The Role of Low-Density Lipoprotein Receptor-Related Protein 1 in Lipid Metabolism, Glucose Homeostasis and Inflammation

{kind=link}
Abstract
:1. Introduction
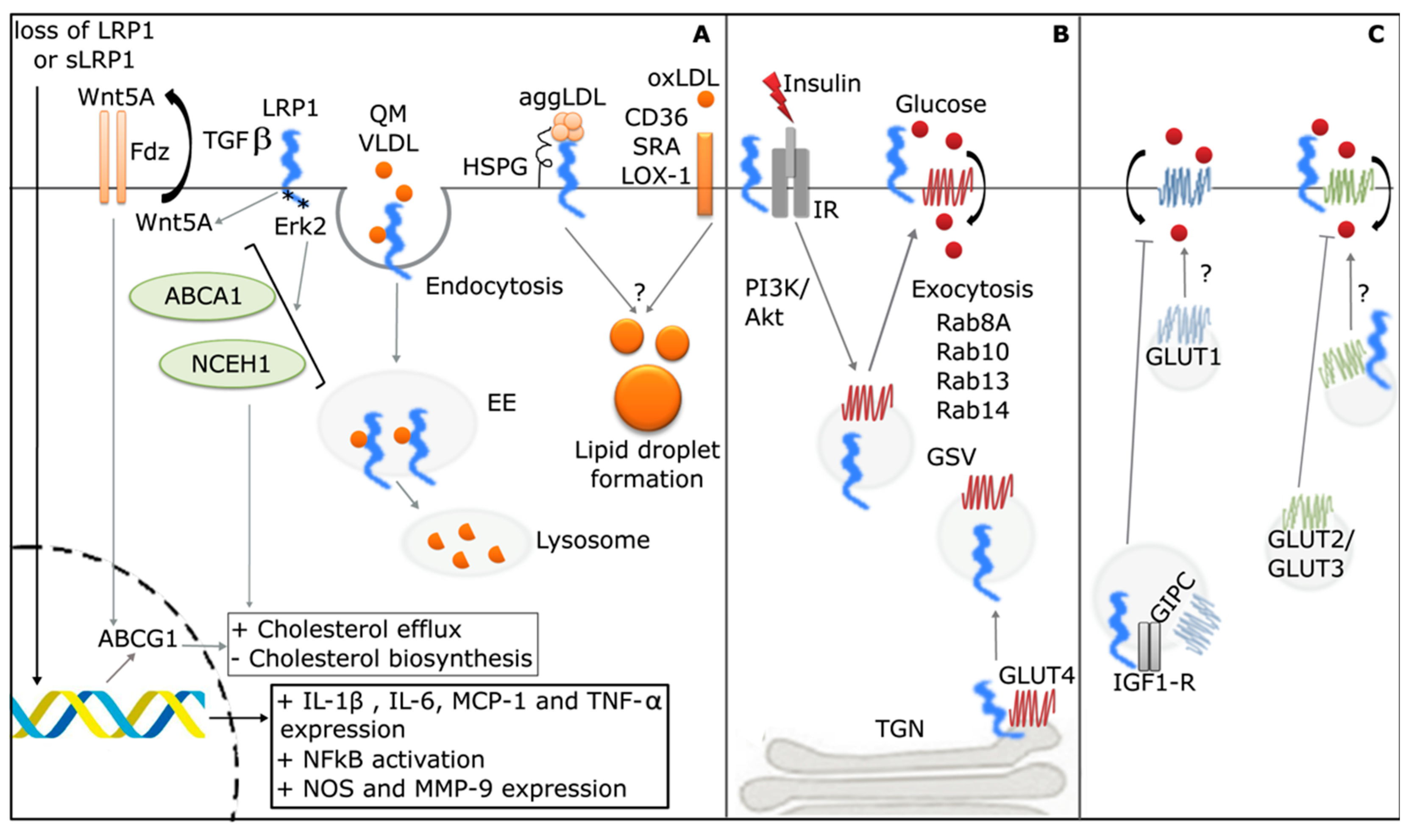
2. LRP1 in Lipoprotein Uptake and Lipid Metabolism
3. LRP1 in the Metabolic Syndrome
3.1. LRP1 in the Tissue Processing of Modified Lipoproteins
3.2. LRP1 in Glucose Homeostasis Control
4. LRP1 in Atherosclerosis
5. Conclusions and Perspectives
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCA1 | ATP binding cassette transporter A1 |
| ApoE | Apolipoprotein E |
| Akt | Protein kinase B (PKB) |
| Erk2 | Serine/threonine kinase |
| GLUTs | Glucose transporters |
| GSVs | GLUT4 storage vesicles |
| HSPG | Heparan sulfate proteoglycan |
| IGF1-R | Insulin-like growth factor 1-receptor |
| IR | Insulin receptor |
| IL-1β | Interleukin 1 β |
| IL-6 | Interleukin 6 |
| LDL | Low-density lipoprotein |
| LOX-1 | Lectin-type oxidized LDL receptor-1 |
| LRP1 | Low-density lipoprotein receptor-related protein-1 |
| MAPK | Mitogen-activated protein kinase |
| MetS | Metabolic syndrome |
| MCP-1 | Monocyte chemoattractant protein 1 |
| PI3K | Phosphatidylinositol-4,5-bisphosphate 3-kinase |
| TNF-α | Tumor necrosis factor-α |
| VLDL | Very low-density lipoprotein |
| VSMC | Vascular smooth muscle cell |
References
- Gonias, S.L.; Campana, W.M. LDL receptor-related protein-1: A regulator of inflammation in atherosclerosis, cancer, and injury to the nervous system. Am. J. Pathol. 2014, 184, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Barcelona, P.F.; Luna, J.D.; Chiabrando, G.A.; Juarez, C.P.; Bhutto, I.A.; Baba, T.; McLeod, D.S.; Sanchez, M.C.; Lutty, G.A. Immunohistochemical localization of low density lipoprotein receptor-related protein 1 and α2-Macroglobulin in retinal and choroidal tissue of proliferative retinopathies. Exp. Eye Res. 2010, 91, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, D.G.; Jaldin-Fincati, J.R.; Amigone, J.L.; Capra, R.H.; Collino, C.J.; Albertini, R.A.; Chiabrando, G.A. Standardized flow cytometry assay for identification of human monocytic heterogeneity and LRP1 expression in monocyte subpopulations: Decreased expression of this receptor in nonclassical monocytes. Cytom. Part A 2014, 85, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickland, D.K.; Gonias, S.L.; Argraves, W.S. Diverse roles for the LDL receptor family. Trends Endocrinol. Metab. 2002, 13, 66–74. [Google Scholar] [CrossRef]
- Boucher, P.; Herz, J. Signaling through LRP1: Protection from atherosclerosis and beyond. Biochem. Pharmacol. 2011, 81, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonias, S.L.; Gaultier, A.; Jo, M. Regulation of the urokinase receptor (uPAR) by LDL receptor-related protein-1 (LRP1). Curr. Pharm. Des. 2011, 17, 1962–1969. [Google Scholar] [CrossRef] [PubMed]
- Barcelona, P.F.; Jaldin-Fincati, J.R.; Sanchez, M.C.; Chiabrando, G.A. Activated α2-macroglobulin induces Muller glial cell migration by regulating MT1-MMP activity through LRP1. FASEB J. 2013, 27, 3181–3197. [Google Scholar] [CrossRef] [PubMed]
