Effect of Inhibition of DNA Methylation Combined with Task-Specific Training on Chronic Stroke Recovery


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
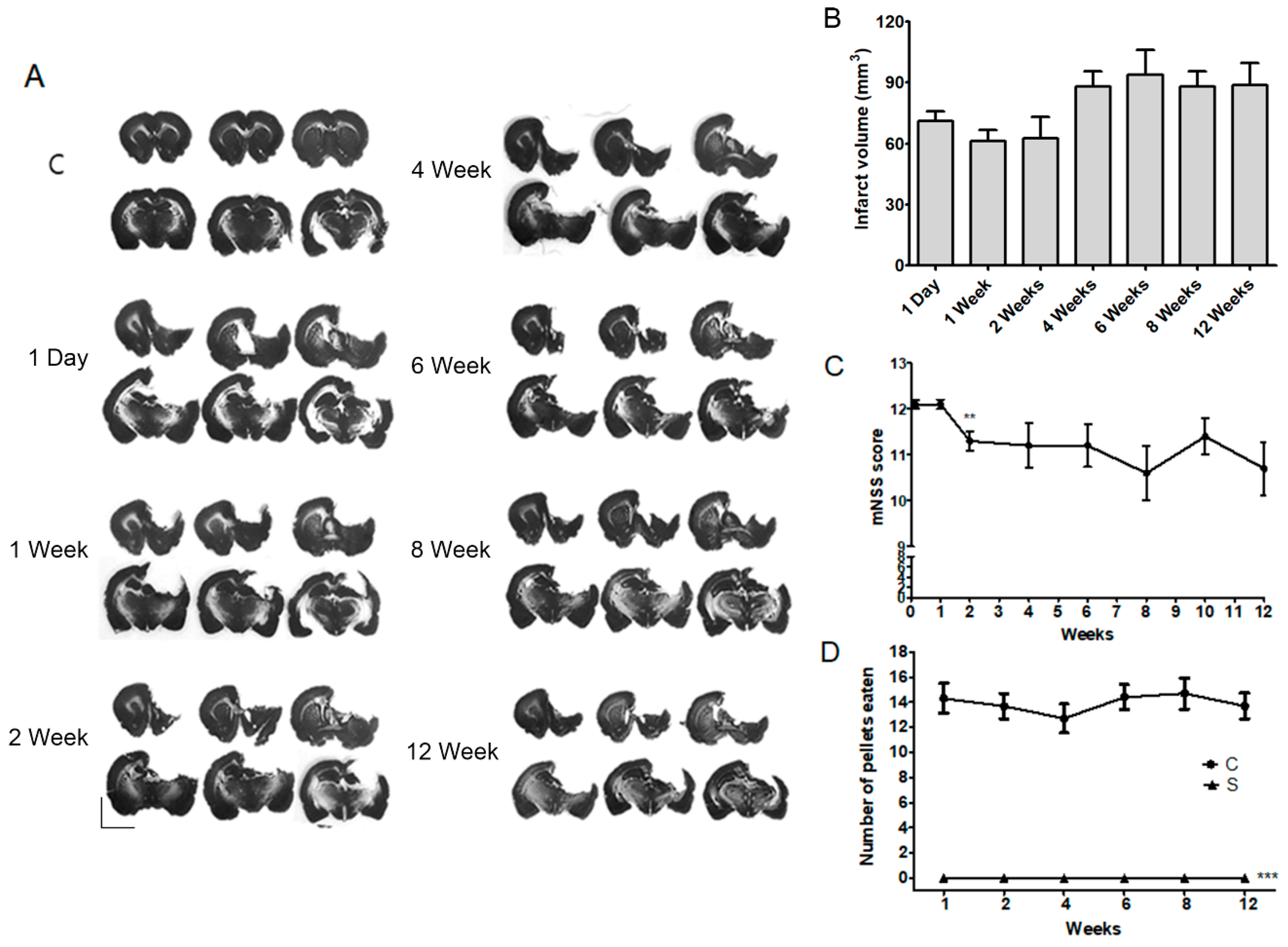
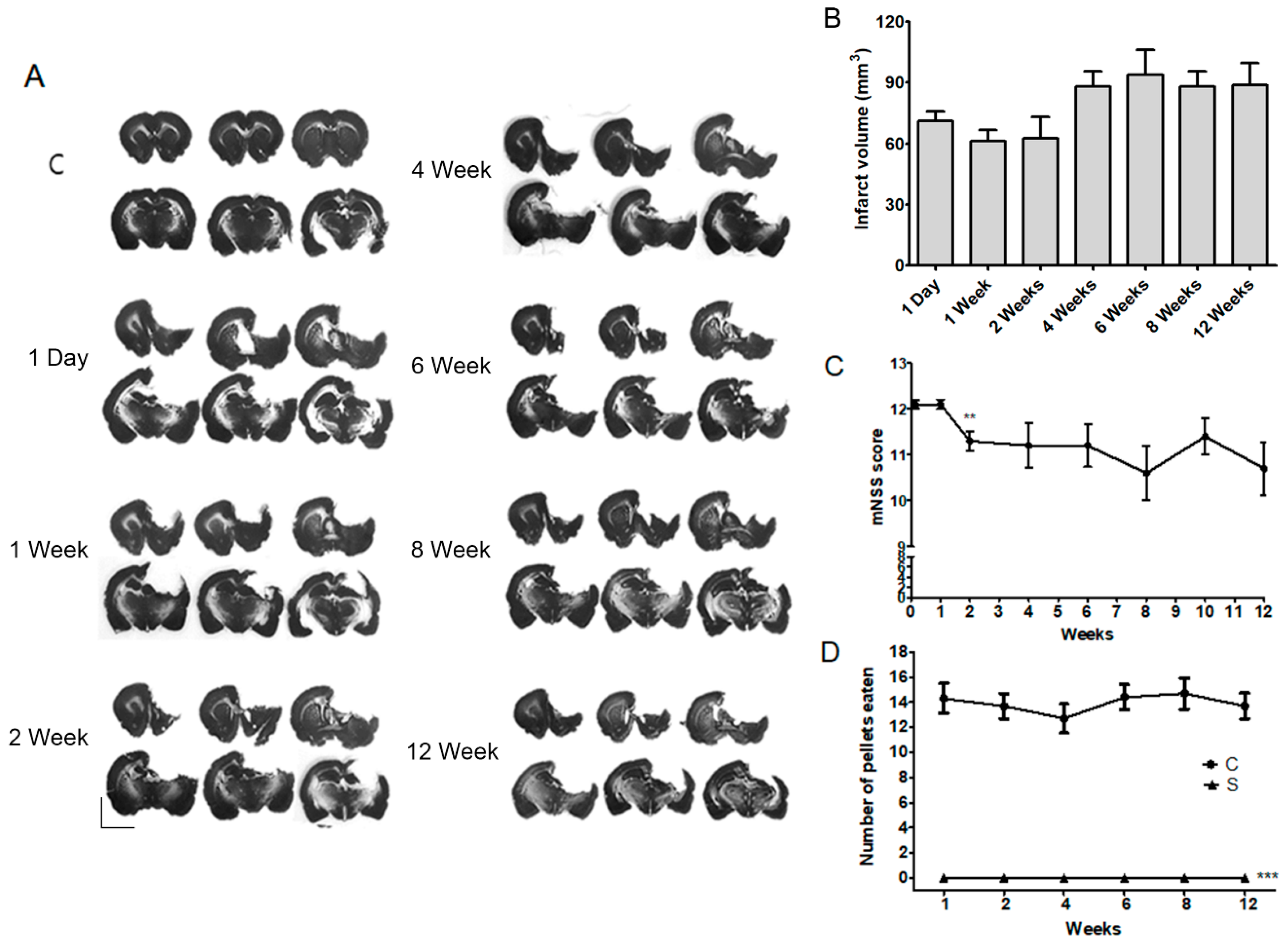
2.1. Long-Term Post-Stroke Changes, Including Functional Outcome and Brain Injury
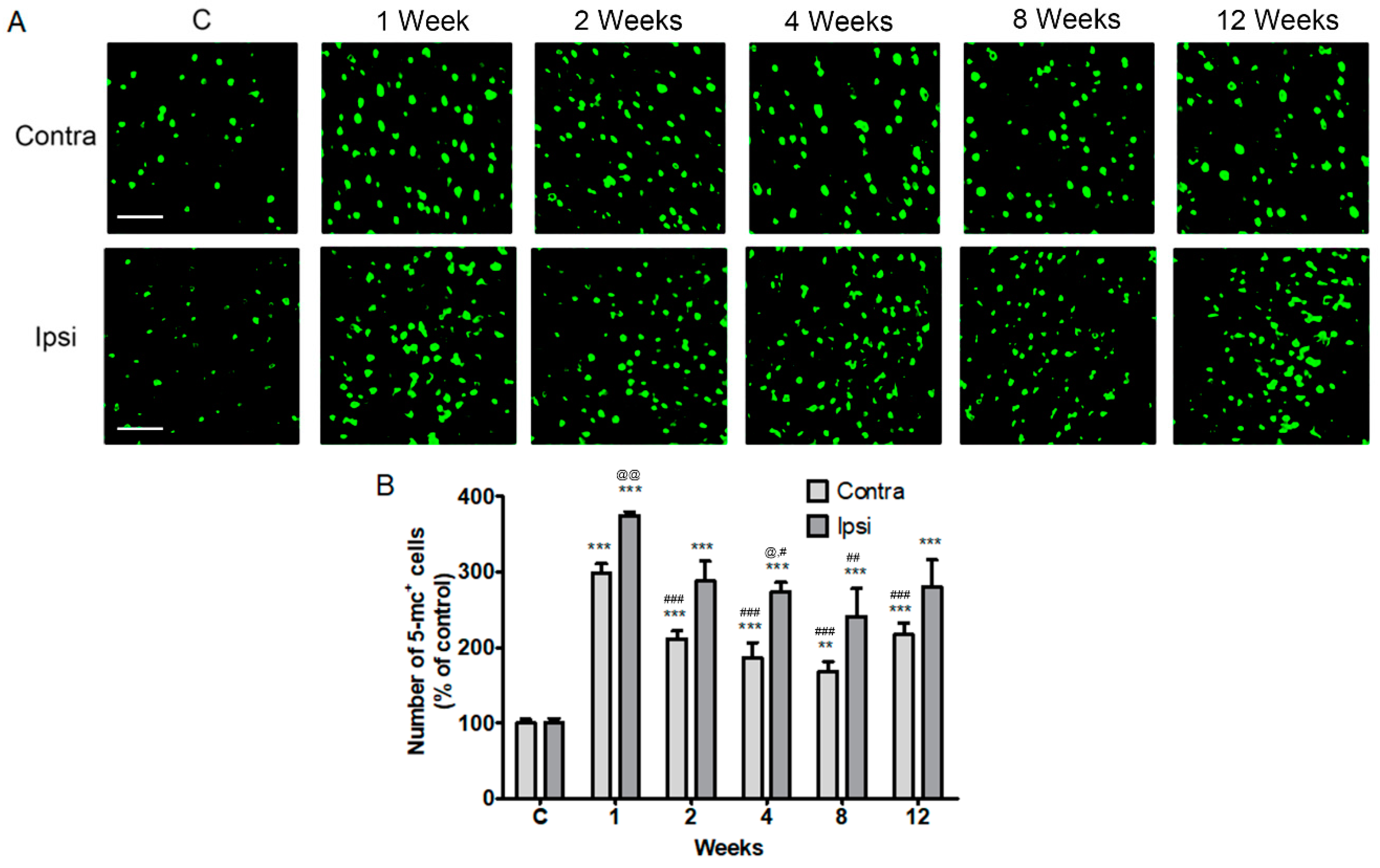
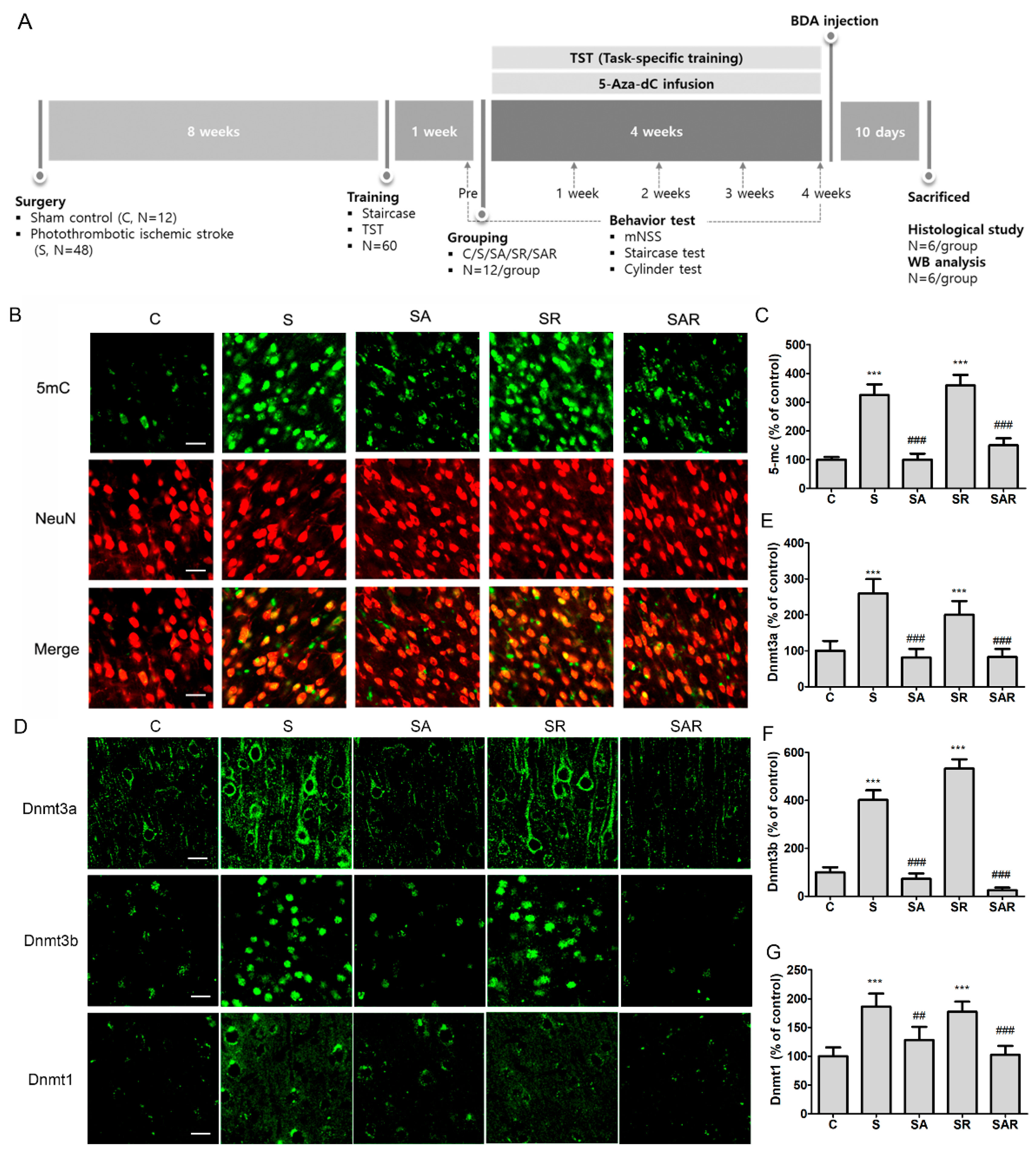
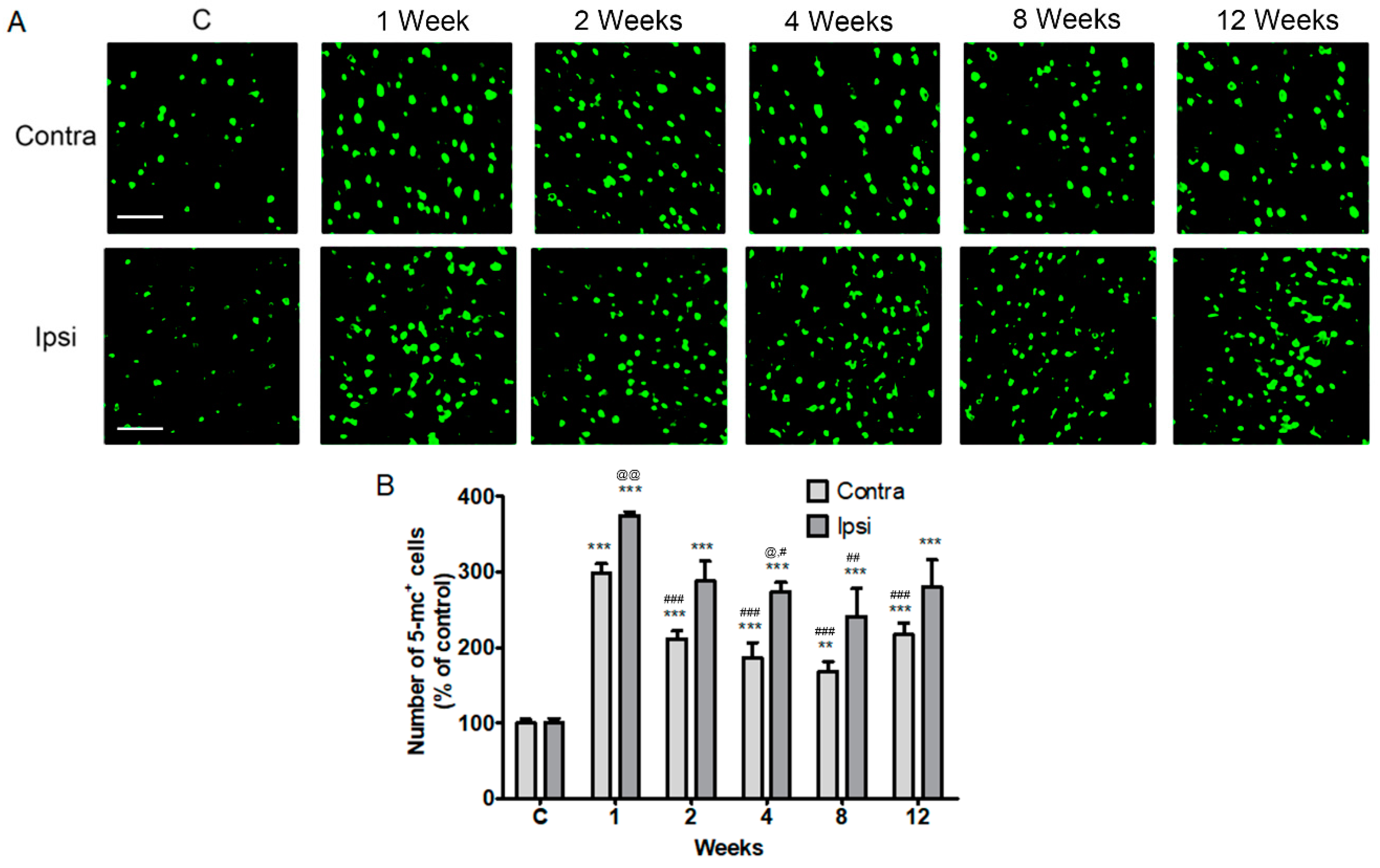
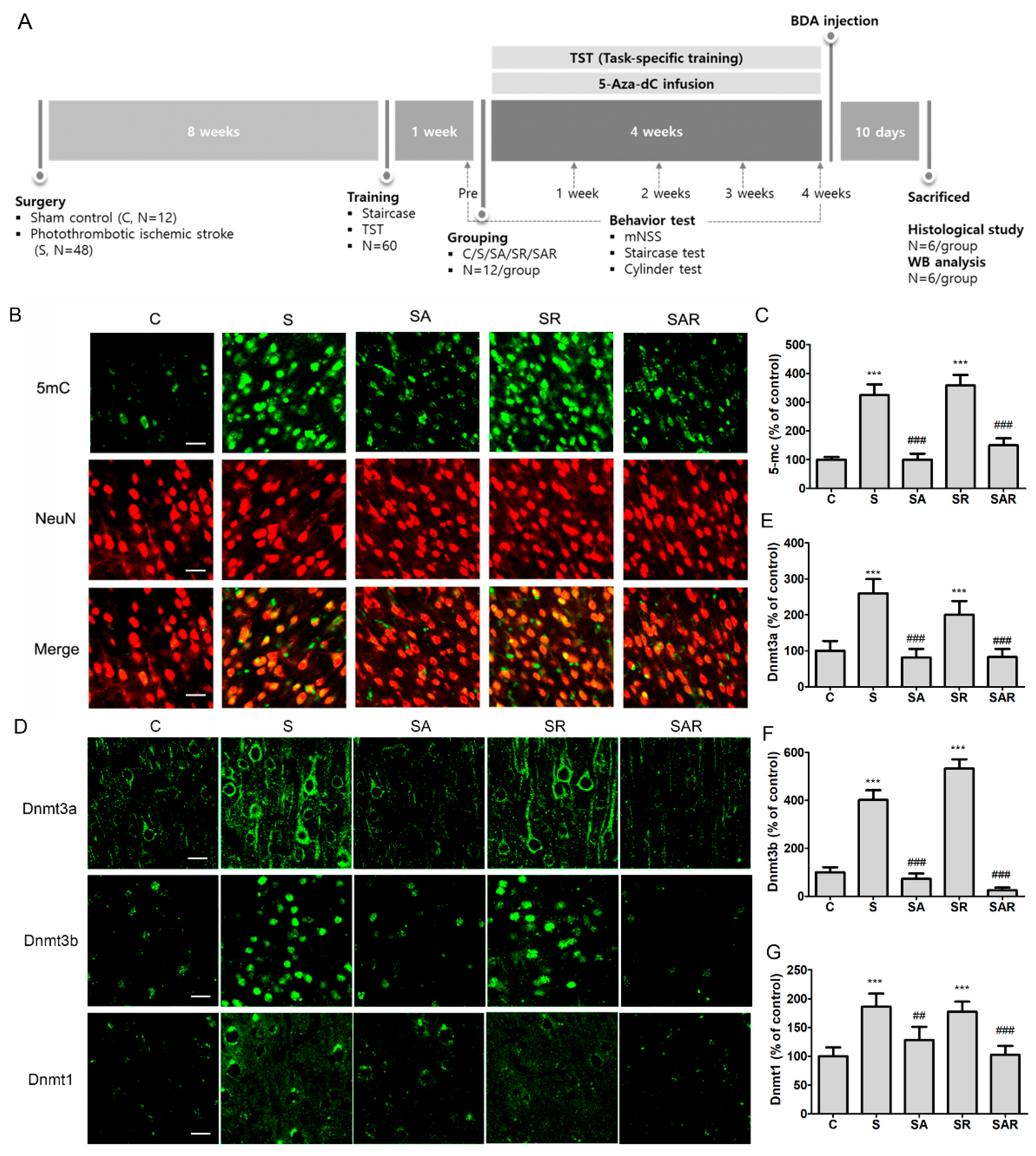
2.2. Increase in Contralateral and Ipsilateral DNA Methylation during the Chronic Phase after a Severe Stroke
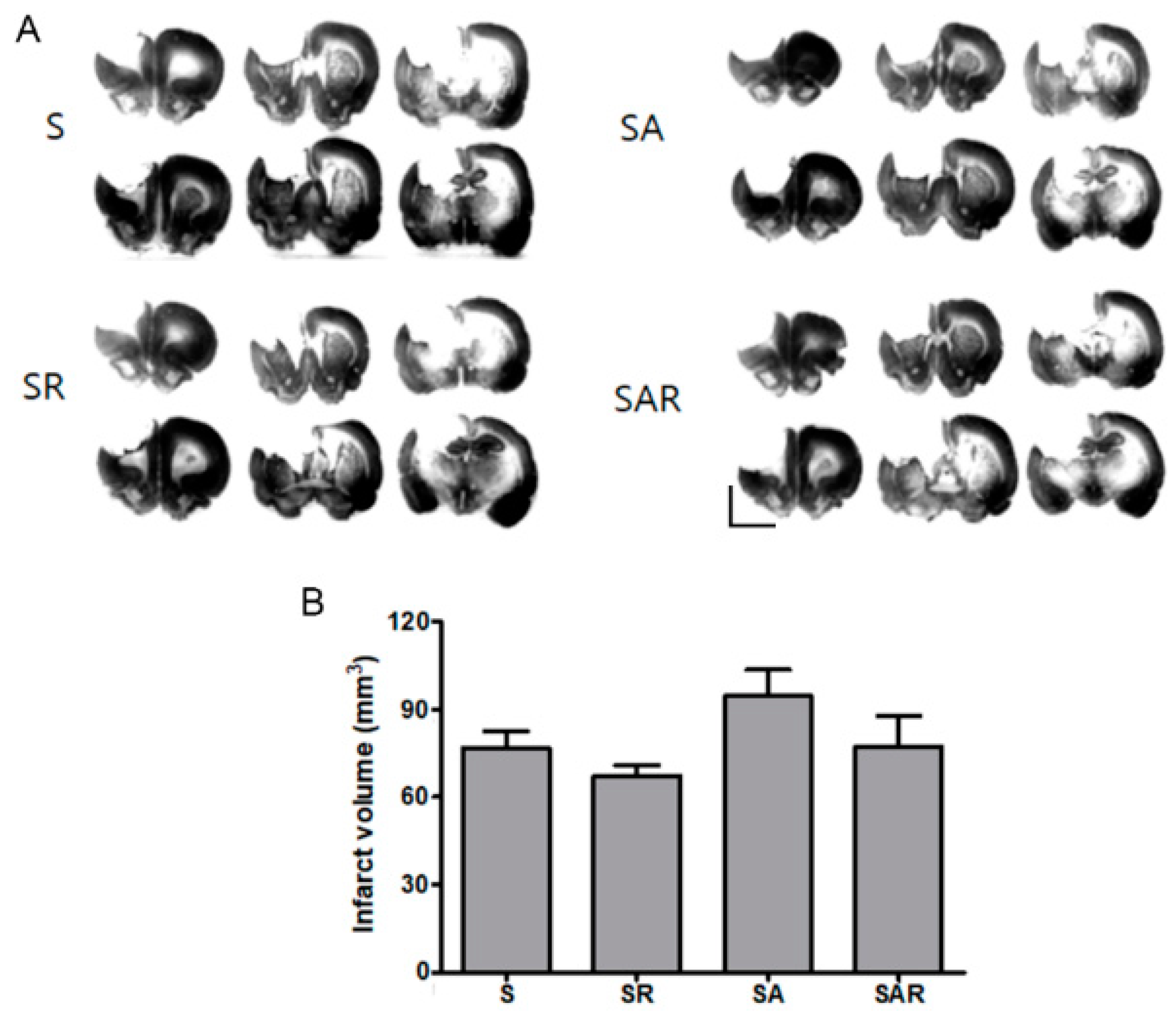
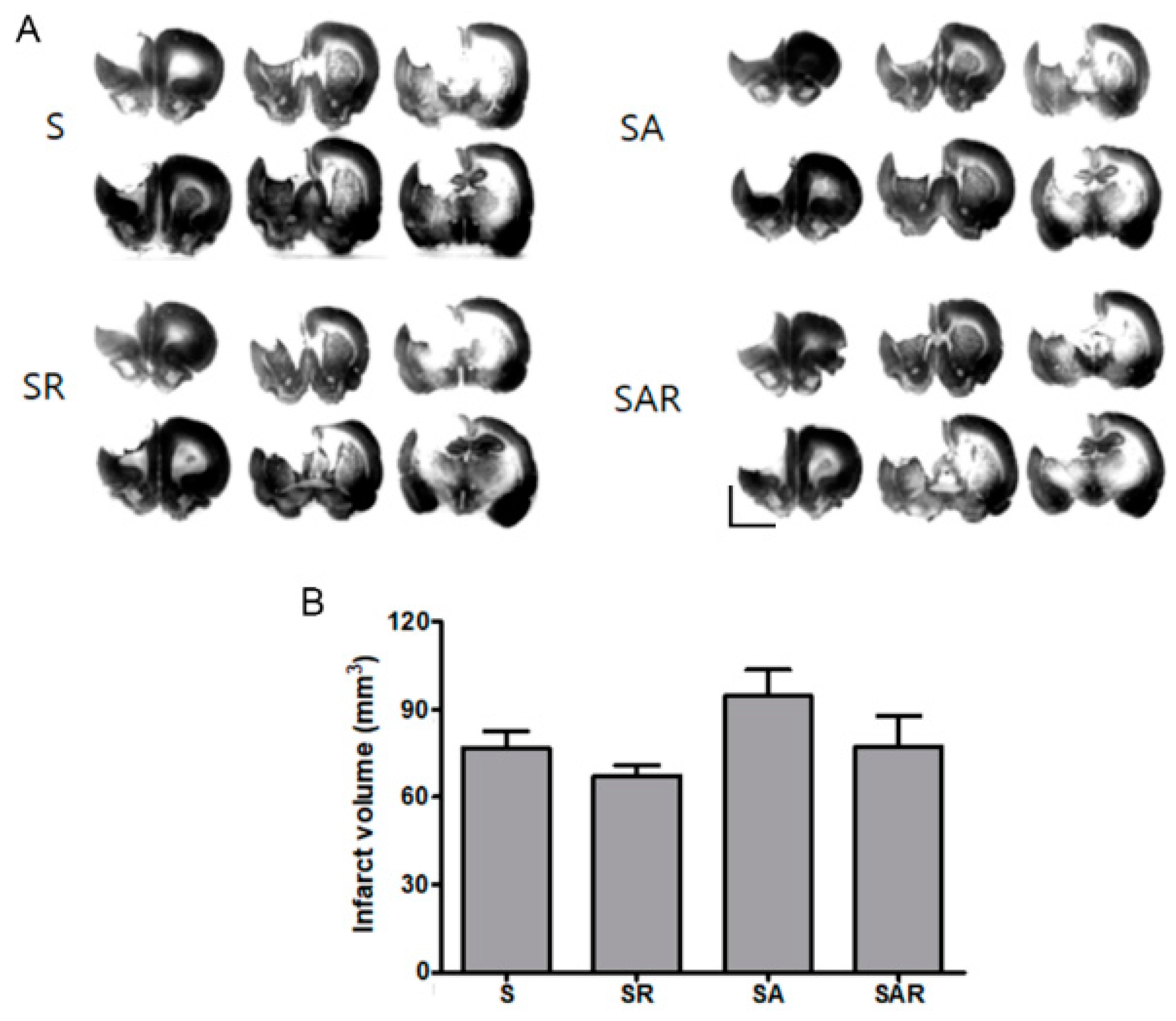
2.3. Infarct Volume Was Not Altered by TST with or without 5-Aza-dc in the Chronic Phase after Severe Stroke
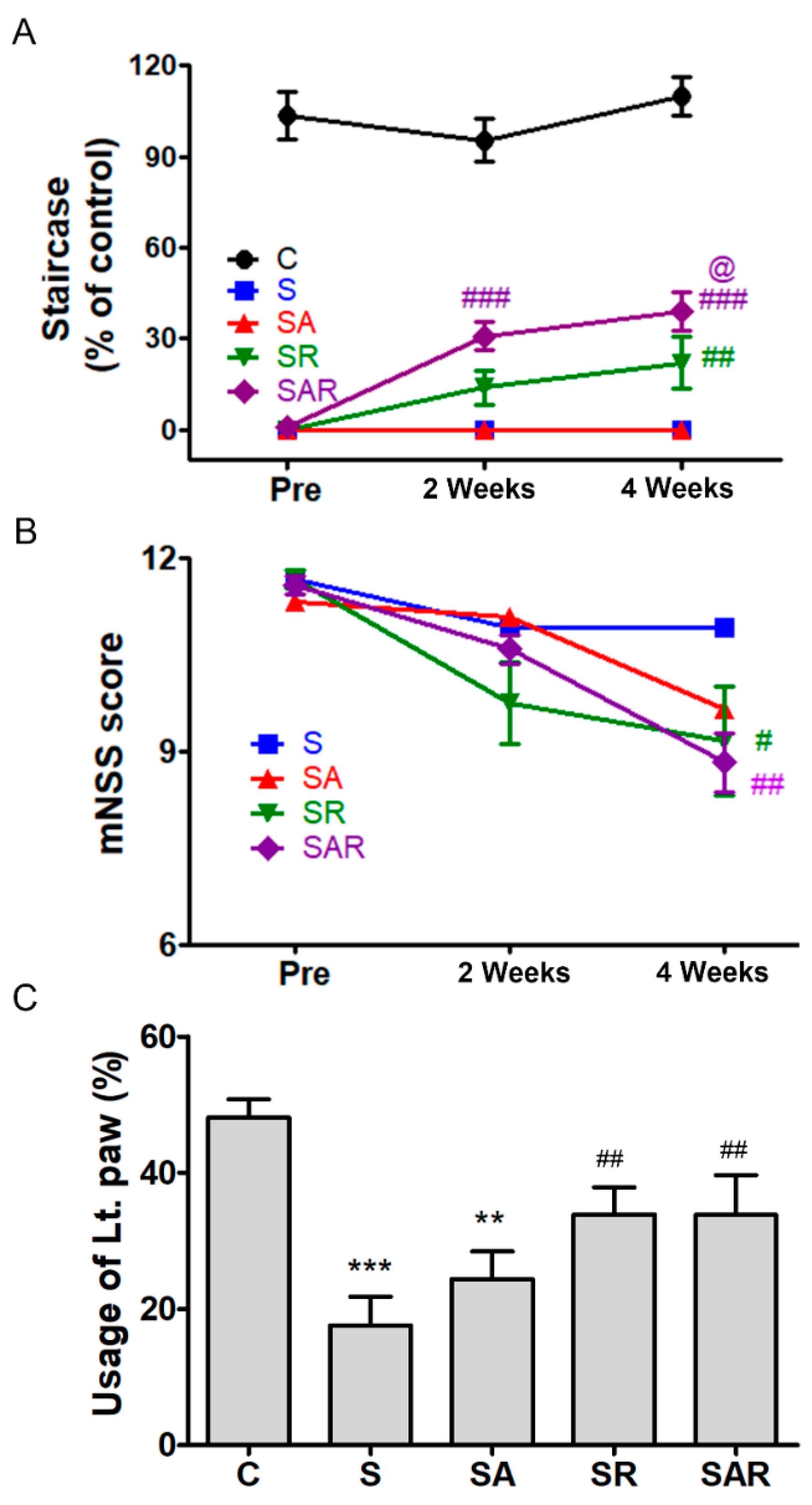
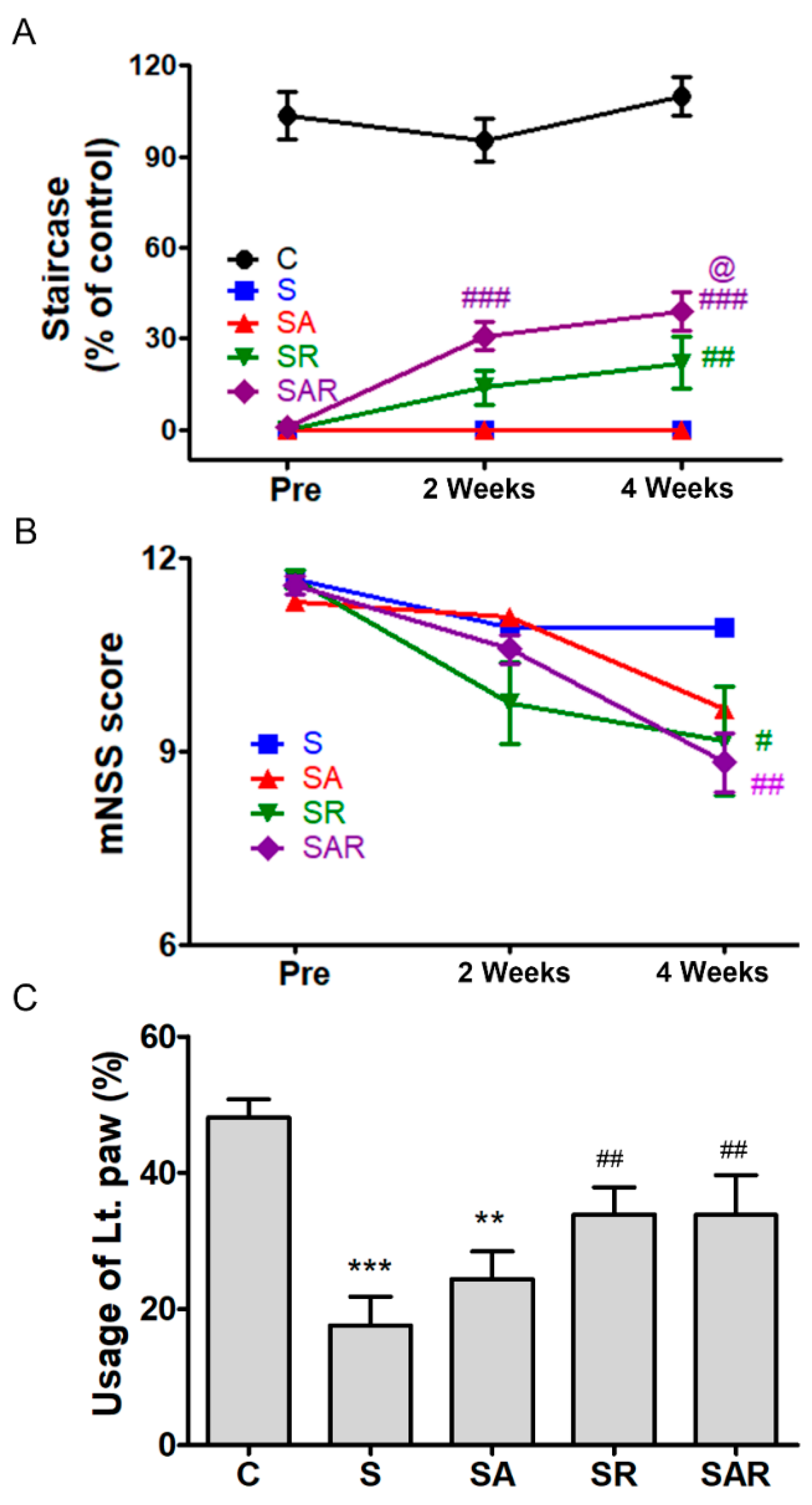
2.4. 5-Aza-dC Combined with TST Treatment Improved the Recovery of Motor Function on the Chronic Phase after Severe Stroke
2.4.1. Montoya Staircase Test
2.4.2. mNSS
2.4.3. Cylinder Test
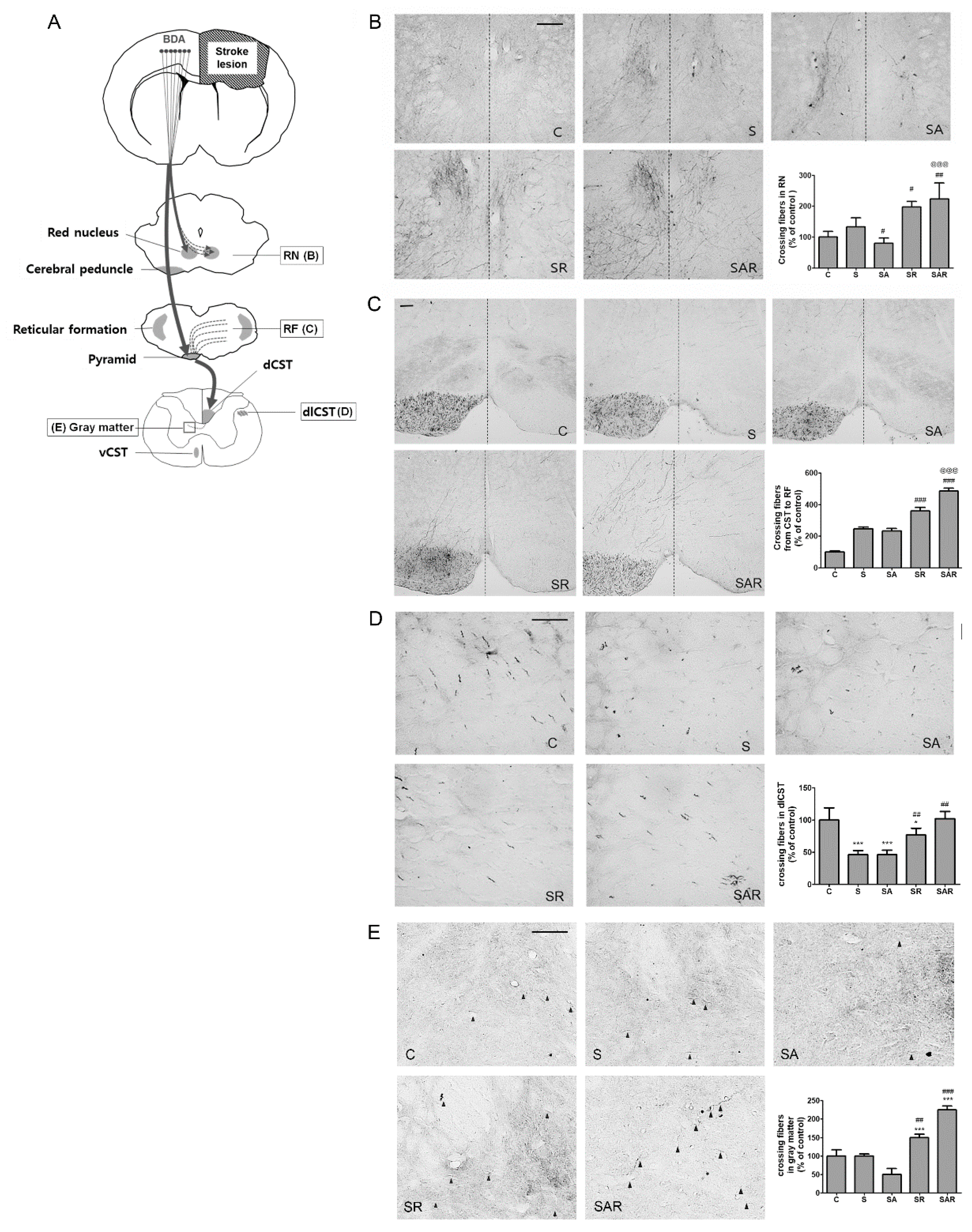
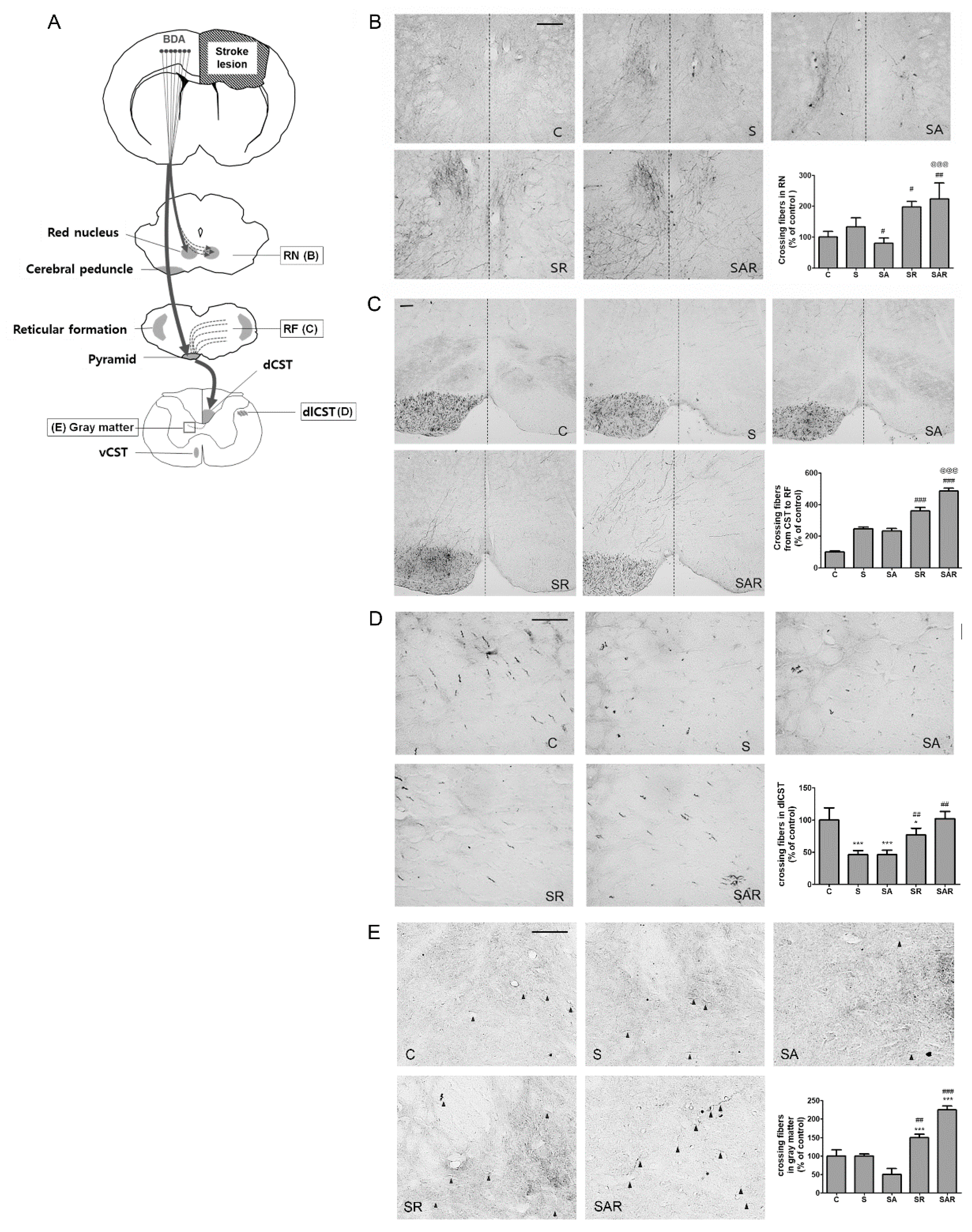
2.5. Aza-dC Treatment with TST Enhanced the Neuronal Plasticity of Motor Pathways
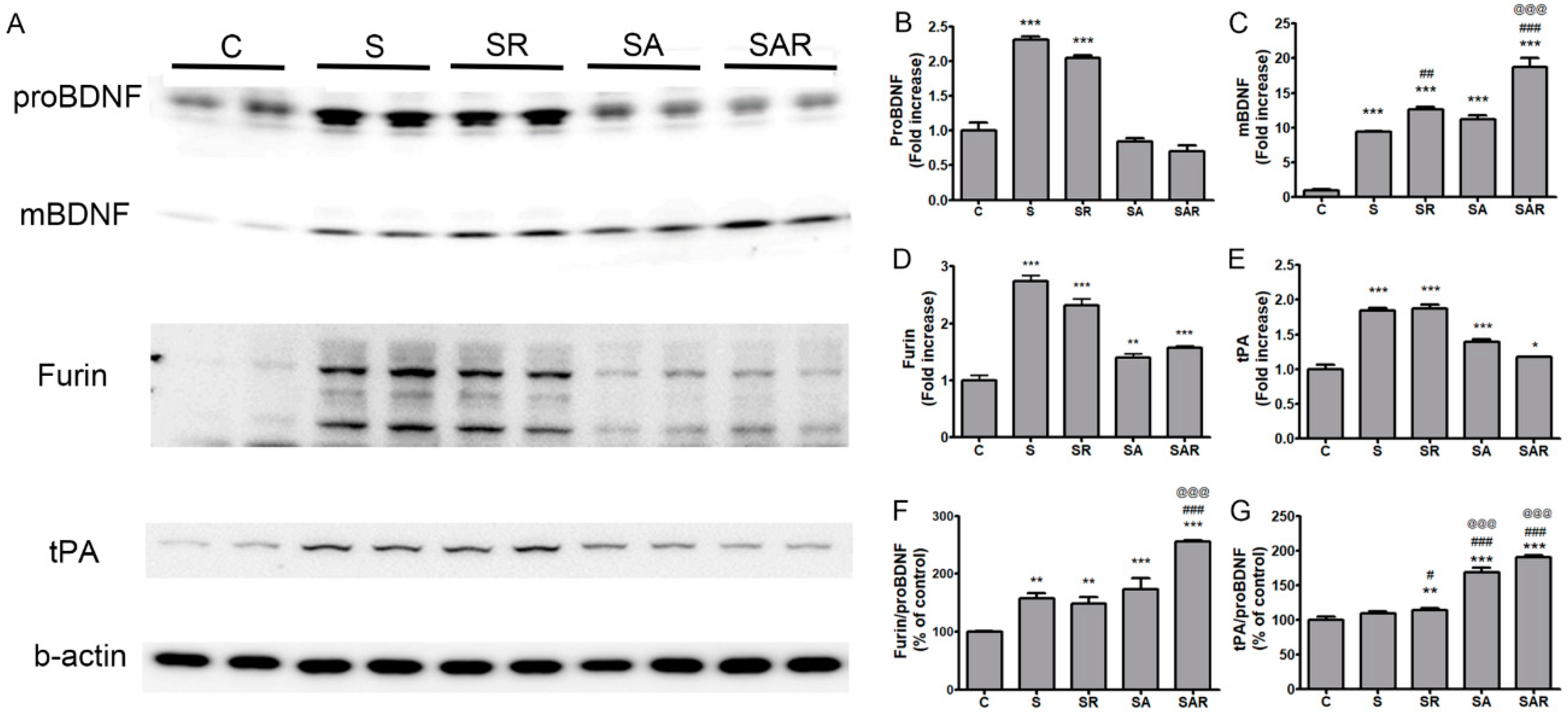
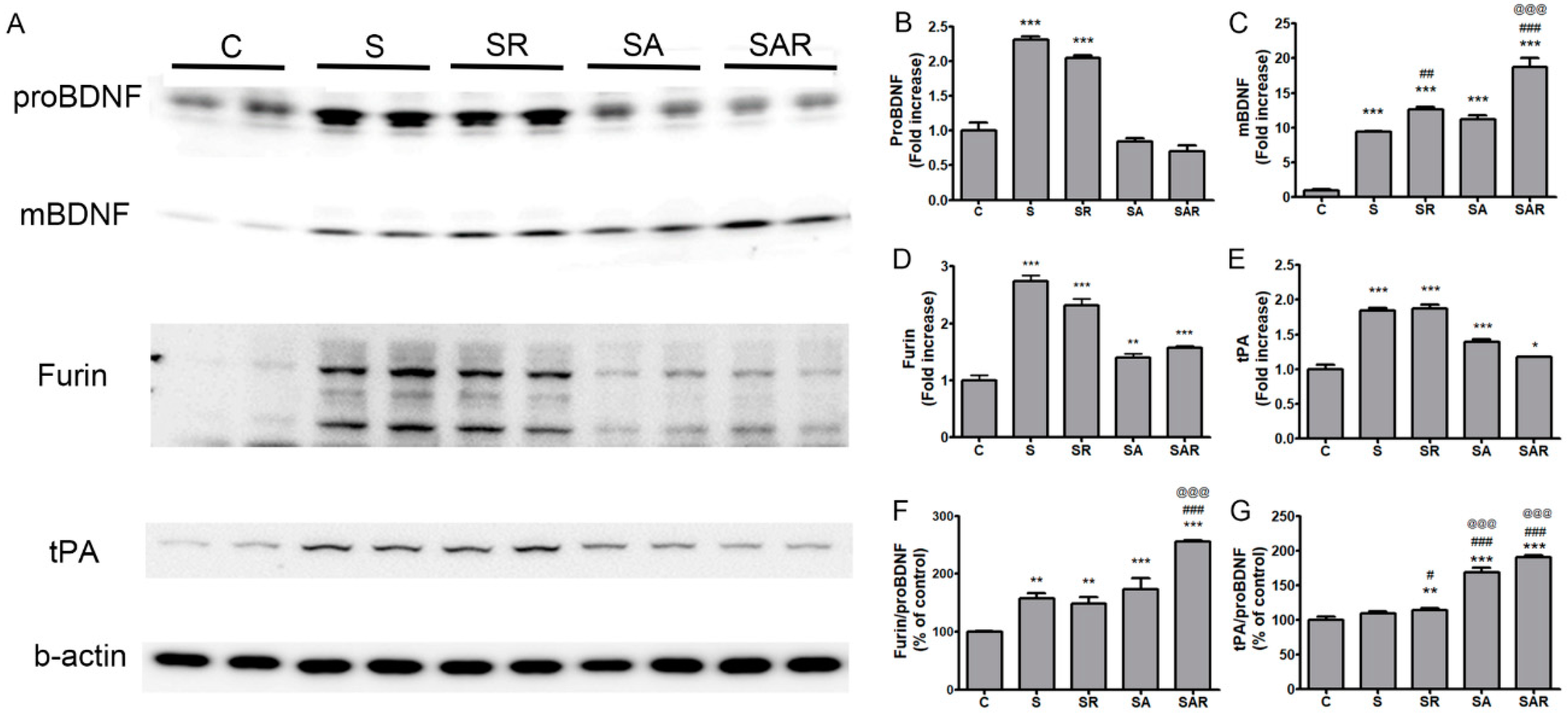
2.6. The Level of Mature Brain-Derived Neurotrophic Factor (BDNF) in the Contralateral Cortex Was Increased by 5-Aza-dc Combined with TST Treatment
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Photothrombotic Ischemia Surgery
4.3. Animal Grouping
4.4. Infusion of 5-Aza-dC in Contralateral Motor Cortex
4.5. Task-Specific Training
4.6. Staircase Test
4.7. Modified Neurological Severity Score
4.8. Cylinder Test (Asymmetrical Forelimb Use)
4.9. Video Recording
4.10. Nissle Staining
4.11. Measurement of Infarct Volume
4.12. CST Projections Using Biotinylated Dextran Amine
4.13. Immunohistochemistry
4.14. Western Blot Analysis
4.15. Double-Fluorescence Immunostaining of Tissues
4.16. Quantitative Analysis
4.17. Analysis of CST Projections
4.18. Data Analysis and Statistics
