Signal Transduction in Plant–Nematode Interactions
by
 and
and
Muhammad Amjad Ali
1,2,*, 
Muhammad Shahzad Anjam
3, 
Muhammad Amjad Nawaz
4, 
Hon-Ming Lam
5 and
Gyuhwa Chung
4,* 1
Department of Plant Pathology, University of Agriculture, Faisalabad 38040, Pakistan
2
Centre of Agricultural Biochemistry and Biotechnology, University of Agriculture, Faisalabad 38040, Pakistan
3
Institute of Molecular Biology & Biotechnology, Bahauddin Zakariya University, Multan 66000, Pakistan
4
Department of Biotechnology, Chonnam National University, Yeosu 59626, Korea
5
School of Life Sciences and Centre for Soybean Research of the Partner State Key Laboratory of Agrobiotechnology, The Chinese University of Hong Kong, Shatin, Hong Kong SAR, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(6), 1648; https://doi.org/10.3390/ijms19061648
Submission received: 26 April 2018
/
Revised: 26 May 2018
/
Accepted: 29 May 2018
/
Published: 2 June 2018
(This article belongs to the Special Issue Signal Transduction Pathways in Plants for Resistance Against Plant Pathogens)
{kind=link}
{kind=link}
Abstract
:To successfully invade and infect their host plants, plant parasitic nematodes (PPNs) need to evolve molecular mechanisms to overcome the defense responses from the plants. Nematode-associated molecular patterns (NAMPs), including ascarosides and certain proteins, while instrumental in enabling the infection, can be perceived by the host plants, which then initiate a signaling cascade leading to the induction of basal defense responses. To combat host resistance, some nematodes can inject effectors into the cells of susceptible hosts to reprogram the basal resistance signaling and also modulate the hosts’ gene expression patterns to facilitate the establishment of nematode feeding sites (NFSs). In this review, we summarized all the known signaling pathways involved in plant–nematode interactions. Specifically, we placed particular focus on the effector proteins from PPNs that mimic the signaling of the defense responses in host plants. Furthermore, we gave an updated overview of the regulation by PPNs of different host defense pathways such as salicylic acid (SA)/jasmonic acid (JA), auxin, and cytokinin and reactive oxygen species (ROS) signaling to facilitate their parasitic successes in plants. This review will enhance the understanding of the molecular signaling pathways involved in both compatible and incompatible plant–nematode interactions.
1. Introduction
Parasitic nematodes are economically important pests of crop plants and can lead to significant yield losses worldwide [1]. Sedentary endoparasitic nematodes belonging to the family Heteroderidae include root knot nematodes from the genus Meloidogyne and cyst-forming nematodes from the genera Heterodera and Globodera [2]. These nematodes have developed very sophisticated mechanisms of parasitizing host plants, which involve the induction of specialized feeding sites in plant roots. These feeding structures are known as giant cells in the case of root knot nematodes and syncytia in cyst-forming nematodes, respectively [3]. Plant parasitic nematodes (PPNs) induce a series of morphological, biochemical, and molecular alterations in the cells of plant roots to facilitate the establishment of these feeding sites [4]. These events are largely affected by nematode secretions through their stylets, setting off a signaling cascade in the host cells.
The first signaling event in plant–nematode interactions involves the movement of second-stage juveniles (J2) towards the plant roots through the process of chemotaxis mediated by the root exudates of the host plants [5]. In the absence of resistance responses from the plant, e.g., production of reactive oxygen species (ROS) and callose deposition, PPNs enter the root cells and locate the vascular tissues for the initiation of feeding sites [6,7]. These nematode feeding sites (NFSs) become the sole nutrient sources for the nematodes as they develop through subsequent sedentary life stages [8,9].
The initiation of NFSs is brought about by nematode secretions that contain cell wall–degrading enzymes (CWDEs), virulence proteins (Avr proteins), and transcription factors [4]. The biochemical analyses of secretions from different PPNs demonstrated a variety of effector proteins that could interact with host genes and proteins to establish successful parasitism [4,10,11]. It has been suggested that the elements in the nematode stylet secretome led to significant changes in the host’s novel gene regulatory cascades, triggering the differentiation of the parasitized root cells into unique feeding cells [12]. Similarly, effectors secreted by the nematodes could reprogramme cellular events in the host plant for the induction of NFSs as well as suppress host defense responses by directly manipulating plant metabolic and developmental pathways [4,11].
PPNs can induce several signaling pathways through nematode-associated molecular patterns (NAMPs) to enable compatible interactions with the plants [13]. Similarly, secretions released by the nematodes can set off different signaling cascades which lead to the activation of plant genes helpful for the establishment of PPNs in plants. At the same time, as a counter measure, the detection of these NAMPs by the host plants also activate the basal immune system to safeguard the plants against nematode invasion [14]. It has also been proposed that as a one-upmanship, nematodes could produce effector proteins to induce changes in the plant’s cell cycle, cytoskeleton, and small RNA production to circumvent the host’s basal defense against parasitism [15,16,17,18]. Furthermore, PPNs are able to modulate different signaling pathways in their hosts, such as gene silencing pathways, jasmonic acid-salicylic acid (JA-SA) pathways, gibberellin (GA) pathways, cytokinin pathways, and post-transcriptional modifications [4,16,19,20,21].
In this review, we have focused on the most recent literature on plant signaling events involved in compatible and incompatible plant nematode interactions, starting from the initial nematode infection to the establishment of the feeding site in the plant root. This review will improve the understanding of various signal transduction pathways involved in plant–nematode molecular interactions by summarizing the most recent discoveries on these topics.
2. Cellular Signal Transduction in Plants
For millennia, plants and pathogens are continuously engaged in co-evolution in the struggle for dominance between hosts and pathogens. The invading pathogens face specific multi-layered defense responses generated by the host plant cells. The plant cell wall is the first physical barrier encountered by pathogens including plant parasitic nematodes (PPNs), fungi, bacteria, and viruses. Once the physical barrier is crossed by the pathogen, the host cytoplasm serves as the battleground where a war is fought between host and pathogen molecules. Next to the cell wall, there are membrane-localized pattern recognition receptors (PRRs) that recognize often-conserved pathogen/microbe-associated molecular patterns (PAMPs/MAMPs) such as proteins, lipids, carbohydrates, and cell wall derivatives [22]. As a consequence of the recognition of PAMPs/MAMPs, PRRs initiate a conserved downstream cellular signaling cascade referred to as PAMP-triggered immunity (PTI) inside the cytoplasm of the host cell [22,23]. The responses include the activation of mitogen-activated protein kinases (MAPKs), the production of reactive oxygen species (ROS), and the induction of signaling pathways via salicylic acid (SA) and jasmonic acid (JA) [24,25,26,27]. In plants, all the PRRs are transmembrane proteins containing a ligand-binding extracellular domain. The PRRs are divided into two groups: the receptor-like kinases (RLKs), which contain an intracellular kinase domain for cytoplasmic signaling, and the receptor-like proteins (RLPs), which lack any apparent cytoplasmic signaling domain [22,28]. It is assumed that RLPs always function in association with one or more RLKs for ligand-specific intracellular signal transduction [29]. PTI-triggered immunity usually protects plants against non-adapted microorganisms [30].
The pathogens that are successful in gaining access to the interior of the host cell release effector proteins to interfere with the host immune response (PTI) and to compromise the plant basal defense system. However, this triggers another mode of signal perception in the host by intracellular nucleotide binding/leucine-rich repeats (NLR) receptors [27,30], which induces a robust and strong response, often leading to localized cell death, thus preventing the spread of the disease beyond the initial site of infection. This defense mechanism is referred to as effector-triggered immunity (ETI). The response leading to cell death is generally referred to as hypersensitive response (HR), which also activates the long-lasting resistance in non-infected tissues as well (termed systemic acquired resistance) [22,27,30]. Nevertheless, plant basal immunity comprises two main responses: PTI and ETI [13,31,32]. These plant immune responses largely match the zig-zag model [32] in which molecular pattern release by PPNs could induce defense responses such as NTI (see Section 4), and nematode effector proteins may interact with host R-proteins to prompt ETI followed by localized HR (see Section 8 and Section 10). Plant immunity against PPNs could also be described on the basis of a multicomponent model [33] in which immunity activation component (IAC) is primarily comprised of NAMPs triggered signaling, followed by modification of gene expression of defense related genes by effector proteins (reviewed in [4]), and ultimately vertical immunity brought about by the interaction of R proteins and their corresponding Avr proteins. The immunity modulation component (IMC) involves the development of hyper-metabolic NFSs and modulation of various signaling cascades by different phytohormones [16].
