Activation of Human Peripheral Basophils in Response to High IgE Antibody Concentrations without Antigens

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Concentration of IgE Antibodies in Serum of Healthy Donors and Patients with Skin Allergic Disorders
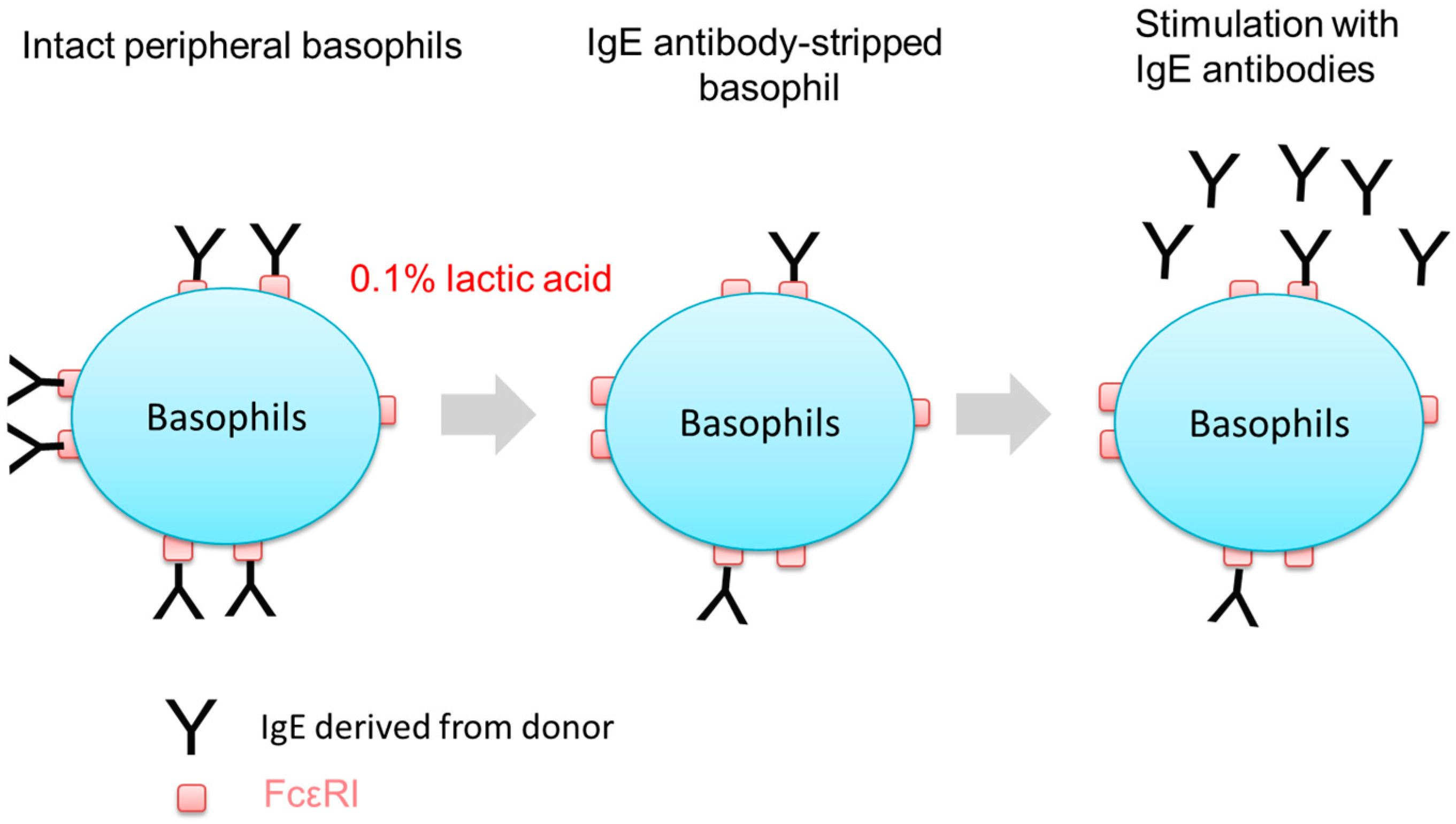
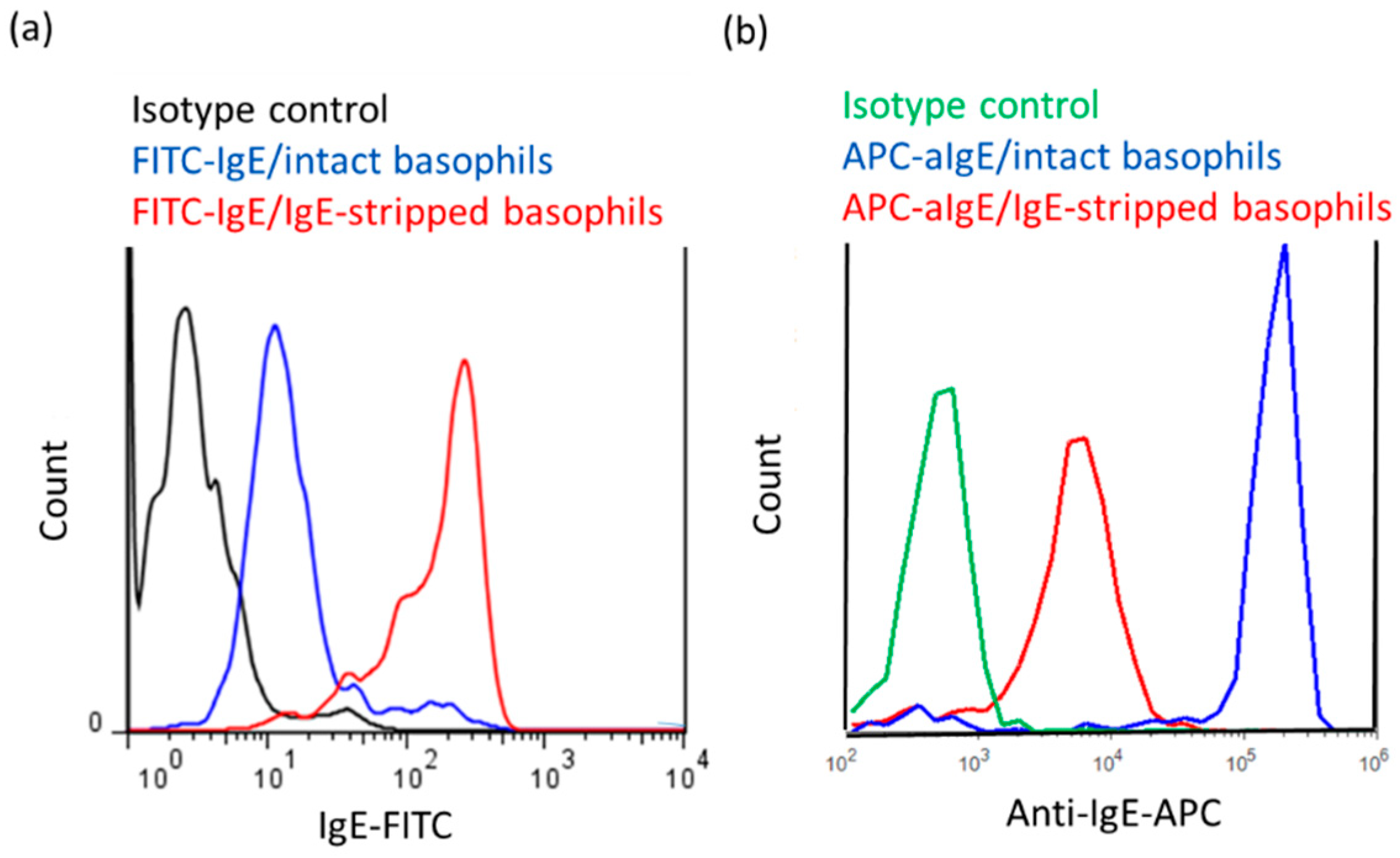
2.2. Stripping of IgE Antibodies Bound to FcεRIs on the Surface of Human Peripheral Basophils
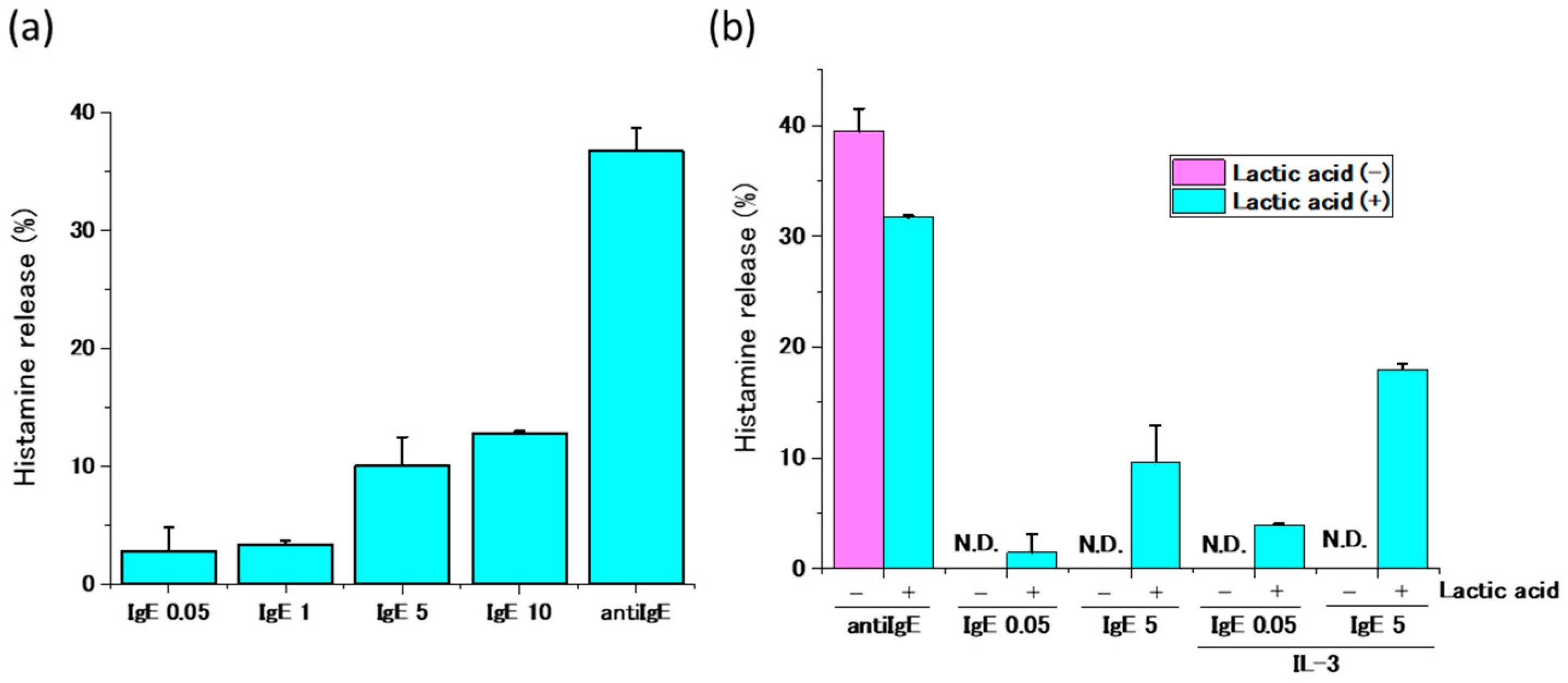
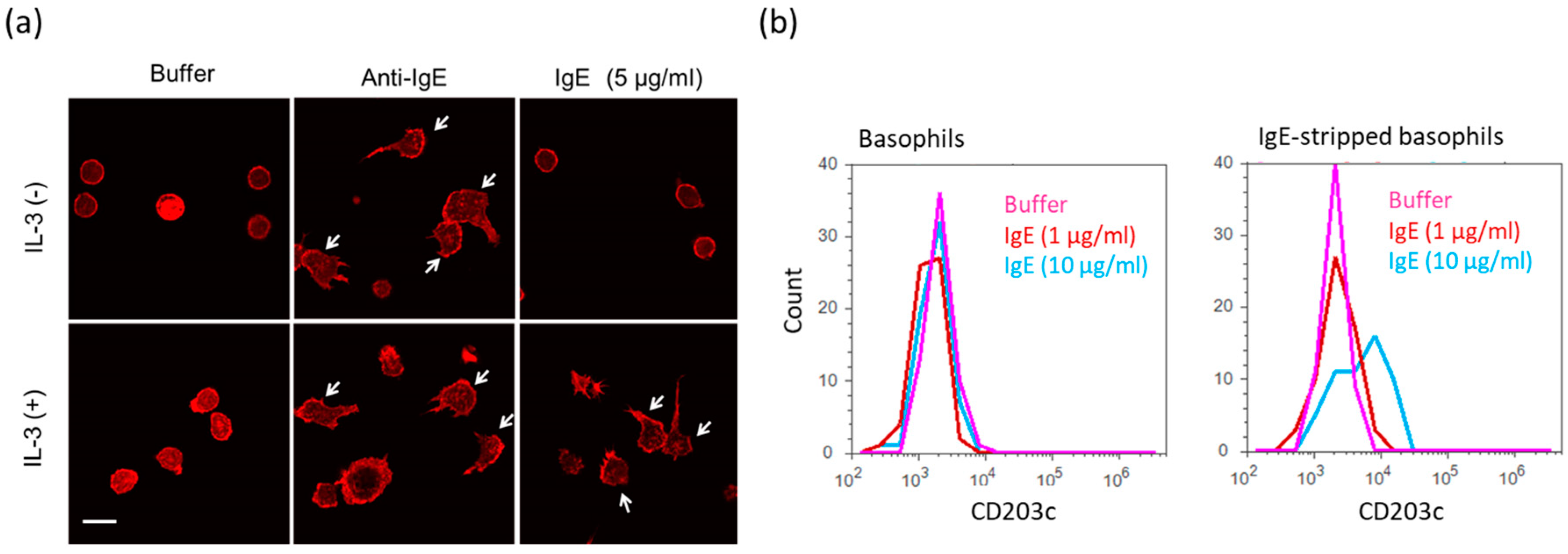
2.3. Activation of Human Peripheral Basophils in Response to Human IgE Antibodies
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Isolation of Peripheral Blood Mononuclear Cells (PBMCs) and Basophils from Human Peripheral Blood
4.3. Detection of IgE Antibodies in Serum
4.4. Labeling of IgE Antibodies with FITC
4.5. Removal of IgEs on the Surface of Basophils
4.6. Detection of IgE-free FcεRI, IgE, Total FcεRI and CD203c on the Surface of Basophils by Means of Flow Cytometry
4.7. Histamine Release Test
4.8. Staining of Actin Cytoskeleton and Observation of Morphology of Human Peripheral Basophils
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FcεRI | high-affinity IgE receptors |
| CSU | chronic spontaneous urticaria |
| AD | atopic dermatitis |
| PBMC | peripheral blood mononuclear cells |