- Rabiej, V.K.; Pflanzner, T.; Wagner, T.; Goetze, K.; Storck, S.E.; Eble, J.A.; Weggen, S.; Mueller-Klieser, W.; Pietrzik, C.U. Low density lipoprotein receptor-related protein 1 mediated endocytosis of β1-integrin influences cell adhesion and cell migration. Exp. Cell Res. 2016, 340, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, D.G.; Dato, V.A.; Fincati, J.R.J.; Lorenc, V.E.; Sanchez, M.C.; Chiabrando, G.A. Activated α2-Macroglobulin Induces Mesenchymal Cellular Migration of Raw264.7 Cells Through Low-Density Lipoprotein Receptor-Related Protein 1. J. Cell. Biochem. 2017, 118, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Hu, J.; Tsai, C.W.; Yue, M.; Melrose, H.L.; Kanekiyo, T.; Bu, G. Neuronal LRP1 regulates glucose metabolism and insulin signaling in the brain. J. Neurosci. 2015, 35, 5851–5859. [Google Scholar] [CrossRef] [PubMed]
- Actis Dato, V.; Grosso, R.A.; Sanchez, M.C.; Fader, C.M.; Chiabrando, G.A. Insulin-induced exocytosis regulates the cell surface level of low density lipoprotein-related protein-1 in Muller Glial cells. Biochem. J. 2018, 475, 1669–1685. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garzon, E.; Fernandez, A.M.; Perez-Alvarez, A.; Genis, L.; Bascunana, P.; Fernandez de la Rosa, R.; Delgado, M.; Angel Pozo, M.; Moreno, E.; McCormick, P.J.; et al. The insulin-like growth factor I receptor regulates glucose transport by astrocytes. Glia 2016, 64, 1962–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaldin-Fincati, J.R.; Pavarotti, M.; Frendo-Cumbo, S.; Bilan, P.J.; Klip, A. Update on GLUT4 Vesicle Traffic: A Cornerstone of Insulin Action. Trends Endocrinol. Metab. 2017, 28, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Pirola, C.J. Metabolic syndrome: From the genetics to the pathophysiology. Curr. Hypertens. Rep. 2011, 13, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Costales, P.; Fuentes-Prior, P.; Castellano, J.; Revuelta-Lopez, E.; Corral-Rodriguez, M.A.; Nasarre, L.; Badimon, L.; Llorente-Cortes, V. K Domain CR9 of Low Density Lipoprotein (LDL) Receptor-related Protein 1 (LRP1) Is Critical for Aggregated LDL-induced Foam Cell Formation from Human Vascular Smooth Muscle Cells. J. Biol. Chem. 2015, 290, 14852–14865. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Xian, X.; Holland, W.L.; Tsai, S.; Herz, J. Low-Density Lipoprotein Receptor-Related Protein-1 Protects Against Hepatic Insulin Resistance and Hepatic Steatosis. EBioMedicine 2016, 7, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Au, D.T.; Strickland, D.K.; Muratoglu, S.C. The LDL Receptor-Related Protein 1: At the Crossroads of Lipoprotein Metabolism and Insulin Signaling. J. Diabetes Res. 2017, 2017, 8356537. [Google Scholar] [CrossRef] [PubMed]
- Bogan, J.S. Regulation of glucose transporter translocation in health and diabetes. Annu. Rev. Biochem. 2012, 81, 507–532. [Google Scholar] [CrossRef] [PubMed]