3. Nematode-Associated Molecular Patterns (NAMPs) and Signaling for Nematode Resistance
Several studies have shown that plants generate PTI and ETI responses, such as the activation of mitogen-activated protein kinases (MAPKs), apoplastic ROS bursts, and JA and SA signaling, when they encounter nematode infections [34,35,36,37,38]. The first detailed study on NAMPs highlighted the perception of nematode-produced ascarosides by plant cells, triggering the gene expressions associated with MAMP-triggered immunity and the activation of MAPKs as well as JA and SA signaling [37]. Ascarosides are small molecules acting as pheromones in the social behavior of nematodes. In spite of diverse nematode phylogeny and ecology, ascaroside biosynthesis and signaling are highly conserved among nematodes [39]. In Caenorhabditis elegans (a free-living nematode) and many other species, ascarosides are involved in social signaling, dauer development, finding mating partners, and coordinating nematode behaviors [39,40,41]. Ascarosides are secreted by nematodes into their environment. Structurally, they are derivatives of the 3, 6-dideoxy-l-sugar, ascarylose, modified with fatty acid-derived side chains, and they are classified according to the number of carbons in their side chains [39,40,41]. Ascr#18 featuring an 11-carbon side chain is the most abundant among three genera of plant parasitic nematodes including root knot and cyst-forming nematodes. The priming of Arabidopsis roots with ascr#18 induced general resistance against bacteria, fungi and nematodes, as well as systemic resistance in the leaves of the plant [37,42]. All this evidence indicates that ascarosides secreted by nematodes are possible NAMPs perceivable by plants for the induction of basal defense mechanisms. However, evidence that ascarosides can be recognized by cell surface-localized receptors in plants is still lacking.
Recently another study claimed that the proteinaceous nature of potential NAMPs is a contributing factor in inducing basal immunity in Arabidopsis [43]. The authors prepared “NemaWater” by incubating pre-infective-stage juveniles of Heterodera schachtii and Meloidogyne incognita in water for 24 h. It was assumed that potential NAMPs present on the nematodes’ surface would be dissolved in the water during incubation. Then Arabidopsis roots were treated with NemaWater and produced apoplastic ROS bursts, which are hallmarks of plant basal immunity. Meanwhile, the Arabidopsis roots treated with NemaWater pre-incubated with proteinase K had significantly reduced ROS bursts. It was pointed out that one or more proteins of nematode origin could have been recognized by the plant cells for the activation of plant immunity. However, the question remains whether such proteins are secreted by pre-infective-stage juveniles on their surfaces. Furthermore, this study showed that the above-mentioned PTI responses were mediated through a membrane-localized leucine-rich repeat receptor-like kinase, NILR1, which was specifically induced upon nematode infection [43]. Therefore, it was the first-ever report on NILR1 as the PRR for perceiving a protein of nematode origin and inducing plant basal immunity against nematode infection. However, further studies are still needed for understanding the downstream signaling of NILR1.
Besides ascarosides and nematode-derived proteins, chitin and cuticle are also potential candidates for NAMPs. Chitin is present in the eggshells of various PPNs but absent in plant cells. Cuticle forms the exoskeleton of the nematode and is very important for its movement and growth [13]. However, the roles of chitin and cuticle have not yet been revealed in the induction of plant basal immunity against nematode infection. On the other hand, exudates released by plant roots are important for egg hatching, nematode chemotaxis towards plants, and trigger early events of signaling between plants and nematodes [44]. For instance, root exudates such as eclepins play pivotal roles in egg hatching of cyst nematodes while others could trigger the chemical mediated movement of nematode towards plant roots.
4. Phytoalexin Pathway and NAMP-Triggered Immunity (NTI)
PPNs have the ability to suppress important genes involved in the early events of defense responses in plants. Phytoalexins are important defense-related plant metabolites in the initial stages of plant–nematode interactions [45,46]. The biosynthesis of these metabolites is a key component of the innate immune system and involves a series of signaling events. The transcription factor, WRKY33, is considered to be the key player in the activation of camalexin-based PTI in plants [47,48]. This type of PTI comprises signal perception by transmembrane receptors followed by the commencement of signaling cascades through various MAPKs such as MPKs, MKKs, MKKKs, etc., involving several phosphorylation and dephosphorylation events [49]. These signaling cascades lead to the upregulation of the WRKY33 protein that in turn activates the PAD3 gene to induce camalexin production [47]. As supporting evidence, WRKY33 was the most downregulated WRKY transcription factors in the syncytia induced by H. schachtii compared to uninfected Arabidopsis roots [45,50]. MKK4 was shown to be the primary signaling kinase that activated WRKY33 and camalexin-mediated PTI. The overexpression of both WRKY33 and MKK4 resulted in enhanced resistance against H. schachtii in Arabidopsis while a T-DNA-insertion mutant of PAD3 led to increased susceptibility [45]. However, the phosphorylation and dephosphorylation events in this signaling cascade have not yet been worked out completely (Figure 1).
5. Nematode Effector Proteins
All PPNs contain a distinctive structure known as a stylet that is employed for puncturing plant cell walls, easing the movement of PPNs into the plant tissues, and for the subsequent injection of esophageal secretions into the host cells to induce NFSs [51]. However, a less significant amount of secretion is also released through the amphids and the cuticle of the nematode [4]. The stylet secretions contain a large number of effector and parasitism proteins which are developmentally controlled in different parasitic life stages of PPNs [51,52,53]. The secretions are collectively called the “secretome,” and the secreted proteins indirectly or directly involved in the development of compatible interactions with plant hosts are collectively known as the “parasitome” [2]. These are the secretions released by the nematode that facilitate the differentiation of host root cells into sophisticated and complex NFSs [12,51,54].
The secretomes of sedentary endoparasitic nematodes are particularly fascinating due to their ability to cause major alterations in host gene expressions. This altered expression pattern leads to metabolic, biochemical and morphological changes in infected root cells to turn these cells into unique NFSs [12]. The nematode secretome comprises highly concentrated proteins with huge diversity originating from divergent gland cell types [54,55]. The first complete parasitome profiling was carried out in the soybean cyst nematode (SCN), Heterodera glycines [56]. Similarly, the secretome of a root knot nematode, Meloidogyne incognita, consisted of 486 proteins [57]. Interestingly, a significant number of these secreted proteins from nematode secretions are homologues of plant proteins that could be stimulated by the nematode secreted proteins to induce plant growth and cell cycle reprogramming of gene expressions in the host cell [57].
The nematode parasitome primarily contains three categories of proteins [4]. The first category includes cell wall-degrading enzymes (CWDEs) and cell wall modification enzymes for localized cell wall degradation to facilitate the movement of PPNs through the plant root tissues. The second category of secreted proteins consists of nematode effectors that are involved in the modification of metabolic events and virulence factors that interact with resistance (R) proteins from the plant host, leading to enhanced resistance and/or susceptibility. The last category is the regulatory proteins that bind to the promoters of plant genes to modulate their expressions in favor of PPNs and the establishment of NFSs. These regulatory proteins are therefore transcription factors which act either as activators of plant genes helpful for the induction of NFSs, e.g., ATPases [58], or, more frequently the case, as suppressors of host genes involved in resistance responses, e.g., AtWRKYs and AtRAP2.6 [45,46].
The secreted proteins are either deposited outside the plasma membrane of the root cells or injected into the cytoplasm of the infected cells. In both situations, particular elicitors present in the nematode secretions could bind to plant cell receptors to induce a signal transduction cascade for the modification of host gene expressions [59,60,61].
