Peiminine Protects against Lipopolysaccharide-Induced Mastitis by Inhibiting the AKT/NF-κB, ERK1/2 and p38 Signaling Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
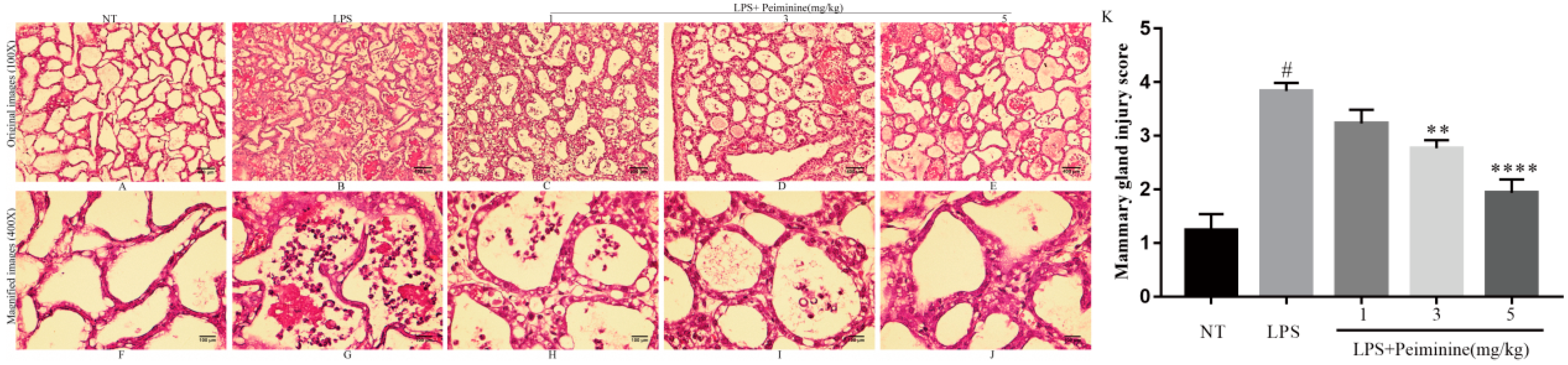
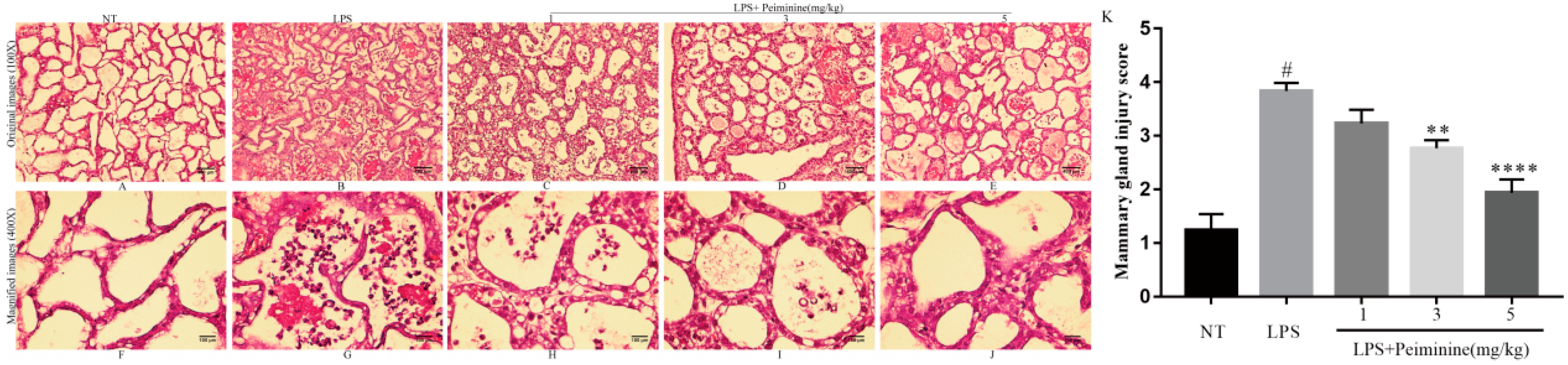
2.1. Effects of Peiminine on the LPS-Induced Histopathological Impairment of the Mammary Gland
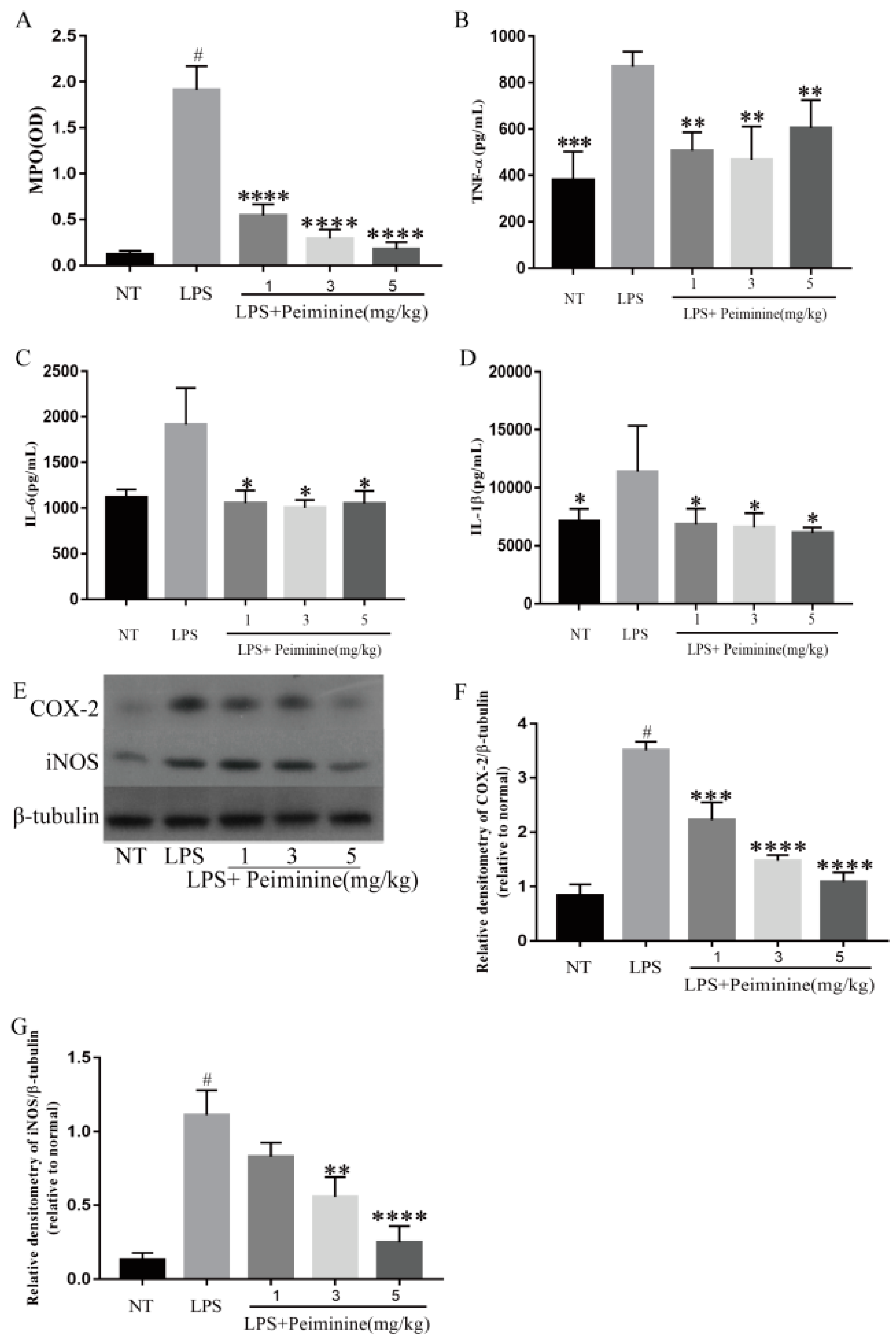
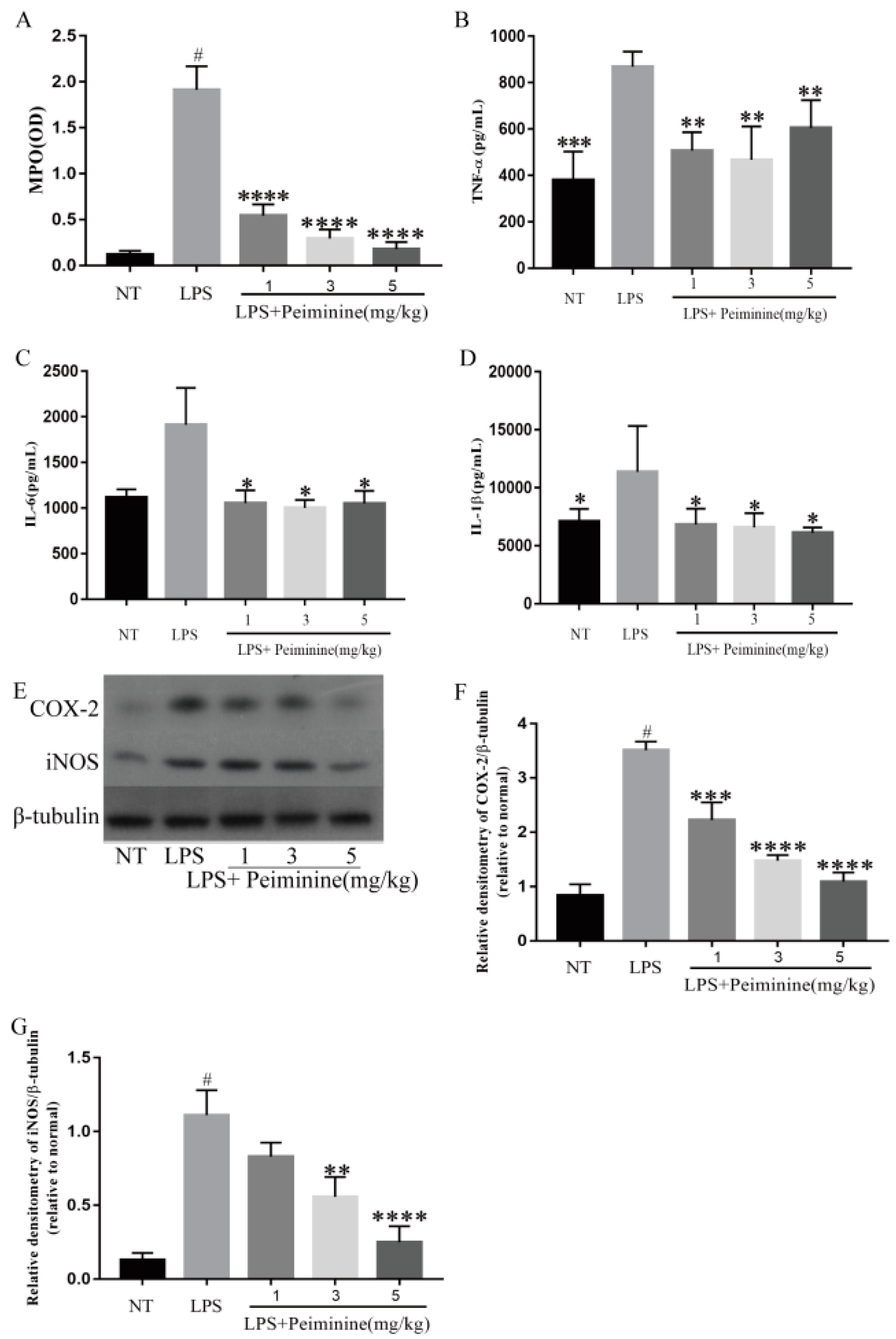
2.2. Effect of Peiminine on Myeloperoxidase (MPO) Activity and Pro-Inflammatory Mediators in the Mammary Gland of LPS-Induced Mastitis Model Mice
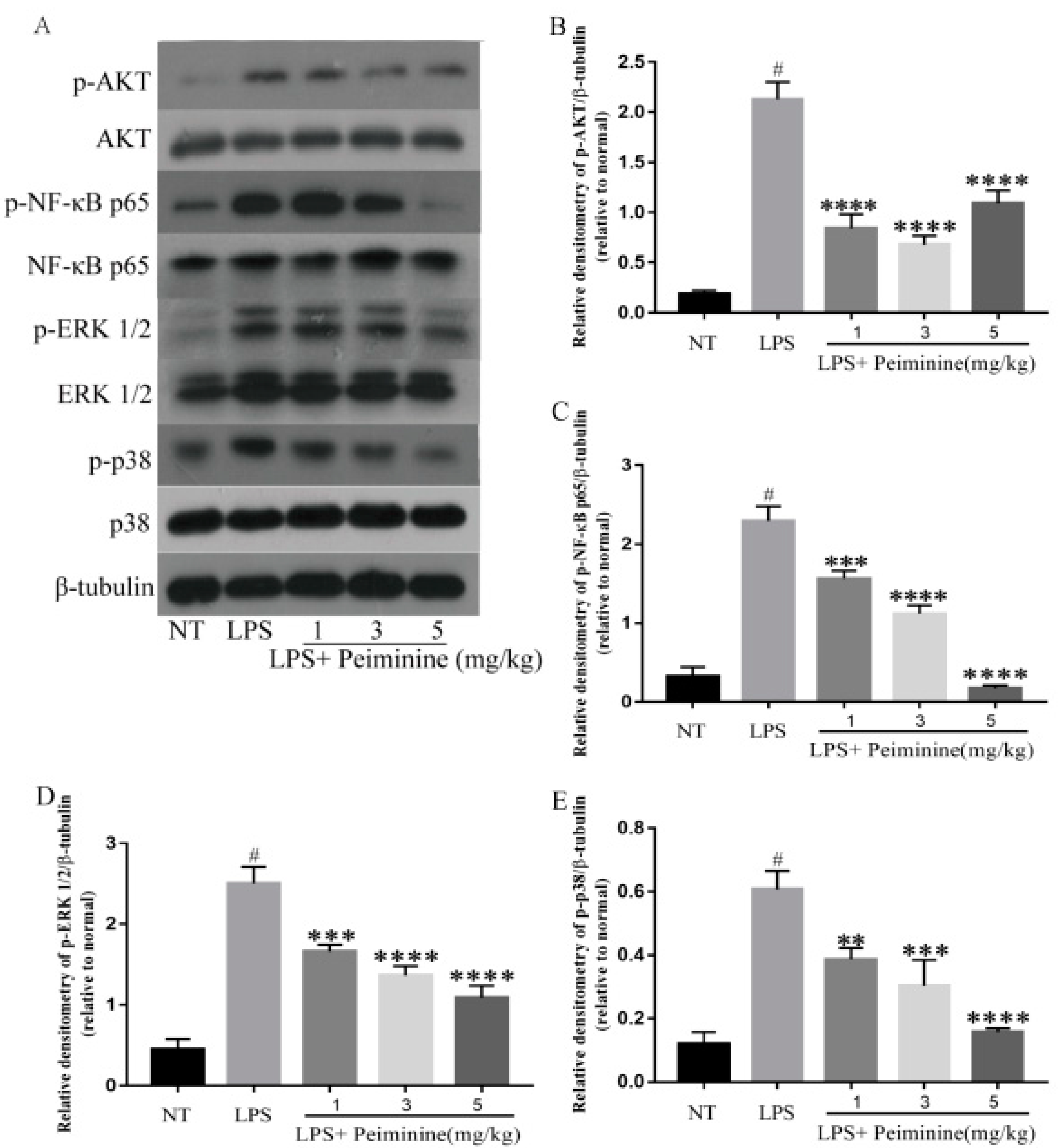
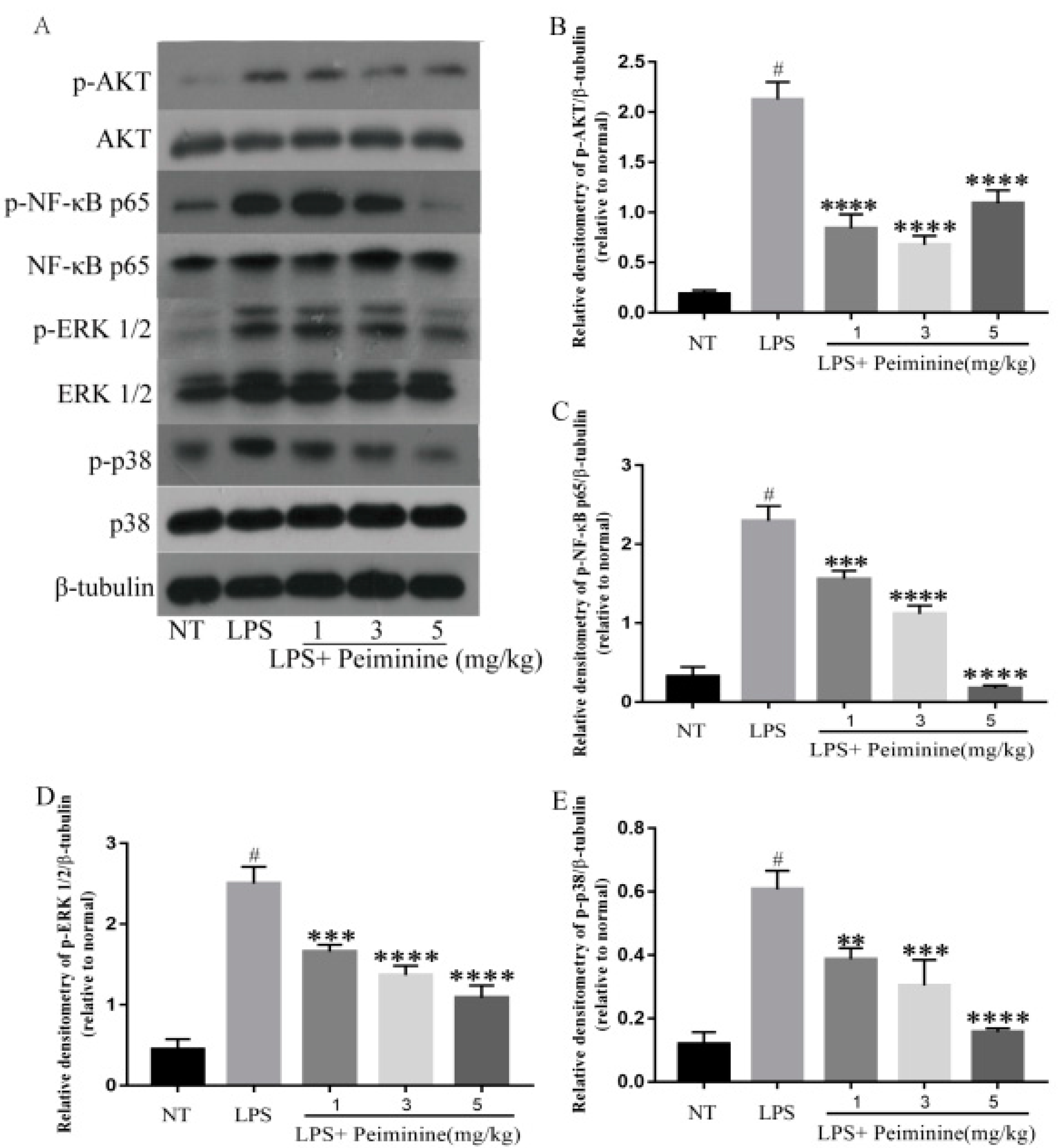
2.3. Effect of Peiminine on the LPS-Induced Activation of AKT, NF-κB p65, ERK1/2, and p38 in Mammary Tissues
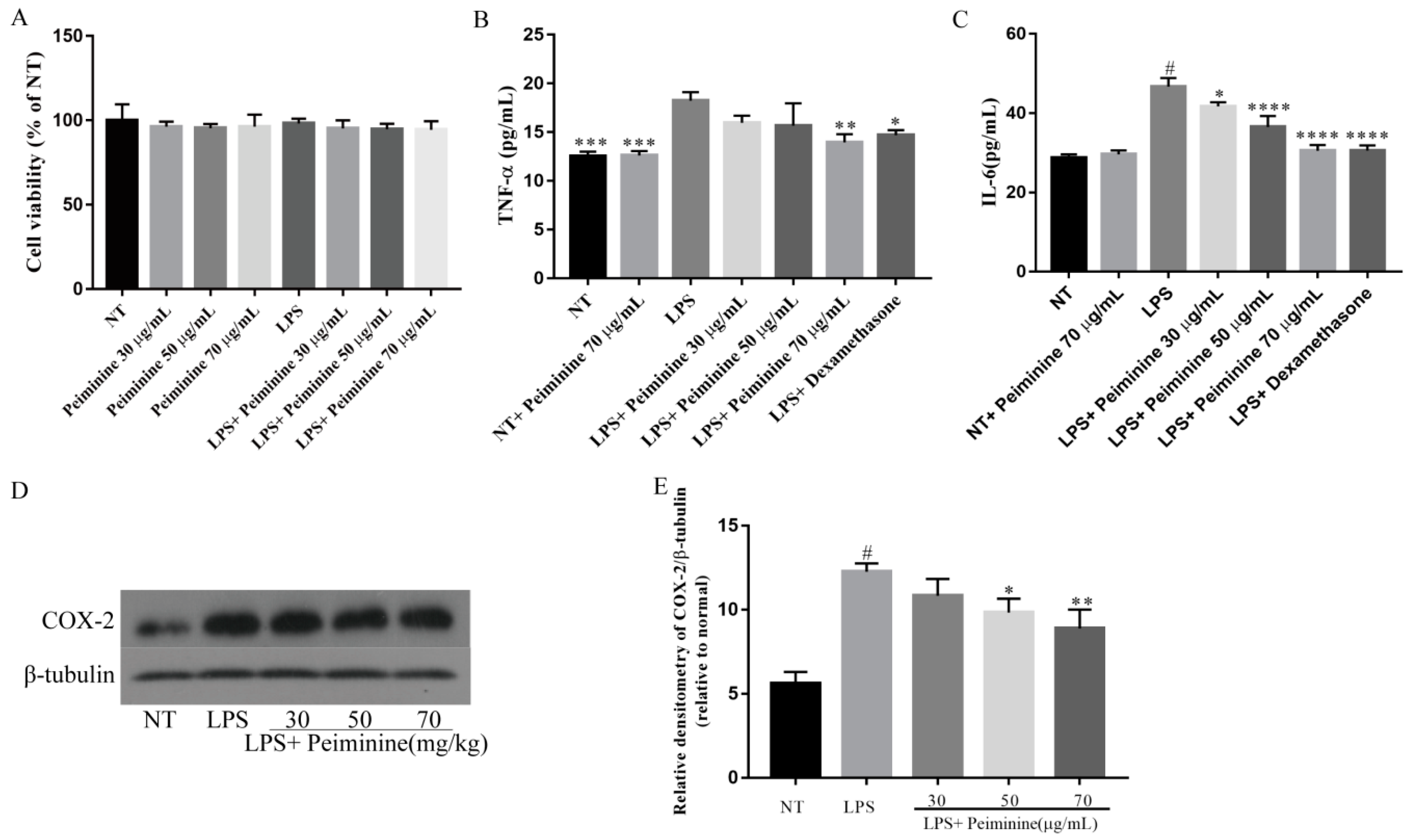
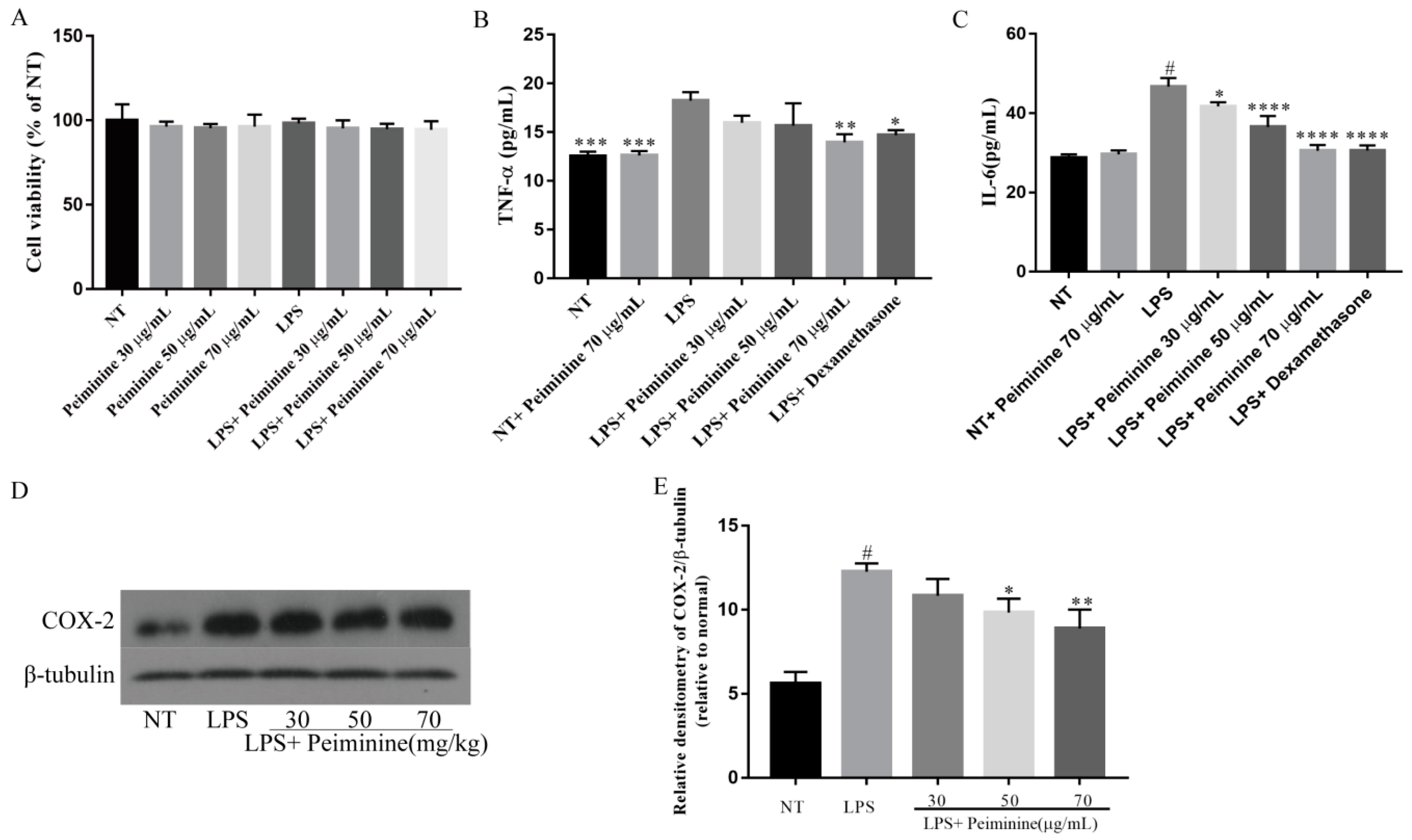
2.4. Effect of Peiminine on Cell Viability and LPS-Induced Inflammatory Response in mMECs
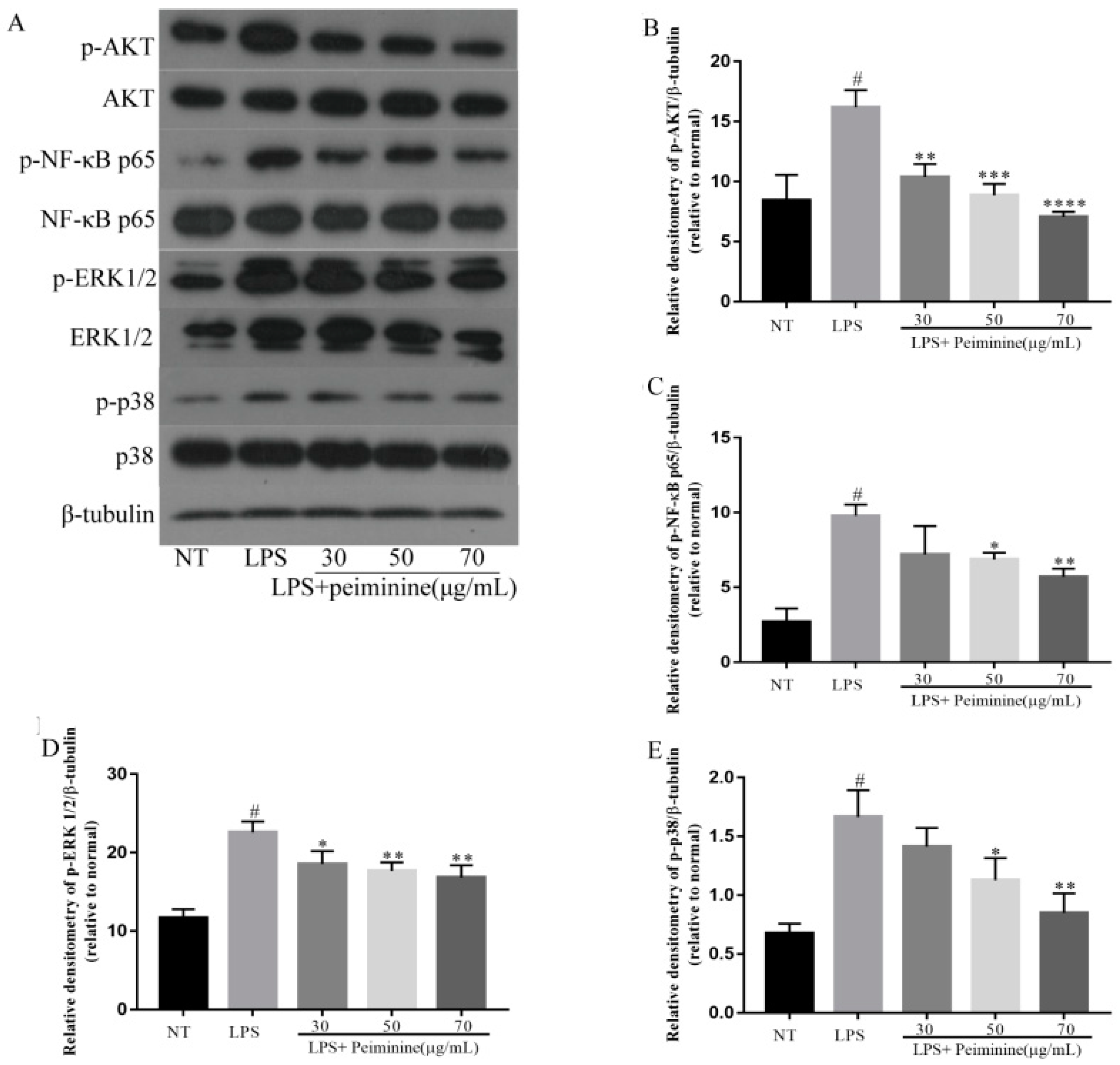
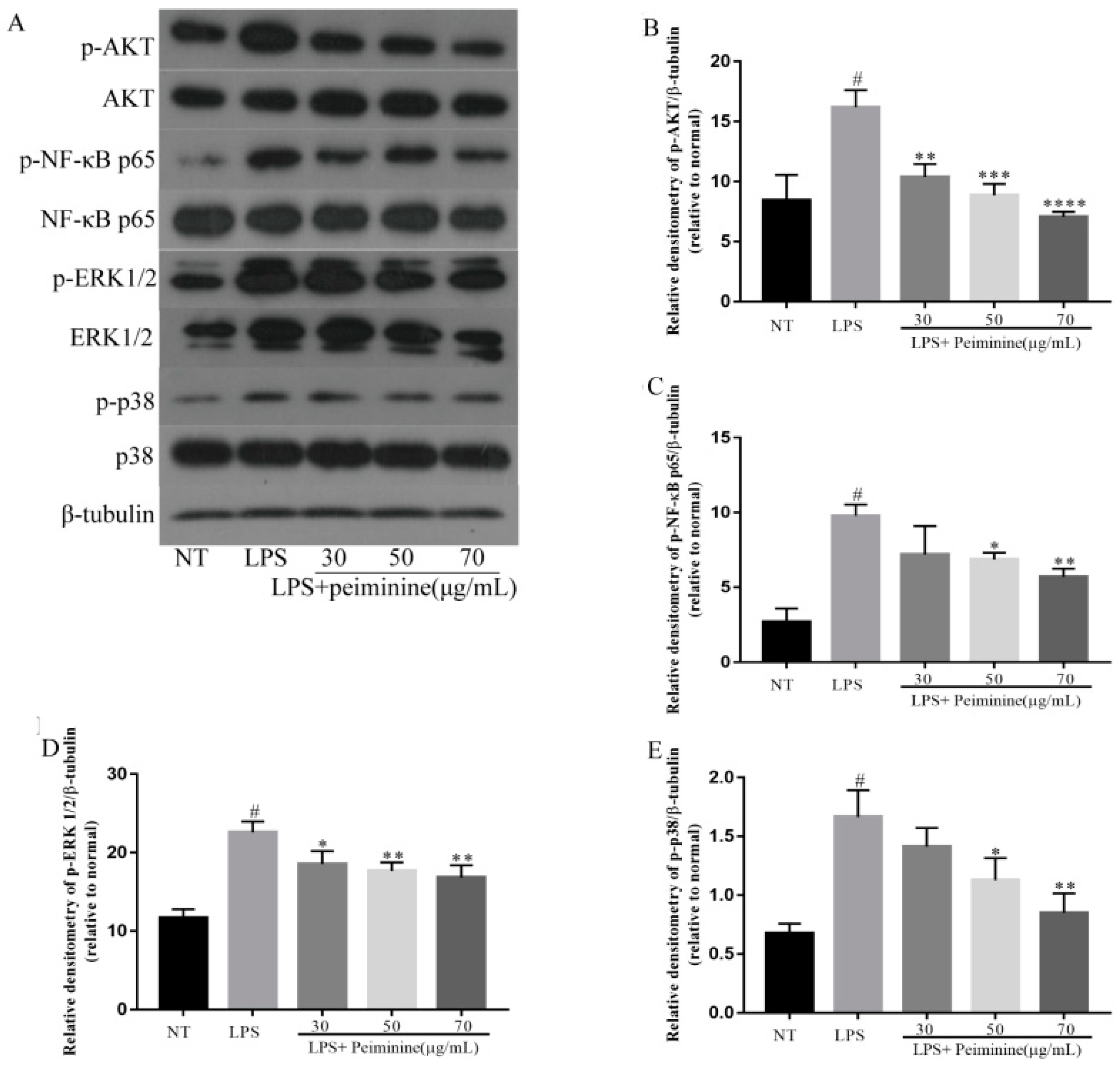
2.5. Effect of Peiminine on the LPS-Induced Activation of AKT, NF-κB p65, ERK1/2, and p38 in mMECs
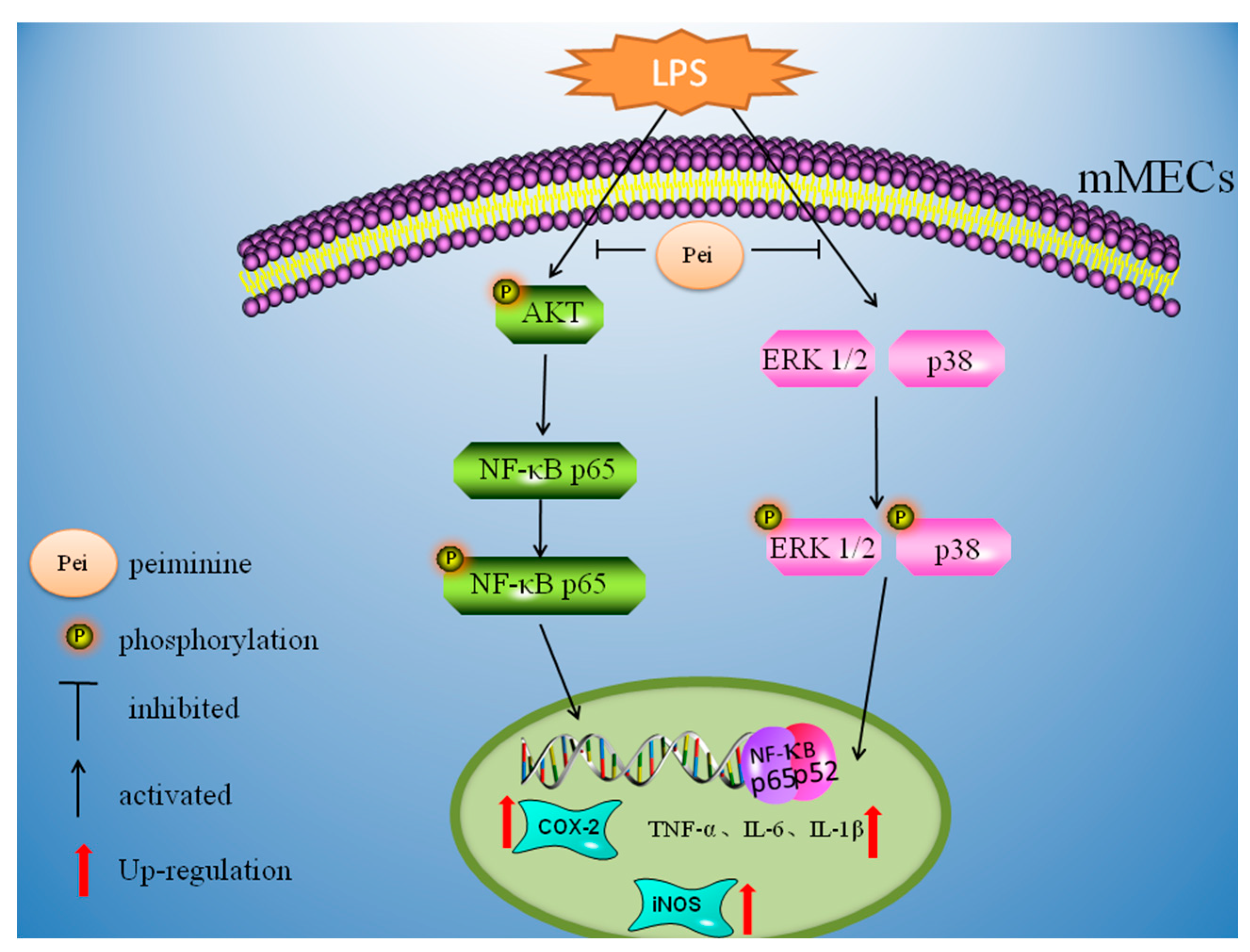
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Mastitis Model
4.3. Cell Culture
4.4. Histopathologic Evaluation of Mammary Tissues
4.5. Tissue Homogenates and MPO Assay
4.6. Enzyme-Linked Immunosorbent Assay
