li, J.; Xie, Z.; Meng, Y.; Li, X. Identification of an aptamer targeting hnrnp a1 by tissue slide-based selex. J. Pathol. 2009, 218, 327–336. [Google Scholar] [CrossRef]
- Park, S.-m.; Ahn, J.-Y.; Jo, M.; Lee, D.-k.; Lis, J.T.; Craighead, H.G.; Kim, S. Selection and elution of aptamers using nanoporous sol-gel arrays with integrated microheaters. Lab Chip 2009, 9, 1206–1212. [Google Scholar] [CrossRef]
- Zykovich, A.; Korf, I.; Segal, D.J. Bind-n-seq: High-throughput analysis of in vitro protein–DNA interactions using massively parallel sequencing. Nucleic Acids Res. 2009, 37, e151. [Google Scholar] [CrossRef]
- Jolma, A.; Kivioja, T.; Toivonen, J.; Cheng, L.; Wei, G.; Enge, M.; Taipale, M.; Vaquerizas, J.M.; Yan, J.; Sillanpää, M.J. Multiplexed massively parallel selex for characterization of human transcription factor binding specificities. Genome Res. 2010, 20, 861–873. [Google Scholar] [CrossRef]
- Slattery, M.; Riley, T.; Liu, P.; Abe, N.; Gomez-Alcala, P.; Dror, I.; Zhou, T.; Rohs, R.; Honig, B.; Bussemaker, H.J. Cofactor binding evokes latent differences in DNA binding specificity between hox proteins. Cell 2011, 147, 1270–1282. [Google Scholar] [CrossRef]
- Lou, X.; Qian, J.; Xiao, Y.; Viel, L.; Gerdon, A.E.; Lagally, E.T.; Atzberger, P.; Tarasow, T.M.; Heeger, A.J.; Soh, H.T. Micromagnetic selection of aptamers in microfluidic channels. Proc. Natl. Acad. Sci. USA 2009, 106, 2989–2994. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Lou, X.; Zhang, Y.; Xiao, Y.; Soh, H.T. Generation of highly specific aptamers via micromagnetic selection. Anal. Chem. 2009, 81, 5490–5495. [Google Scholar] [CrossRef]
- Oh, S.S.; Qian, J.; Lou, X.; Zhang, Y.; Xiao, Y.; Soh, H.T. Generation of highly specific aptamers via micromagnetic selection. Anal. Chem. 2011, 83, 1866. [Google Scholar] [CrossRef]
- Huang, C.-J.; Lin, H.-I.; Shiesh, S.-C.; Lee, G.-B. Integrated microfluidic system for rapid screening of crp aptamers utilizing systematic evolution of ligands by exponential enrichment (selex). Biosens. Bioelectron. 2010, 25, 1761–1766. [Google Scholar] [CrossRef]
- Cho, M.; Xiao, Y.; Nie, J.; Stewart, R.; Csordas, A.T.; Oh, S.S.; Thomson, J.A.; Soh, H.T. Quantitative selection of DNA aptamers through microfluidic selection and high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 15373–15378. [Google Scholar] [CrossRef] [Green Version]
- Mi, J.; Liu, Y.; Rabbani, Z.N.; Yang, Z.; Urban, J.H.; Sullenger, B.A.; Clary, B.M. In vivo selection of tumor-targeting RNA motifs. Nat. Chem. Biol. 2010, 6, 22. [Google Scholar] [CrossRef]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [PubMed]
- Savory, N.; Abe, K.; Sode, K.; Ikebukuro, K. Selection of DNA aptamer against prostate specific antigen using a genetic algorithm and application to sensing. Biosens. Bioelectron. 2010, 26, 1386–1391. [Google Scholar] [CrossRef]
- Nonaka, Y.; Yoshida, W.; Abe, K.; Ferri, S.; Schulze, H.; Bachmann, T.T.; Ikebukuro, K. Affinity improvement of a vegf aptamer by in silico maturation for a sensitive vegf-detection system. Anal. Chem. 2012, 85, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Savory, N.; Lednor, D.; Tsukakoshi, K.; Abe, K.; Yoshida, W.; Ferri, S.; Jones, B.V.; Ikebukuro, K. In silico maturation of binding-specificity of DNA aptamers against proteus mirabilis. Biotechnol. Bioeng. 2013, 110, 2573–2580. [Google Scholar] [CrossRef]
- Savory, N.; Takahashi, Y.; Tsukakoshi, K.; Hasegawa, H.; Takase, M.; Abe, K.; Yoshida, W.; Ferri, S.; Kumazawa, S.; Sode, K. Simultaneous improvement of specificity and affinity of aptamers against streptococcus mutans by in silico maturation for biosensor development. Biotechnol. Bioeng. 2014, 111, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Bowser, M.T. Isolation of DNA aptamers using micro free flow electrophoresis. Lab Chip 2011, 11, 3703–3709. [Google Scholar] [CrossRef]
- Hoon, S.; Zhou, B.; Janda, K.D.; Brenner, S.; Scolnick, J. Aptamer selection by high-throughput sequencing and informatic analysis. Biotechniques 2011, 51, 413–416. [Google Scholar] [CrossRef] [Green Version]
- Lauridsen, L.H.; Shamaileh, H.A.; Edwards, S.L.; Taran, E.; Veedu, R.N. Rapid one-step selection method for generating nucleic acid aptamers: Development of a DNA aptamer against α-bungarotoxin. PLoS ONE 2012, 7, e41702. [Google Scholar] [CrossRef]
- Thiel, W.H.; Bair, T.; Peek, A.S.; Liu, X.; Dassie, J.; Stockdale, K.R.; Behlke, M.A.; Miller Jr, F.J.; Giangrande, P.H. Rapid identification of cell-specific, internalizing RNA aptamers with bioinformatics analyses of a cell-based aptamer selection. PLoS ONE 2012, 7, e43836. [Google Scholar] [CrossRef]
- Stoltenburg, R.; Nikolaus, N.; Strehlitz, B. Capture-selex: Selection of DNA aptamers for aminoglycoside antibiotics. J. Anal. Methods Chem. 2012, 2012, 415697. [Google Scholar] [CrossRef] [PubMed]
- Waybrant, B.; Pearce, T.R.; Wang, P.; Sreevatsan, S.; Kokkoli, E. Development and characterization of an aptamer binding ligand of fractalkine using domain targeted selex. Chem. Commun. 2012, 48, 10043–10045. [Google Scholar] [CrossRef] [PubMed]
- WooáKim, D.; BockáGu, M. Immobilization-free screening of aptamers assisted by graphene oxide. Chem. Commun. 2012, 48, 2071–2073. [Google Scholar]
- Gong, Q.; Wang, J.; Ahmad, K.M.; Csordas, A.T.; Zhou, J.; Nie, J.; Stewart, R.; Thomson, J.A.; Rossi, J.J.; Soh, H.T. Selection strategy to generate aptamer pairs that bind to distinct sites on protein targets. Anal. Chem. 2012, 84, 5365–5371. [Google Scholar] [CrossRef]