- Schaftenaar, F.; Frodermann, V.; Kuiper, J.; Lutgens, E. Atherosclerosis: The interplay between lipids and immune cells. Curr. Opin. Lipidol. 2016, 27, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Mantuano, E.; Brifault, C.; Lam, M.S.; Azmoon, P.; Gilder, A.S.; Gonias, S.L. LDL receptor-related protein-1 regulates NFκB and microRNA-155 in macrophages to control the inflammatory response. Proc. Natl. Acad. Sci. USA 2016, 113, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, A.; Vassiliou, G.; Benoist, F.; McPherson, R. Adipocyte low density lipoprotein receptor-related protein gene expression and function is regulated by peroxisome proliferator-activated receptor γ. J. Biol. Chem. 2003, 278, 11945–11953. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Hiatt, W.R.; Kaul, S.; Smith, R.J. The cardiovascular safety of diabetes drugs—Insights from the rosiglitazone experience. N. Engl. J. Med. 2013, 369, 1285–1287. [Google Scholar] [CrossRef] [PubMed]
- Rondon-Ortiz, A.N.; Lino Cardenas, C.L.; Martinez-Malaga, J.; Gonzales-Urday, A.L.; Gugnani, K.S.; Bohlke, M.; Maher, T.J.; Pino-Figueroa, A.J. High Concentrations of Rosiglitazone Reduce mRNA and Protein Levels of LRP1 in HepG2 Cells. Front. Pharmacol. 2017, 8, 772. [Google Scholar] [CrossRef] [PubMed]
- Vucinic, N.; Stokic, E.; Djan, I.; Obreht, D.; Velickovic, N.; Stankov, K.; Djan, M. The LRP1 Gene Polymorphism is associated with Increased Risk of Metabolic Syndrome Prevalence in the Serbian Population. Balkan J. Med. Genet. 2017, 20, 51–58. [Google Scholar] [PubMed]
- Delgado-Lista, J.; Perez-Martinez, P.; Solivera, J.; Garcia-Rios, A.; Perez-Caballero, A.I.; Lovegrove, J.A.; Drevon, C.A.; Defoort, C.; Blaak, E.E.; Dembinska-Kieć, A. Top single nucleotide polymorphisms affecting carbohydrate metabolism in metabolic syndrome: From the LIPGENE study. J. Clin. Endocrinol. Metab. 2014, 99, E384–E389. [Google Scholar] [CrossRef] [PubMed]
- Mahley, R.W.; Huang, Y. Atherogenic remnant lipoproteins: Role for proteoglycans in trapping, transferring, and internalizing. J. Clin. Investig. 2007, 117, 94–98. [Google Scholar] [CrossRef] [PubMed]
- May, P.; Bock, H.H.; Herz, J. Integration of endocytosis and signal transduction by lipoprotein receptors. Sci. Signal. 2003, 2003, PE12. [Google Scholar] [CrossRef] [PubMed]
- Rensen, P.C.; Jong, M.C.; van Vark, L.C.; van der Boom, H.; Hendriks, W.L.; van Berkel, T.J.; Biessen, E.A.; Havekes, L.M. Apolipoprotein E is resistant to intracellular degradation in vitro and in vivo. Evidence for retroendocytosis. J. Biol. Chem. 2000, 275, 8564–8571. [Google Scholar] [CrossRef] [PubMed]
- Laatsch, A.; Panteli, M.; Sornsakrin, M.; Hoffzimmer, B.; Grewal, T.; Heeren, J. Low density lipoprotein receptor-related protein 1 dependent endosomal trapping and recycling of apolipoprotein E. PLoS ONE 2012, 7, e29385. [Google Scholar] [CrossRef] [PubMed]
- Lillis, A.P.; Mikhailenko, I.; Strickland, D.K. Beyond endocytosis: LRP function in cell migration, proliferation and vascular permeability. J. Thromb. Haemost. 2005, 3, 1884–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeren, J.; Beisiegel, U.; Grewal, T. Apolipoprotein E recycling: Implications for dyslipidemia and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Kowal, R.C.; Herz, J.; Goldstein, J.L.; Esser, V.; Brown, M.S. Low density lipoprotein receptor-related protein mediates uptake of cholesteryl esters derived from apoprotein E-enriched lipoproteins. Proc. Natl. Acad. Sci. USA 1989, 86, 5810–5814. [Google Scholar] [CrossRef] [PubMed]