4.7. CCK8 Assay
4.8. Cell Culture Experimental Design
4.9. Western Blot Analysis
4.10. Data and Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zadoks, R.N.; Middleton, J.R.; McDougall, S.; Katholm, J.; Schukken, Y.H. Molecular epidemiology of mastitis pathogens of dairy cattle and comparative relevance to humans. J. Mammary Gland Biol. Neoplasia 2011, 16, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Kauf, A.C.; Vinyard, B.T.; Bannerman, D.D. Effect of intramammary infusion of bacterial lipopolysaccharide on experimentally induced Staphylococcus aureus intramammary infection. Res. Vet. Sci. 2007, 82, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hartlage-Rubsamen, M.; Lemke, R.; Schliebs, R. Interleukin-1β, inducible nitric oxide synthase, and nuclear factor-κB are induced in morphologically distinct microglia after rat hippocampal lipopolysaccharide/interferon-γ injection. J. Neurosci. Res. 1999, 57, 388–398. [Google Scholar] [CrossRef]
- Takeuchi, H.; Jin, S.J.; Wang, J.Y.; Zhang, G.Q.; Kawanokuchi, J.; Kuno, R.; Sonobe, Y.; Mizuno, T.; Suzumura, A. Tumor necrosis factor-α induces neurotoxicity via glutamate release from hemichannels of activated microglia in an autocrine manner. J. Biol. Chem. 2006, 281, 21362–21368. [Google Scholar] [CrossRef] [PubMed]