- Szeto, K.; Latulippe, D.R.; Ozer, A.; Pagano, J.M.; White, B.S.; Shalloway, D.; Lis, J.T.; Craighead, H.G. Rapid-selex for RNA aptamers. PLoS ONE 2013, 8, e82667. [Google Scholar] [CrossRef]
- Kimoto, M.; Yamashige, R.; Matsunaga, K.-i.; Yokoyama, S.; Hirao, I. Generation of high-affinity DNA aptamers using an expanded genetic alphabet. Nat. Biotechnol. 2013, 31, 453. [Google Scholar] [CrossRef] [PubMed]
- Sefah, K.; Yang, Z.; Bradley, K.M.; Hoshika, S.; Jiménez, E.; Zhang, L.; Zhu, G.; Shanker, S.; Yu, F.; Turek, D. In vitro selection with artificial expanded genetic information systems. Proc. Natl. Acad. Sci. USA 2014, 111, 1449–1454. [Google Scholar] [CrossRef]
- Lao, Y.-H.; Chiang, H.-Y.; Yang, D.-K.; Peck, K.; Chen, L.-C. Selection of aptamers targeting the sialic acid receptor of hemagglutinin by epitope-specific selex. Chem. Commun. 2014, 50, 8719–8722. [Google Scholar] [CrossRef]
- Lai, J.-C.; Hong, C.-Y. Magnetic-assisted rapid aptamer selection (maras) for generating high-affinity DNA aptamer using rotating magnetic fields. ACS Comb. Sci. 2014, 16, 321–327. [Google Scholar] [CrossRef]
- Wang, J.; Gong, Q.; Maheshwari, N.; Eisenstein, M.; Arcila, M.L.; Kosik, K.S.; Soh, H.T. Particle display: A quantitative screening method for generating high-affinity aptamers. Angew. Chem. Int. Ed. 2014, 53, 4796–4801. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, Y.; Li, C.; Zou, Y.; Zhu, L.; An, Y.; Yang, C.J. Monoclonal surface display selex for simple, rapid, efficient, and cost-effective aptamer enrichment and identification. Anal. Chem. 2014, 86, 5881–5888. [Google Scholar] [CrossRef]
- Jacoby, K.; Scharenberg, A.M. Homing endonuclease target determination using selex adapted for yeast surface display. In Homing Endonucleases; Springer: Berlin/Heidelberg, Germany, 2014; pp. 165–190. [Google Scholar]
- Ouellet, E.; Foley, J.H.; Conway, E.M.; Haynes, C. Hi-fi selex: A high-fidelity digital-PCR based therapeutic aptamer discovery platform. Biotechnol. Bioeng. 2015, 112, 1506–1522. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G.; Tolle, F. Method of identifying or producing an aptamer. Google Patents WO2015049356A1, 19 October 2017. [Google Scholar]
- Tolle, F.; Brändle, G.M.; Matzner, D.; Mayer, G. A versatile approach towards nucleobase-modified aptamers. Angew. Chem. Int. Ed. 2015, 54, 10971–10974. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Sakota, E.; Nakamura, Y. The efficient cell-selex strategy, icell-selex, using isogenic cell lines for selection and counter-selection to generate RNA aptamers to cell surface proteins. Biochimie 2016, 131, 77–84. [Google Scholar] [CrossRef]
- Zumrut, H.E.; Ara, M.N.; Fraile, M.; Maio, G.; Mallikaratchy, P. Ligand-guided selection of target-specific aptamers: A screening technology for identifying specific aptamers against cell-surface proteins. Nucleic Acid Ther. 2016, 26, 190–198. [Google Scholar] [CrossRef]
- Yu, H.; Yang, W.; Alkhamis, O.; Canoura, J.; Yang, K.; Xiao, Y. In vitro isolation of small-molecule-binding aptamers with intrinsic dye-displacement functionality. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef]
- Damase, T.R.; Miura, T.A.; Parent, C.E.; Allen, P.B. Application of the open qPCR instrument for the in vitro selection of DNA aptamers against epidermal growth factor receptor and drosophila c virus. ACS Comb. Sci. 2018. [Google Scholar] [CrossRef]
- Kulbachinskiy, A. Methods for selection of aptamers to protein targets. Biochemistry 2007, 72, 1505–1518. [Google Scholar] [CrossRef]
- Greenleaf, W.J.; Frieda, K.L.; Foster, D.A.; Woodside, M.T.; Block, S.M. Direct observation of hierarchical folding in single riboswitch aptamers. Science 2008, 319, 630–633. [Google Scholar] [CrossRef]
- Gu, M.B.; Kim, H.-S. Biosensors Based on Aptamers and Enzymes; Springer: Berlin/Heidelberg, Germany, 2014; Volume 140. [Google Scholar]
- Huang, B.; Wang, W.; Bates, M.; Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science 2008, 319, 810–813. [Google Scholar] [CrossRef]
- Lippincott-Schwartz, J.; Patterson, G.H. Photoactivatable fluorescent proteins for diffraction-limited and super-resolution imaging. Trends Cell Biol. 2009, 19, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Senoner, M.; Unger, W.E. Sims imaging of the nanoworld: Applications in science and technology. J. Anal. At. Spectrom. 2012, 27, 1050–1068. [Google Scholar] [CrossRef]
- Newman, C.F.; Havelund, R.; Passarelli, M.K.; Marshall, P.S.; Francis, I.; West, A.; Alexander, M.R.; Gilmore, I.S.; Dollery, C.T. Intracellular drug uptake a comparison of single cell measurements using tof-sims imaging and quantification from cell populations with lc/ms/ms. Anal. Chem. 2017, 89, 11944–11953. [Google Scholar] [CrossRef]
- Huang, B.; Babcock, H.; Zhuang, X. Breaking the diffraction barrier: Super-resolution imaging of cells. Cell 2010, 143, 1047–1058. [Google Scholar] [CrossRef]
- Mairal, T.; Özalp, V.C.; Sánchez, P.L.; Mir, M.; Katakis, I.; O’Sullivan, C.K. Aptamers: Molecular tools for analytical applications. Anal. Bioanal. Chem. 2008, 390, 989–1007. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.Y.; Citartan, M.; Gopinath, S.C.; Tang, T.-H. Aptamers as a replacement for antibodies in enzyme-linked immunosorbent assay. Biosens. Bioelectron. 2015, 64, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yang, S. Replacing antibodies with aptamers in lateral flow immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef]