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| TST | Task specific training |
| DNMTs | DNA methyltransferases |
| 5-mc | 5-methylcytosine |
| mNSS | Modified neurological severity score |
| 5-Aza-dC | 5-aza-2′-deoxycytidine |
| C | Sham Control group |
| S | Stroke group |
| SA | 5-Aza-dC treated stroke group |
| SR | TST treated stroke group |
| SAR | 5-Aza-dC combined with TST treated stroke group |
| ANOVA | Analysis of variance |
| RN | Red nucleus |
| RF | Reticular formation |
| CST | Corticospinal tract |
| BDA | Biotinylated dextran amine |
| dlCST | Dorsolateral parts of the spinal cord |
| Pro-BDNF | Pro-brain-derived neurotrophic factor |
| mBDNF | Mature BDNF |
| t-PA | Tissue plasminogen activator |
References
- Ovbiagele, B.; Nguyen-Huynh, M.N. Stroke epidemiology: Advancing our understanding of disease mechanism and therapy. Neurotherapeutics 2011, 8, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Maulden, S.A.; Gassaway, J.; Horn, S.D.; Smout, R.J.; DeJong, G. Timing of initiation of rehabilitation after stroke. Arch. Phys. Med. Rehabil. 2005, 86, S34–S40. [Google Scholar] [CrossRef] [PubMed]
- Langhorne, P.; Bernhardt, J.; Kwakkel, G. Stroke rehabilitation. Lancet 2011, 377, 1693–1702. [Google Scholar] [CrossRef]
- Jorgensen, H.S.; Kammersgaard, L.P.; Houth, J.; Nakayama, H.; Raaschou, H.O.; Larsen, K.; Hubbe, P.; Olsen, T.S. Who benefits from treatment and rehabilitation in a stroke unit? A community-based study. Stroke 2000, 31, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Kim, J.H.; Choi, D.H.; Lee, J. Effect of task-specific training on functional recovery and corticospinal tract plasticity after stroke. Restor. Neurol. Neurosci. 2013, 31, 773–785. [Google Scholar] [PubMed]
- Petrosyan, T. Initial training facilitates posttraumatic motor recovery in rats after pyramidal tract lesion and in conditions of induced regeneration. Somatosens. Mot. Res. 2015, 32, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Okabe, N.; Himi, N.; Maruyama-Nakamura, E.; Hayashi, N.; Narita, K.; Miyamoto, O. Rehabilitative skilled forelimb training enhances axonal remodeling in the corticospinal pathway but not the brainstem-spinal pathways after photothrombotic stroke in the primary motor cortex. PLoS ONE 2017, 12, e0187413. [Google Scholar] [CrossRef] [PubMed]