| IL | interleukin |
| IgE | imunoglobulin E |
References
- Kubo, M. Mast cells and basophils in allergic inflammation. Curr. Opin. Immunol. 2018, 54, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Kurotaki, D.; Tamura, T. Regulation of basophil and mast cell development by transcription factors. Allergol. Int. 2016, 65, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J. Mast cells and basophils. Curr. Opin. Hematol. 2000, 7, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Raap, U.; Rivellese, F.; Marone, G.; Gibbs, B.F. Human mast cells and basophils-How are they similar how are they different? Immunol. Rev. 2018, 282, 8–34. [Google Scholar] [CrossRef] [PubMed]
- Hirai, K.; Morita, Y.; Misaki, Y.; Ohta, K.; Takaishi, T.; Suzuki, S.; Motoyoshi, K.; Miyamoto, T. Modulation of human basophil histamine release by hemopoietic growth factors. J. Immunol. 1988, 141, 3958–3964. [Google Scholar] [PubMed]
- Yanase, Y.; Hide, I.; Mihara, S.; Shirai, Y.; Saito, N.; Nakata, Y.; Hide, M.; Sakai, N. A critical role of conventional protein kinase C in morphological changes of rodent mast cells. Immunol. Cell Biol. 2011, 89, 149–159. [Google Scholar] [CrossRef]