- Tan, W.; Wang, H.; Chen, Y.; Zhang, X.; Zhu, H.; Yang, C.; Yang, R.; Liu, C. Molecular aptamers for drug delivery. Trends Biotechnol. 2011, 29, 634–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, J.; Nilsen-Hamilton, M. Aptamers: Multifunctional molecules for biomedical research. J. Mol. Med. 2013, 91, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. Selex—a (r) evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 24, 381–403. [Google Scholar] [CrossRef]
- Fang, X.; Tan, W. Aptamers generated from cell-selex for molecular medicine: A chemical biology approach. Acc. Chem. Res. 2009, 43, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, V.J.; Levisson, M.; Eppink, M.H.; Smidt, H.; Van Der Oost, J. Alternative affinity tools: More attractive than antibodies? Biochem. J. 2011, 436, 1–13. [Google Scholar] [CrossRef]
- Lee, J.-O.; So, H.-M.; Jeon, E.-K.; Chang, H.; Won, K.; Kim, Y.H. Aptamers as molecular recognition elements for electrical nanobiosensors. Anal. Bioanal. Chem. 2008, 390, 1023–1032. [Google Scholar] [CrossRef]
- Kedzierski, S.; Khoshnejad, M.; Caltagirone, G.T. Synthetic antibodies: The emerging field of aptamers. Bioprocess. J. 2012, 11, 46–49. [Google Scholar] [CrossRef]
- McKeague, M.; DeRosa, M.C. Challenges and opportunities for small molecule aptamer development. J. Nucleic Acids 2012, 2012. [Google Scholar] [CrossRef]
- Que, E.L.; Domaille, D.W.; Chang, C.J. Metals in neurobiology: Probing their chemistry and biology with molecular imaging. Chem. Rev. 2008, 108, 1517–1549. [Google Scholar] [CrossRef] [PubMed]
- Keiser, M.J.; Setola, V.; Irwin, J.J.; Laggner, C.; Abbas, A.I.; Hufeisen, S.J.; Jensen, N.H.; Kuijer, M.B.; Matos, R.C.; Tran, T.B. Predicting new molecular targets for known drugs. Nature 2009, 462, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezovski, M.V.; Lechmann, M.; Musheev, M.U.; Mak, T.W.; Krylov, S.N. Aptamer-facilitated biomarker discovery (aptabid). J. Am. Chem. Soc. 2008, 130, 9137–9143. [Google Scholar] [CrossRef]
- Kalani, M.Y.S.; Theodore, N. Personalized medicine and neurosurgery. Front. Surg. 2017, 4, 19. [Google Scholar] [CrossRef]
- De Castro, M.A.G.; Höbartner, C.; Opazo, F. Aptamers provide superior stainings of cellular receptors studied under super-resolution microscopy. PLoS ONE 2017, 12, e0173050. [Google Scholar] [CrossRef]
- de Castro, M.A.; Rammner, B.; Opazo, F. Aptamer stainings for super-resolution microscopy. Methods Mol. Biol. 2016, 1380, 197–210. [Google Scholar]
- Smolyaninov, I.I.; Hung, Y.-J.; Davis, C.C. Magnifying superlens in the visible frequency range. Science 2007, 315, 1699–1701. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Babcock, H.P.; Zhuang, X. Dual-objective storm reveals three-dimensional filament organization in the actin cytoskeleton. Nat. Methods 2012, 9, 185–188. [Google Scholar] [CrossRef]
- Uhlén, M. Affinity as a tool in life science. Biotechniques 2008, 44, 649. [Google Scholar] [CrossRef] [PubMed]
- Ries, J.; Kaplan, C.; Platonova, E.; Eghlidi, H.; Ewers, H. A simple, versatile method for gfp-based super-resolution microscopy via nanobodies. Nat. Methods 2012, 9, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylova, M.; Cloin, B.M.; Finan, K.; Van Den Berg, R.; Teeuw, J.; Kijanka, M.M.; Sokolowski, M.; Katrukha, E.A.; Maidorn, M.; Opazo, F. Resolving bundled microtubules using anti-tubulin nanobodies. Nat. Commun. 2015, 6, 7933. [Google Scholar] [CrossRef] [PubMed]
- Opazo, F.; Levy, M.; Byrom, M.; Schäfer, C.; Geisler, C.; Groemer, T.W.; Ellington, A.D.; Rizzoli, S.O. Aptamers as potential tools for super-resolution microscopy. Nat. Methods 2012, 9, 938. [Google Scholar] [CrossRef]
- Ylera, F.; Lurz, R.; Erdmann, V.A.; Furste, J.P. Selection of RNA aptamers to the alzheimer’s disease amyloid peptide. Biochem. Biophys. Res. Commun. 2002, 290, 1583–1588. [Google Scholar] [CrossRef]
- Rahimi, F.; Murakami, K.; Summers, J.L.; Chen, C.H.; Bitan, G. RNA aptamers generated against oligomeric abeta40 recognize common amyloid aptatopes with low specificity but high sensitivity. PLoS ONE 2009, 4, e7694. [Google Scholar] [CrossRef]
- Janas, T.; Sapon, K.; Stowell, M.H.B.; Janas, T. Selection of membrane RNA aptamers to amyloid beta peptide: Implications for exosome-based antioxidant strategies. Int. J. Mol. Sci. 2019, 20, 299. [Google Scholar] [CrossRef]
- Mahmoud, M.; Deigner, H.-P. Aptamers in diagnostics: Replacing or complementing antibodies? J. Biochips Tissue Chips 2015, 5, 1. [Google Scholar] [CrossRef]
- Kanwar, J.R.; Roy, K.; Maremanda, N.G.; Subramanian, K.; Veedu, R.N.; Bawa, R.; Kanwar, R.K. Nucleic acid-based aptamers: Applications, development and clinical trials. Curr. Med. Chem. 2015, 22, 2539–2557. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Leslie, M.; Kameyama, H.; Volk, D.E.; Tanaka, T. Aptamer therapeutics in cancer: Current and future. Cancers 2018, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.L.; Nascimento, I.C.; Santos, A.P.; Ogusuku, I.E.Y.; Lameu, C.; Mayer, G.; Ulrich, H. Aptamers: Novelty tools for cancer biology. Oncotarget 2018, 9, 26934–26953. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.I.; Herrera, A.; Rossi, J.J.; Zhou, J. Current advances in aptamers for cancer diagnosis and therapy. Cancers 2018, 10, 9. [Google Scholar] [CrossRef]