- Beisiegel, U.; Weber, W.; Ihrke, G.; Herz, J.; Stanley, K.K. The LDL-receptor-related protein, LRP, is an apolipoprotein E-binding protein. Nature 1989, 341, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Beisiegel, U.; Weber, W.; Bengtsson-Olivecrona, G. Lipoprotein lipase enhances the binding of chylomicrons to low density lipoprotein receptor-related protein. Proc. Natl. Acad. Sci. USA 1991, 88, 8342–8346. [Google Scholar] [CrossRef] [PubMed]
- Heeren, J.; Niemeier, A.; Merkel, M.; Beisiegel, U. Endothelial-derived lipoprotein lipase is bound to postprandial triglyceride-rich lipoproteins and mediates their hepatic clearance in vivo. J. Mol. Med. 2002, 80, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Laatsch, A.; Merkel, M.; Talmud, P.J.; Grewal, T.; Beisiegel, U.; Heeren, J. Insulin stimulates hepatic low density lipoprotein receptor-related protein 1 (LRP1) to increase postprandial lipoprotein clearance. Atherosclerosis 2009, 204, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.Y.; Yu, B.L.; Xie, Y.F.; Zhao, S.P.; Wu, C.L. Apolipoprotein A5 regulates intracellular triglyceride metabolism in adipocytes. Mol. Med. Rep. 2017, 16, 6771–6779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillis, A.P.; Muratoglu, S.C.; Au, D.T.; Migliorini, M.; Lee, M.J.; Fried, S.K.; Mikhailenko, I.; Strickland, D.K. LDL Receptor-Related Protein-1 (LRP1) Regulates Cholesterol Accumulation in Macrophages. PLoS ONE 2015, 10, e0128903. [Google Scholar] [CrossRef] [PubMed]
- Xian, X.; Ding, Y.; Dieckmann, M.; Zhou, L.; Plattner, F.; Liu, M.; Parks, J.S.; Hammer, R.E.; Boucher, P.; Tsai, S.; et al. LRP1 integrates murine macrophage cholesterol homeostasis and inflammatory responses in atherosclerosis. eLife 2017, 6, e29292. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Khalil, A.; Larbi, A. The role of elastin peptides in modulating the immune response in aging and age-related diseases. Pathol. Biol. 2012, 60, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Samouillan, V.; Dandurand, J.; Nasarre, L.; Badimon, L.; Lacabanne, C.; Llorente-Cortes, V. Lipid loading of human vascular smooth muscle cells induces changes in tropoelastin protein levels and physical structure. Biophys. J. 2012, 103, 532–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samouillan, V.; Revuelta-Lopez, E.; Dandurand, J.; Nasarre, L.; Badimon, L.; Lacabanne, C.; Llorente-Cortés, V. Cardiomyocyte intracellular cholesteryl ester accumulation promotes tropoelastin physical alteration and degradation: Role of LRP1 and cathepsin S. Int. J. Biochem. Cell Biol. 2014, 55, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Asmar, Z.; Terrand, J.; Jenty, M.; Host, L.; Mlih, M.; Zerr, A.; Justiniano, H.; Matz, R.L.; Boudier, C.; Scholler, E.; et al. Convergent Signaling Pathways Controlled by LRP1 (Receptor-related Protein 1) Cytoplasmic and Extracellular Domains Limit Cellular Cholesterol Accumulation. J. Biol. Chem. 2016, 291, 5116–5127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alique, M.; Luna, C.; Carracedo, J.; Ramirez, R. LDL biochemical modifications: A link between atherosclerosis and aging. Food Nutr. Res. 2015, 59, 29240. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.C.; Chang, P.Y.; Kuo, T.L.; Lu, S.C. Electronegative L5-LDL induces the production of G-CSF and GM-CSF in human macrophages through LOX-1 involving NF-κB and ERK2 activation. Atherosclerosis 2017, 267, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.C.; Chang, P.Y.; Lu, S.C. L5-LDL from ST-elevation myocardial infarction patients induces IL-1β production via LOX-1 and NLRP3 inflammasome activation in macrophages. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H265–H274. [Google Scholar] [CrossRef] [PubMed]
- Saneipour, M.; Ghatreh-Samani, K.; Heydarian, E.; Farrokhi, E.; Abdian, N. Adiponectin inhibits oxidized low density lipoprotein-induced increase in matrix metalloproteinase 9 expression in vascular smooth muscle cells. ARYA Atheroscler. 2015, 11, 191–195. [Google Scholar] [PubMed]
- Lin, J.; Zhou, S.; Zhao, T.; Ju, T.; Zhang, L. TRPM7 channel regulates ox-LDL-induced proliferation and migration of vascular smooth muscle cells via MEK-ERK pathways. FEBS Lett. 2016, 590, 520–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boren, J.; Williams, K.J. The central role of arterial retention of cholesterol-rich apolipoprotein-B-containing lipoproteins in the pathogenesis of atherosclerosis: A triumph of simplicity. Curr. Opin. Lipidol. 2016, 27, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.Y.; Chan, H.C.; Chan, H.C.; Kalu, F.C.U.; Lee, H.C.; Lin, I.L.; Jhuo, S.J.; Lai, W.T.; Tsao, C.R.; Sawamura, T.; et al. Electronegative Low-Density Lipoprotein L5 Induces Adipose Tissue Inflammation Associated With Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2017, 102, 4615–4625. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Su, W.; Ding, Z.; Wang, X.; Mercanti, F.; Chen, M.; Raina, S.; Mehta, J.L. Regulation of MSR-1 and CD36 in macrophages by LOX-1 mediated through PPAR-γ. Biochem. Biophys. Res. Commun. 2013, 431, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, R.; Henkels, K.M.; Wrenshall, L.E.; Kanaho, Y.; Paolo, G.D.; Frohman, M.A.; Gomez-Cambronero, J. Oxidized LDL phagocytosis during foam cell formation in atherosclerotic plaques relies on a PLD2-CD36 functional interdependence. J. Leukoc. Biol. 2018, 103, 867–883. [Google Scholar] [CrossRef] [PubMed]
- Llorente-Cortes, V.; Otero-Vinas, M.; Badimon, L. Differential role of heparan sulfate proteoglycans on aggregated LDL uptake in human vascular smooth muscle cells and mouse embryonic fibroblasts. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Dissmore, T.; Seye, C.I.; Medeiros, D.M.; Weisman, G.A.; Bradford, B.; Mamedova, L. The P2Y2 receptor mediates uptake of matrix-retained and aggregated low density lipoprotein in primary vascular smooth muscle cells. Atherosclerosis 2016, 252, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Buckler-Pena, D.; Nauta, T.; Singh, M.; Asmar, A.; Shi, J.; Kim, J.Y.; Kandror, K.V. Insulin responsiveness of glucose transporter 4 in 3T3-L1 cells depends on the presence of sortilin. Mol. Biol. Cell 2013, 24, 3115–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorens, B. GLUT2, glucose sensing and glucose homeostasis. Diabetologia 2015, 58, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Jedrychowski, M.P.; Gartner, C.A.; Gygi, S.P.; Zhou, L.; Herz, J.; Kandror, K.V.; Pilch, P.F. Proteomic analysis of GLUT4 storage vesicles reveals LRP1 to be an important vesicle component and target of insulin signaling. J. Biol. Chem. 2010, 285, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zaarur, N.; Singh, M.; Morin, P.; Kandror, K.V. Sortilin and retromer mediate retrograde transport of Glut4 in 3T3-L1 adipocytes. Mol. Biol. Cell 2017, 28, 1667–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muratoglu, S.C.; Belgrave, S.; Lillis, A.P.; Migliorini, M.; Robinson, S.; Smith, E.; Zhang, L.; Strickland, D.K. Macrophage LRP1 suppresses neo-intima formation during vascular remodeling by modulating the TGF-β signaling pathway. PLoS ONE 2011, 6, e28846. [Google Scholar] [CrossRef] [PubMed]
- Baitsch, D.; Bock, H.H.; Engel, T.; Telgmann, R.; Muller-Tidow, C.; Varga, G.; Bot, M.; Herz, J.; Robenek, H.; von Eckardstein, A.; et al. Apolipoprotein E induces antiinflammatory phenotype in macrophages. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Overton, C.D.; Yancey, P.G.; Major, A.S.; Linton, M.F.; Fazio, S. Deletion of macrophage LDL receptor-related protein increases atherogenesis in the mouse. Circ. Res. 2007, 100, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Giunzioni, I.; Tavori, H.; Covarrubias, R.; Ding, L.; Zhang, Y.; Ormseth, M.; Major, A.S.; Stafford, J.M.; Linton, M.F.; et al. Loss of Macrophage Low-Density Lipoprotein Receptor-Related Protein 1 Confers Resistance to the Antiatherogenic Effects of Tumor Necrosis Factor-α Inhibition. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1483–1495. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.; Chatterjee, T.K.; Tang, Y.; Hui, D.Y.; Weintraub, N.L. Proinflammatory phenotype of perivascular adipocytes. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.K.; Zhou, Z.; Zhang, J.; Zeng, R.; Wu, J.; Eitzman, D.T.; Chen, Y.E.; Chang, L. Perivascular adipose tissue in vascular function and disease: A review of current research and animal models. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Konaniah, E.S.; Kuhel, D.G.; Basford, J.E.; Weintraub, N.L.; Hui, D.Y. Deficiency of LRP1 in Mature Adipocytes Promotes Diet-Induced Inflammation and Atherosclerosis-Brief Report. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1046–1049. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.G.; Blakemore, J.; Ding, L.; Fan, D.; Overton, C.D.; Zhang, Y.; Linton, M.F.; Fazio, S. Macrophage LRP-1 controls plaque cellularity by regulating efferocytosis and Akt activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, M.; Hayes, C.D.; Thome, J.J.; Thorp, E.; Matsushima, G.K.; Herz, J.; Farber, D.L.; Liu, K.; Lakshmana, M.; Tabas, I. An AXL/LRP-1/RANBP9 complex mediates DC efferocytosis and antigen cross-presentation in vivo. J. Clin. Investig. 