Evaluation of BMP-2 Enhances the Osteoblast Differentiation of Human Amnion Mesenchymal Stem Cells Seeded on Nano-Hydroxyapatite/Collagen/Poly(l-Lactide)

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
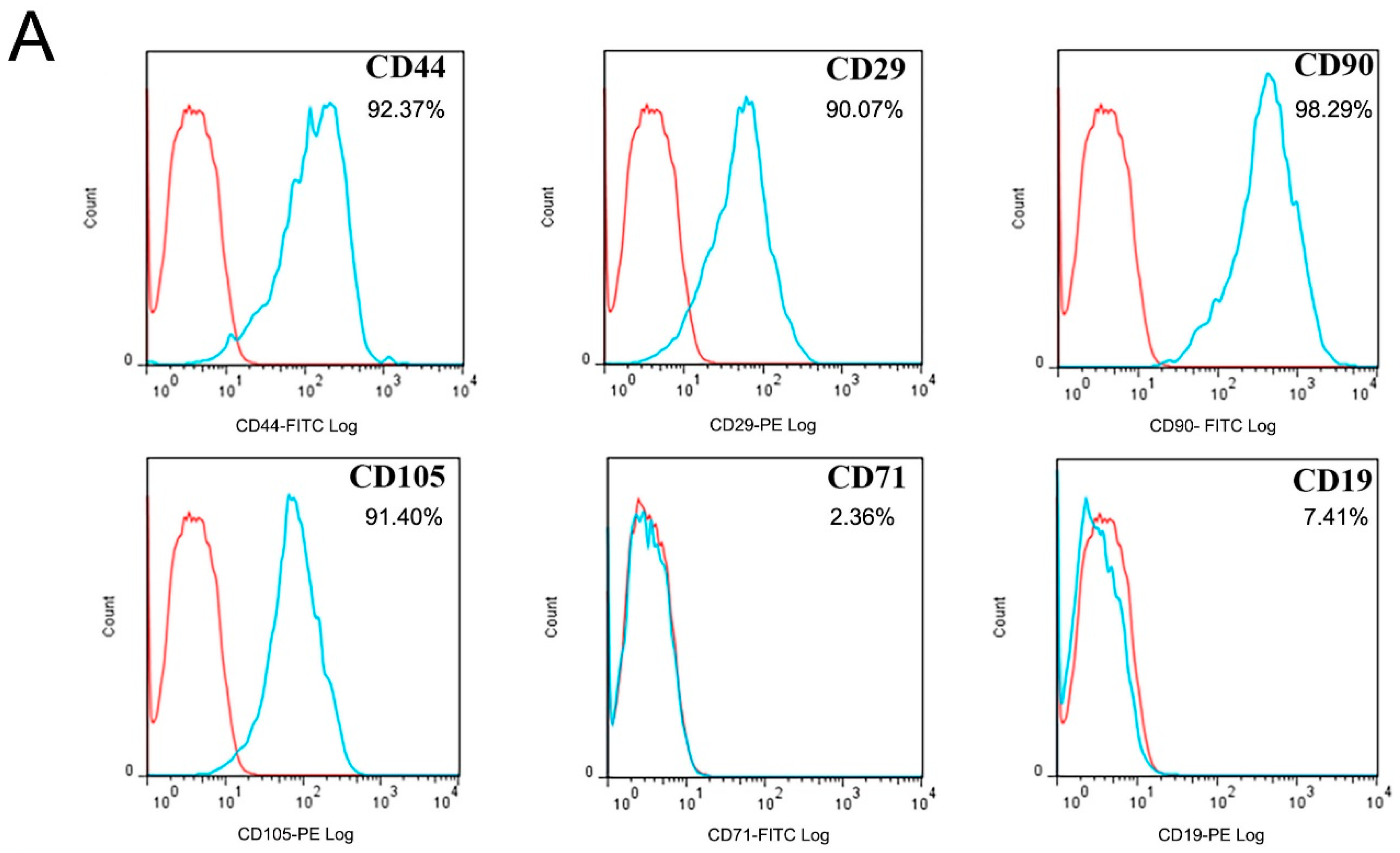
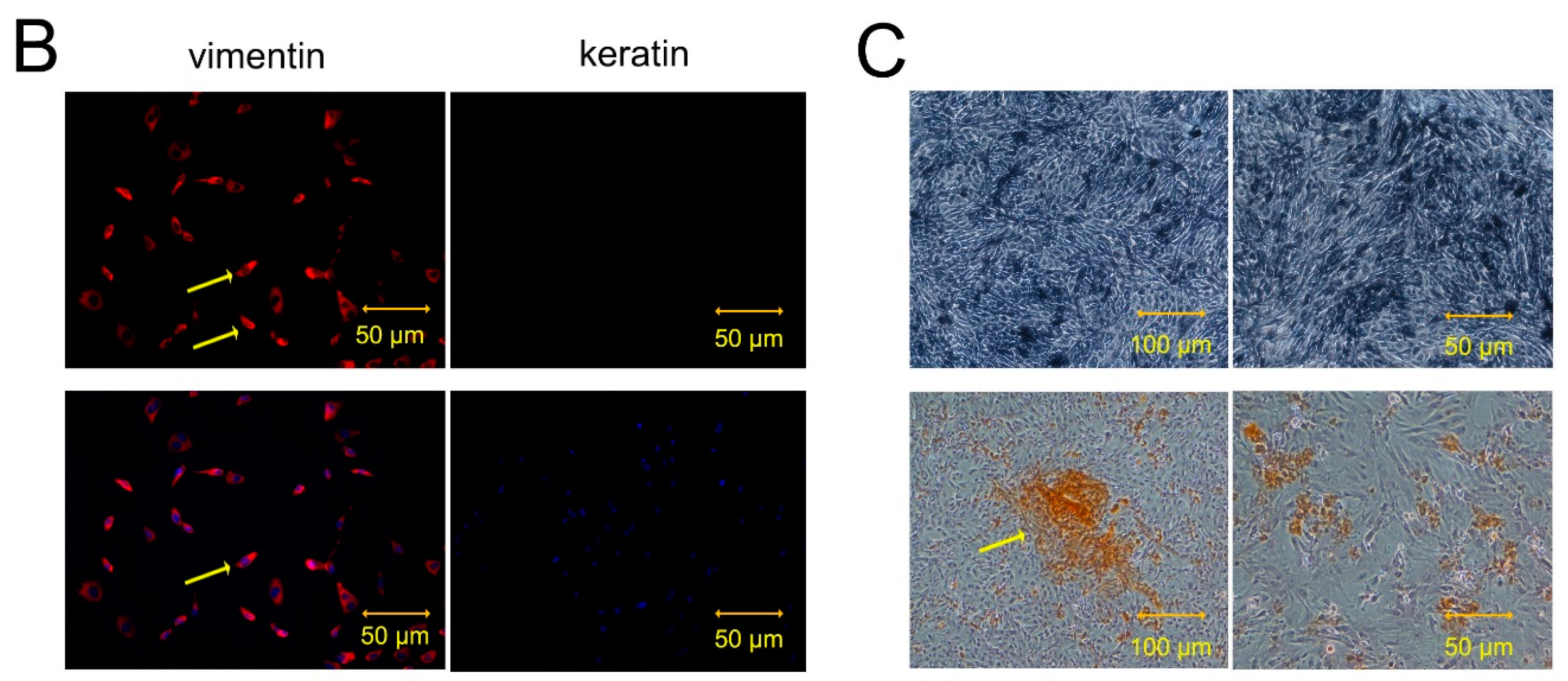
2.1. Cultural and OB Differentiation Identification of hAMSCs
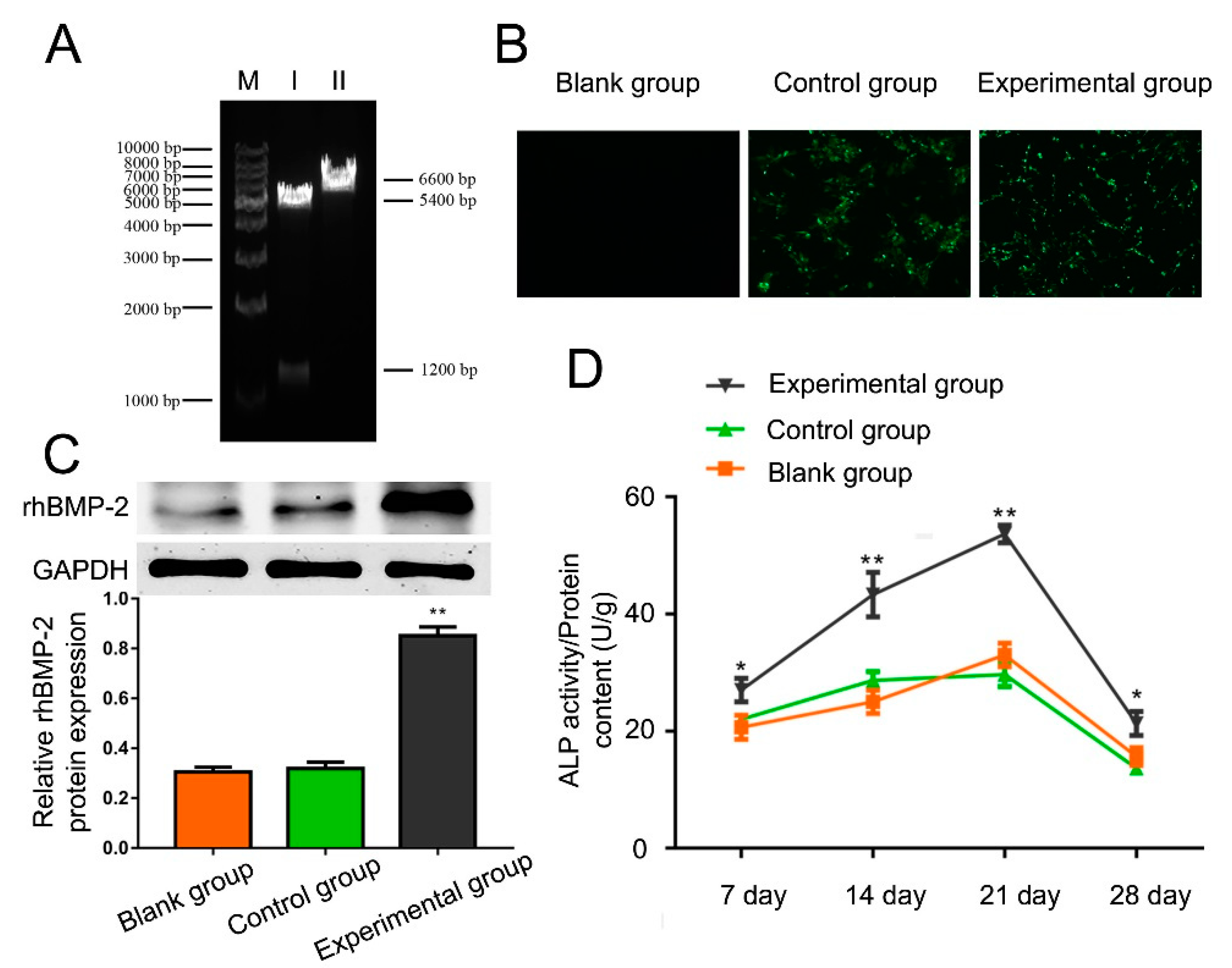
2.2. Identification of the Recombinant Plasmid
2.3. Cell Transfection Efficiency
2.4. Effect of rhBMP-2 on ALP Activity of hAMSCs
2.5. Scanning Electron Microscope (SEM) Analysis
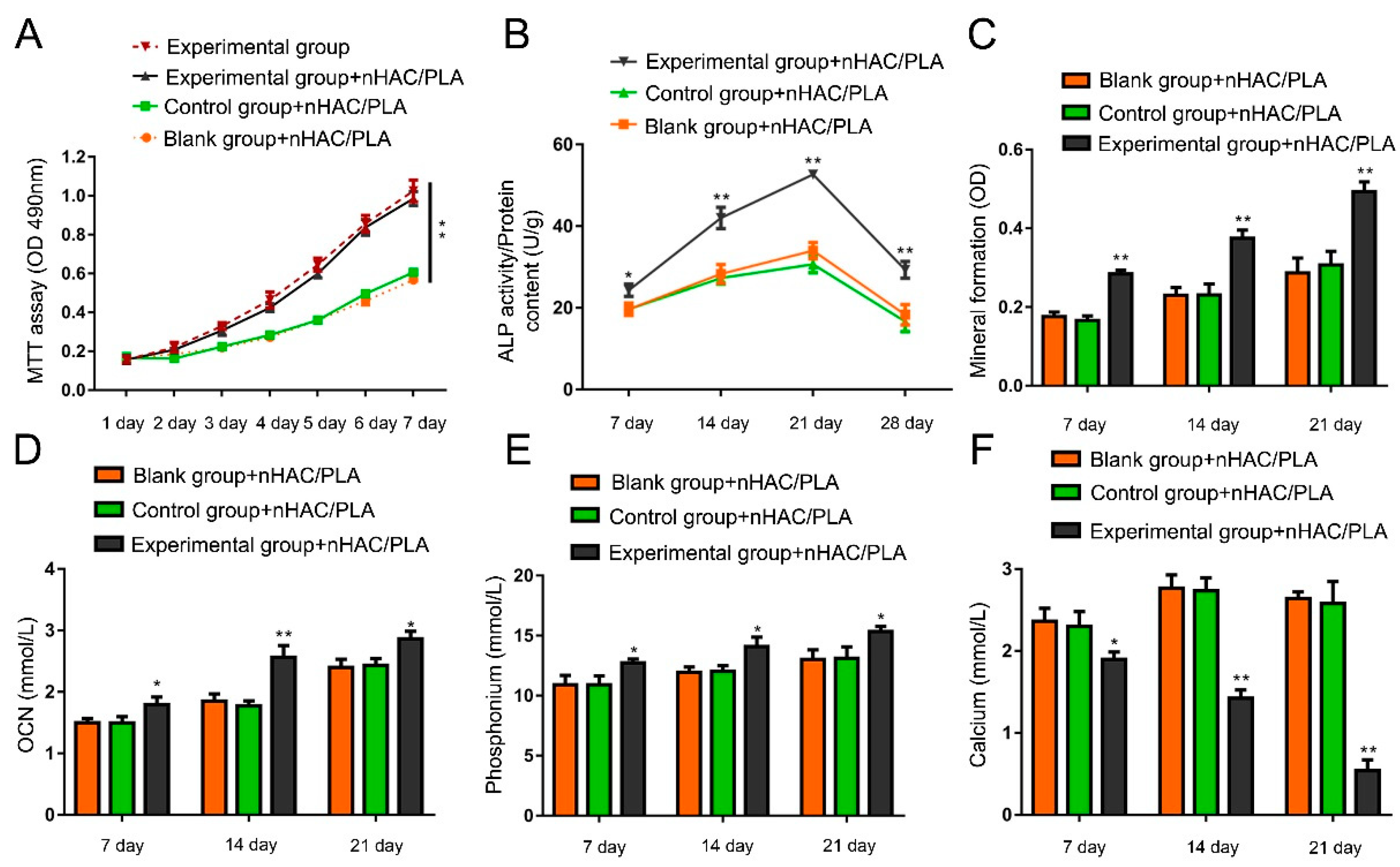
2.6. Effect of rhBMP-2 on Proliferation in the hAMSCs Seeded on the nHAC/PLA
2.7. Effect of rhBMP-2 on ALP Activity of hAMSCs Seeded on nHAC/PLA
2.8. Effect of rhBMP-2 on Mineral Formation in the hAMSCs Seeded on the nHAC/PLA
2.9. Effect of rhBMP-2 on Calcium, Phosphate, and OCN Content of hAMSCs Seeded on nHAC/PLA
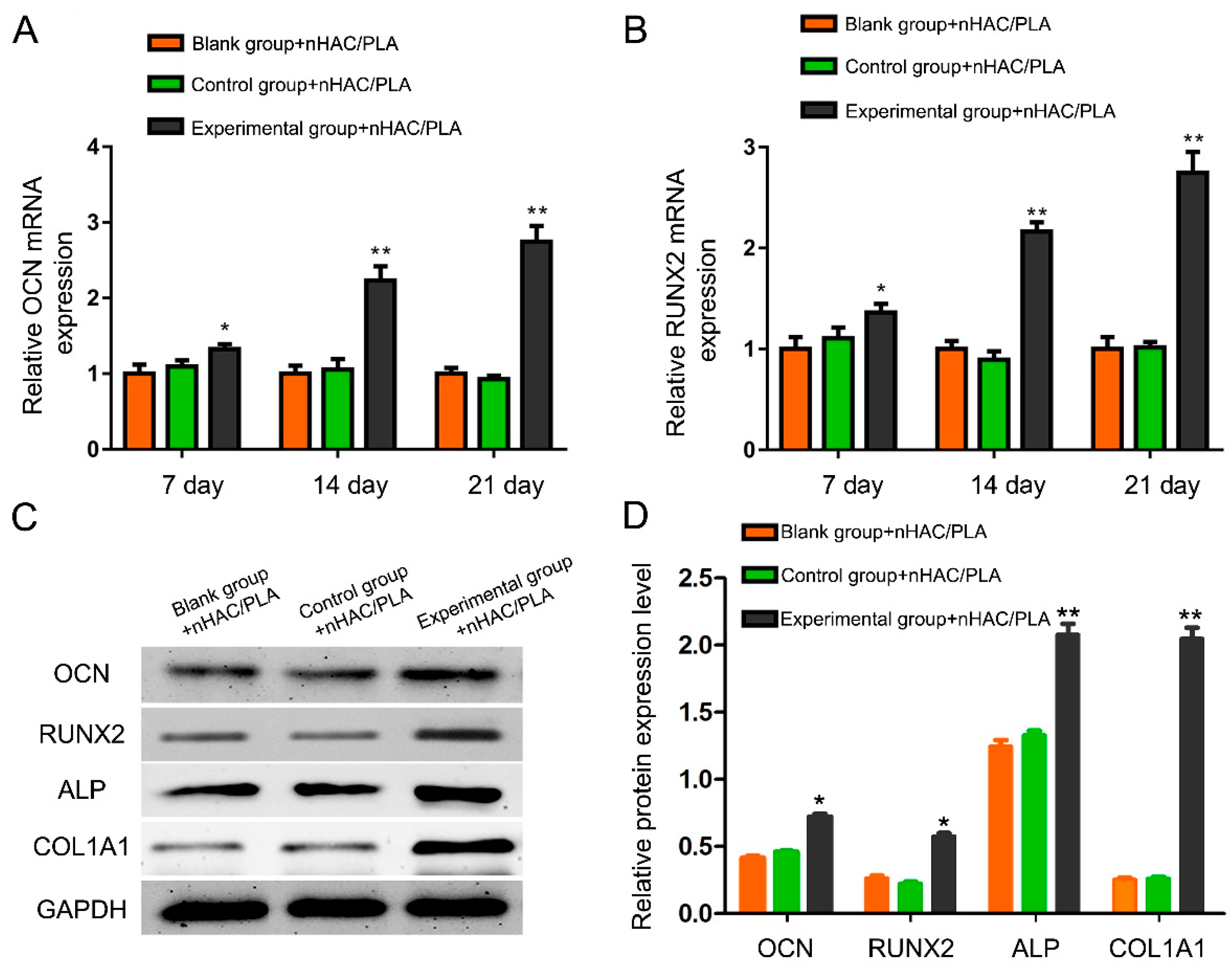
2.10. Effect of rhBMP-2 on OCN, RUNX2, ALP, and COL1A1 Expression of hAMSCs Seeded on nHAC/PLA
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Cultures
4.2. Flow Cytometry and Immunofluorescence
4.3. Construction of pcDNA3.1-rhBMP-2
4.4. Transfection Efficiency Detection
4.5. Biometrics Preparation and Seeding of nHAC/PLA Scaffolds
4.6. Scanning Electron Microscopy (SEM)
4.7. Alkaline Phosphatase Staining and Activity Assay
4.8. Alizarin Red Staining and Quantification
4.9. MTT Assay
4.10. Quantitative Real-Time Polymerase Chain Reaction
4.11. Western Blot Analysis
4.12. Calcium, Phosphate, and OCN Content Assays
4.13. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | alkaline phosphatase |
| ANOVA | analysis of variance |
| BMP | bone morphogenetic protein |
| BMP-2 | bone morphogenetic protein 2 |
| Ct | cycle threshold |
| GAPDH | glyceraldehyde-3-phosphate-dehydrogenase |
| GFP | green fluorescent protein |