- Hell, S.W. Microscopy and its focal switch. Nat. Methods 2009, 6, 24–32. [Google Scholar] [CrossRef]
- Harke, B.; Keller, J.; Ullal, C.K.; Westphal, V.; Schönle, A.; Hell, S.W. Resolution scaling in sted microscopy. Opt. Express 2008, 16, 4154–4162. [Google Scholar] [CrossRef]
- Chen, W.; Delaloye, S.; Silverman, D.H.; Geist, C.; Czernin, J.; Sayre, J.; Satyamurthy, N.; Pope, W.; Lai, A.; Phelps, M.E. Predicting treatment response of malignant gliomas to bevacizumab and irinotecan by imaging proliferation with [18f] fluorothymidine positron emission tomography: A pilot study. J. Clin. Oncol. 2007, 25, 4714–4721. [Google Scholar] [CrossRef]
- Sartori, A.; Gatz, R.; Beck, F.; Rigort, A.; Baumeister, W.; Plitzko, J.M. Correlative microscopy: Bridging the gap between fluorescence light microscopy and cryo-electron tomography. J. Struct. Biol. 2007, 160, 135–145. [Google Scholar] [CrossRef]
- Pavani, S.R.P.; Thompson, M.A.; Biteen, J.S.; Lord, S.J.; Liu, N.; Twieg, R.J.; Piestun, R.; Moerner, W. Three-dimensional, single-molecule fluorescence imaging beyond the diffraction limit by using a double-helix point spread function. Proc. Natl. Acad. Sci. USA 2009, 106, 2995–2999. [Google Scholar] [CrossRef] [Green Version]
- Holly, T.A.; Abbott, B.G.; Al-Mallah, M.; Calnon, D.A.; Cohen, M.C.; DiFilippo, F.P.; Ficaro, E.P.; Freeman, M.R.; Hendel, R.C.; Jain, D. Single photon-emission computed tomography. J. Nucl. Cardiol. 2010, 17, 941–973. [Google Scholar] [CrossRef] [PubMed]
- Rego, E.H.; Shao, L.; Macklin, J.J.; Winoto, L.; Johansson, G.A.; Kamps-Hughes, N.; Davidson, M.W.; Gustafsson, M.G. Nonlinear structured-illumination microscopy with a photoswitchable protein reveals cellular structures at 50-nm resolution. Proc. Natl. Acad. Sci. USA 2012, 109, E135–E143. [Google Scholar] [CrossRef] [PubMed]
- Rossy, J.; Cohen, E.; Gaus, K.; Owen, D.M. Method for co-cluster analysis in multichannel single-molecule localisation data. Histochem. Cell Biol. 2014, 141, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Vira, S.; Mekhedov, E.; Humphrey, G.; Blank, P.S. Fluorescent-labeled antibodies: Balancing functionality and degree of labeling. Anal. Biochem. 2010, 402, 146–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, M.; Huang, B.; Dempsey, G.T.; Zhuang, X. Multicolor super-resolution imaging with photo-switchable fluorescent probes. Science 2007, 317, 1749–1753. [Google Scholar] [CrossRef]
- Heilemann, M.; Van De Linde, S.; Schüttpelz, M.; Kasper, R.; Seefeldt, B.; Mukherjee, A.; Tinnefeld, P.; Sauer, M. Subdiffraction-resolution fluorescence imaging with conventional fluorescent probes. Angew. Chem. Int. Ed. 2008, 47, 6172–6176. [Google Scholar] [CrossRef] [PubMed]
- Granov, A.; Tiutin, L.; Schwarz, T. Positron Emission Tomography; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Zwan, M.D.; Bouwman, F.H.; Konijnenberg, E.; van der Flier, W.M.; Lammertsma, A.A.; Verhey, F.R.; Aalten, P.; van Berckel, B.N.; Scheltens, P. Diagnostic impact of [18F] flutemetamol pet in early-onset dementia. Alzheimers Res. 2017, 9, 2. [Google Scholar] [CrossRef]
- Marcus, C.; Mena, E.; Subramaniam, R.M. Brain pet in the diagnosis of alzheimer’s disease. Clin. Nucl. Med. 2014, 39, e413–e426. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.D.; Rabinovici, G.D.; Mathis, C.A.; Jagust, W.J.; Klunk, W.E.; Ikonomovic, M.D. Using pittsburgh compound b for in vivo pet imaging of fibrillar amyloid-beta. Adv. Pharm. 2012, 64, 27–81. [Google Scholar]
- Nickell, S.; Beck, F.; Korinek, A.; Mihalache, O.; Baumeister, W.; Plitzko, J.M. Automated cryoelectron microscopy of “single particles” applied to the 26s proteasome. Febs Lett. 2007, 581, 2751–2756. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Hirokawa, N.; Kikkawa, M. Three-dimensional structures of the flagellar dynein–microtubule complex by cryoelectron microscopy. J. Cell Biol. 2007, 177, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Hoenger, A. High-resolution cryo-electron microscopy on macromolecular complexes and cell organelles. Protoplasma 2014, 251, 417–427. [Google Scholar] [CrossRef]
- Yan, Q.; Cai, M.; Zhou, L.; Xu, H.; Shi, Y.; Sun, J.; Jiang, J.; Gao, J.; Wang, H. Using an RNA aptamer probe for super-resolution imaging of native egfr. Nanoscale Adv. 2019, 1, 291–298. [Google Scholar] [CrossRef]
- Fornasiero, E.F.; Opazo, F. Super-resolution imaging for cell biologists. Bioessays 2015, 37, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.Y.; Lee, T.S.; Song, I.H.; Cho, Y.L.; Chae, J.R.; Kang, H.; Lim, J.H.; Lee, J.H.; Kang, W.J. Pet imaging of her2 expression with an 18f-fluoride labeled aptamer. PLoS ONE 2019, 14, e0211047. [Google Scholar] [CrossRef]
- Hassanzadeh, L.; Chen, S.; Veedu, R.N. Radiolabeling of nucleic acid aptamers for highly sensitive disease-specific molecular imaging. Pharmaceuticals 2018, 11, 106. [Google Scholar] [CrossRef]
- Serrano-Santos, M.B.; Llobet, E.; Özalp, V.C.; Schäfer, T. Characterization of structural changes in aptamer films for controlled release nanodevices. Chem. Commun. 2012, 48, 10087–10089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, G. The Chemical Biology of Nucleic Acids. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9780470664001 (accessed on 14 May 2010).
