Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks
by
 ,
,
Martin Černý
1,2,*,† ,
,
Hana Habánová
1,3,4,†,
Miroslav Berka
1,
Markéta Luklová
1,3 and
Břetislav Brzobohatý
1,3,5 1
Department of Molecular Biology and Radiobiology, Faculty of AgriSciences Mendel University in Brno, 613 00 Brno, Czech Republic
2
Phytophthora Research Centre, Faculty of AgriSciences, Mendel University in Brno, 613 00 Brno, Czech Republic
3
CEITEC—Central European Institute of Technology, Faculty of AgriSciences Mendel University in Brno, 613 00 Brno, Czech Republic
4
Brno Ph.D. Talent, South Moravian Centre for International Mobility, 602 00 Brno, Czech Republic
5
Institute of Biophysics AS CR, 613 00 Brno, Czech Republic
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2018, 19(9), 2812; https://doi.org/10.3390/ijms19092812
Submission received: 24 July 2018
/
Revised: 13 September 2018
/
Accepted: 15 September 2018
/
Published: 18 September 2018
(This article belongs to the Special Issue Phytohormones and Their Crosstalk during Plant Growth, Development and Environmental Stress Adaptation)
Abstract
:Hydrogen peroxide (H2O2) is steadily gaining more attention in the field of molecular biology research. It is a major REDOX (reduction–oxidation reaction) metabolite and at high concentrations induces oxidative damage to biomolecules, which can culminate in cell death. However, at concentrations in the low nanomolar range, H2O2 acts as a signalling molecule and in many aspects, resembles phytohormones. Though its signalling network in plants is much less well characterized than are those of its counterparts in yeast or mammals, accumulating evidence indicates that the role of H2O2-mediated signalling in plant cells is possibly even more indispensable. In this review, we summarize hydrogen peroxide metabolism in plants, the sources and sinks of this compound and its transport via peroxiporins. We outline H2O2 perception, its direct and indirect effects and known targets in the transcriptional machinery. We focus on the role of H2O2 in plant growth and development and discuss the crosstalk between it and phytohormones. In addition to a literature review, we performed a meta-analysis of available transcriptomics data which provided further evidence for crosstalk between H2O2 and light, nutrient signalling, temperature stress, drought stress and hormonal pathways.
1. Introduction
Hydrogen peroxide, a chemical compound discovered by Louis Jacques Thenard a hundred years ago, has properties that could justify classifying it as a phytohormone. In nature, it can be of inorganic origin, for example, via reactions in the atmosphere [1] but H2O2 from this source has only an indirect effect on living organisms. Thenard was the first to discover not only that H2O2 decomposes into water but also that it can cause skin blistering at a high concentration. However, oxidative stress is not the sole effect of this molecule. It is an evolutionarily conserved signalling molecule and in plants, it has gained attention also for its role in the regulation of growth and development. Indeed, the number of H2O2-related research articles published each year has doubled since 2008, with Web of Science listing over 3000 plant science publications on this topic in the last five years. In this review, we summarize different aspects of H2O2-mediated responses in plants, starting with the sources, catabolism and transport of H2O2. We then describe mechanisms for its perception and discuss its role in plant signalling networks and its effects on plant growth and development.
2. Metabolism
Hydrogen peroxide H2O2 is a non-radical reactive oxygen species (ROS) and it, like singlet oxygen 1O2 and free radicals such as superoxide anion O2− and hydroxyl radical •OH, is one of the major members of the ROS family [2]. In contrast to other ROS, H2O2 is relatively stable, with a half-life of ms and its level in a plant leaf oscillates around 1 µmol per gram of fresh weight under natural conditions [3]. There are numerous routes, both enzymatic and non-enzymatic, for H2O2 production in plant cells. The key sources include photorespiration, electron transport chains and redox reactions in the apoplast [4,5]. The KEGG (Kyoto Encyclopedia of Genes and Genomes) database lists 150 classes of enzyme that produce or utilize hydrogen peroxide. Of these, only 29 enzymes encoded by 227 genes are annotated in Arabidopsis and the largest enzyme family formed by peroxidases has 75 entries (Figure 1, Supplementary tables). However, not all of these enzymes necessarily participate in peroxide metabolism in plants. For instance, a flavin-containing monooxygenase like YUC6 may produce hydrogen peroxide in the absence of its substrate but in vitro experiments indicate that in this case the uncoupled reaction represents less than 4% of the enzyme’s activity [6]. In contrast, mammalian flavin-containing monooxygenases are clearly a source of hydrogen peroxide [7]. The key enzymes that are involved in Arabidopsis H2O2 metabolism reside in the apoplast, peroxisome, chloroplast and mitochondria and they will be described in detail.
2.1. Electron. Transport Chains and Superoxide Dismutase
Under favourable conditions, the majority of intracellular H2O2 is produced from molecular oxygen by a stepwise reaction via a superoxide anion intermediate which undergoes enzymatic reduction to H2O2. Excessive energy and/or malfunctioning of chloroplast and mitochondrial energetic metabolism are key causes of superoxide anion generation in plant cells. In chloroplasts, superoxide anions are produced when the electron-transport chain of photosystem I is oversaturated by excessive irradiation and electrons are transmitted by the Mehler reaction to oxygen molecules [8]. The resulting superoxide anions are then converted to H2O2. This dismutation step is a pH-dependent non-enzymatic event (for details see for example, [9]) but cells also catalyse the process by means of superoxide dismutase (SOD) in order to rapidly remove the toxic superoxide radical. Besides photosystem I, H2O2 may also originate at the manganese-containing, oxygen-evolving complex which is the donor site of photosystem II and by the reduction of singlet oxygen or superoxide anions by photosynthetic electron transport chain components such as plastoquinol [10]. In seeds and non-photosynthetic parts of plants, the main sources of superoxide anion are coupled with the processes of cell respiration in mitochondria. Electron leakage occurs especially in complexes I, II and III and it is estimated that 1–5% of the oxygen entering the plant respiratory chain is converted into H2O2 [11,12,13]. The Arabidopsis genome encodes eight SOD isozymes which can be divided into three classes according to their metal cofactor (Fe2+, Mn2+, Cu2+). There are three chloroplastic Fe-SODs and two Mn-SODs localized in mitochondria. The Fe-SODs are considered to be the oldest in evolutionary terms but the two classes share structural similarities and can also be found in prokaryotes. In contrast, the Cu/Zn-SOD class, which has three isozymes in Arabidopsis, most likely emerged after oxygen saturated the atmosphere. It is specific to eukaryotes and can be present in different cell compartments [14,15].
2.2. NADPH Oxidase
The second largest group of H2O2-producing enzymes consists of the respiratory burst oxidases (Figure 1), which are also known as respiratory burst oxidase homologs (RBOHs) based on their homology to mammalian phagocyte NADPH oxidase (nicotinamide adenine dinucleotide phosphate oxidase). RBOHs, together with the type III cell wall peroxidases, are associated with the so-called “oxidative burst,” which is considered to be one of the main responses of plant cells to biotic or abiotic stress [19,20] but is also a crucial part of normal plant growth and development [21]. RBOHs are plasma membrane-localized proteins which oxidize cytosolic NADPH, transferring the released electron to O2 and producing superoxide which is then dismutated. In Arabidopsis, there are ten RBOH genes which are divided into three classes according to their tissue-specificity [22,23]. RBOHs are probably the best studied enzymatic ROS-generating system in plants and different regulatory mechanisms have been described. RBOHs undergo multiple post-translational modifications (PTMs), including S-nitrosylation and phosphorylation, that are required for enzyme activity and are regulated by calcium ions and phosphatidic acid [24,25].
2.3. Polyamine Oxidase
Hydrogen peroxide is an end product of oxidative degradation of amines and polyamine degradation is considered to be an especially important source of hydrogen peroxide in plants (e.g., [26]). Plant polyamines are catabolized by two distinct classes of amine oxidases, the flavin adenine dinucleotide (FAD)-dependent polyamine oxidases and the copper amine oxidases, of which there are, respectively, five and eight putative functional isozymes encoded by the Arabidopsis genome [27]. The copper amine oxidases oxidize primary amino groups, producing ammonia, H2O2 and an aminoaldehyde, whereas the polyamine oxidases oxidize the secondary amino groups and the reaction products depend on the catalytic mechanism and substrate specificity of a given isozyme. All five Arabidopsis polyamine oxidases are reportedly intracellular and oxidize the carbon on the exo-side of the N4 atom of spermine and spermidine to produce 1,3-diaminopropane, H2O2 and an aminoaldehyde [28]. Polyamines play an important role in plant tolerance of abiotic stress and at least part of this tolerance is associated with hydrogen peroxide production (see for example, review [29]). Furthermore, polyamines represent a direct link between H2O2 and hormonal pathways, as it has been shown that cytokinin increases the polyamine content of plants [30].
2.4. Peroxisomal Production of H2O2
Peroxisomal enzymes represent a major site of H2O2 production in a plant cell. In Arabidopsis, in addition to SOD and amine oxidases that are present in multiple compartments, peroxisomes contain acyl-CoA oxidases, glycolate oxidases, uricase, sulphite oxidase, aldehyde oxidase and sarcosine oxidase (Figure 1). Xanthine oxidase, which converts xanthine to urate and H2O2, can be also localized in peroxisomes [2] but a putative Arabidopsis homolog that preferentially accepts NAD+ as the electron acceptor [31] reportedly resides in the cytosol. A significant proportion of peroxisomal H2O2 originates during the beta-oxidation of long-chain fatty acids via acyl-CoA oxidase [32], which is an especially important process in germinating seeds that contain glyoxysomes, specialized peroxisome-like organelles. However, in photosynthetic tissues, the role of peroxisomes in H2O2 metabolism is predominantly via photorespiration reactions that may contribute up to 70% of the total production of H2O2 in a plant cell [33,34]. In this reaction, glycolate produced in chloroplasts is converted to glyoxylate by glycolate oxidase in peroxisomes. The Arabidopsis genome contains five genes encoding glycolate oxidase and their combined relative expression in photosynthetic tissues is the highest of all H2O2-producing enzymes (Figure 1). However, the actual levels of H2O2 in peroxisomes are kept in check by catalase and it is estimated that the peroxisomal H2O2 concentration is under 10 μM [35].
2.5. The H2O2 Scavenging System
Plant cells survive with H2O2 levels that would kill animal cells and the estimated endogenous H2O2 content of plant cells is usually much higher than that found in animals and bacteria [36]. H2O2 accumulation increases the probability of hydroxyl radical production via the Fenton reaction and this would cause significant oxidative damage to cellular structures if it were not for the presence of a highly efficient antioxidant system. Higher plants contain several types of peroxidases, including catalases, ascorbate peroxidases (APX), thiol-specific peroxidases and classical secretory plant peroxidase. Furthermore, non-enzymatic compounds like tocopherols, ascorbic acid and flavonoids and glutathione play significant roles in H2O2 scavenging [37,38]. The plastoquinone and ubiquinone pool also contribute to the ROS scavenging process as illustrated in recent reports [39,40]. In accordance, inhibition of enzymes that maintain the oxidized plastoquinone and ubiquinone pool, plastid terminal oxidases and mitochondrial alternative oxidases, respectively, stimulates H2O2 production [41,42].
2.6. Catalases
Though catalase belongs to the peroxidase family, it is usually considered separately due to its unique ability to convert two molecules of H2O2 into water and molecular oxygen without the need for any reductant. This heme-containing enzyme is first oxidized to a high-valence iron intermediate, which is then reduced by a further reaction with H2O2 [43]. Under specific circumstances, the intermediate may also react with a different substrate and catalase may oxidize donors such as alcohols or phenols. Catalase has a high turnover rate but a low substrate affinity, with a Km value in the millimolar range, a far greater concentration of H2O2 that that expected to be present in the cell [35]. As an illustration, the activity of a single molecule of rice catalase (kcat 80,000; Km 100 mM) [44] would be equivalent to more than 2200% of tobacco APX (kcat 1800; Km 0.022 mM) [45] at 100 mM H2O2 but to only 1% at concentrations below 1 µM H2O2, which would render catalase redundant. Of course, the constants determined in vitro may be misleading; the active form of catalase is a tetramer and it has been shown that, for example, PTMs may significantly affect the kinetics of a multimeric enzyme (e.g., [46]). Nevertheless, even though catalase activity has also been reported in the cytosol and mitochondria, its predominant localization is in peroxisomes, compartments with a high H2O2 concentration where its efficiency should be greatest (e.g., [47]). There are three functionally conserved classes of catalase with different spatial and developmental localizations in plants. For example, in tobacco catalase class I detoxifies H2O2 produced in photorespiration reactions, class II is localized in the vascular system and class III is present predominantly in flowers and fruits [48].
2.7. Ascorbate and Thiol-Specific Peroxidases
APX and glutathione peroxidases belong to the most important group of intracellular peroxidases [49]. Several types of APX have been described in plants; they include soluble enzymes in the cytosol, chloroplast and mitochondria and membrane-bound peroxidases in peroxisomes, glyoxysomes and thylakoids [50]. APX is the first enzyme in the so-called ascorbate-glutathione cycle, which includes monodehydroascorbate reductase, dehydroascorbate reductase and glutathione reductase and reduces H2O2 and regenerates ascorbate via NAD(P)H [49]. The Arabidopsis genome encodes seven different APX isozymes and as indicated above, APX may be more important than catalase for H2O2 metabolism. Indeed, it has been shown that in the absence of cytosolic APX1, the entire chloroplastic H2O2-scavenging system in Arabidopsis collapses, H2O2 levels increase and protein oxidation occurs [51]. The thiol-specific peroxidases peroxiredoxins and glutathione peroxidases detoxify a broad spectrum of peroxide substrates [8]. However, recent evidence from S. cerevisiae indicates that this could be a secondary role and that thiol peroxidases perceive and transfer oxidative signals to signalling proteins and regulate transcription [52]. In plants and bacteria, six groups of peroxiredoxins are recognized on the basis of differences in sequence, structure and positions of conserved cysteinyl residues [53].
2.8. Peroxidases (Class III)
Peroxidases are by far the most abundant family of enzymes in H2O2 metabolism (Figure 1). These so-called class III peroxidases probably have a correspondingly diverse range of functions, of which only a few, in certain plant species, have been revealed (see for example [54,55] for details). From the point of view of this review, it is important to note that the class III peroxidases participate not only in H2O2 catabolism via oxidation of phenolic compounds but also in producing it via an oxidative cycle using apoplastic reductants. For instance, it has been shown that in Arabidopsis cell culture they contribute to ca. 50% of the H2O2 produced during the oxidative burst in pathogen defence [56]. Class III peroxidases can be found in vacuoles but the majority are apoplastic or associated with cell walls in the apoplast as they play a key role in maintaining cell wall integrity by catalysing its cross-linking and loosening, lignification and suberization [57].