| hAMSCs | human amnion mesenchymal stem cells |
| MSCs | mesenchymal stem cells |
| MTS | 3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium |
| nHAC/PLA | nano-hydroxyapatite/collagen/poly(l-lactide) |
| OB | osteoblast |
| OC | osteoclast |
| OCN | osteocalcin |
| OD | optical density |
| PBS | phosphate buffered saline |
| rhBMP-2 | recombinant human BMP-2 |
| RT-qPCR | Reverse transcription quantitative real-time polymerase chain reaction |
| SD | standard deviation |
| SDS/PAGE | sodium dodecyl sulphate polyacrylamide gel electrophoresis |
| TGF-β | transforming growth factor-β |
References
- Manolagas, S.C. Birth and death of bone cells: Basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr. Rev. 2000, 21, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Christenson, R.H. Biochemical markers of bone metabolism: An overview. Clin. Biochem. 1997, 30, 573–593. [Google Scholar] [CrossRef]
- August, M.; Tompach, P.; Chang, Y.; Kaban, L. Factors influencing the long-term outcome of mandibular reconstruction. J. Oral Maxillofac. Surg. 2000, 58, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Hartman, E.H.; Spauwen, P.H.; Jansen, J.A. Donor-site complications in vascularized bone flap surgery. J. Investig. Surg. 2002, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Ahlmann, E.; Patzakis, M.; Roidis, N.; Shepherd, L.; Holtom, P. Comparison of anterior and posterior iliac crest bone grafts in terms of harvest-site morbidity and functional outcomes. J. Bone Jt. Surg. Am. 2002, 84-A, 716–720. [Google Scholar] [CrossRef]
- Boone, D.W. Complications of iliac crest graft and bone grafting alternatives in foot and ankle surgery. Foot Ankle Clin. 2003, 8, 1–14. [Google Scholar] [CrossRef]
- Velchuru, V.R.; Satish, S.G.; Petri, G.J.; Sturzaker, H.G. Hernia through an iliac crest bone graft site: Report of a case and review of the literature. Bull. Hosp. Jt. Dis. 2006, 63, 166–168. [Google Scholar] [PubMed]