- Stanlis, K.K.; McIntosh, J.R. Single-strand DNA aptamers as probes for protein localization in cells. J Histochem. Cytochem. 2003, 51, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Schermelleh, L.; Heintzmann, R.; Leonhardt, H. A guide to super-resolution fluorescence microscopy. J. Cell Biol. 2010, 190, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempsey, G.T.; Vaughan, J.C.; Chen, K.H.; Bates, M.; Zhuang, X. Evaluation of fluorophores for optimal performance in localization-based super-resolution imaging. Nat. Methods 2011, 8, 1027–1036. [Google Scholar] [CrossRef]
- Lukinavicius, G.; Mitronova, G.Y.; Schnorrenberg, S.; Butkevich, A.N.; Barthel, H.; Belov, V.N.; Hell, S.W. Fluorescent dyes and probes for super-resolution microscopy of microtubules and tracheoles in living cells and tissues. Chem. Sci. 2018, 9, 3324–3334. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.Y.; Byun, J. Nucleic acid aptamers: New methods for selection, stabilization, and application in biomedical science. Biomol. Ther. 2013, 21, 423. [Google Scholar] [CrossRef]
- Ozer, A.; Pagano, J.M.; Lis, J.T. New technologies provide quantum changes in the scale, speed, and success of selex methods and aptamer characterization. Mol. Ther. Nucleic Acids 2014, 3, e183. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.A. In Vivo Selex Strategies. Handbook of RNA Biochemistry: Second, Completely Revised and Enlarged Edition. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9783527647064 (accessed on 28 March 2014).
- Szeto, K.; Craighead, H.G. Devices and approaches for generating specific high-affinity nucleic acid aptamers. Appl. Phys. Rev. 2014, 1, 031103. [Google Scholar] [CrossRef] [Green Version]
- Tombelli, S.; Minunni, M.; Mascini, M. Aptamers-based assays for diagnostics, environmental and food analysis. Biomol. Eng. 2007, 24, 191–200. [Google Scholar] [CrossRef] [PubMed]
- White, R.J.; Phares, N.; Lubin, A.A.; Xiao, Y.; Plaxco, K.W. Optimization of electrochemical aptamer-based sensors via optimization of probe packing density and surface chemistry. Langmuir 2008, 24, 10513–10518. [Google Scholar] [CrossRef]
- Nair, P.R.; Alam, M.A. Theory of "selectivity" of label-free nanobiosensors: A geometro-physical perspective. J. Appl. Phys. 2010, 107, 64701. [Google Scholar] [CrossRef]
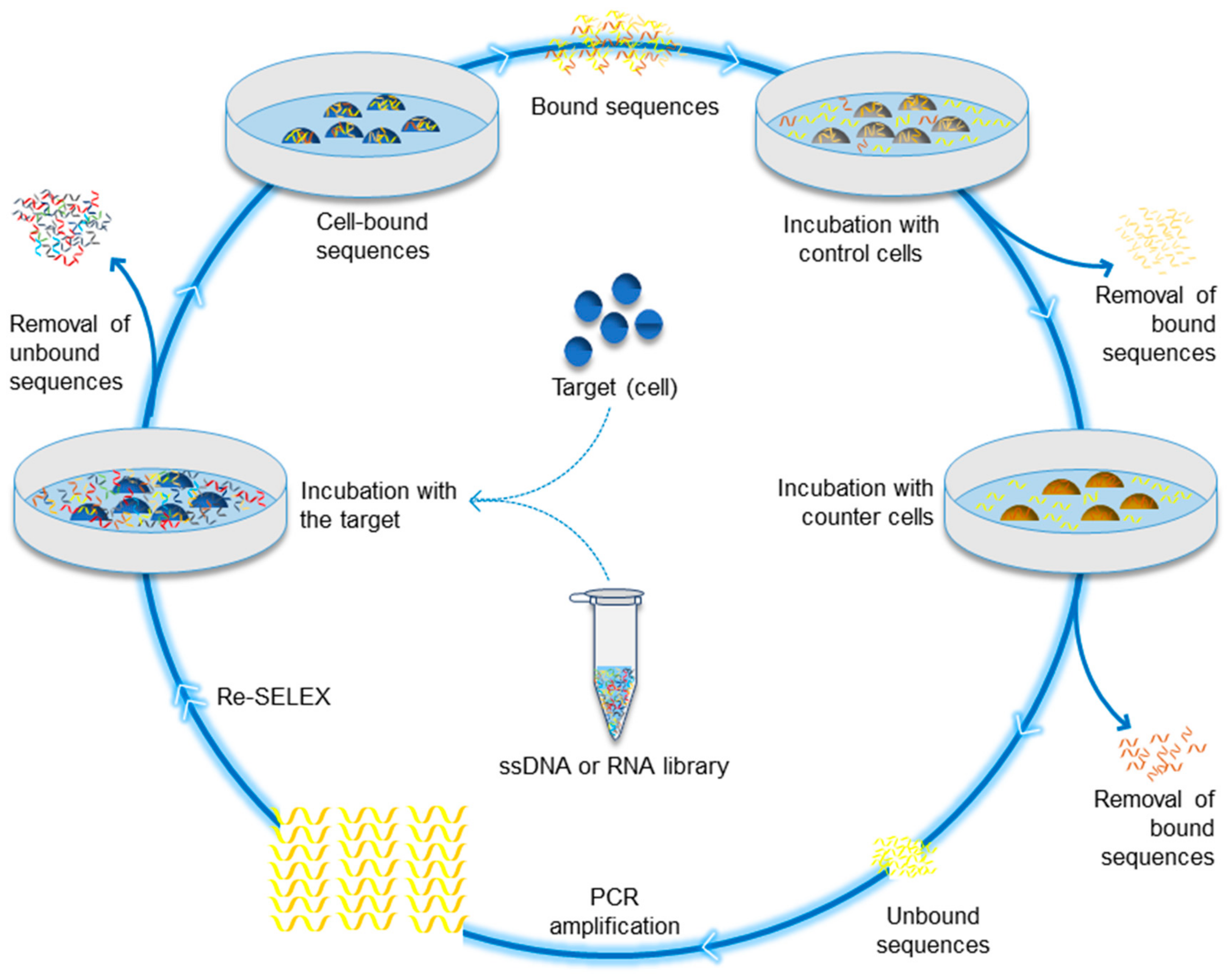
Figure 1.