- Higo, N. Effects of rehabilitative training on recovery of hand motor function: A review of animal studies. Neurosci. Res. 2014, 78, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, I.J.; Parsons, M.W.; Neilson, C.; Carey, L.M. Task-specific training: Evidence for and translation to clinical practice. Occup. Ther. Int. 2009, 16, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Buetefisch, C.M. Role of the contralesional hemisphere in post-stroke recovery of upper extremity motor function. Front. Neurol. 2015, 6, 214. [Google Scholar] [CrossRef] [PubMed]
- Dancause, N.; Touvykine, B.; Mansoori, B.K. Inhibition of the contralesional hemisphere after stroke: Reviewing a few of the building blocks with a focus on animal models. Prog. Brain Res. 2015, 218, 361–387. [Google Scholar] [PubMed]
- Classen, J.; Liepert, J.; Wise, S.P.; Hallett, M.; Cohen, L.G. Rapid plasticity of human cortical movement representation induced by practice. J. Neurophysiol. 1998, 79, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Buga, A.M.; Sascau, M.; Pisoschi, C.; Herndon, J.G.; Kessler, C.; Popa-Wagner, A. The genomic response of the ipsilateral and contralateral cortex to stroke in aged rats. J. Cell. Mol. Med. 2008, 12, 2731–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweizer, S.; Meisel, A.; Marschenz, S. Epigenetic mechanisms in cerebral ischemia. J. Cereb. Blood Flow Metab. 2013, 33, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Kogure, K.; Kato, H. Altered gene expression in cerebral ischemia. Stroke 1993, 24, 2121–2127. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, M.C.; Giffard, R.G.; Bell, B.A. An introduction to the changes in gene expression that occur after cerebral ischaemia. Br. J. Neurosurg. 2000, 14, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.; Meisel, A.; Biniszkiewicz, D.; Namura, S.; Prass, K.; Ruscher, K.; Lipski, A.; Jaenisch, R.; Moskowitz, M.A.; Dirnagl, U. DNA methyltransferase contributes to delayed ischemic brain injury. J. Neurosci. 2000, 20, 3175–3181. [Google Scholar] [CrossRef] [PubMed]