6. CLE Signaling and Parasitic Success of Nematodes
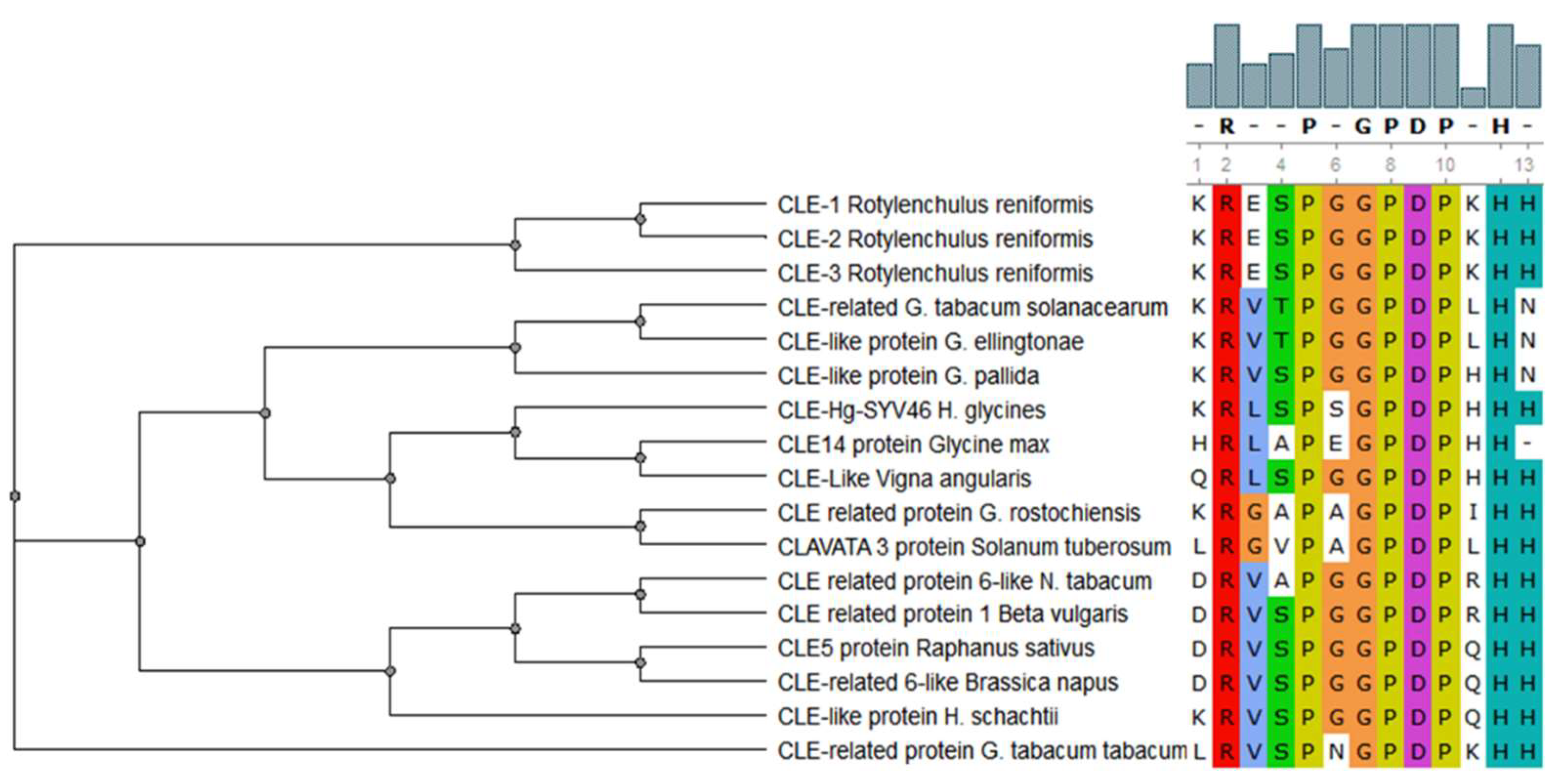
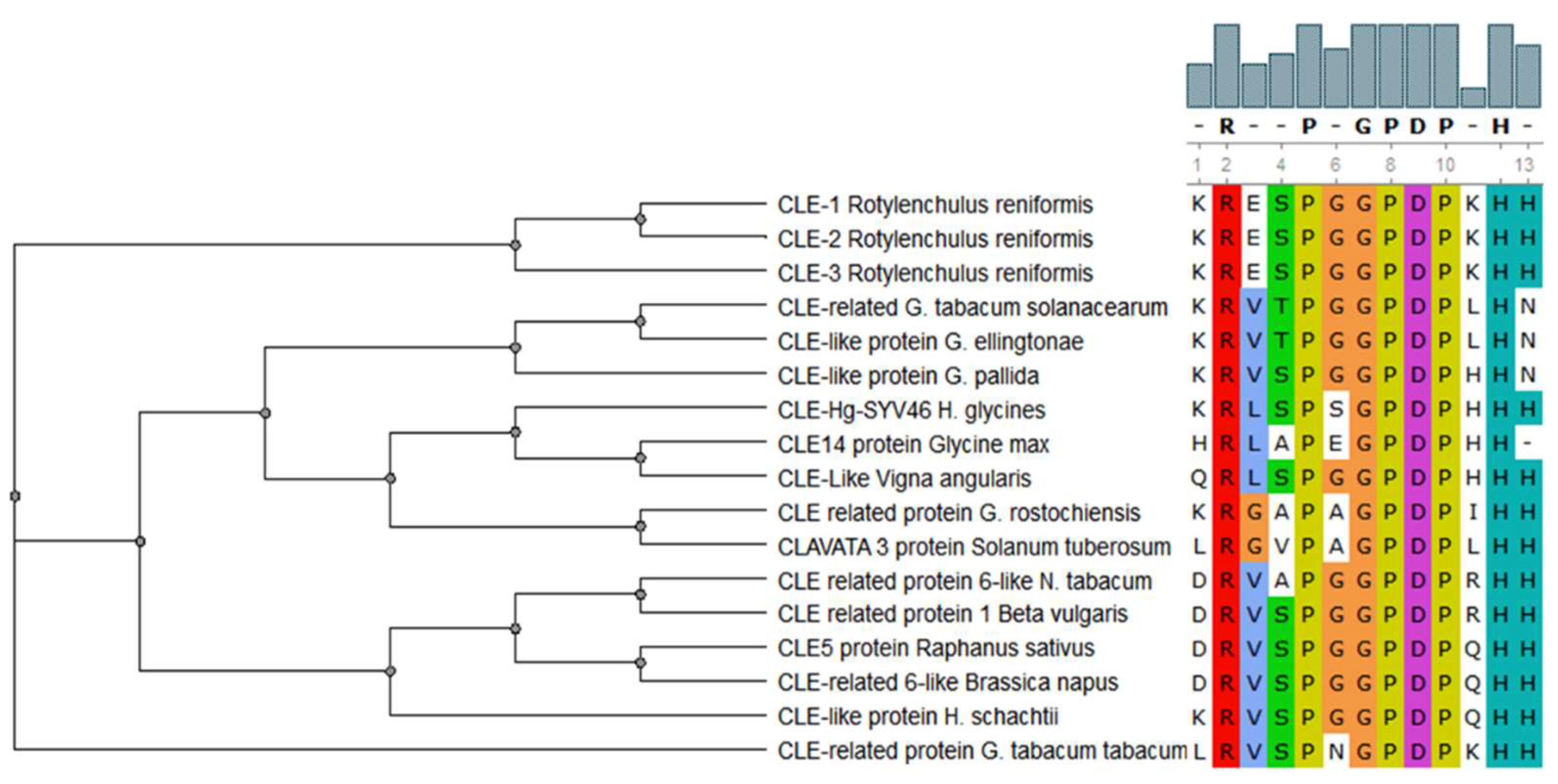
CLAVATA/ESR (CLE) peptides are involved in both the promotion and the inhibition of cell differentiation in meristematic tissues in plants, and members of this family of signaling peptides are also present in PPNs [61]. CLE peptides contain two highly conserved motifs: KRLVPSGPNPLHH and LxLxxxLILxLLLxS [62]. The CLE motifs from various PPNs are very similar to the CLE motifs found in their corresponding host plants [4] (Figure 2). This suggests that CLE peptides from PPNs are possible stimulators of cell differentiation and/or proliferation in host plants. This is why the establishment of syncytia and the differentiation of vascular tissues, e.g., xylem, so closely resemble each other [63]. There is a detailed review regarding the involvement of CLE signaling in plant development and plant–nematode interactions [64].
It has been demonstrated that receptor kinases like CORYNE (CRN) and CLAVATA2 (CLV2) are essential for CLE signaling for compatible plant–nematode interactions and the establishment of syncytia induced by H. schachtii in Arabidopsis [65]. The receptor kinases such as CLV1 and RECEPTOR-LIKE PROTEIN KINASE 2/TOADSTOOL2 (RPK2) are primarily involved in the maintenance of shoot apical meristem in Arabidopsis. These two receptors are also able to transmit the CLV3-independent signaling of CLV2/CRN upon CLE perception for the parasitic success of PPNs in plants [66]. Similarly, the CLE peptide from G. rostochiensis, GrCLE1, was glycosylated into a 12-amino acid arabinosylated glycopeptide resistant to hydrolytic degradation and with high-affinity binding to the potato CLV2-like receptor (StCLV2) [67]. This shows that the process of glycosylation is important in the CLE signaling process for compatible plant–nematode interactions. Moreover, the processing of GrCLE1 by host plant proteases enables this peptide to directly bind to the plant CLE receptors CLV2, BAM1, and BAM2, to modulate their functions [68]. CLE peptides also interact with plant genes to establish NFSs in roots [69]. A new class of CLE peptides known as B-type CLE peptides from Arabidopsis was found to involve in TDIF (tracheary element differentiation inhibitory factor)-TDR (TDIF receptor)-WOX4 pathway to induce the development of NFSs [70]. Primarily, this pathway is involved in proliferation of procambial meristematic cells. However, it was demonstrated that the TDIF pathway could be activated in syncytia induced by H. schachtii in Arabidopsis roots [70]. Loss-of-function mutants of various genes involved in this pathway (cle41, tdr-1, wox4-1, and double mutant tdr-1 wox4-1) exhibited lower number of nematodes along with compromised development of syncytia. These reports showed that CLE peptides are important in the developmental events in the meristem and are also involved in CLE signaling for the promotion of nematode parasitism in plants.
7. Nematode Effectors Are Targeted to Plant Cell Nuclei to Manipulate Host Functions
PPNs are able to alter various signaling pathways in their hosts with the help of effectors. For instance, 10A06 from H. schachtii modulates the biosynthesis of spermidine, which leads to enhanced antioxidant protection coupled with the interruption of salicylic acid (SA) signaling [71]. Some nematode effectors are localized in the plant cell nucleus and act as transcriptional activators or suppressors of plant genes. Recently, an M. incognita effector protein, Mi-EFF1, was found in the nuclei of the multinucleate NFSs and was postulated to be involved in the manipulation of gene expressions of host cells [72]. Nonetheless, the precise function of this effector is still unknown. Similar subcellular localization was observed for a novel M. javanica effector, Mj-NULG1a, which was targeted to the nuclei of giant cells to induce compatible interactions with the host [73]. Similarly, a M. incognita calreticulin, Mi-CRT, was found in the apoplasm of plant cells and was a key player in the suppression of basal defense of infected Arabidopsis plants [74,75,76]. This suppression in basal defense was demonstrated in the Mi-CRT overexpression lines by the downregulation of genes involved in inducing JA-SA signaling and callose production following the application of a PAMP, elf18 [76]. The homologous protein from B. xylophilus, Bx-CRT1, was found to be important for the differentiation of NFSs and cell-to-cell trafficking [77]. Similarly, the 10A07 effector from H. schachtii was found to recruit Interacting Plant Kinase (IPK) and the IAA16 transcription factor for its phosphorylation at serine-144 and 231 to mediate its trafficking from the cytoplasm to the nucleus [78].