3. Transport
Normal levels of H2O2 leaf extracts are reported to be in the μmol per gram of fresh weight range but they may significantly vary within the same plant [3]. For instance, localization of hydrogen peroxide in different regions of the leaf reveals a pattern of increasing accumulation from the base to the leaf tip [58]. There is no clear evidence for long distance transport of H2O2 but it is the least reactive ROS and this allows it to travel at least among neighbouring cells or cellular compartments and to serve as an important signalling molecule [59]. Thus, if it is able to escape the H2O2-scavenging mechanisms described above and is not reduced to the highly reactive hydroxyl radical, it may freely diffuse from the site of its generation and reach its putative target. Questions of how it overcomes the competing H2O2-scavengers that prevent the targeted oxidation of redox-regulated proteins are still not fully answered [60] but it is now clear that transport mediated by simple diffusion would not explain, for example, rapid stress-induced transfer of H2O2 generated in apoplast by NADPH oxidases into cytosol and that a H2O2-specific transporter or channel must therefore exist.
Peroxiporins
Henzer and Steudle found that treatment with HgCl2 (an aquaporin activity inhibitor) caused a rapid decrease in H2O2 and water influx and they postulated the existence of an aquaporin subclass, peroxiporins [61]. The similarity of H2O2 to the water molecule indicates that aquaporins could have such a function. Plant aquaporins are recognized as multifunctional proteins transporting not only water but also many other small uncharged molecules (e.g., CO2 and nutrients) and they thus play a role in the regulation of plant growth and development and in responses to a wide range of stresses. Aquaporins belong to the ancient superfamily of major intrinsic proteins (MIPs) and are present throughout living organisms with the exception of some Archea and bacteria [62]. Plant aquaporins are divided into five subfamilies: plasma membrane intrinsic proteins (PIPs), tonoplast intrinsic proteins (TIPs), nodulin26-like intrinsic proteins (NIPs), small basic intrinsic proteins (SIPs) and uncategorized intrinsic proteins (XIPs). The latter two groups, which were discovered more recently, are not present in some plant species [63]. H2O2 has a higher polarity than water and thus not all aquaporins are peroxiporins. For instance, Hooijmaijers et al. employed heterologous expression of all 13 Arabidopsis PIPs in yeast and found that only five of them inhibited yeast growth in the presence of H2O2 [64]. Since the first report of H2O2 transport by an aquaporin appeared, this phenomenon has been studied in diverse plant species, including maize [65], rice and barley [66], Arabidopsis [64,67,68,69], tulip [70], tobacco, potato and tomato [71]. Kim and Steudle (2009) suggested the occurrence of feedback regulation in aquaporin-facilitated H2O2 transport, based on the observed inhibition of aquaporin transport capacity after H2O2 treatment [72]. Further studies showed that this inhibition may occur indirectly by the internalization of aquaporin into vesicles that is caused by the change in the phosphorylation status of aquaporins [73,74]. Hooijmaijers et al. (2012) also found that H2O2 treatment can alter aquaporin expression, indicating a feedback loop between H2O2 concentration and peroxiporin expression [64].
4. Signalling
It has been widely reported that H2O2 effects are dose-specific and that at low concentrations it serves as a signalling molecule. Despite H2O2 being rapidly removed by protective enzymes, the scavenging mechanisms are less effective at concentrations of around 10 nM, enabling H2O2 to be a second messenger [59,75]. In general, proteins are primary targets of all oxidative species and there are two modes of action by which H2O2 is perceived: direct oxidation of amino acid residues or reaction with reactive intermediates (e.g., [76]). The latter represents an indirect effect mediated via peroxide decomposition products (hydroxyl radical and singlet oxygen) and is usually considered to be a non-specific oxidative stress response. However, it has been shown that the transcription factor PerR, a major regulator of the peroxide inducible stress response in bacteria, senses H2O2 via this pathway, employing metal-catalysed histidine oxidation [77]. The complexity of ROS-mediated processes in plants somewhat limits our understanding of H2O2 signalling circuits and the present state of this understanding lags far behind that for bacteria, yeasts or mammalian cells. For instance, the ratio of superoxide radical to hydrogen peroxide may regulate the respiratory chain in mitochondria [78] and it is believed that the ratio of singlet oxygen plus superoxide radical to hydrogen peroxide determines the activation of cell death programs [79]. Some mechanisms have been conserved during evolution, whereas others seem to be plant-specific. Here, we summarize the main circuits that have been found to operate in plants.
4.1. Oxidation of Cysteine Residues
Targets of direct oxidation are predominantly cysteinyl residues and reactive thiol side chains can act as sensors or switches in both signal transduction and regulation of enzyme activity [76]. Depending on H2O2 concentration a cysteinyl residue can react to undergo several reversible or irreversible modifications, starting with sulfenic acid, which is highly reactive and reacts with other proximal thiolates resulting in the formation of inter/intramolecular disulphide bonds or S-glutathionylation. The reduction of disulphide bonds and the removal of glutathione are regulated by members of the thioredoxin and glutaredoxin enzyme families. Sulfenic acid can be also further oxidized by H2O2 to sulfinic or even sulfonic acid [76,80]. Some signalling models predict that a hypothetical receptor may undergo successive oxidation steps and that each step would correspond to a physiological response but it remains to be seen whether such a receptor exists. Experiments carried out in vitro have shown that the rate of reaction of hydrogen peroxide with cysteine is relatively low but this does not apply to H2O2-scavenging enzymes. The reaction of the cysteinyl residue in peroxiredoxin has an apparent second order rate constant seven orders of magnitude higher than that for cysteinyl in BSA [81] and Marinho et al. calculated that the H2O2 concentration needed for a peroxiredoxin-mediated response time of 5 min is as low as 0.2 nM [82]. The thiol-specific peroxidases thus act as receptors and, upon oxidation, interact with and oxidize effector proteins, forming a redox relay. For example, Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in stomatal closure [83]. Key enzymes in photosynthesis and carbohydrate metabolism are oxidized in response to H2O2, including RuBisCO, phosphoribulokinase, glyceraldehyde-3-phosphate dehydrogenase, transketolase and sedoheptulose-1,7-bisphosphatase [84]. It is very likely that this is also a redox relay mediated by peroxiredoxins present in the chloroplast but evidence for this is lacking.
4.2. Oxidation of Methionine Residues
Methionine, the second proteinaceous sulphur-containing amino acid, is usually not considered to be a regulatory target in H2O2 signalling but its first oxidized form (methionine sulfoxide) is the product of a PTM that can be reversed via the action of a specific reductase [76]. The fact that this enzyme increases H2O2 tolerance indicates that methionine residues have a role at least in the H2O2-induced stress response [85]. Jacques et al. studied protein methionine sulfoxide dynamics in catalase knock-out Arabidopsis and found that 51 proteins were significantly more oxidized compared to wild-type. They also demonstrated that the activity of glutathione S-transferase is reduced upon methionine oxidation [86].
4.3. Other Protein PTMs
It should be noted that the direct effect of H2O2 on protein PTMs is not limited to cysteine or methionine residues. In fact, the presence of oxidative PTMs has been shown to interfere with other PTMs close to the oxidized site [87]. An alteration in the PTM pattern can play a crucial role in signalling. The well-known regulator TP53, which participates in mammalian H2O2 signalling, has to integrate a complex network of PTMs [82]. Its Arabidopsis orthologue SOG1 (suppressor of gamma response 1) is hyperphosphorylated in response to ROS and it has been proposed that H2O2 regulates its hyperphosphorylation, ultimately leading to cell cycle regulation [88]. Examples from mammalian systems also indicate that PTM by ubiquitination and targeted protein degradation is key to the H2O2 response [82]. However, our knowledge about its role in plant H2O2 circuits is limited. It has been found that UPL5 ubiquitin ligase mediates degradation of the transcription factor WRKY53 [89] but there are more than 1500 E3 enzymes in Arabidopsis and this, together with extensive crosstalk with phytohormonal networks (which all to some extent converge on the proteasome) [90], represents a substantial obstacle to the elucidation of H2O2 signal transduction.
4.4. Transcription Factors
4.4.1. HsfA
Heat-shock transcription factors are transcriptional activators that, once trimerized, specifically bind cis-elements called heat shock elements, palindromic DNA sequences that are found in the promoters of heat stress-inducible genes of all eukaryotes, including that encoding APX, the major catabolic enzyme in Arabidopsis H2O2 metabolism [91,92]. The trimerization mechanism requires intramolecular disulphide bonds and it can be directly induced by H2O2 (reviewed in, for example, [93]). In Arabidopsis, HsfA2 was found to be involved in H2O2 signalling and it was shown that both its transcript and the transcript levels of its target genes were induced by treating with exogenous H2O2 [94].
4.4.2. NAC Domain-Containing Protein
NAC (No apical meristem Arabidopsis transcription activation factor Cup-shaped cotyledon) domain-containing proteins constitute one of the largest transcription factor families in plants and they are involved in multiple developmental and physiological processes, including senescence and abiotic stress responses. Multiple genes of this family have been found to be upregulated in response to H2O2 [95] treatment and it has been suggested that NAC042 (JUB1) functions as a modulator of cellular H2O2 levels [96]. NAC059-dependent gene expression was triggered after H2O2 treatment [97] indicating that NAC could be a primary target of H2O2. Furthermore, two transcription factors (NAC013 and NAC017) that apparently shuttle between the nucleus and endoplasmic reticulum membrane mediate redox-related retrograde signalling [98,99].
4.4.3. Mediators of RNA Polymerase
In yeast and mammals, an RNA polymerase inhibitor localized in cytosol is activated by H2O2 through the thioredoxin system and translocated into the nucleus [82]. Its putative orthologue in Arabidopsis is not known to be a H2O2-responsive protein but mediators of RNA polymerase II have been found to be upregulated in response to H2O2, including MED37C [94]. Shaikhali et al. showed that members of this family readily form oligomers in vitro via intramolecular disulphide bonds [100] and showed that root growth in the knock-out mutant med32 was significantly less affected by H2O2 than that in wild-type plants [101].
4.4.4. WRKY and ZAT (Zinc finger of Arabidopsis thaliana) Transcription Factors
There are 74 WRKY amino acid signature sequence-containing transcription factors in Arabidopsis that contain four-stranded β-sheet WRKY DNA binding domain/s ca 60 amino acids in length and zinc-finger motifs [102]. Like Nascent polypeptide-Associated Complex NAC domain-containing proteins, these transcription factors participate in stress-related responses and some have been found to be upregulated in response to H2O2. WRKY30 and WRKY53 were found to be upregulated in response to ozone and H2O2 exposure, with WRKY53 being much more responsive to H2O2 than WRKY30 and vice versa for ozone [103]. WRKY46 was upregulated by H2O2 [104] and WRKY70 is a putative interactor of the H2O2-responsive zinc finger protein ZAT7 [105]. ZAT12, another H2O2-responsive transcription factor, was proposed to mediate iron uptake control via its interaction with the FIT protein and with H2O2 as a signal in iron deficiency responses [106]. The present evidence indicates that WRKY transcription factors and ZAT zinc finger proteins participate in responses to H2O2 but a more detailed analysis of WRKY/ZAT-mediated transcription is needed in order to test the hypothesis that they play a role as the primary target. The fact that ZAT12 and ZAT5 respond positively to both ascorbate and H2O2 [107] indicates that this is probably not the case, at least for the ZATs.
4.5. Calcium Ions
Calcium ions play a key role in a vast array of signalling pathways in plants (e.g., [108]). Ca2+ is a second messenger like H2O2 and multiple characterized cascades require their combined effect, for example, via the opening of H2O2-dependent Ca2+ channels [109,110,111]. The Ca2+-binding protein calmodulin is an activator of catalase [112] and calmodulin-binding transcription activators have been found to be upregulated by H2O2. BT2, another calmodulin-binding protein which is upregulated by H2O2, is also part of an E3 ligase complex [113,114]. Moreover, Ca2+-dependent phosphorylation activates NADPH oxidases (e.g., [115]) and plays a role in the so-called ROS-Ca2+ hubs described in Section 5.11.
5. H2O2 in Growth and Development
The role of H2O2 in the life of plants is illustrated in Figure 2 and outlined in the following text, which presents examples from different developmental stages.
5.1. The Crosstalk between H2O2 and Phytohormones
The first genome-wide analyses of plant H2O2 signalling revealed a connection between ethylene and H2O2. Ethylene signalling is induced in response to H2O2 accumulation [116] but the ethylene receptor ETR1 itself perceives H2O2 directly in an ethylene-independent manner that does not require its kinase domain [117]. ROS is a key component of phytohormonal signalling networks and does not only mediate stress-related pathways. From the proteome-wide point of view, catalases, peroxiredoxins, disulphide isomerases and thioredoxins have been detected at high frequencies in phytohormone-responsive proteomics studies and APX, glutathione S-transferase and class III peroxidase were found at least once in all reported hormone-responsive proteomes; see Table 1 [90]. H2O2 mediates hormonal homeostasis (e.g., auxin conjugation [118] and degradation [119]) but enzymes involved in hormone metabolism may produce H2O2. These include abscisic acid aldehyde oxidases, enzymes that catalyse the final step in abscisic acid biosynthesis producing H2O2 in the process. Auxin aldehyde oxidases are also present in Arabidopsis but it is not clear to what extent these enzymes contribute to auxin metabolism [120]. Furthermore, monooxygenases may catalyse a H2O2-producing side reaction, as illustrated above for the auxin biosynthetic enzyme YUC6, which is encoded by a member of a multigene family in Arabidopsis. Similar enzymes are present in the pathways of cytokinin metabolism (hydroxylases cytochrome P450 735A1 and 735A2), gibberellin and brassinosteroid biosynthesis (ent-kaurene oxidase, ent-kaurenoic acid oxidase) and abscisic acid metabolism (hydroxylases cytochrome P450 707A1-707A4). Our comparison of expression profiles revealed that multiple hormonal metabolism genes share patterns of expression with those of H2O2 metabolism; the former include ABA4 (31 similar patterns), tryptophan aminotransferases TAR3 (29) and TAR4 (29), methyl esterase MES1 (28), cytokinin biosynthetic genes IPT2 (26), IPT6 (22) and LOG2 (24), ethylene metabolism genes ACO2 (23) and ACS4 (20) and Ent-copalyl diphosphate synthase GA1 (27) (see Table 2 and Supplementary Materials for details).