- Zijderveld, S.A.; ten Bruggenkate, C.M.; van Den Bergh, J.P.; Schulten, E.A. Fractures of the iliac crest after splitthickness bone grafting for preprosthetic surgery: Report of 3 cases and review of the literature. J. Oral Maxillofac. Surg. 2004, 62, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Hölzle, F.; Kesting, M.R.; Hölzle, G.; Watola, A.; Loeffelbein, D.J.; Ervens, J.; Wolff, K.D. Clinical outcome and patient satisfaction after mandibular reconstruction with free fibula flaps. Int. J. Oral Maxillofac. Surg. 2007, 36, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R.; Silverman, B.F.; Büring, K.; Dubuc, F.L.; Rosenberg, J.M. The bone induction principle. Clin. Orthop. Relat. Res. 1967, 53, 243–283. [Google Scholar] [CrossRef] [PubMed]
- Brantigan, J.W.; Cunningham, B.W.; Warden, K.; McAfee, P.C.; Steffee, A.D. Compression strength of donor bone for posterior lumbar interbody fusion. Spine 1993, 18, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Marks, S.C., Jr.; Odgren, P.R. Structure and development of the skeleton. In Principles of Bone Biology, 2nd ed.; Bilezikian, J.P., Raisz, L.G., Rodan, G.A., Eds.; Academic Press: San Diego, CA, USA, 2002; p. 3. [Google Scholar]
- Cui, F.Z.; Zhang, W.; Liao, S.S. Hierarchical self-assembly of nano-fibrils in mineralized collagen. Chem. Mater. 2003, 15, 3221–3226. [Google Scholar]
- Liao, S.; Wang, W.; Uo, M.; Ohkawa, S.; Akasaka, T.; Tamura, K.; Cui, F.; Watari, F. A three-layered nano-carbonated hydroxyapatite/collagen/PLGA composite membrane for guided tissue regeneration. Biomaterials 2005, 26, 7564–7571. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.C.; E, L.L.; Wang, D.S.; Su, F.; Wu, X.; Shi, Z.P.; Lv, Y.; Wang, J.Z. Reconstruction of alveolar bone defects using bone morphogenetic protein 2 mediated rabbit dental pulp stem cells seeded on nano-hydroxyapatite/collagen/poly(l-lactide). Tissue Eng. Part A 2011, 17, 2417–2433. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, C.; Brandi, M.L. Adipose mesenchymal stem cells in the field of bone tissue engineering. World J. Stem Cells 2014, 6, 144. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- De Ugarte, D.A.; Morizono, K.; Elbarbary, A.; Alfonso, Z.; Zuk, P.A.; Zhu, M.; Dragoo, J.L.; Ashjian, P.; Thomas, B.; Benhaim, P.; et al. Comparison of multi-lineage cells from human adipose tissue and bone marrow. Cells Tissues Organs 2003, 174, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Vilalta, M.; Dégano, I.R.; Bagó, J.; Gould, D.; Santos, M.; García-Arranz, M.; Ayats, R.; Fuster, C.; Chernajovsky, Y.; García-Olmo, D.; et al. Biodistribution, long-term survival, and safety of human adipose tissue-derived mesenchymal stem cells transplanted in nude mice by high sensitivity non-invasive bioluminescence imaging. Stem Cells Dev. 2008, 17, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.M.; Rosen, V.; Celeste, A.J.; Mitsock, L.M.; Whitters, M.J.; Kriz, R.W.; Hewick, R.M.; Wang, E.A. Novel regulators of bone formation: Molecular clones and activities. Science 1988, 242, 1528. [Google Scholar] [CrossRef] [PubMed]
- Govender, S.; Csimma, C.; Genant, H.K.; Valentin-Opran, A.; Amit, Y.; Arbel, R.; Aro, H.; Atar, D.; Bishay, M.; Börner, M.G.; et al. Recombinant human bone morphogenetic protein-2 for treatment of open tibial fractures: A prospective, controlled, randomized study of four hundred and fifty patients. J. Bone Jt. Surg. Am. 2002, 84-A, 2123–2134. [Google Scholar] [CrossRef]
- Grgurevic, L.; Macek, B.; Mercep, M.; Jelic, M.; Smoljanovic, T.; Erjavec, I.; Dumic-Cule, I.; Prgomet, S.; Durdevic, D.; Vnuk, D.; et al. Cytokines and BMP-4 promote hematopoietic differentiation of human embryonic stem cells. Blood 2003, 102, 906–915. [Google Scholar] [Green Version]