- Pandey, V.; Mihara, S.; Fensome-Green, A.; Bolsover, S.; Cockcroft, S. Monomeric IgE stimulates NFAT translocation into the nucleus, a rise in cytosol Ca2+, degranulation, and membrane ruffling in the cultured rat basophilic leukemia-2H3 mast cell line. J. Immunol. 2004, 172, 4048–4058. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, J.; Kinoshita, T.; Matsumoto, M.; Chung, S.; Kawakami, Y.; Leitges, M.; Wu, D.; Lowell, C.A.; Kawakami, T. IgE- and IgE+Ag-mediated mast cell migration in an autocrine/paracrine fashion. Blood 2005, 105, 3222–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwakura, J.; Xiao, W.; Kitaura, J.; Kawakami, Y.; Maeda-Yamamoto, M.; Pfeiffer, J.R.; Wilson, B.S.; Blank, U.; Kawakami, T. Pivotal advance: IgE accelerates in vitro development of mast cells and modifies their phenotype. J. Leukoc. Biol. 2008, 84, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Kasperkiewicz, M.; Schmidt, E.; Ludwig, R.J.; Zillikens, D. Targeting IgE Antibodies by Immunoadsorption in Atopic Dermatitis. Front. Immunol. 2018, 9, 254. [Google Scholar] [CrossRef] [PubMed]
- Kessel, A.; Helou, W.; Bamberger, E.; Sabo, E.; Nusem, D.; Panassof, J.; Toubi, E. Elevated serum total IgE--a potential marker for severe chronic urticaria. Int. Arch. Allergy Immunol. 2010, 153, 288–293. [Google Scholar] [CrossRef]