5.2. Light Signalling
Light signal transduction is involved in H2O2 metabolism and/or signalling. It has been demonstrated that blue-light perception by cryptochrome is directly coupled with H2O2 generation [140,141,142]. It has also been proposed that phytochrome B modulates homeostasis of reactive oxygen species in roots via synthesis and transport of abscisic acid [143]. Our comparison of expression profiles revealed that genes participating in light signalling share patterns of expression with H2O2 metabolism genes; the former include MED25 which acts in the repression of phytochrome B-mediated light signalling (26 similarities), COP1 (32), phytochrome A (29), PIF1 (28), phytochrome B (26), phytochrome C (25) and cryptochrome 1 (18) (see Table 2 and Supplementary Materials for details).
5.3. Dry Seed
The majority of plants from temperate climate zones produce so-called orthodox seeds which pass through a phase of intensive desiccation and in this state, they are able to survive for periods ranging from months to tens of years (or even hundreds of years in some cases) [144]. The quiescent state limits enzymatic activity to a minimal level but H2O2 and other ROS are still produced and can be accumulated during seed ageing. H2O2 accumulation in seeds may cause significant damage to storage molecules and loss of viability but the degree to which it accumulates and the sensitivity to oxidative damage is species specific. For example, H2O2 does not accumulate in Brassica napus seeds [145].
5.4. Germination
Seed germination is defined as a three-phase process, starting with rapid water intake and ending with seed coat rupture, usually by radicle protuberance. In imbibed and germinating seeds, high levels of H2O2 are produced mainly as a product of intensive metabolism in mitochondria, peroxisomes and glyoxysomes but also by NADPH oxidases and through lipid peroxidation [146]. Though seeds contain both enzymatic and nonenzymatic ROS scavenging machinery to prevent oxidative damage [147], H2O2 is also needed to remove mechanical and hormonal barriers that inhibit germination. H2O2 promotes endosperm weakening [148,149] and triggers an increase in gibberellin biosynthesis and a decrease in abscisic acid levels [150,151,152,153,154]. It also mediates selective oxidation of mRNA and proteins [155,156], for example irreversible carbonylation of storage proteins that enables their rapid mobilization via proteasomes [157]. Another key aspect of seed germination, especially in cereals, is the activation of α-amylase and the promotion of programmed cell death (PCD) in the aleurone layer. Here, H2O2 is produced by NADPH oxidase and it functions via interplay with DELLA proteins (proteins with the highly conserved amino acid sequence motif DELLA), key components of the gibberellin signalling pathway [158,159,160]. In many respects, the role of H2O2 in germination is similar to that of a growth regulator and studies of exogenous H2O2 application have demonstrated that its effect is dose dependent [161,162,163].
5.5. Root Development
Ample evidence showed that ROS regulates root development via NADPH oxidases [111,164,165]. The phytohormone that is key to the regulation of root growth is auxin, which is well known to mediate changes in H2O2 levels and thus promote cell growth and lateral root formation [166,167,168]. However, a recent study indicated that in mediating the induction of lateral roots, H2O2 acts downstream of melatonin, an auxin-like indoleamine compound [169]. Root tip growth is also known to be affected by H2O2 [170,171]. Polar auxin transport seems to regulate H2O2-induced root gravitropism [163] and exogenous H2O2 treatment can disrupt this sensing, probably due to a change in auxin receptor distribution [172]. Abscisic acid inhibits primary root growth by activating NADPH oxidases and thus reducing auxin sensitivity [173] and a RBOH was proposed to interact with abscisic acid in the regulation of lateral root growth in Arabidopsis under drought stress [174]. H2O2-mediated root growth in response to stress was also found in cucumber [175], cotton [176] and rice [177].
5.6. Shoot Development
Shoot growth and development of shoot architecture are driven by phytohormones, especially auxin and cytokinin, levels of which are highly correlated with environmental conditions. Auxin participates in cell growth by inducing cell wall peroxidases (peroxidases class III) and NADPH oxidases to produce ROS and promote cell wall loosening and further cell elongation [178,179]. H2O2 has been reported to mediate apical dominance [180], photosynthesis [181] and leaf epinasty [182].
5.7. Stomatal Movement
Stomata are formed as gaps between pairs of guard cells and changes in guard cell turgor mediate the opening and closure of the stomatal pore. Stomatal closure is an example of rapid leaf-to-leaf communication mediated by ROS (e.g., [183]). Under conditions of excessive irradiation, an autopropagating wave of ROS rapidly transfers a signal to leaves that are not directly exposed to light and initiates stomatal closure. This closure can be induced by multiple stimuli, including brassinosteroids [184,185], strigolactones [186], jasmonic acid and salicylic acid [187], CO2 [188], ethylene [189,190], glucose [191] and interactions among them [192]. The best-described mechanism is that mediated by abscisic acid that recruits calcium ions, nitric oxide (NO), H2O2 and regulatory phosphorylation [193,194]. Guard cells generate H2O2 by means of amine oxidases [195], peroxidases and RBOHs [196,197]. The activity of RBOHs is regulated by Ca2+ binding [198] and phosphorylation by protein kinase OST1 (OPEN STOMATA 1) [199], which in turn is regulated by abscisic acid and interacts with a peroxiporin [200,201]. The overall H2O2-mediated machinery is much more complex. For example, hydrogen sulphide promotes H2O2 production by stimulating RBOH activity [202] but the presence of flavonols in guard cells and H2O2 scavenging inhibits stomatal closure [203] and it has been shown that this flavanol accumulation is induced by 5-aminolevulinic acid [204,205].
5.8. Pollination
In generatively propagating plants, H2O2 and other ROS play a key role in pollen navigation and gametophyte fusion. Angiosperms have developed different reproductive barriers to avoid self-fertilization, one of the most widespread being self-incompatibility [206]. H2O2 level is elevated during the incompatible reaction, triggering PCD. In contrast, the compatible reaction decreases the level of H2O2 in the stigma and the development of the pollen tube is promoted. The further growth of and the penetration of the ovule by, the pollen tube is guided by quite complicated signalling machinery, including the FERONIA protein which modulates NADPH oxidase activity [207,208]. ROS accumulation, especially that of the hydroxyl radical which is largely generated from H2O2, is then crucial for pollen tube rupture and the release of sperm cells [209].
5.9. Fruit Ripening
The involvement of H2O2 in the ripening process is known but not fully understood. Huan et al. proposed that H2O2 acts as a signalling molecule in the middle stage of peach fruit development but that it serves as an important toxic molecule, stimulating lipid peroxidation and oxidative stress, during the late stage of fruit ripening [210]. Kumar et al. analysed ripening in tomato and found changes in the redox state during different stages of ripening with a significant increase of H2O2 at the so-called breaker point (defined by the initial change in fruit colour) [211]. The increase in H2O2 is most likely regulated by ethylene, the key regulator of fruit ripening that enhances respiration rate and ROS production [212].
5.10. Senescence and Cell Death
Senescence ultimately leads to the death of plant organs or whole plants. It is a multistep process by which the plant recovers and recycles valuable nutrient components that would otherwise be lost [213]. The role of H2O2 in plant senescence was investigated by Bieker at al., who showed time-dependent levels of H2O2 and H2O2-scavenging enzymes in senescent leaves [214]. In such tissues H2O2 mediates PCD together with stress phytohormones like ethylene [215] or salicylic acid [216]. H2O2 levels are transiently elevated at the initial point of leaf senescence and peak again during the terminal stage [217,218] and this accumulation is reportedly more pronounced inside interveinal tissue [219]. Furthermore, transgenic lines with lower H2O2 levels display delayed senescence [96,214].
5.11. Stress
The key phytohormones orchestrating plant stress responses are abscisic acid, salicylic acid, jasmonates and ethylene and all of these phytohormones employ H2O2 in their signalling cascades in an either upstream or downstream manner [220]. Putative markers of nutrient status, temperature stress and drought stress share patterns of expression with those of H2O2 metabolism (Table 2) and H2O2 has been implicated in cold acclimation [221], salt stress responses and salt stress tolerance [222,223,224] and hypoxia stress [225]. Important targets in these responses are RBOHs [177,226,227]. Recently, maintenance of acquired thermotolerance was found to be interlinked with generation of H2O2 by RBOHs [228] and these NADH oxidases also participate in H2O2 production in biotic interactions. Under pathogen attack, ROS accumulation is involved in PCD of infected and surrounding cells [229]. This hypersensitive response is orchestrated by the phytohormones ethylene, JA and SA (e.g., [214]) but high cytokinin levels also induce H2O2 accumulation [230]. H2O2 has been implicated in the susceptibility of Brassica napus to Leptosphaeria maculans [231], resistance to root-knot nematode in tomato [232], systemic virus resistance in Nicotiana benthamiana [233] and reduction of rot in postharvest citrus fruits [234]. In accordance, plants primed with H2O2 or with a higher basal level of H2O2 formation display enhanced resistance to stressors [42,235].
It is well established that a significant proportion of H2O2-mediated stress response originates from its decomposition products. This decomposition is enhanced by the presence of transient metal catalysts through the so-called Haber-Weiss reaction. It is widely postulated that this reaction accounts for the in vivo generation of the highly reactive hydroxyl radical, which is a prime cause of oxidative damage to biomolecules (e.g., [9,236]). The hydroxyl radical is one of the strongest oxidants known and reacts at nearly diffusion-limited rates near the site of its formation [237]. Besides its ability to damage anything in its close vicinity and generate further radicals, the hydroxyl radical seems to be a potent effector in calcium and potassium homeostasis, regulation of cell elongation and stress-induced cell death [111,238,239,240,241]. Furthermore, hydroxyl radical-mediated activation of calcium channels is also proposed to be a part of the so-called ROS-Ca2+ hub, the mechanism that is utilized to perceive and amplify signal. This self-amplifying system employs Ca2+-dependent phosphorylation of NADPH oxidases and promotes hydroxyl radical production that, in turn, stimulates Ca2+-influx and NADPH oxidases’ activity (see for example [242]). The ROS-Ca2+ hub is believed to be central to hypersensitive response, phytohormonal signalling or abiotic stress responses [115,158,243,244]
Organelles like chloroplasts or mitochondria are key cellular sensors of environmental fluctuations and integral parts of plant stress responses. They communicate information by signalling to nuclei via stress-triggered retrograde signals, including ROS (reviewed in Reference [245]). Recent reports show not only that H2O2 participates indirectly via ROS triggered signals but also that it can transfer from chloroplasts to nuclei and facilitate photosynthetic control over gene expression [246].
6. Conclusions
H2O2 represents a key signalling molecule, connecting the signalling pathways of multiple phytohormones and acting as a second messenger in response to diverse conditions modulating plant growth and development. Its dose-dependent effect on growth clearly indicates that H2O2 is a growth regulator but can we also refer to H2O2 as a putative phytohormone? It is produced and degraded by the plant in response to stimuli and it is perceived by specialized proteins and elicits a response at low nanomolar concentrations. However, the limiting factor is its transport. Though it can be readily transported within a single cell and exported to extracellular space, it is not believed to serve as a long-distance signal due to its low stability and the presence of H2O2 scavengers. Exogenous treatment with H2O2 elicits a response and H2O2 gradients are established in plant organs but it is believed that signal propagation is sequential and that H2O2 reaches only neighbouring cells [248]. In conclusion, the recent literature offers multiple examples that reveal H2O2 as a versatile mediator of molecular communication in plants and whether we classify it as a phytohormone or not, this does not change its importance in the life of plants. There are new perspectives emerging in the field of H2O2 research with tools being developed for the detection of low micromolar and even picomolar H2O2 concentrations [249,250] and it is likely that their eventual application in plant sciences will provide answers to some of our questions about H2O2 transport and concentration dynamics. Similarly, we may expect that increasing sensitivity in proteomics approaches combined with imaging or laser microdissection techniques (e.g., [251]) will reveal more H2O2 targets and their spatio-temporal distribution.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/19/9/2812/s1.
Author Contributions
M.Č., H.H. performed the analytical and systematic search of the literature, analysed and interpreted the data and wrote the manuscript. M.L. performed and interpreted meta analyses, M.Č., H.H. and M.B. prepared figures, B.B. reviewed the final text and provided critical comments.