- Endres, M.; Fan, G.; Meisel, A.; Dirnagl, U.; Jaenisch, R. Effects of cerebral ischemia in mice lacking DNA methyltransferase 1 in post-mitotic neurons. Neuroreport 2001, 12, 3763–3766. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhong, B.; Tan, J.; Chen, C.; Lei, Q.; Zeng, L. The emerging role of epigenetics in cerebral ischemia. Mol. Neurobiol. 2017, 54, 1887–1905. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Ba, M.; Liang, H.; Ma, L.; Yu, Q.; Yu, T.; Wang, Y. 5′-Aza-dc sensitizes paraquat toxic effects on PC12 cell. Neurosci. Lett. 2012, 524, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Doerfler, W. In pursuit of the first recognized epigenetic signal–DNA methylation: A 1976 to 2008 synopsis. Epigenetics 2008, 3, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Narlikar, G.J.; Fan, H.Y.; Kingston, R.E. Cooperation between complexes that regulate chromatin structure and transcription. Cell 2002, 108, 475–487. [Google Scholar] [CrossRef]
- Yin, K.J.; Deng, Z.; Hamblin, M.; Xiang, Y.; Huang, H.; Zhang, J.; Jiang, X.; Wang, Y.; Chen, Y.E. Peroxisome proliferator-activated receptor delta regulation of miR-15a in ischemia-induced cerebral vascular endothelial injury. J. Neurosci. 2010, 30, 6398–6408. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liang, J.; Li, Y.; Li, J.; Yang, X.; Zhang, X.; Han, S.; Li, S.; Li, J. Down-regulation of miRNA-30a alleviates cerebral ischemic injury through enhancing beclin 1-mediated autophagy. Neurochem. Res. 2014, 39, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Han, Z.; Ji, X.; Luo, Y. Epigenetic regulation of oxidative stress in ischemic stroke. Aging Dis. 2016, 7, 295–306. [Google Scholar] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Luo, H.; Sims, N.R.; Bobrovskaya, L.; Zhou, X.F. Investigation of mature BDNF and proBDNF signaling in a rat photothrombotic ischemic model. Neurochem. Res. 2018, 43, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Takase, H.; Kurihara, Y.; Yokoyama, T.A.; Kawahara, N.; Takei, K. Lotus overexpression accelerates neuronal plasticity after focal brain ischemia in mice. PLoS ONE 2017, 12, e0184258. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, A.M.; Fouad, K.; Winship, I.R. Enhancing spinal plasticity amplifies the benefits of rehabilitative training and improves recovery from stroke. J. Neurosci. 2017, 37, 10983–10997. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Dong, J.; Tang, Y.; Ni, H.Y.; Zhang, Y.; Su, P.; Liang, H.Y.; Yao, M.C.; Yuan, H.J.; Wang, D.L.; et al. Opening a new time window for treatment of stroke by targeting HDAC2. J. Neurosci. 2017, 37, 6712–6728. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Lin, Y.H.; Ni, H.Y.; Dong, J.; Yuan, H.J.; Zhang, Y.; Liang, H.Y.; Yao, M.C.; Zhou, Q.G.; Wu, H.Y.; et al. Inhibiting histone deacetylase 2 (HDAC2) promotes functional recovery from stroke. J. Am. Heart Assoc. 2017, 6, e007236. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.C.; Crafton, K.R. Somatotopy and movement representation sites following cortical stroke. Exp. Brain Res. 2006, 168, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ueno, M.; Yamashita, T. Axonal remodeling for motor recovery after traumatic brain injury requires downregulation of gamma-aminobutyric acid signaling. Cell Death Dis. 2011, 2, e133. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.H.; Corbett, D. Plasticity during stroke recovery: From synapse to behaviour. Nat. Rev. Neurosci. 2009, 10, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, L.I.; Carmichael, S.T. Promoting axonal rewiring to improve outcome after stroke. Neurobiol. Dis. 2010, 37, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, S.T.; Wei, L.; Rovainen, C.M.; Woolsey, T.A. New patterns of intracortical projections after focal cortical stroke. Neurobiol. Dis. 2001, 8, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kim, J.E.; Sivula, M.; Strittmatter, S.M. Nogo receptor antagonism promotes stroke recovery by enhancing axonal plasticity. J. Neurosci. 2004, 24, 6209–6217. [Google Scholar] [CrossRef] [PubMed]