- Toubiana, M.; Gerdol, M.; Rosani, U.; Pallavicini, A.; Venier, P.; Roch, P. Toll-like receptors and MyD88 adaptors in Mytilus: Complete CDS and gene expression levels. Dev. Comp. Immunol. 2013, 40, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Wang, W.; Li, D.; Li, S.; Zhang, L.; Fu, Y.; Zhang, N. Mangiferin inhibits mastitis induced by LPS via suppressing NF-κB and NLRP3 signaling pathways. Int. Immunopharmacol. 2017, 43, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Xiao, C.; Wei, Z.; Wang, J.; Yang, Z.; Fu, Y. Resveratrol inhibits LPS-induced mice mastitis through attenuating the MAPK and NF-κB signaling pathway. Microb. Pathog. 2017, 107, 462–467. [Google Scholar] [CrossRef] [PubMed]
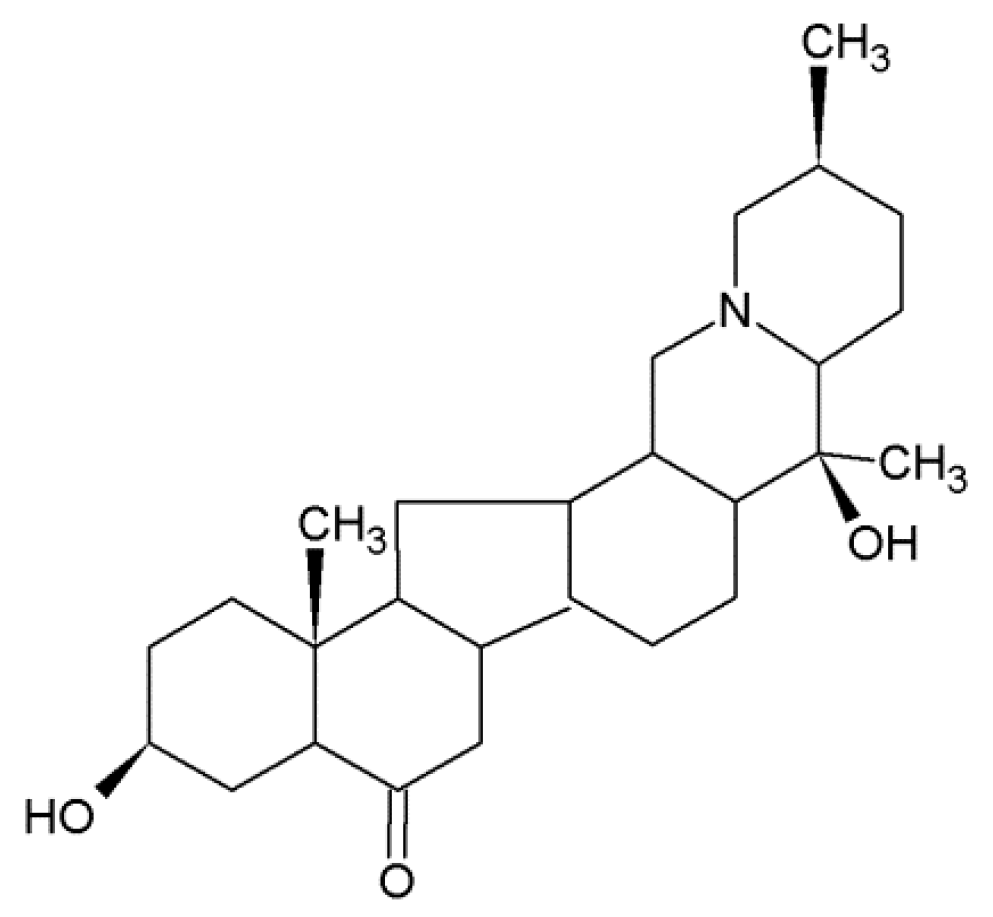
- Ruan, X.; Yang, L.; Cui, W.X.; Zhang, M.X.; Li, Z.H.; Liu, B.; Wang, Q. Optimization of supercritical fluid extraction of total alkaloids, peimisine, peimine and peiminine from the bulb of Fritillaria thunbergii Miq, and evaluation of antioxidant activities of the extracts. Materials 2016, 9, 524. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Tou, F.; Su, H.; Wu, X.; Chen, X.; Zheng, Z. The natural product peiminine represses colorectal carcinoma tumor growth by inducing autophagic cell death. Biochem. Biophys. Res. Commun. 