The beet cyst nematode was shown to secret an ANNEXIN-like protein, Hs4F01, with high sequence similarity to plant annexins, and able to mimic plant annexins during the parasitic interaction, through binding phospholipids and calcium, which are important modulators of plant defense [79]. However, a recent report demonstrated that Ha-annexin from cereal cyst nematode H. avenae was targeted to the plant nucleus to suppress basal defense responses of the host [80]. Stable silencing of this gene led to abnormal nematode development on wheat roots, whereas transient expression of Ha-annexin resulted in the suppression of programmed cell death (PCD) induced by different PAMPs and the BAX protein [80]. Similarly, silencing and overexpression of MiMsp40 in Arabidopsis led to the suppression and establishment of parasitism by M. incognita, respectively, through the respective enhancement and suppression of PAMP-triggered immunity and callose deposition [81]. In this case, both PTI and ETI were reduced, through the interactions between BAX and cognate elicitors R3a/Avr3a to inhibit HR and facilitate nematode establishment in the plant root [81].
Among different types of effector proteins released by the nematodes, transcription factors are very important players involved in modulation of host gene expression [4]. A total of 46 differential transcription factor binding sites (TFBSs) in soybean lines with contrasting resistance against soybean cyst nematode, H. glycines were identified [82]. HAHB4, DOF, MYB, and ARF boxes are some of the important examples of these differentially represented TFBSs [82]. The authors proposed that these over–represented TFBSs in soybean could play important roles in cis-regulatory dynamics.
8. Nematode Effectors Mimic Defense Responses in Host Plants
At the same time that effector proteins originating from PPNs are mimicking various plant proteins and suppressing plant defense responses, their perception by the host plant would induce host defense against their invasion. The very first step of these responses is the recognition of nematode effectors (Avr proteins) by resistance (R) proteins from the host plant, leading to the activation of ETI [4]. PPNs contain numerous Avr proteins that result in gene-for-gene interactions and play a central role in incompatible plant–nematode interactions. Important examples of these Avr proteins are SPRYSEC-19 [83,84], map-1.2 [85], chorismate mutase from H. glycines (Hg-cm-1) [85,86,87], Cg-1 from M. incognita [88], and Gr-VAP1 [89] and Gp-Rbp-1 [90,91] from pale and white potato cyst nematodes, respectively.
Cg-1 and map-1.2 proteins interact with the Mi-1 R protein, a member of CC-NBS-LRR (CNL) class of R-proteins, to induce Mi-1-mediated ETI in tomato plants [85,88,92]. Experimental evidence shows that SPRYSEC-19 from G. rostochiensis is recognized by the leucine-rich repeat (LRR) motif of the tomato CNL family R protein, SW5-F [83]. However, surprisingly, SPRYSEC-19 did not induce PCD in tobacco leaves when transiently co-expressed with the SW5-F protein [84]. This suggests that SPRYSEC-19 somehow suppresses CNL-mediated resistance responses in plants. Conversely, a SPRYSEC gene from G. pallida (Gp-Rbp-1) led to the activation of HR after it was co-expressed with the potato resistance genes, Gpa2 and RanGAP2 (Ran GTPase Activating Protein 2) [90,91]. The variability in the amino acid sequence of potato RanGAP2 is a main contributing factor for the recognition of Avr proteins by Gpa2 to enhance nematode resistance in potato [93].
The venom allergen-like proteins (Vap) are also important PPN components recognized by host plants to mount a defense response against the nematode infection [89,94,95,96]. A strong interaction was demonstrated between the G. rostochiensis Vap protein (Gr-Vap1) and the tomato cysteine protease, Rcr3pim, and immune receptor protein, Cf-2, to induce PCD and enhanced resistance against the infection by G. rostochiensis [89,97]. It seems that both of these genes in tomato are involved in signal perception and interaction with unknown R proteins to confer resistance responses. As Gr-VAP1 has also been shown to activate basal innate immunity in the potato hairy roots following the infection with G. rostochiensis, it may be regarded as a PAMP to elicit PTI as well. Another effector protein from G. rostochiensis, GrEXPB2, can both suppress and elicit defense responses in the plant. On one hand, this effector is involved in the suppression of PCD induced by a Phytophthora infestans extracellular elicitor, PiNPP, and NB-LRR mediated immune responses by the inhibition of an immune receptor of Rx protein in tobacco leaves [98]. On the other hand, GrEXPB2 was also found to elicit defense responses in a number of potato and tomato lines [98].
It has been reported that the nematode effector protein, Mj-FAR-1, from M. javanica could be involved in the manipulation of lipid-based molecular signaling and inhibition of JA-mediated signaling in plant–nematode interactions [99]. Likewise, similar studies have also demonstrated the participation of different effector proteins in both the suppression and activation of basal immune responses of the host plants [100]. Overall, the majority of reported effectors function by manipulating the hosts’ nuclear functions to develop NFSs [4].
In addition to well characterized R genes, a few quantitative trait loci (QTLs) regulating nematode resistance were also reported. Among them, Cre and Rhg are well known examples in wheat and soybean conferring resistance against cereal cyst nematodes and soybean cyst nematodes (SCN) respectively. Rhg1 and Rhg4 are naturally occurring QTLs identified in soybean that control resistance responses against SCN [35,101]. Experimental evidence showed that Rhg4 encodes a serine hydroxymethyltransferase and overexpression of this enzyme in soybean roots led to 45% reduction in the number of mature cyst nematodes [102]. Genome-wide association study (GWAS) could also be an effective way to identity resistance QTLs. For instance, 13 single nucleotide polymorphisms (SNPs) associated with soybean cyst nematode resistance in soybean were identified, in which 3 SNPs were linked to the previously reported QTLs Rhg1 and Rhg4 [103].
9. Roles of SA/JA, Auxin, and Cytokinin Signaling in Plant–Nematode Interactions
Plant hormones such as auxins and cytokinins are important in plant–nematode interactions because they are involved in the establishment of NFSs in plant roots [21,104,105]. SA and JA are key players involved in the activation of different signaling pathways in defense responses in plants [19,20,34]. Callose production is one of the earliest defense responses of plants against the invading PPNs [9]. Overexpression of an Arabidopsis ethylene-responsive transcription factor, RAP2.6, led to enhanced production of JA and callose in NFSs as compared to control plant roots. However, resistance in tomato against M. incognita is primarily associated with SA signaling instead of the typical JA signaling [19,20]. In the same way, HeroA-based resistance responses in potato involves elevated expressions of SA-induced PR genes following the infection by potato cyst nematode, while in susceptible potato lines, the SA signaling pathway was inhibited [106]. Comparable results were found in a resistant wheat line expressing the Cre2 gene against cereal cyst nematode, H. avenae [107].
Nematode effector proteins can modulate auxin-based signaling and defense responses. For instance, the H. schachtii effector, 19C07, can bind the Arabidopsis auxin transporter, LAX3, to control auxin influx and the formation of syncytia [108]. Likewise, a chorismate mutase-coding gene from M. javanica, MjCM-1, regulates auxin signaling through altering the levels of indole-3-acetic acid (IAA) in host cells, which leads to alterations in chorismate metabolism as well as chorismate-derived metabolites to promote cell enlargement and the induction of NFSs [109]. Moreover, the upregulation of auxin response transcription factors (ARFs) in NFSs suggests the putative role of auxin signaling in compatible plant–nematode interactions [110]. By studying the transcriptome of H. glycines-resistant soybean lines, it has been demonstrated that auxin-induced biosynthesis of ROS is associated with increased Ca2+ conductance across the cell membrane in the resistant lines. Furthermore, DELLA-like proteins are probably involved in auxin-regulated ROS signaling to produce resistance responses in soybean [111].
Cytokinins are required for the successful development of syncytia in Arabidopsis. This was discovered by infecting cytokinin-deficient Arabidopsis mutants with H. schachtii. The results revealed decreased susceptibility of cytokinin-deficient mutants against the PPN [112]. It was also reported that isopentenyl transferase, the key player involved in the biosynthesis of cytokinin, could modulate nematode virulence, the division of host cells, and the establishment of NFSs in Arabidopsis roots [112].
Recently, the expressions of various genes involved in the defense mechanisms in resistant versus susceptible rice cultivars were compared after infection with M. graminicola [113]. The expressions of several genes involved in lignin biosynthesis and callose deposition were enhanced in the resistant cultivar compared to the susceptible one. Moreover, MAPKs such as OsMAPK5, OsMAPK6 and OsMAPK20 were activated in the early stages of M. graminicola infection in a nematode-resistant rice cultivar [113]. Furthermore, SA-JA as well as ethylene-related genes in the resistant line were also activated, suggesting the activation of various defense mechanisms involving these pathways in rice. Similar results were shown in wild soybean–H. glycines interactions in which several MAPKs and SA-JA signaling genes were upregulated during resistance responses [114]. More recently, two transcription factors, DAF-16 and SKN-1 from various free-living and parasitic nematodes, were identified as key players involved in important signaling pathways including SA, JA, MTI, and ROS biosynthesis [115].