- Lind, M.; Eriksen, E.F.; Bünger, C. Bone morphogenetic protein-2 but not bone morphogenetic protein-4 and -6 stimulates chemotactic migration of human osteoblasts, human marrow osteoblasts, and U2-OS cells. Bone 1996, 18, 53–57. [Google Scholar] [CrossRef]
- Reddi, A.H.; Cunningham, N.S. Initiation and promotion of bone differentiation by bone morphogenetic proteins. J. Bone Miner. Res. 1993, 8 (Suppl. 2), S499–S502. [Google Scholar] [CrossRef] [PubMed]
- Grgurevic, L.; Macek, B.; Mercep, M.; Jelic, M.; Smoljanovic, T.; Erjavec, I.; Dumic-Cule, I.; Prgomet, S.; Durdevic, D.; Vnuk, D.; et al. Bone morphogenetic protein (BMP)1-3 enhances bone repair. Biochem. Biophys. Res. Commun. 2011, 408, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musial, K.; Fornalczyk, K.; Zwolinska, D. Osteopontin (OPN), PDGF-BB (platelet-derived growth factor) and BMP-7 (bone morphogenetic protein) as markers of atherogenesis in children with chronic kidney disease (CKD) treated conservatively preliminary results. Polski Merkuriusz Lekarski 2008, 24, 25–27. [Google Scholar] [PubMed]
- Davis, H.; Raja, E.; Miyazono, K.; Tsubakihara, Y.; Moustakas, A. Mechanisms of action of bone morphogenetic proteins in cancer. Cytokine Growth Factor Rev. 2016, 27, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Song, S.H.; Yun, Y.P.; Choi, B.J.; Kwon, I.K.; Bae, M.S.; Moon, H.J.; Kwon, Y.D. The effect of immobilization of heparin and bone morphogenic protein-2 (BMP-2) to titanium surfaces on inflammation and osteoblast function. Biomaterials 2011, 32, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.E.; Choi, J.; Joung, Y.K.; Park, K.; Han, D.K. Controlled release of bone morphogenetic protein (BMP)-2 from nanocomplex incorporated on hydroxyapatite-formed titanium surface. J. Control. Release 2012, 160, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Pyo, S.J.; Song, W.W.; Kim, I.R.; Park, B.S.; Kim, C.H.; Shin, S.H.; Chung, I.K.; Kim, Y.D. Low-level