- Okabe, N.; Shiromoto, T.; Himi, N.; Lu, F.; Maruyama-Nakamura, E.; Narita, K.; Iwachidou, N.; Yagita, Y.; Miyamoto, O. Neural network remodeling underlying motor map reorganization induced by rehabilitative training after ischemic stroke. Neuroscience 2016, 339, 338–362. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Ueno, M.; Itokazu, T.; Yamashita, T. Bilateral movement training promotes axonal remodeling of the corticospinal tract and recovery of motor function following traumatic brain injury in mice. Cell Death Dis. 2013, 4, e534. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; Campbell, K.L.; Ross, C.J.; Boyd, L.A. Promoting neuroplasticity for motor rehabilitation after stroke: Considering the effects of aerobic exercise and genetic variation on brain-derived neurotrophic factor. Phys. Ther. 2013, 93, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Berretta, A.; Tzeng, Y.C.; Clarkson, A.N. Post-stroke recovery: The role of activity-dependent release of brain-derived neurotrophic factor. Expert Rev. Neurother. 2014, 14, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Ploughman, M.; Granter-Button, S.; Chernenko, G.; Attwood, Z.; Tucker, B.A.; Mearow, K.M.; Corbett, D. Exercise intensity influences the temporal profile of growth factors involved in neuronal plasticity following focal ischemia. Brain Res. 2007, 1150, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, F.; Zhuang, Y.; Feng, J.; Ying, Z.; Fan, G. Exercise impacts brain-derived neurotrophic factor plasticity by engaging mechanisms of epigenetic regulation. Eur. J. Neurosci. 2011, 33, 383–390. [Google Scholar] [CrossRef] [PubMed]