10. Parasitic Nematodes Modulate ROS Signaling and PCD in Plants
Programmed cell death (PCD) and hypersensitive response (HR) are the main results of ROS signaling in plants. Plants are able to program the suicidal death of their cells upon pathogen invasion through the biosynthesis of ROS. This leads to HR that restricts the pathogen’s spread through living tissue and kills the pathogen at the site of infection. However, the biosynthesis of ROS is largely reliant on the accumulation of SA at the site of infection [116,117]. It has been reported that the production of ROS is one of the earliest events in molecular plant–nematode interactions [9]. R proteins frequently lead to the upregulation of genes involved in ROS production which results in HR. Soybean lines harboring the Rhg1 gene displayed elevated expressions of ROS-related genes and enhanced resistance to soybean cyst nematode, H. glycines [35]. Moreover, DELLA-like proteins were found to be involved in auxin-induced signaling for the biosynthesis of ROS in soybean lines resistant to H. glycines [111].
As mentioned earlier, higher concentrations of SA stimulate the biosynthesis of ROS and result in PCD in plants [118,119]. However, the SA-dependent ROS biosynthesis pathway and the ROS biosynthesis pathway mediated by Respiratory Burst Oxidase Homologs (RBOHs) are mutually antagonistic in Arabidopsis [120]. It has been reported that PPNs can manipulate the RBOH-dependent ROS pathway for parasitic success in plants. This study demonstrated that H. schachtii was able to stimulate the expression of RBohD and RBohF in Arabidopsis to induce the synthesis of ROS that surprisingly resulted in the inhibition of PCD and therefore supported the development of syncytia [36].
Plants have a variety of enzymes (e.g., peroxidases) to scavenge excessive ROS in the plant tissue to prevent self-damage. In the same way, secretomes of PPNs contain superoxide dismutases (SODs) to detoxify ROS produced by the plant cells and dampen resistance responses [57,121]. Similarly, PPNs can scavenge the excess ROS with enzymes and antioxidants, such as glutathione, peroxidases, thioredoxins, cytochrome C-peroxidases, ascorbate peroxidases, and catalases, which protect the nematode cells from ROS-mediated injuries [57,122]. Additionally, the overexpression of a peroxiredoxin-family gene from potato cyst nematode, G. rostochiensis, protected it against defense responses from the potato plant through an array of redox reactions [123].
11. Nematodes Modulate RNA Silencing Pathways to Promote Infection Process
RNA silencing is an important strategy of plant responses against abiotic and biotic stresses, in which microRNAs (miRNAs) are important modulators [124]. Using deep sequencing to compare miRNA profiles, 60 differentially expressed miRNA from 25 diverse miRNA families associated with responses against SCN were identified [125]. Moreover, an Arabidopsis microRNA, mi396, was reported to interact with transcription factors like Growth-Regulating Factor, GRF1/GRF3 to modulate the reprogramming of root development under H. schachtii infection [126]. The same technique was used to analyze miRNAs in soybean after infection with SCN, which led to the identification of 20 miRNAs with diverse expression pattern in resistant and susceptible soybean cultivars [127]. It has been shown recently that the majority of these miRNAs were predominantly upregulated in susceptible cultivars as compared to the resistant counterparts [128]. The expression of these upregulated miRNAs (i.e., miR156, miR159, miR164, and miR396) was found to be negatively correlated with the expression of their target transcription factors such as SBP, GAMYB-like, NAC, and GRF1, respectively. These studies suggest a pivotal role of miRNAs in the regulation of regulatory transcription factors responsible for plant resistance responses.
12. Conclusions and Future Perspectives
PPNs are able to induce and modulate different signaling pathways in plants with the help of their effector proteins. These effectors can not only induce the auxin and cytokinin signaling for the development of NFSs but can also suppress the SA and JA signaling to avoid host defense responses. PPNs release NAMPs that lead to the signaling for camalexin-based NTI in plants. Similarly, PPNs could regulate different defense pathways, e.g., ROS production, to their own advantage to establish compatible interactions with the plant hosts. In-planta RNA silencing of genes from PPNs as well as those of host plants involved in various signaling pathways could be helpful for understanding molecular signaling in plant–nematode interactions [129]. The use of recently developed transcriptomic technologies such as high-throughput RNA-seq will be helpful for the identification of interacting genes in various signaling pathways related to plant defense mechanisms [114,130]. The secretomes of PPNs and transcriptomes of numerous host plants reveal a significant number of differentially regulated plant genes [4], which could be potential candidates for understanding the site-specific silencing in syncytia to enhance nematode resistance in plants [131].
Emerging molecular technologies such as deep sequencing could be of potential use for high throughput studies to provide a holistic view of signaling events involved in the regulation of plant nematode interactions [114]. Gland cell RNA sequencing could be exploited to understand the functioning of various effectors associated with signaling process [132]. Similarly, genome editing approaches like CRISPR-Cas9 could be employed for exploring the function of various plant and nematode genes involved in compatible and incompatible plant–nematode interactions. These genome editing technologies (GETs) could be helpful in understanding the interaction of nematode effector proteins with R proteins from plants [133].
Acknowledgments
This work was supported by grants from the Hong Kong Research Grants Council Area of Excellence Scheme (AoE/M-403/16), CUHK VC Discretionary Fund VCF2014004, and the Lo Kwee-Seong Biomedical Research Fund to H.-M.L. Jee Yan Chu copy-edited the manuscript. MAA is highly thankful to Higher Education Commission of Pakistan for support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Abbas, A.; Azeem, F.; Javed, N.; Bohlmann, H. Plant–nematode Interactions: From Genomics to Metabolomics. Int. J. Agric. Biol. 2015, 17, 1071–1082. [Google Scholar]
- Ali, M.A.; Azeem, F.; Abbas, A.; Joyia, F.A.; Li, H.; Dababat, A. Transgenic strategies for enhancement of nematode resistance in plants. Front. Plant Sci. 2017, 8, 750. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Azeem, F.; Li, H.; Bohlmann, H. Smart Parasitic Nematodes Use Multifaceted Strategies to Parasitize Plants. Front. Plant Sci. 2017, 8, 1699. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The Good, the Bad, and the Ugly of Rhizosphere Microbiome. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Wyss, U.; Grundler, F.M.W. Feeding-Behavior of Sedentary Plant Parasitic Nematodes. Neth. J. Plant. Pathol. 1992, 98, 165–173. [Google Scholar] [CrossRef]
- Golinowski, W.; Grundler, F.M.W.; Sobczak, M. Changes in the structure of Arabidopsis thaliana during female development of the plant-parasitic nematode Heterodera schachtii. Protoplasma 1996, 194, 103–116. [Google Scholar] [CrossRef]
- Endo, B.Y.; Wyss, U. Ultrastructure of Cuticular Exudations in Parasitic Juvenile Heterodera-Schachtii, as Related to Cuticle Structure. Protoplasma 1992, 166, 67–77. [Google Scholar] [CrossRef]
- Grundler, F.M.W.; Sobczak, M.; Lange, S. Defence responses of Arabidopsis thaliana during invasion and feeding site induction by the plant-parasitic nematode Heterodera glycines. Physiol. Mol. Plant Pathol. 1997, 50, 419–429. [Google Scholar] [CrossRef]
- Bellafiore, S.; Briggs, S.P. Nematode effectors and plant responses to infection. Curr. Opin. Plant Biol. 2010, 13, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Gheysen, G.; Mitchum, M.G. How nematodes manipulate plant development pathways for infection. Curr. Opin. Plant Biol. 2011, 14, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Gheysen, G.; Fenoll, C. Gene expression in nematode feeding sites. Ann. Rev. Phytopathol. 2002, 40, 191–219. [Google Scholar] [CrossRef] [PubMed]