Funding
This research was funded partially by grant TE02000177 (TACR), AF-IGA-IP-2018/014 (Internal Grant Agency of Faculty of AgriSciences, Mendel University in Brno) and the LQ1601 (CEITEC 2020) project with financial contribution made by the Ministry of Education, Youths and Sports of the Czech Republic from within special support paid from the National Programme for Sustainability II funds.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Möller, D. Atmospheric hydrogen peroxide: Evidence for aqueous-phase formation from a historic perspective and a one-year measurement campaign. Atmos. Environ. 2009, 43, 5923–5936. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Cheeseman, J.M. Hydrogen peroxide concentrations in leaves under natural conditions. J. Exp. Bot. 2006, 57, 2435–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrzaczek, M.; Brosché, M.; Kangasjärvi, J. ROS signalling loops—Production, perception, regulation. Curr. Opin. Plant Biol. 2013, 16, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Bloom, A.J.; Queval, G.; Noctor, G. Photorespiratory Metabolism: Genes, Mutants, Energetics and Redox Signalling. Annu. Rev. Plant Biol. 2009, 60, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Mashiguchi, K.; Chen, Q.; Kasahara, H.; Kamiya, Y.; Ojha, S.; DuBois, J.; Ballou, D.; Zhao, Y. The biochemical mechanism of auxin biosynthesis by an arabidopsis YUCCA flavin-containing monooxygenase. J. Biol. Chem. 2013, 288, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Siddens, L.K.; Krueger, S.K.; Henderson, M.C.; Williams, D.E. Mammalian flavin-containing monooxygenase (FMO) as a source of hydrogen peroxide. Biochem. Pharmacol. 2014, 89, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, K.-J.; Turkan, I.; Krieger-Liszkay, A. Redox- and Reactive Oxygen Species-Dependent Signaling into and out of the Photosynthesizing Chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Khorobrykh, S.A.; Karonen, M.; Tyystjärvi, E. Experimental evidence suggesting that H2O2 is produced within the thylakoid membrane in a reaction between plastoquinol and singlet oxygen. FEBS Lett. 2015, 589, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The Roles of Mitochondrial Reactive Oxygen Species in Cellular Signaling and Stress Response in Plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Mignolet-Spruyt, L.; Xu, E.; Idänheimo, N.; Hoeberichts, F.A.; Mühlenbock, P.; Brosché, M.; Van Breusegem, F.; Kangasjärvi, J. Spreading the news: Subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant Mitochondria and Oxidative Stress: Electron Transport, NADPH Turnover and Metabolism of Reactive Oxygen Species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—Mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef]
- Tanz, S.K.; Castleden, I.; Hooper, C.M.; Vacher, M.; Small, I.; Millar, H.A. SUBA3: A database for integrating experimentation and prediction to define the SUBcellular location of proteins in Arabidopsis. Nucleic Acids Res. 2012, 41, D1185–D1191. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, V.; Hanlon, M.R.; Contrino, S.; Ferlanti, E.S.; Karamycheva, S.; Kim, M.; Rosen, B.D.; Cheng, C.-Y.; Moreira, W.; Mock, S.A.; et al. Araport: The Arabidopsis Information Portal. Nucleic Acids Res. 2015, 43, D1003–D1009. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Dunand, C.; Puppo, A.; Pauly, N. A burst of plant NADPH oxidases. Trends Plant Sci. 2012, 17, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Swanson, S.; Gilroy, S. ROS in plant development. Physiol. Plant. 2010, 138, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagi, M.; Fluhr, R. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.-W.; Feechan, A.; Yin, M.; Saidi, N.B.B.; Le Bihan, T.; Yu, M.; Moore, J.W.; Kang, J.-G.; Kwon, E.; Spoel, S.H.; et al. S-nitrosylation of NADPH oxidase regulates cell death in plant immunity. Nature 2011, 478, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Yan, M.; Zhang, Q. Functional regulation of plant NADPH oxidase and its role in signaling. Plant Signal. Behav. 2017, 12, e1356970. [Google Scholar] [CrossRef] [PubMed]
- Yoda, H.; Yamaguchi, Y.; Sano, H. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant Physiol. 2003, 132, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-Containing Amine Oxidases and FAD-Dependent Polyamine Oxidases Are Key Players in Plant Tissue Differentiation and Organ Development. Front. Plant Sci. 2016, 7, 824. [Google Scholar] [CrossRef] [PubMed]
- Fincato, P.; Moschou, P.N.; Spedaletti, V.; Tavazza, R.; Angelini, R.; Federico, R.; Roubelakis-Angelakis, K.A.; Tavladoraki, P. Functional diversity inside the Arabidopsis polyamine oxidase gene family. J. Exp. Bot. 2011, 62, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen Peroxide and Polyamines Act as Double Edged Swords in Plant Abiotic Stress Responses. Front. Plant Sci. 2016, 7, 1343. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Kuklová, A.; Hoehenwarter, W.; Fragner, L.; Novák, O.; Rotková, G.; Jedelský, P.L.; Žáková, K.; Šmehilová, M.; Strnad, M.; et al. Proteome and metabolome profiling of cytokinin action in Arabidopsis identifying both distinct and similar responses to cytokinin down- and up-regulation. J. Exp. Bot. 2013, 64, 4193–4206. [Google Scholar] [CrossRef] [PubMed]
- Hesberg, C.; Hänsch, R.; Mendel, R.R.; Bittner, F. Tandem Orientation of Duplicated Xanthine Dehydrogenase Genes from Arabidopsis thaliana: Differential gene expression and enzyme activities. J. Biol. Chem. 2004, 279, 13547–13554. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Baker, A.; Bartel, B.; Linka, N.; Mullen, R.T.; Reumann, S.; Zolman, B.K. Plant Peroxisomes: Biogenesis and Function. Plant Cell 2012, 24, 2279–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauwe, H.; Hagemann, M.; Fernie, A.R. Photorespiration: Players, partners and origin. Trends Plant Sci. 2010, 15, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Maurino, V.G.; Peterhansel, C. Photorespiration: Current status and approaches for metabolic engineering. Curr. Opin. Plant Biol. 2010, 13, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Drago, I.; Behera, S.; Zottini, M.; Pizzo, P.; Schroeder, J.I.; Pozzan, T.; Schiavo, F.L. H2O2 in plant peroxisomes: An in vivo analysis uncovers a Ca2+-dependent scavenging system. Plant J. 2010, 62, 760–772. [Google Scholar] [CrossRef] [PubMed]
- del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.D.; Van Breusegem, F. Hydrogen peroxide—A central hub for information flow in plant cells. AoB Plants 2012, 2012, pls014. [Google Scholar] [CrossRef] [PubMed]
- Ksas, B.; Légeret, B.; Ferretti, U.; Chevalier, A.; Pospíšil, P.; Alric, J.; Havaux, M. The plastoquinone pool outside the thylakoid membrane serves in plant photoprotection as a reservoir of singlet oxygen scavengers. Plant Cell Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Khorobrykh, S.; Tyystjärvi, E. Plastoquinol generates and scavenges reactive oxygen species in organic solvent: Potential relevance for thylakoids. Biochim. Biophys. Acta-Bioenerg. 2018, 1859, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Popov, V.; Simonian, R.; Skulachev, V.; Starkov, A. Inhibition of the alternative oxidase stimulates H2O2 production in plant mitochondria. FEBS Lett. 1997, 415, 87–90. [Google Scholar] [CrossRef]
- Wiciarz, M.; Gubernator, B.; Kruk, J.; Niewiadomska, E. Enhanced chloroplastic generation of H2O2 in stress-resistant Thellungiella salsuginea in comparison to Arabidopsis thaliana. Physiol. Plant. 2015, 153, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Prieto, M.; Biarnés, X.; Vidossich, P.; Rovira, C. The molecular mechanism of the catalase reaction. J. Am. Chem. Soc. 2009, 131, 11751–11761. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.; Mishra, P.; Das, P.; Sabat, S.C. Expression and purification of soluble bio-active rice plant catalase-A from recombinant Escherichia coli. J. Biotechnol. 2012, 157, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, S.; Kitamura, M.; Koja, N. Triple mutation of Cys26, Trp35 and Cys126 in stromal ascorbate peroxidase confers H2O2 tolerance comparable to that of the cytosolic isoform. Biochem. Biophys. Res. Commun. 2008, 372, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Doubnerová, V.; Müller, K.; Ryšlavá, H. Characterization of phosphoenolpyruvate carboxylase from mature maize seeds: Properties of phosphorylated and dephosphorylated forms. Biochimie 2010, 92, 1362–1370. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant catalases: Peroxisomal redox guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Fomenko, D.E.; Koc, A.; Agisheva, N.; Jacobsen, M.; Kaya, A.; Malinouski, M.; Rutherford, J.C.; Siu, K.-L.; Jin, D.-Y.; Winge, D.R.; et al. Thiol peroxidases mediate specific genome-wide regulation of gene expression in response to hydrogen peroxide. Proc. Natl. Acad. Sci. USA 2011, 108, 2729–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebthal, M.; Maynard, D.; Dietz, K.-J. Peroxiredoxins and Redox Signaling in Plants. Antioxid. Redox Signal. 2018, 28, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Podgórska, A.; Burian, M.; Szal, B. Extra-Cellular But Extra-Ordinarily Important for Cells: Apoplastic Reactive Oxygen Species Metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.A.; Daudi, A.; Finch, P.; Butt, V.S.; Whitelegge, J.P.; Souda, P.; Ausubel, F.M.; Bolwell, G.P. A peroxidase-dependent apoplastic oxidative burst in cultured Arabidopsis cells functions in MAMP-elicited defense. Plant Physiol. 2012, 158, 2013–2027. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, S.V.; Kukavica, B.; Vidović, M.; Morina, F.; Menckhoff, L. Class III Peroxidases: Functions, Localization and Redox Regulation of Isoenzymes. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer International Publishing: Cham, Switzerland, 2018; pp. 269–300. [Google Scholar]
- Tewari, R.K.; Singh, P.K.; Watanabe, M. The spatial patterns of oxidative stress indicators co-locate with early signs of natural senescence in maize leaves. Acta Physiol. Plant. 2013, 35, 949–957. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Biological Production, Detection and Fate of Hydrogen Peroxide. Antioxid. Redox Signal. 2017, 29, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Stöcker, S.; Van Laer, K.; Mijuskovic, A.; Dick, T.P. The Conundrum of Hydrogen Peroxide Signaling and the Emerging Role of Peroxiredoxins as Redox Relay Hubs. Antioxid. Redox Signal. 2018, 28, 558–573. [Google Scholar] [CrossRef] [PubMed]
- Henzler, T.; Steudle, E. Transport and metabolic degradation of hydrogen peroxide in Chara corallina: Model calculations and measurements with the pressure probe suggest transport of H2O2 across water channels. J. Exp. Bot. 2000, 51, 2053–2066. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. Biochim. Biophys. Acta-Gen. Subj. 2014, 1840, 1468–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooijmaijers, C.; Rhee, J.Y.; Kwak, K.J.; Chung, G.C.; Horie, T.; Katsuhara, M.; Kang, H. Hydrogen peroxide permeability of plasma membrane aquaporins of Arabidopsis thaliana. J. Plant Res. 2012, 125, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Heinen, R.B.; Berny, M.C.; Chaumont, F. Maize plasma membrane aquaporin ZmPIP2;5 but not ZmPIP1;2, facilitates transmembrane diffusion of hydrogen peroxide. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Katsuhara, M.; Sasano, S.; Horie, T.; Matsumoto, T.; Rhee, J.; Shibasaka, M. Functional and molecular characteristics of rice and barley NIP aquaporins transporting water, hydrogen peroxide and arsenite. Plant Biotechnol. 2014, 31, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Bienert, G.P.; Møller, A.L.B.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific Aquaporins Facilitate the Diffusion of Hydrogen Peroxide across Membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Dynowski, M.; Schaaf, G.; Loque, D.; Moran, O.; Ludewig, U. Plant plasma membrane water channels conduct the signalling molecule H2O2. Biochem. J. 2008, 414, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wang, X.; Li, P.; Wang, H.; Ji, H.; Xie, J.; Qiu, Q.; Shen, D.; Dong, H. Plant Aquaporin AtPIP1;4 Links Apoplastic H2O2 Induction to Disease Immunity Pathways. Plant Physiol. 2016, 171, 1635–1650. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.K.; Yoshikawa, N.; Ishikawa, T.; Sawa, Y.; Shibata, H. Substitution of a single amino acid residue in the aromatic/arginine selectivity filter alters the transport profiles of tonoplast aquaporin homologs. Biochim. Biophys. Acta 2012, 1818, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J. 2011, 66, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.X.; Steudle, E. Gating of aquaporins by light and reactive oxygen species in leaf parenchyma cells of the midrib of Zea mays. J. Exp. Bot. 2009, 60, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Chaumont, F. Aquaporin-facilitated transmembrane diffusion of hydrogen peroxide. Biochim. Biophys. Acta 2014, 1840, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Verdoucq, L.; Rodrigues, O.; Martinière, A.; Luu, D.T.; Maurel, C. Plant aquaporins on the move: Reversible phosphorylation, lateral motion and cycling. Curr. Opin. Plant Biol. 2014, 22, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Skalák, J.; Cerna, H.; Brzobohatý, B. Advances in purification and separation of posttranslationally modified proteins. J. Proteomics 2013, 92, 2–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-W.; Helmann, J.D. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature 2006, 440, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Andronis, E.A.; Moschou, P.N.; Toumi, I.; Roubelakis-Angelakis, K.A. Peroxisomal polyamine oxidase and NADPH-oxidase cross-talk for ROS homeostasis which affects respiration rate in Arabidopsis thaliana. Front. Plant Sci. 2014, 5, 132. [Google Scholar] [CrossRef] [PubMed]