laser therapy induces the expressions of BMP-2, osteocalcin, and TGF-beta1 in hypoxic-cultured human osteoblasts. Lasers Med. Sci. 2013, 28, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Samara, S.; Dailiana, Z.; Varitimidis, S.; Chassanidis, C.; Koromila, T.; Malizos, K.N.; Kollia, P. Bone morphogenetic proteins (BMPs) expression in the femoral heads of patients with avascular necrosis. Mol. Biol. Rep. 2013, 40, 4465–4472. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.Y.; On, S.W.; Song, S.I. Bone regenerative effect of recombinant human bone morphogenetic protein-2 after cyst enucleation. Maxillofac. Plast. Reconstr. Surg. 2016, 38, 22. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.; Hu, J.; Song, D.; Jiang, X.; Zhu, S. Recombinant human bone morphogenetic protein-2 suspended in fibrin glue enhances bone formation during distraction osteogenesis in rabbits. Arch. Med. Sci. 2016, 12, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Chatzinikolaidou, M.; Pontikoglou, C.; Terzaki, K.; Kaliva, M.; Kalyva, A.; Papadaki, E.; Vamvakaki, M.; Farsari, M. Recombinant human bone morphogenetic protein 2 (rhBMP-2) immobilized on laser-fabricated 3D scaffolds enhance osteogenesis. Colloids Surf. B Biointerfaces 2016, 149, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Ries, J.; Gelse, K.; Kloss, F.; von der Mark, K.; Wiltfang, J.; Neukam, F.W.; Schneider, H. Bone regeneration in critical size defects by cell-mediated BMP-2 gene transfer: A comparison of adenoviral vectors and liposomes. Gene Ther. 2003, 10, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Tong, W.; Luria, J.S.; Wang, Z.; Nussenbaum, B.; Krebsbach, P.H. Effects of bone morphogenetic protein-2 on proliferation and angiogenesis in oral squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2010, 39, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Wada, T.; Yamashita, T.; Hamada, H. Enhanced