- Holbein, J.; Grundler, F.M.W.; Siddique, S. Plant basal resistance to nematodes: An update. J. Exp. Bot. 2016, 67, 2049–2061. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, M.C.; Dubreuil, G.; Quentin, M.; Perfus-Barbeoch, L.; Lecomte, P.; de Almeida Engler, J.; Abad, P.; Rosso, M.N.; Favery, B. Root-knot nematodes manipulate plant cell functions during a compatible interaction. J. Plant Physiol. 2008, 165, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T. Cellular Signaling Pathways and Posttranslational Modifications Mediated by Nematode Effector Proteins. Plant Physiol. 2015, 169, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Baum, T. Manipulation of Plant Cells by Cyst and Root-Knot Nematode Effectors. Mol. Plant Microbe Interact 2012, 26, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Howe, P.; Maier, T.R.; Baum, T.J. Arabidopsis Small RNAs and Their Targets During Cyst Nematode Parasitism. Mol. Plant Microbe Interact 2008, 21, 1622–1634. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.K.; Xie, Q.G.; Mantelin, S.; Bishnoi, U.; Girke, T.; Navarre, D.A.; Kaloshian, I. Tomato susceptibility to root-knot nematodes requires an intact jasmonic acid signaling pathway. Mol. Plant Microbe Interact 2008, 21, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Branch, C.; Hwang, C.F.; Navarre, D.A.; Williamson, V.M. Salicylic acid is part of the Mi-1-mediated defense response to root-knot nematode in tomato. Mol. Plant Microbe Interact 2004, 17, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Absmanner, B.; Stadler, R.; Hammes, U.Z. Phloem development in nematode-induced feeding sites: The implications of auxin and cytokinin. Front. Plant Sci. 2013, 4, 241. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Kmiecik, P.; Willems, P.; Van Der Kelen, K.; Coll, N.S.; Teige, M.; Van Breusegem, F. Plant innate immunity—Sunny side up? Trends Plant Sci. 2015, 20, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More Than Just jasmonate-salicylate Antagonism. Ann. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Ann. Rev. Cell Developmental Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zipfel, C. Plant PRRs and the Activation of Innate Immune Signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-Triggered Immunity: From Pathogen Perception to Robust Defense. Ann. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, H.; Li, X.; Li, Y.; Cui, H.; Wen, C.-K.; Tang, X.; Su, Z.; Zhou, J.-M. Effector-Triggered and Pathogen-Associated Molecular Pattern–Triggered Immunity Differentially Contribute to Basal Resistance to Pseudomonas syringae. Mol. Plant Microbe Interact 2010, 23, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfo, G.; Ercolano, M.R. Plant Innate Immunity Multicomponent Model. Front. Plant Sci. 2015, 6, 987. [Google Scholar] [CrossRef] [PubMed]
- Hamamouch, N.; Li, C.Y.; Seo, P.J.; Park, C.M.; Davis, E.L. Expression of Arabidopsis pathogenesis-related genes during nematode infection. Mol. Plant Pathol. 2011, 12, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Kandoth, P.K.; Ithal, N.; Recknor, J.; Maier, T.; Nettleton, D.; Baum, T.J.; Mitchum, M.G. The Soybean Rhg1 Locus for Resistance to the Soybean Cyst Nematode Heterodera glycines Regulates the Expression of a Large Number of Stress- and Defense-Related Genes in Degenerating Feeding Cells. Plant Physiol. 2011, 155, 1960–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, S.; Matera, C.; Radakovic, Z.S.; Shamim Hasan, M.; Gutbrod, P.; Rozanska, E.; Sobczak, M.; Angel Torres, M.; Grundler, F.M. Parasitic worms stimulate host NADPH oxidases to produce reactive oxygen species that limit plant cell death and promote infection. Sci. Signal 2014, 7, ra33. [Google Scholar] [CrossRef] [PubMed]
- Manosalva, P.; Manohar, M.; von Reuss, S.H.; Chen, S.; Koch, A.; Kaplan, F.; Choe, A.; Micikas, R.J.; Wang, X.; Kogel, K.-H.; et al. Conserved nematode signalling molecules elicit plant defenses and pathogen resistance. Nat. Commun. 2015, 6, 7795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidonskaya, E.; Schweighofer, A.; Shubchynskyy, V.; Kammerhofer, N.; Hofmann, J.; Wieczorek, K.; Meskiene, I. Plant resistance against the parasitic nematodeHeterodera schachtiiis mediated by MPK3 and MPK6 kinases, which are controlled by the MAPK phosphatase AP2C1 in Arabidopsis. J. Exp. Bot. 2016, 67, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Choe, A.; von Reuss, S.H.; Kogan, D.; Gasser, R.B.; Platzer, E.G.; Schroeder, F.C.; Sternberg, P.W. Ascaroside Signaling Is Widely Conserved among Nematodes. Curr. Biol. 2012, 22, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Panda, O.; Akagi, A.E.; Artyukhin, A.B.; Judkins, J.C.; Le, H.H.; Mahanti, P.; Cohen, S.M.; Sternberg, P.W.; Schroeder, F.C. Biosynthesis of Modular Ascarosides in C. elegans. Angew. Chem. Int. Ed. 2017, 56, 4729–4733. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Srinivasan, J.; Mahanti, P.; Ajredini, R.; Durak, O.; Nimalendran, R.; Sternberg, P.W.; Teal, P.E.A.; Schroeder, F.C.; Edison, A.S.; et al. Ascaroside Expression in Caenorhabditis elegans Is Strongly Dependent on Diet and Developmental Stage. PLoS ONE 2011, 6, e17804. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, X.; Wei, Y.; Zhou, J.; Zhang, W.; Qin, P.; Chinta, S.; Kong, X.; Liu, Y.; Yu, H.; et al. Ascarosides coordinate the dispersal of a plant-parasitic nematode with the metamorphosis of its vector beetle. Nat. Commun. 2016, 7, 12341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendy, B.; Wang’ombe, M.W.; Radakovic, Z.S.; Holbein, J.; Ilyas, M.; Chopra, D.; Holton, N.; Zipfel, C.; Grundler, F.M.W.; Siddique, S. Arabidopsis leucine-rich repeat receptor–like kinase NILR1 is required for induction of innate immunity to parasitic nematodes. PLoS Pathog. 2017, 13, e1006284. [Google Scholar] [CrossRef] [PubMed]
- Rasmann, S.; Ali, J.G.; Helder, J.; van der Putten, W.H. Ecology and Evolution of Soil Nematode Chemotaxis. J. Chem. Ecol. 2012, 38, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Wieczorek, K.; Kreil, D.P.; Bohlmann, H. The beet cyst nematode Heterodera schachtii modulates the expression of WRKY transcription factors in syncytia to favour its development in Arabidopsis roots. PLoS ONE 2014, 9, e102360. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Abbas, A.; Kreil, D.P.; Bohlmann, H. Overexpression of the transcription factor RAP2.6 leads to enhanced callose deposition in syncytia and enhanced resistance against the beet cyst nematode Heterodera schachtii in Arabidopsis roots. BMC Plant Biol. 2013, 13, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin biosynthesis in Arabidopsis. Plant Cell 2011, 23, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.; Fiil, B.K.; Mundy, J.; Petersen, M. Downstream targets of WRKY33. Plant Signal Behav. 2008, 3, 1033–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Szakasits, D.; Heinen, P.; Wieczorek, K.; Hofmann, J.; Wagner, F.; Kreil, D.P.; Sykacek, P.; Grundler, F.M.; Bohlmann, H. The transcriptome of syncytia induced by the cyst nematode Heterodera schachtii in Arabidopsis roots. Plant J. 2009, 57, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.L.; Hussey, R.S.; Baum, T.J.; Bakker, J.; Schots, A. Nematode parasitism genes. Ann. Rev. Phytopathol. 2000, 38, 365–396. [Google Scholar] [CrossRef] [PubMed]
- Hussey, R.S. Disease-Inducing Secretions of Plant-Parasitic Nematodes. Ann. Rev. Phytopathol. 1989, 27, 123–141. [Google Scholar] [CrossRef]
- Hussey, R.S.; Davis, E.L.; Baum, T.J. Secrets in secretions: Genes that control nematode parasitism of plants. Braz. J. Plant Physiol. 2002, 14, 183–194. [Google Scholar] [CrossRef]
- Burgess, T.L.; Kelly, R.B. Constitutive and Regulated Secretion of Proteins. Ann. Rev. Cell Biol. 1987, 3, 243–293. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, B.; De Meutter, J.; Tytgat, T.; Van Montagu, M.; Coomans, A.; Gheysen, G. Secretions of plant-parasitic nematodes: A molecular update. Gene 2004, 332, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.L.; Allen, R.; Maier, T.; Davis, E.L.; Baum, T.J.; Hussey, R.S. The parasitome of the phytonematode Heterodera glycines. Mol. Plant Microbe Interact 2003, 16, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Bellafiore, S.; Shen, Z.X.; Rosso, M.N.; Abad, P.; Shih, P.; Briggs, S.P. Direct Identification of the Meloidogyne incognita Secretome Reveals Proteins with Host Cell Reprogramming Potential. PLoS Pathog. 2008, 4, e1000192. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Plattner, S.; Radakovic, Z.; Wieczorek, K.; Elashry, A.; Grundler, F.M.; Ammelburg, M.; Siddique, S.; Bohlmann, H. An Arabidopsis ATPase gene involved in nematode-induced syncytium development and abiotic stress responses. Plant J. 2013, 74, 852–866. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.L.; Hussey, R.S.; Baum, T.J. Getting to the roots of parasitism by nematodes. Trends Parasitol. 2004, 20, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.L.; Mitchum, M.G. Nematodes. Sophisticated parasites of legumes. Plant Physiol. 2005, 137, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Wang, X.H.; Davis, E.L. Diverse and conserved roles of CLE peptides. Curr. Opin. Plant Biol. 2008, 11, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Skriver, K. Ligand mimicry? Plant-parasitic nematode polypeptide with similarity to CLAVATA3. Trends Plant Sci. 2003, 8, 55–57. [Google Scholar] [CrossRef]