- Sabater, B.; Martín, M. Hypothesis: Increase of the ratio singlet oxygen plus superoxide radical to hydrogen peroxide changes stress defense response to programmed leaf death. Front. Plant Sci. 2013, 4, 479. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Chibani, K.; Jacquot, J.-P.; Rouhier, N. Cysteine–based redox regulation and signaling in plants. Front. Plant Sci. 2013, 4, 105. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Lv, D.; Wang, P.; Wang, X.-C.; Chen, J.; Miao, C.; Song, C.-P. An Arabidopsis Glutathione Peroxidase Functions as Both a Redox Transducer and a Scavenger in Abscisic Acid and Drought Stress Responses. Plant Cell Online 2006, 18, 2749–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthuramalingam, M.; Matros, A.; Scheibe, R.; Mock, H.-P.; Dietz, K.-J. The hydrogen peroxide-sensitive proteome of the chloroplast in vitro and in vivo. Front. Plant Sci. 2013, 4, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.-W.; Lee, S.-H.; Chieh, P.-S.; Lin, C.-S.; Wang, Y.-C.; Chan, M.-T. Arabidopsis Root-Abundant Cytosolic Methionine Sulfoxide Reductase B Genes MsrB7 and MsrB8 are Involved in Tolerance to Oxidative Stress. Plant Cell Physiol. 2012, 53, 1707–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacques, S.; Ghesquière, B.; De Bock, P.-J.; Demol, H.; Wahni, K.; Willems, P.; Messens, J.; Van Breusegem, F.; Gevaert, K. Protein Methionine Sulfoxide Dynamics in Arabidopsis thaliana under Oxidative Stress. Mol. Cell. Proteomics 2015, 14, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Hardin, S.C.; Larue, C.T.; Oh, M.-H.; Jain, V.; Huber, S.C. Coupling oxidative signals to protein phosphorylation via methionine oxidation in Arabidopsis. Biochem. J. 2009, 422, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, D.; Alvim Kamei, C.L.; Cools, T.; Vanderauwera, S.; Takahashi, N.; Okushima, Y.; Eekhout, T.; Yoshiyama, K.O.; Larkin, J.; Van den Daele, H.; et al. The Arabidopsis SIAMESE-RELATED cyclin-dependent kinase inhibitors SMR5 and SMR7 regulate the DNA damage checkpoint in response to reactive oxygen species. Plant Cell 2014, 26, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zentgraf, U. A HECT E3 ubiquitin ligase negatively regulates Arabidopsis leaf senescence through degradation of the transcription factor WRKY53. Plant J. 2010, 63, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Černý, M.; Novák, J.; Habánová, H.; Cerna, H.; Brzobohatý, B. Role of the proteome in phytohormonal signaling. Biochim. Biophys. Acta-Proteins Proteom. 2016, 1864, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Schöffl, F.; Rieping, M.; Baumann, G.; Bevan, M.; Angermüller, S. The function of plant heat shock promoter elements in the regulated expression of chimaeric genes in transgenic tobacco. Mol. Gen. Genet. 1989, 217, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Panchuk, I.I.; Volkov, R.A.; Schöffl, F. Heat stress- and heat shock transcription factor-dependent expression and activity of ascorbate peroxidase in Arabidopsis. Plant Physiol. 2002, 129, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could Heat Shock Transcription Factors Function as Hydrogen Peroxide Sensors in Plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, A.; Yabuta, Y.; Yoshida, E.; Maruta, T.; Yoshimura, K.; Shigeoka, S. Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress. Plant J. 2006, 48, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balazadeh, S.; Wu, A.; Mueller-Roeber, B. Salt-triggered expression of the ANAC092-dependent senescence regulon in Arabidopsis thaliana. Plant Signal. Behav. 2010, 5, 733–735. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.-I.; Asensi-Fabado, M.A.; Munné-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.-P.; Mueller-Roeber, B. ORS1, an H2O2-responsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; Van Aken, O.; De Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A Membrane-Bound NAC Transcription Factor, ANAC017, Mediates Mitochondrial Retrograde Signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clercq, I.; Vermeirssen, V.; Van Aken, O.; Vandepoele, K.; Murcha, M.W.; Law, S.R.; Inzé, A.; Ng, S.; Ivanova, A.; Rombaut, D.; et al. The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis. Plant Cell 2013, 25, 3472–3490. [Google Scholar] [CrossRef] [PubMed]
- Shaikhali, J.; Davoine, C.; Brännström, K.; Rouhier, N.; Bygdell, J.; Björklund, S.; Wingsle, G. Biochemical and redox characterization of the mediator complex and its associated transcription factor GeBPL, a GLABROUS1 enhancer binding protein. Biochem. J. 2015, 468, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Shaikhali, J.; Davoine, C.; Björklund, S.; Wingsle, G. Redox regulation of the MED28 and MED32 mediator subunits is important for development and senescence. Protoplasma 2016, 253, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY Transcription Factors: Molecular Regulation and Stress Responses in Plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [PubMed]
- Besseau, S.; Li, J.; Palva, E.T. WRKY54 and WRKY70 co-operate as negative regulators of leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 2667–2679. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.J.; Yan, J.Y.; Xu, X.Y.; Yu, D.Q.; Li, G.X.; Zhang, S.Q.; Zheng, S.J. Transcription factor WRKY46 regulates osmotic stress responses and stomatal movement independently in Arabidopsis. Plant J. 2014, 79, 13–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciftci-Yilmaz, S.; Morsy, M.R.; Song, L.; Coutu, A.; Krizek, B.A.; Lewis, M.W.; Warren, D.; Cushman, J.; Connolly, E.L.; Mittler, R. The EAR-motif of the Cys2/His2-type zinc finger protein Zat7 plays a key role in the defense response of Arabidopsis to salinity stress. J. Biol. Chem. 2007, 282, 9260–9268. [Google Scholar] [CrossRef] [PubMed]
- Le, C.T.T.; Brumbarova, T.; Ivanov, R.; Stoof, C.; Weber, E.; Mohrbacher, J.; Fink-Straube, C.; Bauer, P. ZINC FINGER OF ARABIDOPSIS THALIANA12 (ZAT12) Interacts with FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR (FIT) Linking Iron Deficiency and Oxidative Stress Responses. Plant Physiol. 2016, 170, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Pavet, V.; Olmos, E.; Kiddle, G.; Mowla, S.; Kumar, S.; Antoniw, J.; Alvarez, M.E.; Foyer, C.H. Ascorbic Acid Deficiency Activates Cell Death and Disease Resistance Responses in Arabidopsis. Plant Physiol. 2005, 139, 1291–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scrase-Field, S.A.M.G.; Knight, M.R. Calcium: Just a chemical switch? Curr. Opin. Plant Biol. 2003, 6, 500–506. [Google Scholar] [CrossRef]
- Peiter, E. The Ever-Closer Union of Signals: Propagating Waves of Calcium and ROS Are Inextricably Linked. Plant Physiol. 2016, 172, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, Z.-M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandadi, K.K.; Misra, A.; Ren, S.; McKnight, T.D. BT2, a BTB protein, mediates multiple responses to nutrients, stresses and hormones in Arabidopsis. Plant Physiol. 2009, 150, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Vierstra, R.D. The Cullin-RING Ubiquitin-Protein Ligases. Annu. Rev. Plant Biol. 2011, 62, 299–334. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.; Brown, I.; Adams, S.; Knight, M.; Ainslie, A.; Mansfield, J. The RPM1 plant disease resistance gene facilitates a rapid and sustained increase in cytosolic calcium that is necessary for the oxidative burst and hypersensitive cell death. Plant J. 2000, 23, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderauwera, S.; Zimmermann, P.; Rombauts, S.; Vandenabeele, S.; Langebartels, C.; Gruissem, W.; Inzé, D.; Van Breusegem, F. Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis. Plant Physiol. 2005, 139, 806–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desikan, R.; Hancock, J.T.; Bright, J.; Harrison, J.; Weir, I.; Hooley, R.; Neill, S.J. A role for ETR1 in hydrogen peroxide signaling in stomatal guard cells. Plant Physiol. 2005, 137, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Tognetti, V.B.; Van Aken, O.; Morreel, K.; Vandenbroucke, K.; van de Cotte, B.; De Clercq, I.; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of Indole-3-Butyric Acid Homeostasis by the UDP-Glucosyltransferase UGT74E2 Modulates Arabidopsis Architecture and Water Stress Tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wu, J.; Yuan, D.; Zhang, D.; Huang, Z.; Xiao, L.; Yang, C. Perturbation of auxin homeostasis caused by mitochondrial FtSH4 gene-mediated peroxidase accumulation regulates arabidopsis architecture. Mol. Plant. 2014, 7, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, A.; Baerenfaller, K.; Loscos, J.; Chételat, A.; Baginsky, S.; Farmer, E.E. Jasmonate controls polypeptide patterning in undamaged tissue in wounded Arabidopsis leaves. Plant Physiol. 2011, 156, 1797–1807. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Jedelský, P.L.; Novák, J.; Schlosser, A.; Brzobohatý, B. Cytokinin modulates proteomic, transcriptomic and growth responses to temperature shocks in Arabidopsis. Plant Cell Environ. 2014, 37, 1641–1655. [Google Scholar] [CrossRef] [PubMed]
- Žd’árská, M.; Zatloukalová, P.; Benítez, M.; Šedo, O.; Potěšil, D.; Novák, O.; Svačinová, J.; Pešek, B.; Malbeck, J.; Vašíčková, J.; et al. Proteome analysis in Arabidopsis reveals shoot- and root-specific targets of cytokinin action and differential regulation of hormonal homeostasis. Plant Physiol. 2013, 161, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Dai, S.; Zhu, N.; Booy, A.; Simons, B.; Yi, S.; Chen, S. Methyl jasmonate responsive proteins in Brassica napus guard cells revealed by iTRAQ-based quantitative proteomics. J. Proteome Res. 2012, 11, 3728–3742. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhu, N.; Song, W.; Harmon, A.C.; Assmann, S.M.; Chen, S. Thiol-based redox proteins in abscisic acid and methyl jasmonate signaling in Brassica napus guard cells. Plant J. 2014, 78, 491–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Blackburn, K.; Lin, Y.; Goshe, M.B.; Williamson, J.D. Absolute protein quantification by LC/MS(E) for global analysis of salicylic acid-induced plant protein secretion responses. J. Proteome Res. 2009, 8, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Comparison of plasma membrane proteomic changes of Arabidopsis suspension-cultured cells (T87 Line) after cold and ABA treatment in association with freezing tolerance development. Plant Cell Physiol. 2012, 53, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Lochmanová, G.; Zdráhal, Z.; Konečná, H.; Koukalová, Š.; Malbeck, J.; Souček, P.; Válková, M.; Kiran, N.S.; Brzobohatý, B. Cytokinin-induced photomorphogenesis in dark-grown Arabidopsis: A. proteomic analysis. J. Exp. Bot. 2008, 59, 3705–3719. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Bertini, L.; Timperio, A.M.; Zolla, L.; Caporale, C.; Caruso, C. Crosstalk between salicylic acid and jasmonate in Arabidopsis investigated by an integrated proteomic and transcriptomic approach. Mol. Biosyst. 2013, 9, 1169–1187. [Google Scholar] [CrossRef] [PubMed]
- Böhmer, M.; Schroeder, J.I. Quantitative transcriptomic analysis of abscisic acid-induced and reactive oxygen species-dependent expression changes and proteomic profiling in Arabidopsis suspension cells. Plant J. 2011, 67, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Dai, S.Y.; Yuan, J.S. Integration of shot-gun proteomics and bioinformatics analysis to explore plant hormone responses. BMC Bioinform. 2012, 13, S8. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Xue, H. A proteomics study of auxin effects in Arabidopsis thaliana. Acta Biochim. Biophys. Sin. (Shanghai) 2012, 44, 783–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueckershoff, K.; Mueller, S.; Mueller, M.J.; Reinders, J. Impact of cyclopentenone-oxylipins on the proteome of Arabidopsis thaliana. Biochim. Biophys. Acta 2008, 1784, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Dyčka, F.; Bobál′ová, J.; Brzobohatý, B. Early cytokinin response proteins and phosphoproteins of Arabidopsis thaliana identified by proteome and phosphoproteome profiling. J. Exp. Bot. 2011, 62, 921–937. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Belghazi, M.; Huguet, R.; Robin, C.; Moreau, A.; Job, C.; Job, D. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol. 2006, 141, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Czarnecki, O.; Chourey, K.; Yang, J.; Tuskan, G.A.; Hurst, G.B.; Pan, C.; Chen, J.-G. Strigolactone-regulated proteins revealed by iTRAQ-based quantitative proteomics in Arabidopsis. J. Proteome Res. 2014, 13, 1359–1372. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, T.; Yasuda, D.; Mori, T.; Yoshimitsu, Y.; Nakamura, Y.; Yoshida, S.; Asami, T.; Okamoto, S.; Matsuo, T. Characterization of brassinosteroid-regulated proteins in a nuclear-enriched fraction of Arabidopsis suspension-cultured cells. Plant Physiol. Biochem. 2011, 49, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xue, L.; Batelli, G.; Lee, S.; Hou, Y.-J.; Van Oosten, M.J.; Zhang, H.; Tao, W.A.; Zhu, J.-K. Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action. Proc. Natl. Acad. Sci. USA 2013, 110, 11205–11210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jourdan, N.; Martino, C.F.; El-Esawi, M.; Witczak, J.; Bouchet, P.-E.; d’Harlingue, A.; Ahmad, M. Blue-light dependent ROS formation by Arabidopsis cryptochrome-2 may contribute toward its signaling role. Plant Signal. Behav. 2015, 10, e1042647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.; Arthaut, L.-D.; Jourdan, N.; d’Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.-E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-light dependent reactive oxygen species formation by Arabidopsis cryptochrome may define a novel evolutionarily conserved signaling mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Kim, J.-H.; Kim, S.-G.; Sim, H.-J.; Lee, G.; Halitschke, R.; Baldwin, I.T.; Kim, J.-I.; Park, C.-M. Shoot phytochrome B modulates reactive oxygen species homeostasis in roots via abscisic acid signaling in Arabidopsis. Plant J. 2018, 94, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Debeaujon, I. Seed longevity: Survival and maintenance of high germination ability of dry seeds. C. R. Biol. 2008, 331, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; He, D.; Gupta, R.; Yang, P. Physiological and proteomic analyses on artificially aged Brassica napus seed. Front. Plant Sci. 2015, 6, 112. [Google Scholar] [CrossRef] [PubMed]