- Youssef, L.A.; Schuyler, M.; Wilson, B.S.; Oliver, J.M. Roles for the High Affinity IgE Receptor, FcεRI, of Human Basophils in the Pathogenesis and Therapy of Allergic Asthma: Disease Promotion, Protection or Both? Open Allergy J. 2010, 3, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Larenas-Linnemann, D.E.S.; Parisi, C.A.S.; Ritchie, C.; Cardona-Villa, R.; Cherrez-Ojeda, I.; Cherrez, A.; Ensina, L.F.; Garcia, E.; Medina, I.V.; Rodríguez-González, M.; et al. Update on Omalizumab for Urticaria: What’s New in the Literature from Mechanisms to Clinic. Curr. Allergy Asthma Rep. 2018, 18, 33. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Goodarzi, H.; Chen, H.Y. IgE, mast cells, and eosinophils in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Metz, M.; Staubach, P.; Bauer, A.; Brehler, R.; Gericke, J.; Kangas, M.; Ashton-Chess, J.; Jarvis, P.; Georgiou, P.; Canvin, J.; et al. Clinical efficacy of omalizumab in chronic spontaneous urticaria is associated with a reduction of FcεRI-positive cells in the skin. Theranostics 2017, 7, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Yanase, Y.; Hiragun, T.; Yanase, T.; Kawaguchi, T.; Ishii, K.; Hide, M. Evaluation of peripheral blood basophil activation by means of surface plasmon resonance imaging. Biosens. Bioelectron. 2012, 32, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Hide, M.; Tanaka, T.; Yamamura, Y.; Koro, O.; Yamamoto, S. IgE-mediated hypersensitivity against human sweat antigen in patients with atopic dermatitis. Acta Derm.