- Fisher, K.; Turner, S. PXY, a receptor-like kinase essential for maintaining polarity during plant vascular-tissue development. Curr. Biol. 2007, 17, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Fiers, M. CLE peptide signaling during plant development. Protoplasma 2009, 240, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Replogle, A.; Wang, J.Y.; Bleckmann, A.; Hussey, R.S.; Baum, T.J.; Sawa, S.; Davis, E.L.; Wang, X.H.; Simon, R.; Mitchum, M.G. Nematode CLE signaling in Arabidopsis requires CLAVATA2 and CORYNE. Plant J. 2011, 65, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Replogle, A.; Wang, J.; Paolillo, V.; Smeda, J.; Kinoshita, A.; Durbak, A.; Tax, F.E.; Wang, X.; Sawa, S.; Mitchum, M.G. Synergistic interaction of CLAVATA1, CLAVATA2, and RECEPTOR-LIKE PROTEIN KINASE 2 in cyst nematode parasitism of Arabidopsis. Mol. Plant Microbe Interact 2013, 26, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lang, P.; Chronis, D.; Zhang, S.; De Jong, W.S.; Mitchum, M.G.; Wang, X. In planta processing and glycosylation of a nematode CLAVATA3/ENDOSPERM SURROUNDING REGION-like effector and its interaction with a host CLAVATA2-like receptor to promote parasitism. Plant Physiol. 2015, 167, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.F.; Ni, J.; Denver, R.; Wang, X.H.; Clark, S.E. Mechanisms of Molecular Mimicry of Plant CLE Peptide Ligands by the Parasitic Nematode Globodera rostochiensis. Plant Physiol. 2011, 157, 476–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyohara, S.; Sawa, S. CLE signaling systems during plant development and nematode infection. Plant Cell Physiol. 2012, 53, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, J.; Gardner, M.; Fukuda, H.; Kondo, Y.; Etchells, J.P.; Wang, X.; Mitchum, M.G. Identification of cyst nematode B-type CLE peptides and modulation of the vascular stem cell pathway for feeding cell formation. PLoS Pathog. 2017, 13, e1006142. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Howe, P.J.; Maier, T.R.; Hussey, R.S.; Mitchum, M.G.; Davis, E.L.; Baum, T.J. Arabidopsis Spermidine Synthase Is Targeted by an Effector Protein of the Cyst Nematode Heterodera schachtii. Plant Physiol. 2010, 152, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Jaouannet, M.; Perfus-Barbeoch, L.; Deleury, E.; Magliano, M.; Engler, G.; Vieira, P.; Danchin, E.G.; Da Rocha, M.; Coquillard, P.; Abad, P.; et al. A root-knot nematode-secreted protein is injected into giant cells and targeted to the nuclei. New Phytol. 2012, 194, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Zhuo, K.; Wu, P.; Cui, R.; Zhang, L.H.; Liao, J. A novel effector protein, MJ-NULG1a, targeted to giant cell nuclei plays a role in Meloidogyne javanica parasitism. Mol. Plant Microbe Interact 2013, 26, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Jaubert, S.; Ledger, T.N.; Laffaire, J.B.; Piotte, C.; Abad, P.; Rosso, M.N. Direct identification of stylet secreted proteins from root-knot nematodes by a proteomic approach. Mol. Biochem. Parasitol. 2002, 121, 205–211. [Google Scholar] [CrossRef]
- Jaubert, S.; Milac, A.L.; Petrescu, A.J.; de Almeida-Engler, J.; Abad, P.; Rosso, M.N. In planta secretion of a calreticulin by migratory and sedentary stages of root-knot nematode. Mol. Plant Microbe Interact 2005, 18, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Jaouannet, M.; Magliano, M.; Arguel, M.J.; Gourgues, M.; Evangelisti, E.; Abad, P.; Rosso, M.N. The root-knot nematode calreticulin Mi-CRT is a key effector in plant defense suppression. Mol. Plant Microbe Interact 2013, 26, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Zhuo, K.; Luo, M.; Sun, L.H.; Liao, J.L. Molecular cloning and characterization of a calreticulin cDNA from the pinewood nematode Bursaphelenchus xylophilus. Exp. Parasitol. 2011, 128, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Juvale, P.S.; Piya, S.; Maier, T.R.; Rambani, A.; Rice, J.H.; Mitchum, M.G.; Davis, E.L.; Hussey, R.S.; Baum, T.J. The Cyst Nematode Effector Protein 10A07 Targets and Recruits Host Posttranslational Machinery to Mediate Its Nuclear Trafficking and to Promote Parasitism in Arabidopsis. Plant Cell 2015, 27, 891–907. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Hamamouch, N.; Li, C.Y.; Hewezi, T.; Hussey, R.S.; Baum, T.J.; Mitchum, M.G.; Davis, E.L. A nematode effector protein similar to annexins in host plants. J. Exp. Bot. 2010, 61, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, S.; Liu, Q.; Niu, J.; Liu, P.; Zhao, J.; Jian, H. An ANNEXIN-Like Protein from the Cereal Cyst Nematode Heterodera avenae Suppresses Plant Defense. PLoS ONE 2015, 10, e0122256. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Liu, P.; Liu, Q.; Chen, C.; Guo, Q.; Yin, J.; Yang, G.; Jian, H. Msp40 effector of root-knot nematode manipulates plant immunity to facilitate parasitism. Sci. Rep. 2016, 6, 19443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, P.; Matthews, B.F. Regulatory interplay between soybean root and soybean cyst nematode during a resistant and susceptible reaction. BMC Plant BIol. 2014, 14, 300. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Postma, W.; Tytgat, T.; Prins, P.; Qin, L.; Overmars, H.; Vossen, J.; Spiridon, L.N.; Petrescu, A.J.; Goverse, A.; et al. A Secreted SPRY Domain-Containing Protein (SPRYSEC) from the Plant-Parasitic Nematode Globodera rostochiensis Interacts with a CC-NB-LRR Protein from a Susceptible Tomato. Mol. Plant Microbe Interact 2009, 22, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Postma, W.J.; Slootweg, E.J.; Rehman, S.; Finkers-Tomczak, A.; Tytgat, T.O.; van Gelderen, K.; Lozano-Torres, J.L.; Roosien, J.; Pomp, R.; van Schaik, C.; et al. The effector SPRYSEC-19 of Globodera rostochiensis suppresses CC-NB-LRR-mediated disease resistance in plants. Plant Physiol. 2012, 160, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Castagnone-Sereno, P.; Semblat, J.P.; Castagnone, C. Modular architecture and evolution of the map-1 gene family in the root-knot nematode Meloidogyne incognita. Mol. Genet Genom. 2009, 282, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Lambert, K.N.; Bekal, S.; Domier, L.L.; Niblack, T.L.; Noel, G.R.; Smyth, C.A. Selection of Heterodera glycines chorismate mutase-1 alleles on nematode-resistant soybean. Mol. Plant Microbe Interact 2005, 18, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Bekal, S.; Niblack, T.L.; Lambert, K.N. A chorismate mutase from the soybean cyst nematode Heterodera glycines shows polymorphisms that correlate with virulence. Mol. Plant Microbe Interact 2003, 16, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.A.; Liu, Q.L.; Williamson, V.M. Silencing a candidate nematode effector gene corresponding to the tomato resistance gene Mi-1 leads to acquisition of virulence. Mol. Plant Microbe Interact 2008, 21, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Torres, J.L.; Wilbers, R.H.P.; Gawronski, P.; Boshoven, J.C.; Finkers-Tomczak, A.; Cordewener, J.H.G.; America, A.H.P.; Overmars, H.A.; Van’t Klooster, J.W.; Baranowski, L.; et al. Dual disease resistance mediated by the immune receptor Cf-2 in tomato requires a common virulence target of a fungus and a nematode. Proc. Natl. Acad. Sci. USA 2012, 109, 10119–10124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacco, M.A.; Koropacka, K.; Grenier, E.; Jaubert, M.J.; Blanchard, A.; Goverse, A.; Smant, G.; Moffett, P. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2-and RanGAP2-Dependent Plant Cell Death. PLoS Pathog. 2009, 5, e1000564. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, A.; Esquibet, M.; Fouville, D.; Grenier, E. Ranbpm homologue genes characterised in the cyst nematodes Globodera pallida and Globodera ‘mexicana’. Physiol. Mol. Plant Pathol. 2005, 67, 15–22. [Google Scholar] [CrossRef]
- Semblat, J.P.; Rosso, M.N.; Hussey, R.S.; Abad, P.; Castagnone-Sereno, P. Molecular cloning of a cDNA encoding an amphid-secreted putative avirulence protein from the root-knot nematode Meloidogyne incognita. Mol. Plant Microbe Interact 2001, 14, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, J.; Grenier, E.; Esquibet, M.; Hamel, L.P.; Moffett, P.; Manzanares-Dauleux, M.J.; Kerlan, M.C. Evolution and variability of Solanum RanGAP2, a cofactor in the incompatible interaction between the resistance protein GPA2 and the Globodera pallida effector Gp-RBP-1. BMC Evol. Biol. 2013, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Shields, J.; Allen, R.; Hussey, R.S. Molecular cloning and characterisation of a venom allergen AG5-like cDNA from Meloidogyne incognita. Int. J. Parasitol. 2000, 30, 77–81. [Google Scholar] [CrossRef]
- Wang, X.H.; Replogle, A.; Davis, E.L.; Mitchum, M.G. The tobacco Cel7 gene promoter is auxin-responsive and locally induced in nematode feeding sites of heterologous plants. Mol. Plant Pathol. 2007, 8, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Koh, Y.H.; Moon, Y.S.; Lee, S.H. Molecular properties of a venom allergen-like protein suggest a parasitic function in the pinewood nematode Bursaphelenchus xylophilus. Int. J. Parasitol. 2012, 42, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Torres, J.L.; Wilbers, R.H.P.; Warmerdam, S.; Finkers-Tomczak, A.; Diaz-Granados, A.; van Schaik, C.C.; Helder, J.; Bakker, J.; Goverse, A.; Schots, A.; et al. Apoplastic Venom Allergen-like Proteins of Cyst Nematodes Modulate the Activation of Basal Plant Innate Immunity by Cell Surface Receptors. PLoS Pathog. 2014, 10, e1004569. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Magne, M.; Chen, S.; Cote, O.; Stare, B.G.; Obradovic, N.; Jamshaid, L.; Wang, X.; Belair, G.; Moffett, P. Analysis of putative apoplastic effectors from the nematode, Globodera rostochiensis, and identification of an expansin-like protein that can induce and suppress host defenses. PLoS ONE 2015, 10, e0115042. [Google Scholar] [CrossRef] [PubMed]