- KOCSY, G. Die or survive? Redox changes as seed viability markers. Plant Cell Environ. 2015, 38, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.-P.; Cueff, G.; Hegedus, D.D.; Rajjou, L.; Bentsink, L. A role for seed storage proteins in Arabidopsis seed longevity. J. Exp. Bot. 2015, 66, 6399–6413. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.; Garcia, Q. Reactive oxygen species and seed germination. Biologia 2013, 68, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, B.; Xu, Z.; Shi, Z.; Chen, S.; Huang, X.; Chen, J.; Wang, X. Involvement of reactive oxygen species in endosperm cap weakening and embryo elongation growth during lettuce seed germination. J. Exp. Bot. 2014, 65, 3189–3200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oracz, K.; Karpiński, S. Phytohormones Signaling Pathways and ROS Involvement in Seed Germination. Front. Plant Sci. 2016, 7, 864. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Kasa, S.; Sakamoto, M.; Aoki, N.; Watabe, G.; Yuasa, T.; Iwaya-Inoue, M.; Ishibashi, Y. Role of reactive oxygen species produced by NADPH oxidase in gibberellin biosynthesis during barley seed germination. Plant Signal. Behav. 2016, 11, e1180492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, Y.; Aoki, N.; Kasa, S.; Sakamoto, M.; Kai, K.; Tomokiyo, R.; Watabe, G.; Yuasa, T.; Iwaya-Inoue, M. The Interrelationship between Abscisic Acid and Reactive Oxygen Species Plays a Key Role in Barley Seed Dormancy and Germination. Front. Plant Sci. 2017, 8, 275. [Google Scholar] [CrossRef] [PubMed]
- Lariguet, P.; Ranocha, P.; De Meyer, M.; Barbier, O.; Penel, C.; Dunand, C. Identification of a hydrogen peroxide signalling pathway in the control of light-dependent germination in Arabidopsis. Planta 2013, 238, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different Modes of Hydrogen Peroxide Action During Seed Germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Barba-Espín, G.; Hernández, J.A.; Diaz-Vivancos, P. Role of H2O2 in pea seed germination. Plant Signal. Behav. 2012, 7, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Bazin, J.; Langlade, N.; Vincourt, P.; Arribat, S.; Balzergue, S.; El-Maarouf-Bouteau, H.; Bailly, C. Targeted mRNA Oxidation Regulates Sunflower Seed Dormancy Alleviation during Dry After-Ripening. Plant Cell 2011, 23, 2196–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Maarouf-Bouteau, H.; Meimoun, P.; Job, C.; Job, D.; Bailly, C. Role of protein and mRNA oxidation in seed dormancy and germination. Front. Plant Sci. 2013, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, Y.; Kasa, S.; Sakamoto, M.; Aoki, N.; Kai, K.; Yuasa, T.; Hanada, A.; Yamaguchi, S.; Iwaya-Inoue, M. A Role for Reactive Oxygen Species Produced by NADPH Oxidases in the Embryo and Aleurone Cells in Barley Seed Germination. PLoS ONE 2015, 10, e0143173. [Google Scholar] [CrossRef] [PubMed]
- Aoki, N.; Ishibashi, Y.; Kai, K.; Tomokiyo, R.; Yuasa, T.; Iwaya-Inoue, M. Programmed cell death in barley aleurone cells is not directly stimulated by reactive oxygen species produced in response to gibberellin. J. Plant Physiol. 2014, 171, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, Y.; Tawaratsumida, T.; Kondo, K.; Kasa, S.; Sakamoto, M.; Aoki, N.; Zheng, S.-H.; Yuasa, T.; Iwaya-Inoue, M. Reactive Oxygen Species Are Involved in Gibberellin/Abscisic Acid Signaling in Barley Aleurone Cells. Plant Physiol. 2012, 158, 1705–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farouk, S.; Abdul Qados, A.M.S. Enhancing seed quality and productivity as well as physio-anatomical responses of pea plants by folic acid and/or hydrogen peroxide application. Sci. Hortic. (Amsterdam) 2018, 240, 29–37. [Google Scholar] [CrossRef]
- Bouallègue, A.; Souissi, F.; Nouairi, I.; Souibgui, M.; Abbes, Z.; Mhadhbi, H. Salicylic acid and hydrogen peroxide pretreatments alleviate salt stress in faba bean (Vicia faba) seeds during germination. Seed Sci. Technol. 2017, 45, 675–690. [Google Scholar] [CrossRef]
- Li, B.; Cai, Q.; Ma, S.; Li, S.; Zhang, X.; Yu, Y. Regulation of NPA and ACC on H2O2-Induced Pea Primary Horizontal Bending Root. J. Plant Growth Regul. 2018, 37, 246–254. [Google Scholar] [CrossRef]
- Carol, R.J.; Takeda, S.; Linstead, P.; Durrant, M.C.; Kakesova, H.; Derbyshire, P.; Drea, S.; Zarsky, V.; Dolan, L. A RhoGDP dissociation inhibitor spatially regulates growth in root hair cells. Nature 2005, 438, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Gapper, C.; Kaya, H.; Bell, E.; Kuchitsu, K.; Dolan, L. Local positive feedback regulation determines cell shape in root hair cells. Science 2008, 319, 1241–1244. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Wang, Q.; Guo, J.; Wang, P.; Song, P.; Jia, Q.; Zhang, X.; Kudla, J.; Zhang, W.; Zhang, Q. Peroxisomal CuAOζ and its product H2O2 regulate the distribution of auxin and IBA-dependent lateral root development in Arabidopsis. J. Exp. Bot. 2017, 68, 4851–4867. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Liu, L.; Liu, H.; Ferguson, B.J.; Zou, Y.; Zhao, Y.; Wang, T.; Wang, Y.; Li, X. H2O2 regulates root system architecture by modulating the polar transport and redistribution of auxin. J. Plant Biol. 2016, 59, 260–270. [Google Scholar] [CrossRef]
- Takáč, T.; Obert, B.; Rolčík, J.; Šamaj, J. Improvement of adventitious root formation in flax using hydrogen peroxide. New Biotechnol. 2016, 33, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gu, Q.; Yu, X.; Huang, L.; Xu, S.; Wang, R.; Shen, W.; Shen, W. Hydrogen peroxide acts downstream of melatonin to induce lateral root formation. Ann. Bot. 2018, 121, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Yang, Y.; Fu, G.; Tao, L. Novel roles of hydrogen peroxide (H2O2) in regulating pectin synthesis and demethylesterification in the cell wall of rice (Oryza sativa) root tips. New Phytol. 2015, 206, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Ivanchenko, M.G.; den Os, D.; Monshausen, G.B.; Dubrovsky, J.G.; Bednárová, A.; Krishnan, N. Auxin increases the hydrogen peroxide (H2O2) concentration in tomato (Solanum lycopersicum) root tips while inhibiting root growth. Ann. Bot. 2013, 112, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Hou, H.; Yang, T.; Lian, Y.; Sun, Y.; Bian, Z.; Wang, C. Exogenous hydrogen peroxide inhibits primary root gravitropism by regulating auxin distribution during Arabidopsis seed germination. Plant Physiol. Biochem. PPB 2018, 128, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Sun, L.; Song, Y.; Wang, L.; Liu, L.; Zhang, L.; Liu, B.; Li, N.; Miao, C.; Hao, F. AtrbohD and AtrbohF positively regulate abscisic acid-inhibited primary root growth by affecting Ca2+ signalling and auxin response of roots in Arabidopsis. J. Exp. Bot. 2013, 64, 4183–4192. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yan, J.; Yu, X.; Liang, Y.; Fang, L.; Scheller, H.V.; Zhang, A. The NADPH-oxidase AtRbohI plays a positive role in drought-stress response in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2017, 491, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; He, J.; Yang, X.; Li, X.; Luo, D.; Wei, C.; Ma, J.; Zhang, Y.; Yang, J.; Zhang, X. Glutathione-dependent induction of local and systemic defense against oxidative stress by exogenous melatonin in cucumber (Cucumis sativus L.). J. Pineal Res. 2016, 60, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Luo, Z.; Dong, H.; Eneji, A.E.; Li, W. H2O2 and ABA signaling are responsible for the increased Na+ efflux and water uptake in Gossypium hirsutum L. roots in the non-saline side under non-uniform root zone salinity. J. Exp. Bot. 2016, 67, 2247–2261. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Yoshioka, M.; Fukazawa, A.; Mori, H.; Nishizawa, N.K.; Tsutsumi, N.; Yoshioka, H.; Nakazono, M. An NADPH Oxidase RBOH Functions in Rice Roots during Lysigenous Aerenchyma Formation under Oxygen-Deficient Conditions. Plant Cell 2017, 29, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Mangano, S.; Denita-Juarez, S.P.; Choi, H.-S.; Marzol, E.; Hwang, Y.; Ranocha, P.; Velasquez, S.M.; Borassi, C.; Barberini, M.L.; Aptekmann, A.A.; et al. Molecular link between auxin and ROS-mediated polar growth. Proc. Natl. Acad. Sci. USA 2017, 114, 5289–5294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voxeur, A.; Höfte, H. Cell wall integrity signaling in plants: “To grow or not to grow that’s the question”. Glycobiology 2016, 26, 950–960. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-J.; Xia, X.-J.; Guo, X.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Yu, J.-Q. Apoplastic H2 O2 plays a critical role in axillary bud outgrowth by altering auxin and cytokinin homeostasis in tomato plants. New Phytol. 2016, 211, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, F.; Xiang, X.; Ahammed, G.J.; Wang, M.; Onac, E.; Zhou, J.; Xia, X.; Shi, K.; Yin, X.; et al. Systemic Induction of Photosynthesis via Illumination of the Shoot Apex Is Mediated Sequentially by Phytochrome, B., Auxin and Hydrogen Peroxide in Tomato. Plant Physiol. 2016, 172, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C. Leaf epinasty and auxin: A biochemical and molecular overview. Plant Sci. 2016, 253, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Zandalinas, S.I.; Gómez-Cadenas, A.; Blumwald, E.; Mittler, R. Coordinating the overall stomatal response of plants: Rapid leaf-to-leaf communication during light stress. Sci. Signal. 2018, 11, eaam9514. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Shang, Y.; Nam, K.H. Brassinosteroids modulate ABA-induced stomatal closure in Arabidopsis. J. Exp. Bot. 2016, 67, 6297–6308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.-J.; Gao, C.-J.; Song, L.-X.; Zhou, Y.-H.; Shi, K.; Yu, J.-Q. Role of H2O2 dynamics in brassinosteroid-induced stomatal closure and opening in Solanum lycopersicum. Plant Cell Environ. 2014, 37, 2036–2050. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Zhang, Y.; Li, C.; Liu, Z.; Yang, N.; Pan, L.; Wu, J.; Wang, J.; Yang, J.; Lv, Y.; et al. Strigolactone-triggered stomatal closure requires hydrogen peroxide synthesis and nitric oxide production in an abscisic acid-independent manner. New Phytol. 2018, 217, 290–304. [Google Scholar] [CrossRef] [PubMed]
- Khokon, M.A.R.; Salam, M.A.; Jammes, F.; Ye, W.; Hossain, M.A.; Okuma, E.; Nakamura, Y.; Mori, I.C.; Kwak, J.M.; Murata, Y. MPK9 and MPK12 function in SA-induced stomatal closure in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2017, 81, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Ehonen, S.; Yarmolinsky, D.; Kollist, H.; Kangasjärvi, J. Reactive Oxygen Species, Photosynthesis and Environment in the Regulation of Stomata. Antioxid. Redox Signal. 2017, ars.2017.7455. [Google Scholar] [CrossRef]
- Ge, X.-M.; Cai, H.-L.; Lei, X.; Zhou, X.; Yue, M.; He, J.-M. Heterotrimeric G protein mediates ethylene-induced stomatal closure via hydrogen peroxide synthesis in Arabidopsis. Plant J. 2015, 82, 138–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.; Qi, C.; Ren, H.; Huang, A.; Hei, S.; She, X. Ethylene mediates brassinosteroid-induced stomatal closure via Gα protein-activated hydrogen peroxide and nitric oxide production in Arabidopsis. Plant J. 2015, 82, 280–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, S.; Wang, Z.; He, L.; Xu, K.; Wang, G. Glucose triggers stomatal closure mediated by basal signaling through HXK1 and PYR/RCAR receptors in Arabidopsis. J. Exp. Bot. 2018, 69, 1471–1484. [Google Scholar] [CrossRef] [PubMed]
- Nazareno, A.L.; Hernandez, B.S. A mathematical model of the interaction of abscisic acid, ethylene and methyl jasmonate on stomatal closure in plants. PLoS ONE 2017, 12, e0171065. [Google Scholar] [CrossRef] [PubMed]
- Dubovskaya, L.V.; Bakakina, Y.S.; Kolesneva, E.V.; Sodel, D.L.; Mcainsh, M.R.; Hetherington, A.M.; Volotovski, I.D. cGMP-dependent ABA-induced stomatal closure in the ABA-insensitive Arabidopsis mutant abi1-1. New Phytol. 2011, 191, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Y.; Miao, W.; Piao, T.; Hao, Y.; Hao, F.-S. NADK2 positively modulates abscisic acid-induced stomatal closure by affecting accumulation of H2O2, Ca2+ and nitric oxide in Arabidopsis guard cells. Plant Sci. 2017, 262, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Agurla, S.; Gayatri, G.; Raghavendra, A.S. Polyamines increase nitric oxide and reactive oxygen species in guard cells of Arabidopsis thaliana during stomatal closure. Protoplasma 2018, 255, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Jardim-Messeder, D.; Caverzan, A.; Rauber, R.; Cunha, J.R.; Carvalho, F.E.L.; Gaeta, M.L.; da Fonseca, G.C.; Costa, J.M.; Frei, M.; Silveira, J.A.G.; et al. Thylakoidal APX modulates hydrogen peroxide content and stomatal closure in rice (Oryza sativa L.). Environ. Exp. Bot. 2018, 150, 46–56. [Google Scholar] [CrossRef]