Cell-SELEX aptamer selection method. It consists of three major steps that can be interspersed with sub-steps: a. preparation of the ssDNA or RNA library, b. incubation of the target molecules with the library as a positive selection step, and c. amplification of the selected and recovered strands. Some sub-steps can be performed, whether before or after the positive selection, such as incubation with counter cells in order to remove the non-specific binding and enhance the selection process.
Figure 1.
Cell-SELEX aptamer selection method. It consists of three major steps that can be interspersed with sub-steps: a. preparation of the ssDNA or RNA library, b. incubation of the target molecules with the library as a positive selection step, and c. amplification of the selected and recovered strands. Some sub-steps can be performed, whether before or after the positive selection, such as incubation with counter cells in order to remove the non-specific binding and enhance the selection process.


{kind=link}
Table 1.
Important strides in SELEX research.
| SELEX Type | Principle | Aim | Application/Result | Year | Ref. |
|---|---|---|---|---|---|
| Negative SELEX | In vitro selection of single-stranded oligonucleotides against a target after removing sequences that bind to partitioning/immobilization matrices |
| Isolated ligand-specific aptamers that selectively bind to small molecules (dyes) | 1992 | [21] |
| In vivo SELEX | The selection and amplification steps occur inside the living cell using retroviral-based replication system | Produce aptamers that are functional in vivo | Provided a method to transfer aptamer selection and amplification from in vitro to in vivo | 1993 | [22,23] |
| Counter SELEX | Uses a second elution step against a molecule of similar target structure (e.g., caffeine, which differs from theophylline at the N-7 position) | Increase aptamer’s specificity and affinity towards target molecules | Isolated aptamer of high-specificity to theophylline | 1994 | [24] |
| cDNA-SELEX | Uses a preselected natural oligonucleotide pool that binds to the protein of interest rather than using a synthetic library | Identify natural library that interact with the protein of interest | Provided a new approach to study the interaction between the nucleotides and the protein of interest | 1995 | [25] |
| Blended SELEX | Forming a blended-pool through linking molecules (e.g., inhibitor) to a specific site on the library that will be subjected to SELEX |
|
| 1995 | [26] |
| Uses photoactivatable aptamers to crosslink and/or photoactive the target |
|
| 1995 2000 | [27,28] |
| Spiegelmer® | Selection of D-oligonucleotides against a mirrored target. Then, the D-oligonucleotides will be synthesized as L-oligonucleotides that can bind to unmirrored target | Increase aptamer stability against nucleases | Identified an endonuclease-stable L-RNAaptamer that binds to D-adenosine | 1996 | [29] |
| Magnetic bead-based SELEX | Uses magnetic beads for immobilization |
| Provided a new method to avoid DNA precipitation | 1997 | [30] |
| EMSA-SELEX | Uses EMSA partitioning at every cycle of the selection process | Identify a method to study oligonucleotides binding to proteins | Identified the role of specific zinc finger motifs in the Roaz protein that help in DNA‒protein binding and protein‒protein interaction | 1998 | [31] |
| Cell-SELEX | Identifies aptamers that bind specifically to different cells or targets on cell membrane |
| Identified aptamers that binds to specific cell (e.g., RBCs) or cell surface protein (e.g., variant surface glycoprotein of Trypanosoma brucei) | 1998 1999 | [32,33,34] |
| Chimeric SELEX | Fuses pre-selected aptamers of different targets together to form a combined small library followed by applying a dual selection pressure to select aptamers that can bind to multiple targets | Generate dual-function aptamers | Generated an engineered dual-function aptamer capable of testing/binding to two distinct features/targets | 1998 | [35] |
| Multi-stage SELEX | Modified chimeric-based SELEX followed by an extra selection with entirety of targets | Develop an allosteric synthetic aptamer | Provided a method to study the allosteric interactions in the DNA | 1999 | [36] |
| Indirect selection | Aptamer binds to a “primary target.” Binding to this target is required for aptamer bind ing to the main target | Develop a transition-state-bound aptamer that facilitates its binding to the main target | Generated Zn2+-dependent aptamer molecule for specific target binding to HIV-1 Tat protein | 2000 | [37] |
| Toggle-SELEX | A “polyclonal” aptamer selection strategy that generates aptamer capable of binding to multiple proteins through incubating the pre-selected aptamer with a second target until the aptamer can identify another region for binding | Select aptamers against homologous targets depending on cross-reactivity |
| 2001 | [38] |
| Truncates fixed regions present in the aptamers sequences that may limit the structure variations or reduce the ability of aptamer binding to the target |
| Provided a method to control aptamer-fixed sequences | 2001 2004 2008 2010 | [39,40,41,42] |
| Expression Cassette SELEX | Fuses a pre-selected aptamer with a polymerase III promoter “expression cassette” in a DNA plasmid | Produce highly expressed levels of functional aptamers |
| 2002 | [43] |
| NECEEM | Uses non-equilibrium conditions to separate the constituents of protein‒DNA mixture (free and bound components) |
| Provided a novel electrophoretic method for studying protein‒DNA interaction and Kd of the DNA‒protein complex | 2002 | [44] |
| Subtractive SELEX | Similar to counter SELEX where it adopts extra SELEX rounds to control target binding | Distinguish cell subtypes of homologous origin | Distinguished between differentiated and normal PC12 cells | 2003 | [45] |
| Tailored SELEX | Isolation of short aptamer through designing of cleavable sites to remove fixed sequences |
| Provided a new method to identify short fixed nucleotides aptamers through ligation and removal of primer | 2003 | [46] |
| CE-SELEX | Uses differential electrophoretic separation at every SELEX round after aptamer binds to the target |
| Standardized the SELEX method and made it more efficient | 2004 | [47] |
| SweepCE | Uses protein non-stopped flow in the capillary electrophoresis to form DNA‒protein complex that causes sweeping of the DNA |