osteoinduction by mesenchymal stem cells transfected with a fiber-mutant adenoviral BMP2 gene. J. Gene Med. 2005, 7, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.S.; Choi, C.H.; Cho, Y.B.; Kang, M.K.; Jang, C.H. Recombinant human BMP-2 enhances osteogenesis of demineralized bone matrix in experimental mastoid obliteration. Acta Otolaryngol. 2014, 134, 785–790. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yu, J.; Cao, J.; E, L.; Wang, D.; Zhang, H.; Liu, H. Biocompatibility and osteogenic capacity of periodontal ligament stem cells on nHAC/PLA and HA/TCP scaffolds. J. Biomater. Sci. Polym. Ed. 2011, 22, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Tamura, K.; Zhu, Y.; Wang, W.; Uo, M.; Akasaka, T.; Cui, F.; Watari, F. Human neutrophils reaction to the biodegraded nano-hydroxyapatite/collagen and nanohydroxyapatite/collagen/poly(l-lactic acid) composites. J. Biomed. Mater. Res. A 2006, 76, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Cheng, N.; Xu, C.; Sun, W.; Yu, C.; Shi, B. The effect of mesoporous bioglass on osteogenesis and adipogenesis of osteoporotic BMSCs. J. Biomed. Mater. Res. A 2016, 104, 3004–3014. [Google Scholar] [CrossRef] [PubMed]
- Guler, Z.; Silva, J.C.; Sarac, A.S. Enhanced osteogenesis on biofunctionalized poly(varepsilon-caprolactone)/poly(m-anthranilic acid) nanofibers. J. Biomater. Appl. 2016, 31, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Zhao, Y.; Sun, W.; Chen, B.; Zhang, J.; Zhao, W.; Xiao, Z.; Dai, J. The effect of crosslinking heparin to demineralized bone matrix on mechanical strength and specific binding to human bone morphogenetic protein-2. Biomaterials 2008, 29, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, R.T.; Ge, C.; Xiao, G.; Roca, H.; Jiang, D. Transcriptional regulation of osteoblasts. Cells Tissues Organs 2009, 189, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Phimphilai, M.; Zhao, Z.; Boules, H.; Roca, H.; Franceschi, R.T. BMP signaling is required for RUNX2-dependent induction of the osteoblast phenotype. J. Bone Miner. Res. 2006, 21, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Jiao, G.; Liu, H.; Wu, W.; Li, S.; Wang, Q.; Xu, D.; Li, X.; Liu, H.; Chen, Y. Biological silicon stimulates collagen type 1 and osteocalcin synthesis in human osteoblast-like cells through the BMP-2/Smad/RUNX2 signaling pathway. Biol. Trace Elem. Res. 2016, 173, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Jiang, M.; Miao, D. Transplanted human amniotic membrane-derived mesenchymal stem cells ameliorate carbon tetrachloride-induced liver cirrhosis in mouse. PLoS ONE 2011, 6, e16789. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, Y.; Jiang, F.; Chen, N. Human amnion mesenchymal stem cells promote proliferation and osteogenic differentiation in human bone marrow mesenchymal stem cells. J. Mol. Histol. 2015, 46, 13–20. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Xiao, Z.; Song, J.; Li, M.; Li, W. Evaluation of BMP-2 Enhances the Osteoblast Differentiation of Human Amnion Mesenchymal Stem Cells Seeded on Nano-Hydroxyapatite/Collagen/Poly(l-Lactide). Int. J. Mol. Sci. 2018, 19, 2171. https://doi.org/10.3390/ijms19082171
Wu S, Xiao Z, Song J, Li M, Li W. Evaluation of BMP-2 Enhances the Osteoblast Differentiation of Human Amnion Mesenchymal Stem Cells Seeded on Nano-Hydroxyapatite/Collagen/Poly(l-Lactide). International Journal of Molecular Sciences. 2018; 19(8):2171. https://doi.org/10.3390/ijms19082171
Chicago/Turabian StyleWu, Shuhong, Zhili Xiao, Jinlin Song, Min Li, and Wenhua Li. 2018. "Evaluation of BMP-2 Enhances the Osteoblast Differentiation of Human Amnion Mesenchymal Stem Cells Seeded on Nano-Hydroxyapatite/Collagen/Poly(l-Lactide)" International Journal of Molecular Sciences 19, no. 8: 2171. https://doi.org/10.3390/ijms19082171