2014, 124, 1296–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, P.A.; Zhu, L.; Tavori, H.; Huynh, K.; Giunzioni, I.; Stafford, J.M.; Linton, M.F.; Fazio, S. Deletion of Macrophage Low-Density Lipoprotein Receptor-Related Protein 1 (LRP1) Accelerates Atherosclerosis Regression and Increases CCR7 Expression in Plaque Macrophages. Circulation 2018. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Calvo, D.; Colom, C.; Vilades, D.; Rivas-Urbina, A.; Moustafa, A.H.; Perez-Cuellar, M.; Sánchez-Quesada, J.L.; Pérez, A.; LLorente-Cortes, V. Soluble LRP1 is an independent biomarker of epicardial fat volume in patients with type 1 diabetes mellitus. Sci. Rep. 2018, 8, 1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gonzalo-Calvo, D.; Vilades, D.; Nasarre, L.; Carreras, F.; Leta, R.; Garcia-Moll, X.; Llorente-Cortes, V. Circulating levels of soluble low-density lipoprotein receptor-related protein 1 (sLRP1) as novel biomarker of epicardial adipose tissue. Int. J. Cardiol. 2016, 223, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Von Arnim, C.A.; Kinoshita, A.; Peltan, I.D.; Tangredi, M.M.; Herl, L.; Lee, B.M.; Spoelgen, R.; Hshieh, T.T.; Ranganathan, S.; Battey, F.D.; et al. The low density lipoprotein receptor-related protein (LRP) is a novel β-secretase (BACE1) substrate. J. Biol. Chem. 2005, 280, 17777–17785. [Google Scholar] [CrossRef] [PubMed]
- Gorovoy, M.; Gaultier, A.; Campana, W.M.; Firestein, G.S.; Gonias, S.L. Inflammatory mediators promote production of shed LRP1/CD91, which regulates cell signaling and cytokine expression by macrophages. J. Leukoc. Biol. 2010, 88, 769–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gonzalo-Calvo, D.; Cenarro, A.; Martinez-Bujidos, M.; Badimon, L.; Bayes-Genis, A.; Ordonez-Llanos, J.; Civeira, F.; Llorente-Cortés, V. Circulating soluble low-density lipoprotein receptor-related protein 1 (sLRP1) concentration is associated with hypercholesterolemia: A new potential biomarker for atherosclerosis. Int. J. Cardiol. 2015, 201, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Shinohara, T.; Adachi, Y.; Asami, T.; Ohtaki, T. A novel LRP1-binding peptide L57 that crosses the blood brain barrier. Biochem. Biophys. Rep. 2017, 12, 135–139. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Actis Dato, V.; Chiabrando, G.A. The Role of Low-Density Lipoprotein Receptor-Related Protein 1 in Lipid Metabolism, Glucose Homeostasis and Inflammation. Int. J. Mol. Sci. 2018, 19, 1780. https://doi.org/10.3390/ijms19061780
Actis Dato V, Chiabrando GA. The Role of Low-Density Lipoprotein Receptor-Related Protein 1 in Lipid Metabolism, Glucose Homeostasis and Inflammation. International Journal of Molecular Sciences. 2018; 19(6):1780. https://doi.org/10.3390/ijms19061780
Chicago/Turabian StyleActis Dato, Virginia, and Gustavo Alberto Chiabrando. 2018. "The Role of Low-Density Lipoprotein Receptor-Related Protein 1 in Lipid Metabolism, Glucose Homeostasis and Inflammation" International Journal of Molecular Sciences 19, no. 6: 1780. https://doi.org/10.3390/ijms19061780