- MacLellan, C.L.; Keough, M.B.; Granter-Button, S.; Chernenko, G.A.; Butt, S.; Corbett, D. A critical threshold of rehabilitation involving brain-derived neurotrophic factor is required for poststroke recovery. Neurorehabil. Neural Repair 2011, 25, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Kuge, Y.; Yokota, C.; Harada, A.; Kokame, K.; Inoue, H.; Kawashima, H.; Hanzawa, H.; Shono, Y.; Saji, H.; et al. Gene and protein analysis of brain derived neurotrophic factor expression in relation to neurological recovery induced by an enriched environment in a rat stroke model. Neurosci. Lett. 2011, 495, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.C.; Sanchez-Mendoza, E.H.; Doeppner, T.R.; Hermann, D.M. Post-acute delivery of memantine promotes post-ischemic neurological recovery, peri-infarct tissue remodeling, and contralesional brain plasticity. J. Cereb. Blood Flow Metab. 2017, 37, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Ploughman, M.; Windle, V.; MacLellan, C.L.; White, N.; Dore, J.J.; Corbett, D. Brain-derived neurotrophic factor contributes to recovery of skilled reaching after focal ischemia in rats. Stroke 2009, 40, 1490–1495. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, A.N.; Overman, J.J.; Zhong, S.; Mueller, R.; Lynch, G.; Carmichael, S.T. Ampa receptor-induced local brain-derived neurotrophic factor signaling mediates motor recovery after stroke. J. Neurosci. 2011, 31, 3766–3775. [Google Scholar] [CrossRef] [PubMed]
- Madinier, A.; Bertrand, N.; Rodier, M.; Quirie, A.; Mossiat, C.; Prigent-Tessier, A.; Marie, C.; Garnier, P. Ipsilateral versus contralateral spontaneous post-stroke neuroplastic changes: Involvement of BDNF? Neuroscience 2013, 231, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Mowla, S.J.; Farhadi, H.F.; Pareek, S.; Atwal, J.K.; Morris, S.J.; Seidah, N.G.; Murphy, R.A. Biosynthesis and post-translational processing of the precursor to brain-derived neurotrophic factor. J. Biol. Chem. 2001, 276, 12660–12666. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.E.; Jung, D.H.; Lee, H.J.; Shin, M.J.; Kim, S.Y.; Shin, Y.B.; Yun, Y.J.; Shin, H.K.; Choi, B.T. Combined therapy involving electroacupuncture and treadmill exercise attenuates demyelination in the corpus callosum by stimulating oligodendrogenesis in a rat model of neonatal hypoxia-ischemia. Exp. Neurol. 