- Mao, X.; Zheng, Y.; Xiao, K.; Wei, Y.; Zhu, Y.; Cai, Q.; Chen, L.; Xie, H.; Zhang, J. OsPRX2 contributes to stomatal closure and improves potassium deficiency tolerance in rice. Biochem. Biophys. Res. Commun. 2018, 495, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Kaya, H.; Kawarazaki, T.; Hiraoka, G.; Senzaki, E.; Michikawa, M.; Kuchitsu, K. Protein phosphorylation is a prerequisite for the Ca2+-dependent activation of Arabidopsis NADPH oxidases and may function as a trigger for the positive feedback regulation of Ca2+ and reactive oxygen species. Biochim. Biophys. Acta 2012, 1823, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Sirichandra, C.; Gu, D.; Hu, H.-C.; Davanture, M.; Lee, S.; Djaoui, M.; Valot, B.; Zivy, M.; Leung, J.; Merlot, S.; et al. Phosphorylation of the Arabidopsis AtrbohF NADPH oxidase by OST1 protein kinase. FEBS Lett. 2009, 583, 2982–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, O.; Reshetnyak, G.; Grondin, A.; Saijo, Y.; Leonhardt, N.; Maurel, C.; Verdoucq, L. Aquaporins facilitate hydrogen peroxide entry into guard cells to mediate ABA- and pathogen-triggered stomatal closure. Proc. Natl. Acad. Sci. USA 2017, 114, 9200–9205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, S.M.; Jegla, T. Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid and CO2. Curr. Opin. Plant Biol. 2016, 33, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; Nietzel, T.; Di Fino, L.M.; Meyer, A.J.; Lamattina, L.; Schwarzländer, M.; Laxalt, A.M.; García-Mata, C. Hydrogen Sulfide Increases Production of NADPH Oxidase-Dependent Hydrogen Peroxide and Phospholipase D-Derived Phosphatidic Acid in Guard Cell Signaling. Plant Physiol. 2018, 176, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.M.; Chapman, J.M.; Muday, G.K. Abscisic Acid-Induced Reactive Oxygen Species Are Modulated by Flavonols to Control Stomata Aperture. Plant Physiol. 2017, 175, 1807–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Feng, X.; Liu, L.; Xiong, L.; Wang, L. ALA-Induced Flavonols Accumulation in Guard Cells Is Involved in Scavenging H2O2 and Inhibiting Stomatal Closure in Arabidopsis Cotyledons. Front. Plant Sci. 2016, 7, 1713. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Liu, L.; Chen, L.; Wang, L. ALA Inhibits ABA-induced Stomatal Closure via Reducing H2O2 and Ca(2+) Levels in Guard Cells. Front. Plant Sci. 2016, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Serrano, I.; Romero-Puertas, M.C.; Sandalio, L.M.; Olmedilla, A. The role of reactive oxygen species and nitric oxide in programmed cell death associated with self-incompatibility. J. Exp. Bot. 2015, 66, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassig, R.; Gutermuth, T.; Bey, T.D.; Konrad, K.R.; Romeis, T. Pollen tube NAD(P)H oxidases act as a speed control to dampen growth rate oscillations during polarized cell growth. Plant J. 2014, 78, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, H.; Nakajima, R.; Iwano, M.; Kanaoka, M.M.; Kimura, S.; Takeda, S.; Kawarazaki, T.; Senzaki, E.; Hamamura, Y.; Higashiyama, T.; et al. Ca2+-activated reactive oxygen species production by Arabidopsis RbohH and RbohJ is essential for proper pollen tube tip growth. Plant Cell 2014, 26, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Kita, D.; Johnson, E.A.; Aggarwal, M.; Gates, L.; Wu, H.-M.; Cheung, A.Y. Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis. Nat. Commun. 2014, 5, 3129. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Jiang, L.; An, X.; Yu, M.; Xu, Y.; Ma, R.; Yu, Z. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol. Biochem. PPB 2016, 104, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Irfan, M.; Ghosh, S.; Chakraborty, N.; Chakraborty, S.; Datta, A. Fruit ripening mutants reveal cell metabolism and redox state during ripening. Protoplasma 2016, 253, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Hurr, B.M.; Huber, D.J.; Vallejos, C.E.; Lee, E.; Sargent, S.A. Ethylene-induced overproduction of reactive oxygen species is responsible for the development of watersoaking in immature cucumber fruit. J. Plant Physiol. 2013, 170, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef] [PubMed]
- Bieker, S.; Riester, L.; Stahl, M.; Franzaring, J.; Zentgraf, U. Senescence-specific Alteration of Hydrogen Peroxide Levels in Arabidopsis thaliana and Oilseed Rape Spring Variety Brassica napus L. cv. MozartF. J. Integr. Plant Biol. 2012, 54, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lin, J.; Chang, Y.; Jiang, C.-Z. Comparative Transcriptomic Analysis Reveals That Ethylene/ H2O2-Mediated Hypersensitive Response and Programmed Cell Death Determine the Compatible Interaction of Sand Pear and Alternaria alternata. Front. Plant Sci. 2017, 8, 195. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, Y.; Zhang, Z.; Wei, J. Hydrogen peroxide promotes programmed cell death and salicylic acid accumulation during the induced production of sesquiterpenes in cultured cell suspensions of Aquilaria sinensis. Funct. Plant Biol. 2015, 42, 337–346. [Google Scholar] [CrossRef]
- Zimmermann, P.; Heinlein, C.; Orendi, G.; Zentgraf, U. Senescence-specific regulation of catalases in Arabidopsis thaliana (L.) Heynh. Plant Cell Environ. 2006, 29, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Jajić, I.; Sarna, T.; Szewczyk, G.; Strzałka, K. Changes in production of reactive oxygen species in illuminated thylakoids isolated during development and senescence of barley. J. Plant Physiol. 2015, 184, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, E.; Polzien, L.; Desel, C.; Rozpadek, P.; Miszalski, Z.; Krupinska, K. Spatial patterns of senescence and development-dependent distribution of reactive oxygen species in tobacco (Nicotiana tabacum) leaves. J. Plant Physiol. 2009, 166, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross Talk between H2O2 and Interacting Signal Molecules under Plant Stress Response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Diao, Q.; Song, Y.; Shi, D.; Qi, H. Interaction of Polyamines, Abscisic Acid, Nitric Oxide and Hydrogen Peroxide under Chilling Stress in Tomato (Lycopersicon esculentum Mill.) Seedlings. Front. Plant Sci. 2017, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.-G.; Kong, C.-C.; Xie, Z.-H. Role of abscisic acid in strigolactone-induced salt stress tolerance in arbuscular mycorrhizal Sesbania cannabina seedlings. BMC Plant Biol. 2018, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Freitas, V.S.; de Souza Miranda, R.; Costa, J.H.; de Oliveira, D.F.; de Oliveira Paula, S.; de Castro Migueld, E.; Freire, R.S.; Prisco, J.T.; Gomes-Filho, E. Ethylene triggers salt tolerance in maize genotypes by modulating polyamine catabolism enzymes associated with H2O2 production. Environ. Exp. Bot. 2018, 145, 75–86. [Google Scholar] [CrossRef]
- 224 Kaur, N.; Kirat, K.; Saini, S.; Sharma, I.; Gantet, P.; Pati, P.K. Reactive oxygen species generating system and brassinosteroids are linked to salt stress adaptation mechanisms in rice. Plant Signal. Behav. 2016, 11, e1247136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-Y.; Huang, Y.-C.; Ou, S.-L. ERF73/HRE1 is involved in H2O2 production via hypoxia-inducible Rboh gene expression in hypoxia signaling. Protoplasma 2017, 254, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.H.; Deng, X.M.; He, J.B.; Song, L.; He, J.; Marchant, A.; Chen, X.-Y.; et al. ETHYLENE RESPONSE FACTOR 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD)-dependent mechanism in response to different stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Smith, J.A.C.; Harberd, N.P.; Jiang, C. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Mol. Biol. 2016, 91, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Jiang, F.; Cen, B.; Wen, J.; Zhou, Y.; Wu, Z. Respiratory burst oxidase homologue-dependent H2O2 and chloroplast H2O2 are essential for the maintenance of acquired thermotolerance during recovery after acclimation. Plant Cell Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Birch, P.R.J.; Avrova, A.O.; Dellagi, A.; Lacomme, C.; Cruz, S.S.; Lyon, G.D. Programmed Cell Death in Plants in Response to Pathogen Attack. In Annual Plant. Reviews; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 184–208. [Google Scholar]
- Novák, J.; Pavlů, J.; Novák, O.; Nožková-Hlaváčková, V.; Špundová, M.; Hlavinka, J.; Koukalová, Š.; Skalák, J.; Černý, M.; Brzobohatý, B. High cytokinin levels induce a hypersensitive-like response in tobacco. Ann. Bot. 2013, 112, 41–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nováková, M.; Šašek, V.; Trdá, L.; Krutinová, H.; Mongin, T.; Valentová, O.; Balesdent, M.-H.; Rouxel, T.; Burketová, L. L eptosphaeria maculans effector AvrLm4-7 affects salicylic acid (SA) and ethylene (ET) signalling and hydrogen peroxide (H2O2) accumulation in Brassica napus. Mol. Plant Pathol. 2016, 17, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Song, L.-X.; Xu, X.-C.; Wang, F.-N.; Wang, Y.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Zhou, J.; Yu, J.-Q. Brassinosteroids act as a positive regulator for resistance against root-knot nematode involving RESPIRATORY BURST OXIDASE HOMOLOG-dependent activation of MAPKs in tomato. Plant Cell Environ. 2018, 41, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.-G.; Zhu, T.; Zou, L.-J.; Han, X.-Y.; Zhou, X.; Xi, D.-H.; Zhang, D.-W.; Lin, H.-H. Orchestration of hydrogen peroxide and nitric oxide in brassinosteroid-mediated systemic virus resistance in Nicotiana benthamiana. Plant J. 2016, 85, 478–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Chen, J.; Xiao, X.; Zhang, M.; Yun, Z.; Zeng, Y.; Xu, J.; Cheng, Y.; Deng, X. Salicylic acid treatment reduces the rot of postharvest citrus fruit by inducing the accumulation of H2O2, primary metabolites and lipophilic polymethoxylated flavones. Food Chem. 2016, 207, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Ellouzi, H.; Sghayar, S.; Abdelly, C. H2O2 seed priming improves tolerance to salinity; drought and their combined effect more than mannitol in Cakile maritima when compared to Eutrema salsugineum. J. Plant Physiol. 2017, 210, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Wilkins, K.A.; Swarbreck, S.M.; Anderson, A.A.; Habib, N.; Smith, A.G.; McAinsh, M.; Davies, J.M. The hydroxyl radical in plants: From seed to seed. J. Exp. Bot. 2015, 66, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demidchik, V.; Shabala, S.N.; Coutts, K.B.; Tester, M.A.; Davies, J.M. Free oxygen radicals regulate plasma membrane Ca2+- and K+-permeable channels in plant root cells. J. Cell Sci. 2003, 116, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Laohavisit, A.; Shang, Z.; Rubio, L.; Cuin, T.A.; Véry, A.-A.; Wang, A.; Mortimer, J.C.; Macpherson, N.; Coxon, K.M.; Battey, N.H.; et al. Arabidopsis annexin1 mediates the radical-activated plasma membrane Ca2+- and K+-permeable conductance in root cells. Plant Cell 2012, 24, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shabala, S. Mechanisms of cytosolic calcium elevation in plants: The role of ion channels, calcium extrusion systems and NADPH oxidase-mediated “ROS-Ca2+ Hub”. Funct. Plant Biol. 2018, 45, 9. [Google Scholar] [CrossRef]
- Sun, J.; Wang, M.-J.; Ding, M.-Q.; Deng, S.-R.; Liu, M.-Q.; Lu, C.-F.; Zhou, X.-Y.; Shen, X.; Zheng, X.-J.; Zhang, Z.-K.; et al. H2O2 and cytosolic Ca2+ signals triggered by the PM H+-coupled transport system mediate K+/Na+ homeostasis in NaCl-stressed Populus euphratica cells. Plant Cell Environ. 2010, 33, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, Calcium and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, T.; Lehotai, N.; Strand, Å. The role of retrograde signals during plant stress responses. J. Exp. Bot. 2018, 69, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Pavlů, J.; Novák, J.; Koukalová, V.; Luklová, M.; Brzobohatý, B.; Černý, M. Cytokinin at the crossroad of abiotic stress signalling pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Pratsinis, A.; Kelesidis, G.A.; Zuercher, S.; Krumeich, F.; Bolisetty, S.; Mezzenga, R.; Leroux, J.-C.; Sotiriou, G.A. Enzyme-Mimetic Antioxidant Luminescent Nanoparticles for Highly Sensitive Hydrogen Peroxide Biosensing. ACS Nano 2017, 11, 12210–12218. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.J.; Gupta, A.; Barkam, S.; Saraf, S.; Das, S.; Cho, H.J.; Seal, S. Picomolar Detection of Hydrogen Peroxide using Enzyme-free Inorganic Nanoparticle-based Sensor. Sci. Rep. 2017, 7, 1324. [Google Scholar] [CrossRef] [PubMed]
- Shabrangy, A.; Roustan, V.; Reipert, S.; Weidinger, M.; Roustan, P.-J.; Stoger, E.; Weckwerth, W.; Ibl, V. Using RT-qPCR, Proteomics and Microscopy to Unravel the Spatio-Temporal Expression and Subcellular Localization of Hordoindolines Across Development in Barley Endosperm. Front. Plant Sci. 2018, 9, 775. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Key enzymes of hydrogen peroxide metabolism in plants. The list shows enzymes that directly catalyse hydrogen peroxide production or degradation in Arabidopsis, including the numbers of different isozymes, a comparison of relative gene expression profiles in seedlings, roots and shoots and the figure indicates subcellular localization. Colour coding: anabolic processes (green), catabolic processes (red), based on UniProt [16], SUBA 3.0 [17] and average gene expression profiles in 45, 24 and 7 NGS experiments for seedlings, leaf and root respectively (ThaleMine [18]).
Figure 1.
Key enzymes of hydrogen peroxide metabolism in plants. The list shows enzymes that directly catalyse hydrogen peroxide production or degradation in Arabidopsis, including the numbers of different isozymes, a comparison of relative gene expression profiles in seedlings, roots and shoots and the figure indicates subcellular localization. Colour coding: anabolic processes (green), catabolic processes (red), based on UniProt [16], SUBA 3.0 [17] and average gene expression profiles in 45, 24 and 7 NGS experiments for seedlings, leaf and root respectively (ThaleMine [18]).

Figure 2.
Hydrogen peroxide-mediated processes in plant growth and development. This figure summarizes our present-day knowledge about the role of H2O2 in the life of plants as described, with references, in Section 5. ABA—abscisic acid, AUX—auxin, BR—brassinosteroids, ET—ethylene, GA—gibberellins, SA—salicylic acid, JA—jasmonic acid, AOX—amine oxidases, PRX—peroxidases, RBOH—NADPH oxidases, PCD—programmed cell death. The water droplet shape indicates flooding and absence of water, for hypoxia and drought, respectively.
Figure 2.
Hydrogen peroxide-mediated processes in plant growth and development. This figure summarizes our present-day knowledge about the role of H2O2 in the life of plants as described, with references, in Section 5. ABA—abscisic acid, AUX—auxin, BR—brassinosteroids, ET—ethylene, GA—gibberellins, SA—salicylic acid, JA—jasmonic acid, AOX—amine oxidases, PRX—peroxidases, RBOH—NADPH oxidases, PCD—programmed cell death. The water droplet shape indicates flooding and absence of water, for hypoxia and drought, respectively.


{kind=link}
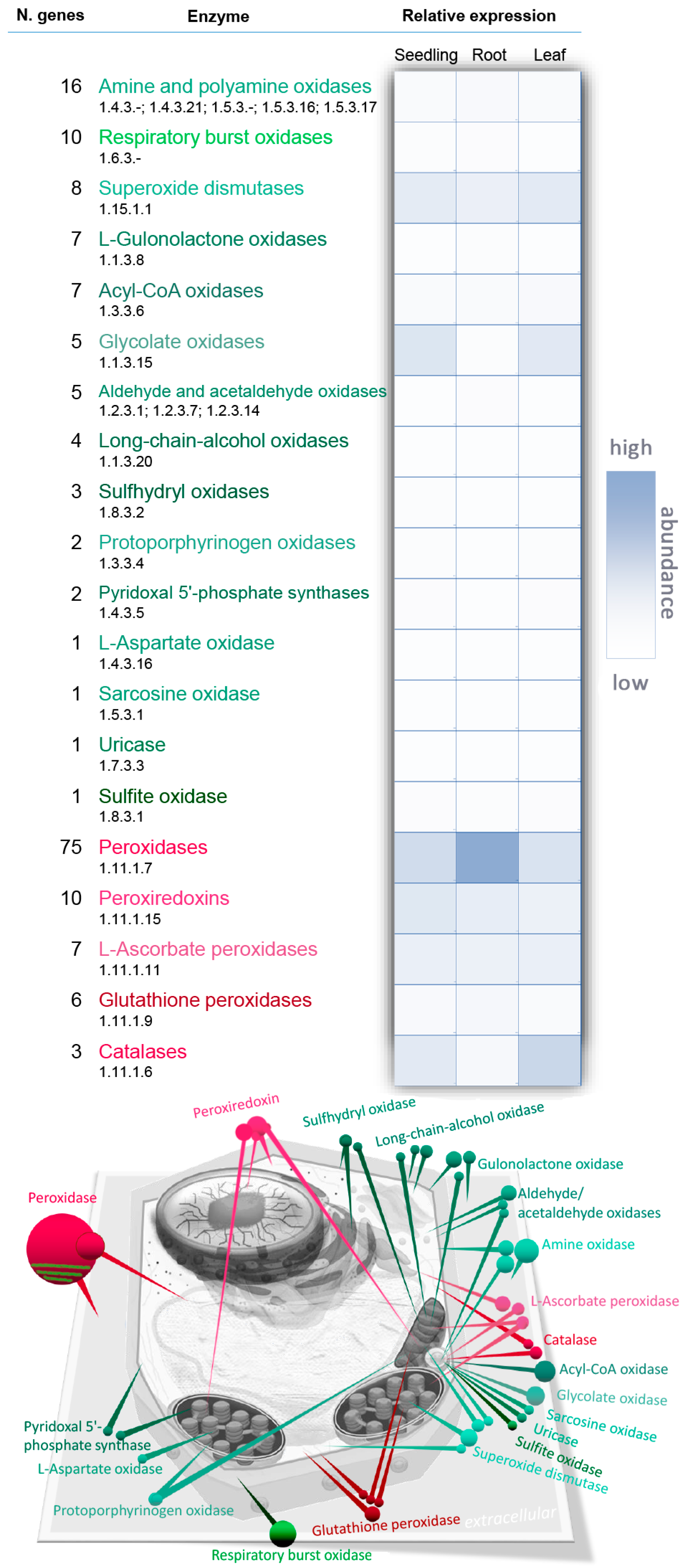{kind=link}
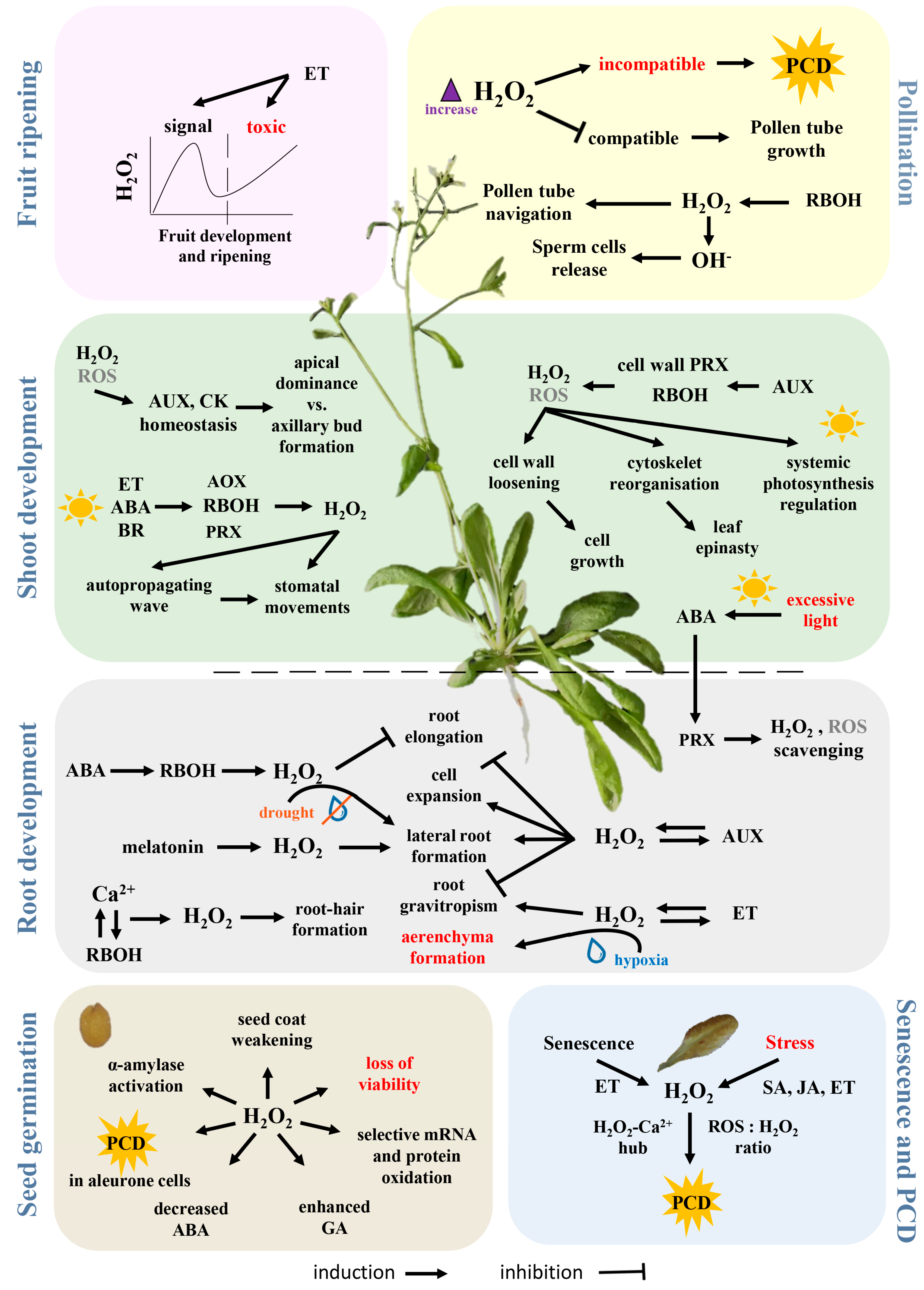{kind=link}
Table 1.