2015, 462, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ji, F.; Liu, B.; Chen, X.; He, J.; Gong, J. Peiminine ameliorates bleomycin-induced acute lung injury in rats. Mol. Med. Rep. 2013, 7, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhu, J.; Wang, S.; Wang, X.; Ou, Y.; Wei, D.; Li, X. Antitussive, expectorant and anti-inflammatory alkaloids from Bulbus Fritillariae Cirrhosae. Fitoterapia 2011, 82, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.F.; Huri, D.A.; Snyder, S.H. Inducible nitric oxide synthase binds, S-nitrosylates, and activates cyclooxygenase-2. Science 2005, 310, 1966–1970. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Li, P.; Li, S.L.; Chan, S.W. Chromatographic analysis of Fritillaria isosteroidal alkaloids, the active ingredients of Beimu, the antitussive traditional Chinese medicinal herb. J. Chromatogr. A 2001, 935, 321–338. [Google Scholar] [CrossRef]
- Li, Y.F.; Li, Y.X.; Lin, J.; Xu, Y.; Yan, F.; Tang, L.; Chen, F. Identification of bulb from Fritillaria cirrhosa by PCR with specific primers. Planta Med. 2003, 69, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Gao, L.M.; Yang, Y.P. Genetic diversity and structure of a traditional Chinese medicinal plant species, Fritillaria cirrhosa (Liliaceae) in southwest China and implications for its conservation. Biochem. Syst. Ecol. 2010, 38, 236–242. [Google Scholar] [CrossRef]