| Provided a method that help in studying the equilibrium and kinetic parameters during protein‒DNA complex formation | 2004 | [48] |
| On-chip selection | On-chip selection method combined with point-mutation of the sequence and usage of genetic algorithm to produce aptamers | Generate different aptamers against different target molecules | Generated aptamers against resorufin | 2004 | [49] |
| ECEEM | Maintains the equilibrium of the aptamer-target while separating the components of the equilibrium mixture by capillary electrophoresis |
| Produced “smart aptamers” with a predefined Kd value matching the theoretically predicted value | 2005 | [50] |
| FluMag-SELEX | Magnetic beads-based method using fluorescent labelling for monitoring the affinity and complex separation |
|
| 2005 | [51] |
| Non-SELEX | NECEEM-based method in which the PCR amplification step is skipped and the recovered target‒DNA complex is incubated with fresh target followed by partitioning |
| Developed a technique to reduce the selection time while producing aptamers of a 4-fold affinity improvement over the CE-SELEX | 2006 | [52] |
| SPR-SELEX | A method that couples the aptamer or the target to a chip then immobilizes the target or the aptamer and measures the change at the surface upon aptamer-target binding |
| Provided a method to measure the aptamer’s real-time binding | 2006 | [53,54] |
| TECS-SELEX | Uses modified cells expressing recombinant form of a surface protein that eliminates the need for the purification step | Produce aptamer against ectopic cell surface protein | Produced aptamer against TGF-β type III receptor that is ectopically expressed on CHO cells | 2006 | [55] |
|
| Provide a method to study in vivo oligonucleotide‒protein binding |
| 2006 2012 | [56,57] |
| DeSELEX and Convergent selection | Complex SELEX methods where many proteins targets are incubated with the library and then deselecting the dominant-protein bound aptamer and redirecting the selection toward the less abundant protein/s | Shift the selection process toward a specific protein (even a less abundant one) in mixture of different protein complexes |
| 2007 | [58] |
| MonoLEX |
|
|
| 2007 | [59] |
| NanoSelection® (nM-AFM SELEX) |
|
| Developed a method to avoid multiple selection cycles and also select aptamers from small library | 2007 | [60] |
| ASExp | Uses quenching and dequenching of aptamers | Rapid selection method | Provided a method to rapidly select aptamer for different types of targets | 2008 | [61] |
| FACS-SELEX | Combines in vitro selection with FACS to separate specific cell population |
| Produced aptamer bound to vital Burkitt lymphoma cells | 2008 | [62,63] |
| Single microbead SELEX | Reduces the amount of the target molecules and expose them to a single microbead | Produce high affinity aptamer using a single target-conjugated microbead |
| 2008 | [64] |
| CLADE | In silico aptamer selection method | Rapid aptamer designing method | Identified aptamers against allophycocyanin in silico | 2008 | [65] |
| In silico selection | In silico secondary structure-based selection followed by a 3D structure prediction and HTP screening of the selected aptamers |
| Provided a computational approach to select aptamers of high-affinity | 2009 | [66] |
| Tissue slide-based SELEX | In situ SELEX method against paraffin tissue sections | Target clinical tumour markers and provide a way to differentiate between the clinical specimens |
| 2009 | [67] |
| Sol–gel SELEX |
| Produce faster, HTP, efficient, and cheap aptamer |
| 2009 | [68] |
|
|
|
| 2009 2010 2011 | [69,70,71] |
| M-SELEX | Target‒DNA sample separation is obtained through applying a high voltage |
| Provided a fast tool for screening aptamers against any targets | 2009 | [72,73,74,75] |
| QSAS | Combines M-SELEX with HTS for efficient and rapid aptamer production | Generate aptamer with high efficiency, high affinity, and high specificity | Introduced an integrated method for rapid, high-affinity, and high-specificity generation of aptamer | 2010 | [76] |
| Cellular-uptake in vivo-variant | Intravenous injection of modified random library into cancerous animal model | Select aptamer for internal target inside the organism depending on cellular uptake | Provided a unique approach to generate aptamer that can specifically localize to tumour cells in vivo | 2010 | [77] |
| SOMAmers |
|
|
| 2010 | [78] |
| ISM | In silico post-SELEX genetic algorithm performed to identify aptamers of high-binding affinity | Improve aptamers binding-affinity and specificity | Selected aptamers of high binding affinity and specificity to prostate specific antigen, VEGF, and Proteus mirabilis | 2010 2012 2013 2014 | [79,80,81,82] |
| μFFE | Applies electric field separation to overcome the limitations of CE-SELEX to separate bound from unbound sequences |
| Improved the library size by a 300-fold over what CE-SELEX has achieved | 2011 | [83] |
| HTS-SELEX | One round of positive selection followed by HTS and informatic analysis |
| Identified aptamers that can bind to thrombin in nM range | 2011 | [84] |
| One-step selection Method | One-step selection of target-immobilized coverslip followed by library binding, extensive washing, and amplification step |
| Provided a rapid one-step generation of aptamer against α-bungarotoxin | 2012 | [85] |
| Cell-internalization SELEX | Combines cell-SELEX with HTS and bioinformatics | Produce aptamers of high intracellular internalization | Provided a new approach to design aptamers that capable of internalizations into the cytoplasm of vascular smooth muscle cells | 2012 | [86] |
| Capture-SELEX (FluMag-based) | Immobilization of specific aptamers library on magnetic beads using a docking sequence linked to the library |
| Provided a method to select aptamer against small molecule (e.g., kanamycin A) | 2012 | [87] |
| Domain targeted SELEX | Uses recombinant protein containing chemokine domain (to extend protein accessibility) immobilized onto magnetic agarose beads | Select aptamer against specific protein’s unfolded domain | Developed aptamer to Fractalkine protein | 2012 | [88] |
| Uses π-π stacking and adsorption of ssDNA on 2D GO sheets to separate unbound DNA | Generation of high-affinity, cost-effective, and target-immobilization-free SELEX | Produced aptamer to Nampt protein without target immobilization | 2012 | [89] |
| MAI-SELEX | A selection method to recognize two separate sites on the target using 2’-fluoro-modified library | Target distinct sites/subunits of the protein | Produced two aptamers that recognize the αV or β3 subunits of integrin αVβ3 | 2012 | [90] |