Proteins of hydrogen peroxide metabolism in Arabidopsis identified in phytohormone-responsive proteomics analyses. Based on a previously published overview of hormone-responsive proteins [90].
Table 1.
Proteins of hydrogen peroxide metabolism in Arabidopsis identified in phytohormone-responsive proteomics analyses. Based on a previously published overview of hormone-responsive proteins [90].
| AGI | Protein Name (UniProt) | Relative Protein Abundance | ||||||
|---|---|---|---|---|---|---|---|---|
| Auxin | Abscisic Acid | Brassinosteroid | Cytokinin | Salicylic Acid | Jasmonate/Oxylipins | Strigolactone | ||
| AT1G05260 | Peroxidase 3 | down [121] | ||||||
| AT1G06290 | Acyl-coenzyme A oxidase 3 | up [122] | ||||||
| AT1G07890 | L-Ascorbate peroxidase 1 | down [121] | down [30,123,124] | up [125,126] | ||||
| AT1G08830 | Superoxide dismutase [Cu-Zn] 1 | up [127] | ||||||
| AT1G20620 | Catalase-3 | up [128] | down [129] | up [130] | up [130] | |||
| AT1G20630 | Catalase-1 | up [131] | ||||||
| AT1G31710 | Amine oxidase | down [121] | ||||||
| AT1G44446 | Chlorophyllide a oxygenase | down [132] | ||||||
| AT1G65980 | Peroxiredoxin-2B | down [121] | up [126] | |||||
| AT1G71695 | Peroxidase 12 | down [121] | down [132] | |||||
| AT1G77490 | L-Ascorbate peroxidase T | up [133] | ||||||
| AT2G18150 | Peroxidase 15 | up [127] | ||||||
| AT2G22420 | Peroxidase 17 | up [127] | ||||||
| AT2G26230 | Uricase | down [121] | ||||||
| AT2G28190 | Superoxide dismutase [Cu-Zn] 2 | up [134] | ||||||
| AT2G30490 | Trans-cinnamate 4-monooxygenase | up [131] | ||||||
| AT2G43350 | Probable glutathione peroxidase 3 | down [121] | ||||||
| AT3G06050 | Peroxiredoxin-2F | up [134] | ||||||
| AT3G10920 | Superoxide dismutase [Mn] 1 | down [135] | up [136] | down [126] | ||||
| AT3G11630 | 2-Cys peroxiredoxin BAS1 | up [30,129] | up [125] | |||||
| AT3G14415 | (S)-2-hydroxy-acid oxidase | down [125] | up [130] | up [125,130] | ||||
| AT3G14420 | (S)-2-hydroxy-acid oxidase GLO1 | up [30] | up [130] | up [126,130] | ||||
| AT3G26060 | Peroxiredoxin Q, chloroplastic | up [134] | ||||||
| AT3G32980 | Peroxidase 32 | down [30] | up [127] | |||||
| AT3G49120 | Peroxidase 34 | up [128,131] | down [30] | up [127] | ||||
| AT3G56350 | Superoxide dismutase [Mn] 2 | up [137] | ||||||
| AT4G08390 | L-Ascorbate peroxidase S | up [126] | down [30] | |||||
| AT4G08770 | Peroxidase 37 | up [127] | ||||||
| AT4G08780 | Peroxidase 38 | up [127] | ||||||
| AT4G15760 | Monooxygenase 1 | up [137] | ||||||
| AT4G16760 | Acyl-coenzyme A oxidase 1 | up [133] | up [122] | |||||
| AT4G25100 | Superoxide dismutase [Fe] 1 | up [127] | up [125] | |||||
| AT4G35000 | L-Ascorbate peroxidase 3 | down [30] | ||||||
| AT4G35090 | Catalase-2 | up [131] | up/down [30,123] | up [130] | up [130] | |||
| AT4G36430 | Peroxidase 49 | up [127] | ||||||
| AT5G06290 | 2-Cys peroxiredoxin BAS1-like | up [126] | ||||||
| AT5G14220 | Protoporphyrinogen oxidase 2 | up [132] | up [132] | |||||
| AT5G17820 | Peroxidase 57 | up [128] | ||||||
| AT5G18100 | Superoxide dismutase [Cu-Zn] 3 | up [127] | ||||||
| AT5G23310 | Superoxide dismutase [Fe] 3 | down [138] | down [122] | |||||
| AT5G49970 | PYRIDOXINE/PYRIDOXAMINE 5′-PHOSPHATE OXIDASE 1 | up [122] | ||||||
| AT5G51100 | Superoxide dismutase [Fe] 2 | up [139] | ||||||
| AT5G64120 | Peroxidase 71 | down [131] | up [122] | down [137] | ||||
| AT5G65110 | Acyl-coenzyme A oxidase 2 | down [132] | ||||||
Table 2.
Hydrogen peroxide metabolism genes have expression patterns similar to those of genes related to light signalling, nutrient status, temperature stress, drought stress and hormonal metabolism. Based on average gene expression profiles in stress-related experiments (ThaleMine [11]) and reference stress-related genes [247]. Numbers indicate the number of analysed genes (numbers in brackets) and the number of detected co-expressed genes (hydrogen peroxide metabolism/candidate signalling and metabolism genes). See Supplementary Materials for the full list of co-expressed genes.
Table 2.
Hydrogen peroxide metabolism genes have expression patterns similar to those of genes related to light signalling, nutrient status, temperature stress, drought stress and hormonal metabolism. Based on average gene expression profiles in stress-related experiments (ThaleMine [11]) and reference stress-related genes [247]. Numbers indicate the number of analysed genes (numbers in brackets) and the number of detected co-expressed genes (hydrogen peroxide metabolism/candidate signalling and metabolism genes). See Supplementary Materials for the full list of co-expressed genes.
| Nutrient Stress (142) | Temperature Stress (43) | Drought Stress (13) | Light Signalling (27) | Abscisic Acid Metabolism (16) | Auxin Metabolism (31) | Brassinosteroid Metabolism (13) | Cytokinin Metabolism (37) | Ethylene Metabolism (12) | Gibberellin Metabolism (23) | Jasmonate Metabolism (17) | Salicylic Acid Metabolism (9) | Strigolactone Metabolism (3) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amine/polyamine oxidase (15) | 11/40 | 4/6 | 1/1 | 7/16 | 6/5 | 5/6 | 5/5 | 10/15 | 5/4 | 4/7 | 3/5 | 2/1 | 0/0 |
| Respiratory burst oxidase (10) | 10/58 | 6/6 | 4/1 | 9/16 | 5/3 | 7/14 | 8/8 | 9/21 | 4/4 | 8/6 | 3/2 | 4/4 | 4/3 |
| Superoxide dismutase (8) | 7/46 | 5/5 | 0/0 | 4/13 | 5/2 | 5/9 | 4/6 | 5/15 | 2/3 | 5/10 | 1/3 | 2/2 | 0/0 |
| L-Gulonolactone oxidase (7) | 7/31 | 2/2 | 2/1 | 4/5 | 4/5 | 4/7 | 2/1 | 7/10 | 3/3 | 3/5 | 3/4 | 3/2 | 2/2 |
| Acyl-coenzyme A oxidase (7) | 5/19 | 4/6 | 4/2 | 4/14 | 4/3 | 6/9 | 4/2 | 5/11 | 0/0 | 5/7 | 6/6 | 1/1 | 2/2 |
| Glycolate oxidase (5) | 5/10 | 2/1 | 0/0 | 5/8 | 0/0 | 3/3 | 4/2 | 3/4 | 0/0 | 2/1 | 3/2 | 0/0 | 3/1 |
| Aldehyde/acetaldehyde oxidase (5) | 4/36 | 2/1 | 2/1 | 4/9 | 3/3 | 5/8 | 4/4 | 4/13 | 1/1 | 3/7 | 1/2 | 3/3 | 1/2 |
| Long-chain-alcohol oxidase (4) | 3/18 | 3/4 | 0/0 | 2/7 | 2/2 | 3/5 | 1/1 | 3/6 | 1/1 | 1/1 | 1/2 | 2/2 | 0/0 |
| Sulfhydryl oxidase (3) | 3/25 | 3/5 | 0/0 | 3/13 | 3/2 | 3/8 | 3/3 | 3/5 | 2/2 | 3/6 | 3/3 | 2/1 | 1/1 |
| Protoporphyrinogen oxidase (2) | 2/21 | 2/1 | 0/0 | 2/5 | 2/2 | 2/6 | 2/3 | 2/6 | 2/1 | 2/3 | 0/0 | 1/2 | 0/0 |
| Pyridoxal 5′-phosphate synthase (2) | 2/28 | 2/3 | 0/0 | 2/9 | 2/2 | 2/7 | 2/3 | 2/8 | 2/2 | 2/5 | 1/1 | 2/2 | 1/1 |
| L-Aspartate oxidase (1) | 1/5 | 1/1 | 1/1 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1 | 0/0 | 0/0 | 1/1 |
| Sarcosine oxidase (1) | 1/1 | 0/0 | 0/0 | 1/1 | 0/0 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| Uricase (1) | 1/6 | 1/1 | 1/1 | 1/3 | 1/1 | 1/1 | 1/1 | 1/1 | 0/0 | 0/0 | 1/1 | 0/0 | 1/1 |
| Sulphite oxidase (1) | 1/12 | 1/3 | 0/0 | 1/9 | 1/1 | 1/5 | 1/1 | 1/4 | 1/1 | 1/3 | 1/1 | 1/1 | 0/0 |
| Peroxidase (73) | 53/106 | 21/9 | 9/2 | 33/25 | 21/7 | 35/21 | 29/13 | 43/30 | 21/6 | 27/15 | 17/8 | 18/7 | 10/3 |
| Peroxiredoxin (10) | 8/43 | 4/7 | 1/1 | 5/14 | 5/3 | 3/7 | 3/5 | 6/15 | 4/4 | 7/12 | 2/4 | 4/3 | 0/0 |
| L-Ascorbate peroxidase (7) | 6/44 | 6/7 | 2/2 | 6/16 | 3/2 | 4/10 | 5/7 | 6/16 | 3/4 | 6/9 | 4/3 | 3/2 | 2/2 |
| Glutathione peroxidase (6) | 5/26 | 3/6 | 0/0 | 3/10 | 1/1 | 5/8 | 3/3 | 3/6 | 3/2 | 3/3 | 1/4 | 1/1 | 2/2 |
| Catalase (3) | 3/13 | 2/2 | 1/1 | 2/13 | 1/1 | 3/5 | 2/3 | 3/8 | 0/0 | 3/3 | 2/4 | 0/0 | 2/1 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks. Int. J. Mol. Sci. 2018, 19, 2812. https://doi.org/10.3390/ijms19092812
AMA Style
Černý M, Habánová H, Berka M, Luklová M, Brzobohatý B. Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks. International Journal of Molecular Sciences. 2018; 19(9):2812. https://doi.org/10.3390/ijms19092812
Chicago/Turabian StyleČerný, Martin, Hana Habánová, Miroslav Berka, Markéta Luklová, and Břetislav Brzobohatý. 2018. "Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks" International Journal of Molecular Sciences 19, no. 9: 2812. https://doi.org/10.3390/ijms19092812
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.