- Gu, B.B.; Miao, J.F.; Zhu, Y.M.; Deng, Y.E.; Zou, S.X. Protective effect of retinoid against endotoxin-induced mastitis in rats. Inflamm. Res. 2009, 58, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.P.; Christensen, R.D.; Rothstein, G. Cellular and extracellular myeloperoxidase in pyogenic inflammation. Blood 1982, 60, 618–622. [Google Scholar] [PubMed]
- Winterbourn, C.C.; Vissers, M.C.; Kettle, A.J. Myeloperoxidase. Curr. Opin. Hematol. 2000, 7, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Haegens, A.; Heeringa, P.; van Suylen, R.J.; Steele, C.; Aratani, Y.; O’Donoghue, R.J.; Mutsaers, S.E.; Mossman, B.T.; Wouters, E.F.; Vernooy, J.H. Myeloperoxidase deficiency attenuates lipopolysaccharide-induced acute lung inflammation and subsequent cytokine and chemokine production. J. Immunol. 2009, 182, 7990–7996. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.; Baldus, S. Myeloperoxidase and its contributory role in inflammatory vascular disease. Pharmacol. Ther. 2006, 111, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [PubMed]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Xiong, H.Z.; Liu, L.B. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2012, 141, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.; Pfaffl, M.W.; Meyer, H.H.D.; Bruckmaier, R.M. Short-term changes of mRNA expression of various inflammatory factors and milk proteins in mammary tissue during LPS-induced mastitis. Domest. Anim. Endocrinol. 2004, 26, 111–126. [Google Scholar] [CrossRef] [PubMed]