| RAPID-SELEX |
| Reduce selection time | Identified aptamer to CHK2 and UBLCP1 in one-third of the time required for the conventional selection | 2013 | [91] |
| Expanded genetic alphabet (Ds-base) | Uses a library containing natural nucleotides and other three unnatural nucleotides with a hydrophobic base | Increase sequence diversity and so expand SELEX selection power | Generated aptamers that bind to VEGF165 and interferon-γ with more than 100-fold improved affinity | 2013 | [92] |
| AEGIS-SELEX | Uses unnatural forms of nucleotides, nonstandard P and Z, beside the normal four bases-based library (GACTZP DNA library) |
| Produced ZAP-2012 aptamer that can bind to MDA-MB-231 with 10-fold higher affinity than achieved using the conventional aptamers | 2014 | [93] |
| ES-SELEX |
| Generate epitope-specific aptamer of a native protein structure | Produced anti-sialic acid receptor aptamers that inhibit hemagglutination at pM range | 2014 | [94] |
| MARAS | Selects aptamers with different affinities to the target molecule using magnetic beads and external rotating magnetic field | Select aptamers based on their different binding affinities | Produced aptamer with high affinity to C-reactive protein | 2014 | [95] |
| Particle Display | Measures the affinity of each aptamer sequence in the library and sorting them using HTS. Then isolating the highest-affinities candidates using FACS | Generate high-affinity, simple, rapid, and cost-effective aptamers | Measured the affinity of 100 million aptamers and obtained high-affinity aptamers for thrombin, ApoE, PAI-1, and 4-1BB proteins | 2014 | [96] |
| MSD-SELEX | Uses a library of monoclonal DNA-displaying beads | Generate high-affinity and rapid aptamers | Obtained high-affinity aptamers against EpCAM and aflatoxin B1 | 2014 | [97] |
| Yeast surface display-SELEX | Target protein is loaded on the surface of yeast | Quick and unexpansive HTS identification of DNA-binding sites of the proteins without prior knowledge of the target site | Provided a way to determine aptamer‒protein binding specificity | 2014 | [98] |
| Hi-Fi SELEX | Introduces fixed-region blocking elements to enhance the functional diversity of the library |
| Identified aptamers of Kd in nM range to α-thrombin | 2015 | [99] |
| Click-SELEX | Uses copper(I)-catalysed alkyne–azide cycloaddition modified nucleic acid libraries | Generate modified nucleobases that increase binding capabilities |
| 2015 | [100,101] |
| Icell SELEX |
| Target various membrane proteins through successful manipulation of the endogenous expression of the target proteins | Produced aptamer to integrin alpha V | 2016 | [102] |
| LIGS | Uses antibody bound to antigen for the partitioning step to compete with aptamers from partially enriched SELEX | Identify highly-specific aptamer sequences that outcompeting with specific antibody for selecting highly specific aptamers | Identified three aptamers that outcompete with mIgM antibody | 2016 | [103] |
| Colorimetric dye-displacement SELEX | Uses target-induced displacement of a small-molecule dye to isolate signal-producing aptamers triggered by binding to the target | Develop an efficient and rapid way to isolate small molecules through colorimetric dye-displacement | Provided a new way of sensitive target isolation based on Cy7-displacement colorimetric assay | 2018 | [104] |
| Open qPCR SELEX | Uses open qPCR to quantify target-aptamer binding |
| Produced aptamers against whole Drosophila C virus particles | 2018 | [105] |
Abbreviations: AEGIS-SELEX, artificially expanded genetic information systems SELEX; AFM, atomic force microscopy; ASExp, aptamer selection express; CE-SELEX, capillary electrophoresis SELEX; CLADE, closed loop aptameric directed evolution; ECEEM, equilibrium capillary electrophoresis of equilibrium mixtures; EMSA-SELEX, electrophoretic mobility shift assay SELEX; ES-SELEX, epitope-specific SELEX; FACS-SELEX, fluorescence-activated cell sorting SELEX; GO-SELEX, graphene oxide SELEX; Hi-Fi SELEX, high-fidelity SELEX; HTS, high-throughput sequencing; HTP, high-throughput; Icell SELEX, isogenic cell SELEX; ISM, in silico maturation; LIGS, ligand-guided selection; MAI-SELEX, multivalent aptamer isolation; MARAS, magnetic-assisted rapid aptamer selection; μFFE, micro free-flow electrophoresis; MP, minimal primer selection; MSD-SELEX, monoclonal surface display SELEX; M-SELEX, microfluidics SELEX; NECEEM, non-equilibrium capillary electrophoresis of equilibrium mixtures; PF, primer-free selection; QSAS, quantitative selection of aptamers through sequencing; RAPID-SELEX, RNA aptamer isolation via dual-cycles SELEX; SELEX, systematic evolution of ligands by exponential enrichment; SOMAmers, slow off-rate modified aptamer; SPR, surface plasmon resonance; SweepCE, sweeping capillary electrophoresis; TECS-SELEX, target expressed on cell surface SELEX.
Table 2.
Comparison between aptamers and antibodies.
| Aptamer | Antibodies | |
|---|---|---|
| Molecular weight | Small (~12–30 kDa) | Relatively big (~150–180 kDa) |
| Secondary structures | Various structures: hairpin, loop, G-quadruplex, etc | β-sheets |
| Generation time | Few hours to months | Several months (~six months) |
| Batches variations | Low | High |
| Immunogenicity | Low | High |
| Minimal target size | Targets small sizes ~60 Da | ~600 Da |
| Targets | Wide range of targets | Immunogenic molecules |
| Shelf life | Long | Short |
| Allowed chemical modifications | Various modifications | Limited modifications |
| Nuclease degradation | Sensitive | Resistant |
| In vivo half-life | Short (~20 min) | Long (~one month) |
| Stability | Very stable | Sensitive to temperature and pH changes |
| Cost | Lower | Higher |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ali, M.H.; Elsherbiny, M.E.; Emara, M. Updates on Aptamer Research. Int. J. Mol. Sci. 2019, 20, 2511. https://doi.org/10.3390/ijms20102511
AMA Style
Ali MH, Elsherbiny ME, Emara M. Updates on Aptamer Research. International Journal of Molecular Sciences. 2019; 20(10):2511. https://doi.org/10.3390/ijms20102511
Chicago/Turabian StyleAli, Mohamed H., Marwa E. Elsherbiny, and Marwan Emara. 2019. "Updates on Aptamer Research" International Journal of Molecular Sciences 20, no. 10: 2511. https://doi.org/10.3390/ijms20102511
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.