2018, 300, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Pollak, J.; Yang, T.; Siddiqui, M.R.; Doyle, K.P.; Taravosh-Lahn, K.; Cekanaviciute, E.; Han, A.; Goodman, J.Z.; Jones, B.; et al. Delayed administration of a small molecule tropomyosin-related kinase B ligand promotes recovery after hypoxic-ischemic stroke. Stroke 2012, 43, 1918–1924. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.M.; Zhao, Y.Y.; Chen, S.D.; Zhang, W.H.; Lou, L.; Jin, X. Functional recovery after transplantation of neural stem cells modified by brain-derived neurotrophic factor in rats with cerebral ischaemia. J. Int. Med. Res. 2011, 39, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, I.A.; Mehler, M.F. The emerging role of epigenetics in stroke: III. Neural stem cell biology and regenerative medicine. Arch. Neurol. 2011, 68, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Ahn, J.H.; Choi, I.A.; Kim, J.H.; Kim, B.R.; Lee, J. Effect of task-specific training on Eph/ephrin expression after stroke. BMB Rep. 2016, 49, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.H.; Kim, J.H.; Lee, K.H.; Kim, H.Y.; Kim, Y.S.; Choi, W.S.; Lee, J. Role of neuronal nadph oxidase 1 in the peri-infarct regions after stroke. PLoS ONE 2015, 10, e0116814. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, I.-A.; Lee, C.S.; Kim, H.Y.; Choi, D.-H.; Lee, J. Effect of Inhibition of DNA Methylation Combined with Task-Specific Training on Chronic Stroke Recovery. Int. J. Mol. Sci. 2018, 19, 2019. https://doi.org/10.3390/ijms19072019
Choi I-A, Lee CS, Kim HY, Choi D-H, Lee J. Effect of Inhibition of DNA Methylation Combined with Task-Specific Training on Chronic Stroke Recovery. International Journal of Molecular Sciences. 2018; 19(7):2019. https://doi.org/10.3390/ijms19072019
Chicago/Turabian StyleChoi, In-Ae, Cheol Soon Lee, Hahn Young Kim, Dong-Hee Choi, and Jongmin Lee. 2018. "Effect of Inhibition of DNA Methylation Combined with Task-Specific Training on Chronic Stroke Recovery" International Journal of Molecular Sciences 19, no. 7: 2019. https://doi.org/10.3390/ijms19072019