- West, M.A.; Seatter, S.C.; Bellingham, J.; Clair, L. Mechanisms of reprogrammed macrophage endotoxin signal transduction after lipopolysaccharide pretreatment. Surgery 1995, 118, 220–228. [Google Scholar] [CrossRef]
- Nilsson, K.; Jernberg, H.; Pettersson, M. IL-6 as a growth factor for human multiple myeloma cells—A short overview. Curr. Top. Microbiol. Immunol. 1990, 166, 3–12. [Google Scholar] [PubMed]
- Wang, D.D.; Du, Q.D.; Li, H.C.; Wang, S. The isosteroid alkaloid imperialine from bulbs of Fritillaria cirrhosa mitigates pulmonary functional and structural impairment and suppresses inflammatory response in a COPD-like rat model. Mediat. Inflamm. 2016, 2016, 4192483. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.M.; Lee, B.; Min, J.H.; Kim, E.Y.; Kim, J.H.; Hong, S.; Kim, J.J.; Sohn, Y.; Jung, H.S. Effect of peiminine on DNCB-induced atopic dermatitis by inhibiting inflammatory cytokine expression in vivo and in vitro. Int. Immunopharmacol. 2018, 56, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-κB pathway in the treatment of inflammation and cancer. J. Clin. Investig. 2001, 107, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Huang, W.; Gu, J.; Du, X.; Lei, L.; Yuan, X.; Sun, G.; Wang, Z.; Li, X.; Liu, G. SREBP-1c overactivates ROS-mediated hepatic NF-κB inflammatory pathway in dairy cows with fatty liver. Cell Signal. 