-Venereol. 2002, 82, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Yanase, Y.; Irifuku, R.; Ishii, K.; Kawaguchi, T.; Takahagi, S.; Hide, I.; Hide, M. The role of adenosine for IgE receptor-dependent degranulation of human peripheral basophils and skin mast cells. Allergol. Int. 2018, 67, 524–526. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanase, Y.; Matsuo, Y.; Kawaguchi, T.; Ishii, K.; Tanaka, A.; Iwamoto, K.; Takahagi, S.; Hide, M. Activation of Human Peripheral Basophils in Response to High IgE Antibody Concentrations without Antigens. Int. J. Mol. Sci. 2019, 20, 45. https://doi.org/10.3390/ijms20010045
Yanase Y, Matsuo Y, Kawaguchi T, Ishii K, Tanaka A, Iwamoto K, Takahagi S, Hide M. Activation of Human Peripheral Basophils in Response to High IgE Antibody Concentrations without Antigens. International Journal of Molecular Sciences. 2019; 20(1):45. https://doi.org/10.3390/ijms20010045
Chicago/Turabian StyleYanase, Yuhki, Yoshimi Matsuo, Tomoko Kawaguchi, Kaori Ishii, Akio Tanaka, Kazumasa Iwamoto, Shunsuke Takahagi, and Michihiro Hide. 2019. "Activation of Human Peripheral Basophils in Response to High IgE Antibody Concentrations without Antigens" International Journal of Molecular Sciences 20, no. 1: 45. https://doi.org/10.3390/ijms20010045