2015, 27, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Seo, H.S.; Ku, J.M.; Kim, G.J.; Jeon, C.Y.; Park, J.H.; Jang, B.H.; Park, S.J.; Shin, Y.C.; Ko, S.G. Silibinin inhibits the production of pro-inflammatory cytokines through inhibition of NF-κB signaling pathway in HMC-1 human mast cells. Inflamm. Res. 2013, 62, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Mo, C.F.; Xiao, H.Y.; Jiang, Y.; Ye, B.G.; Wang, S. Imperialine and verticinone from bulbs of Fritillaria wabuensis inhibit pro-inflammatory mediators in LPS-stimulated RAW264.7 macrophages. Planta Med. 2015, 81, 821–829. [Google Scholar] [PubMed]
- Lee, B.; Kim, E.Y.; Kim, J.H.; Min, J.H.; Jeong, D.W.; Jun, J.Y.; Cho, C.Y.; Sohn, Y.; Jung, H.S. Antiallergic effects of peiminine through the regulation of inflammatory mediators in HMC-1 cells. Immunopharm. Immunol. 2015, 37, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, N.; Gu, J.; Fu, S.; Peng, Z.; Zhao, C.; Zhang, Y.; Li, X.; Wang, Z.; Li, X.; et al. β-Hydroxybutyrate induces bovine hepatocyte apoptosis via an ROS-p38 signaling pathway. J. Dairy Sci. 2016, 99, 9184–9198. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, J.; Jiang, L.; Ran, X.; He, D.; Li, Y.; Huang, B.; Wang, W.; Liu, D.; Fu, S. Peiminine protects dopaminergic neurons from inflammation-induced cell death by inhibiting the ERK1/2 and NF-κB signalling pathways. Int. J. Mol. Sci. 2018, 19, 821. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Lian, S.; Zhen, L.; Zang, S.; Chen, Y.; Lang, L.; Xu, B.; Guo, J.; Ji, H.; Wang, J.; et al. The favored mechanism for coping with acute cold stress: Upregulation of miR-210 in rats. Cell. Physiol. Biochem. 2018, 46, 2090–2102. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, Q.; Li, Y.; Ma, H.; Guo, W.; Kan, X.; Xu, D.; Liu, J.; Fu, S. Peiminine Protects against Lipopolysaccharide-Induced Mastitis by Inhibiting the AKT/NF-κB, ERK1/2 and p38 Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 2637. https://doi.org/10.3390/ijms19092637
Gong Q, Li Y, Ma H, Guo W, Kan X, Xu D, Liu J, Fu S. Peiminine Protects against Lipopolysaccharide-Induced Mastitis by Inhibiting the AKT/NF-κB, ERK1/2 and p38 Signaling Pathways. International Journal of Molecular Sciences. 2018; 19(9):2637. https://doi.org/10.3390/ijms19092637
Chicago/Turabian StyleGong, Qian, Yanwei Li, He Ma, Wenjin Guo, Xingchi Kan, Dianwen Xu, Juxiong Liu, and Shoupeng Fu. 2018. "Peiminine Protects against Lipopolysaccharide-Induced Mastitis by Inhibiting the AKT/NF-κB, ERK1/2 and p38 Signaling Pathways" International Journal of Molecular Sciences 19, no. 9: 2637. https://doi.org/10.3390/ijms19092637