- Iberkleid, I.; Vieira, P.; de Almeida Engler, J.; Firester, K.; Spiegel, Y.; Horowitz, S.B. Fatty acid-and retinol-binding protein, Mj-FAR-1 induces tomato host susceptibility to root-knot nematodes. PLoS ONE 2013, 8, e64586. [Google Scholar] [CrossRef] [PubMed]
- Mantelin, S.; Thorpe, P.; Jones, J.T. Suppression of Plant Defences by Plant-Parasitic Nematodes. 2015, 73, 325–337. [Google Scholar]
- Liu, S.; Kandoth, P.K.; Warren, S.D.; Yeckel, G.; Heinz, R.; Alden, J.; Yang, C.; Jamai, A.; El-Mellouki, T.; Juvale, P.S.; et al. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature 2012, 492, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.F.; Beard, H.; MacDonald, M.H.; Kabir, S.; Youssef, R.M.; Hosseini, P.; Brewer, E. Engineered resistance and hypersusceptibility through functional metabolic studies of 100 genes in soybean to its major pathogen, the soybean cyst nematode. Planta 2013, 237, 1337–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wen, Z.; Li, W.; Zhang, Y.; Zhang, L.; Dai, H.; Wang, D.; Xu, R. Genome-wide association study for soybean cyst nematode resistance in Chinese elite soybean cultivars. Mol. Breed. 2017, 37, 60. [Google Scholar] [CrossRef]
- Grunewald, W.; Cannoot, B.; Friml, J.; Gheysen, G. Parasitic Nematodes Modulate PIN-Mediated Auxin Transport to Facilitate Infection. PLoS Pathog. 2009, 5, e1000266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, W.; van Noorden, G.; van Isterdael, G.; Beeckman, T.; Gheysen, G.; Mathesius, U. Manipulation of Auxin Transport in Plant Roots during Rhizobium Symbiosis and Nematode Parasitism. Plant Cell 2009, 21, 2553–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, T.; Sugiyama, S.; Matsuura, H.; Arie, T.; Masuta, C. Resistant and susceptible responses in tomato to cyst nematode are differentially regulated by salicylic acid. Plant Cell Physiol. 2010, 51, 1524–1536. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, E.; Alba, E.; Montes, M.J.; Delibes, A.; Lopez-Brana, I. Analysis of ascorbate peroxidase genes expressed in resistant and susceptible wheat lines infected by the cereal cyst nematode, Heterodera avenae. Plant Cell Rep. 2010, 29, 1169–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Chronis, D.; Kenning, C.; Peret, B.; Hewezi, T.; Davis, E.L.; Baum, T.J.; Hussey, R.; Bennett, M.; Mitchum, M.G. The Novel Cyst Nematode Effector Protein 19C07 Interacts with the Arabidopsis Auxin Influx Transporter LAX3 to Control Feeding Site Development. Plant Physiol. 2011, 155, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Doyle, E.A.; Lambert, K.N. Meloidogyne javanicaChorismate Mutase 1 Alters Plant Cell Development. Mol. Plant Microbe Interact 2003, 16, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Piya, S.; Richard, G.; Rice, H.J. Spatial and temporal expression patterns of auxin response transcription factors in the syncytium induced by the beet cyst nematode Heterodera schachtii in Arabidopsis. Mol. Plant Pathol. 2014, 15, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Beneventi, M.A.; da Silva, O.B., Jr.; de Sa, M.E.; Firmino, A.A.; de Amorim, R.M.; Albuquerque, E.V.; da Silva, M.C.; da Silva, J.P.; Campos Mde, A.; Lopes, M.J.; et al. Transcription profile of soybean-root-knot nematode interaction reveals a key role of phythormones in the resistance reaction. BMC Genom. 2013, 14, 322. [Google Scholar] [CrossRef] [PubMed]
- Siddique, S.; Radakovic, Z.S.; De La Torre, C.M.; Chronis, D.; Novák, O.; Ramireddy, E.; Holbein, J.; Matera, C.; Hütten, M.; Gutbrod, P.; et al. A parasitic nematode releases cytokinin that controls cell division and orchestrates feeding site formation in host plants. Proc. Natl. Acad. Sci. USA 2015, 112, 12669–12674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, C.; Dutta, T.K.; Banakar, P.; Rao, U. Comparing the defence-related gene expression changes upon root-knot nematode attack in susceptible versus resistant cultivars of rice. Sci. Rep. 2016, 6, 22846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Kjemtrup-Lovelace, S.; Li, C.; Luo, Y.; Chen, L.P.; Song, B.-H. Comparative RNA-Seq Analysis Uncovers a Complex Regulatory Network for Soybean Cyst Nematode Resistance in Wild Soybean (Glycine soja). Sci. Rep. 2017, 7, 9699. [Google Scholar] [CrossRef] [PubMed]
- Gillet, F.-X.; Bournaud, C.; de Souza Júnior, J.D.A.; Fatima Grossi-de-Sa, M. Plant-parasitic nematodes: Towards understanding molecular players in stress responses. Ann. Bot. 2017, 119, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Dong, X. Systemic Acquired Resistance. Ann. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Durner, J.; Shah, J.; Klessig, D.F. Salicylic acid and disease resistance in plants. Trends Plant Sci. 1997, 2, 266–274. [Google Scholar] [CrossRef]
- Draper, J. Salicylate, superoxide synthesis and cell suicide in plant defence. Trends Plant Sci. 1997, 2, 162–165. [Google Scholar] [CrossRef]
- Overmyer, K.; Brosche, M.; Kangasjarvi, J. Reactive oxygen species and hormonal control of cell death. Trends Plant Sci. 2003, 8, 335–342. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Pathogen-induced, NADPH oxidase–derived reactive oxygen intermediates suppress spread of cell death in Arabidopsis thaliana. Nat. Genet. 2005, 37, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Roze, E.; Hanse, B.; Mitreva, M.; Vanholme, B.; Bakker, J.; Smant, G. Mining the secretome of the root-knot nematode Meloidogyne chitwoodi for candidate parasitism genes. Mol. Plant Pathol. 2008, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.G.; Jesuino, R.S.; Dantas Ada, S.; Brigido Mde, M.; Felipe, M.S. Oxidative stress response in Paracoccidioides brasiliensis. Genet. Mol. Res. 2005, 4, 409–429. [Google Scholar] [PubMed]
- Robertson, L.; Robertson, W.M.; Sobczak, M.; Helder, J.; Tetaud, E.; Ariyanayagam, M.R.; Ferguson, M.A.J.; Fairlamb, A.; Jones, J.T. Cloning, expression and functional characterisation of a peroxiredoxin from the potato cyst nematode Globedera rostochiensis. Mol. Biochem. Parasit. 2000, 111, 41–49. [Google Scholar] [CrossRef]
- Sunkar, R. MicroRNAs in Plant Development and Stress Responses; Springer: Heidelberg/Berlin, Germany, 2012; Volume 15. [Google Scholar]
- Tian, B.; Wang, S.; Todd, T.C.; Johnson, C.D.; Tang, G.; Trick, H.N. Genome-wide identification of soybean microRNA responsive to soybean cyst nematodes infection by deep sequencing. BMC Genom. 2017, 18, 572. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The Arabidopsis MicroRNA396-GRF1/GRF3 Regulatory Module Acts as a Developmental Regulator in the Reprogramming of Root Cells during Cyst Nematode Infection. Plant Physiol. 2012, 159, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Zhang, S.; Liu, D.; Duan, Y.; Dong, W. Identification of soybean microRNAs involved in soybean cyst nematode infection by deep sequencing. PLoS ONE 2012, 7, e39650. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Shukla, N.; Joshi, G.; VijayaKumar, C.; Jagannath, A.; Agarwal, M.; Goel, S.; Kumar, A. Genome-wide identification and characterization of miRNAome from tomato (Solanum lycopersicum) roots and root-knot nematode (Meloidogyne incognita) during susceptible interaction. PLoS ONE 2017, 12, e0175178. [Google Scholar] [CrossRef] [PubMed]
- Gheysen, G.; Vanholme, B. RNAi from plants to nematodes. Trends Biotechnol. 2007, 25, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Paiva, G.; Proença, D.N.; Francisco, R.; Verissimo, P.; Santos, S.S.; Fonseca, L.; Abrantes, I.M.O.; Morais, P.V. Nematicidal Bacteria Associated to Pinewood Nematode Produce Extracellular Proteases. PLoS ONE 2013, 8, e79705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klink, V.P.; Matthews, B.F. Emerging approaches to broaden resistance of soybean to soybean cyst nematode as supported by gene expression studies. Plant Physiol. 2009, 151, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.R.; Hewezi, T.; Peng, J.; Baum, T.J. Isolation of whole esophageal gland cells from plant-parasitic nematodes for transcriptome analyses and effector identification. Mol. Plant Microbe Interact 2013, 26, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Andolfo, G.; Iovieno, P.; Frusciante, L.; Ercolano, M.R. Genome-Editing Technologies for Enhancing Plant Disease Resistance. Front. Plant. Sci. 2016, 7, 1813. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hypothetical overview of nematode-associated molecular pattern (NAMP)-triggered immunity (NTI) in the Arabidopsis-beet cyst nematode H. schachtii model system. NAMPs are specific molecular patterns released by plant parasitic nematodes which are recognized by transmembrane receptors such as kinases on the host to induce a signaling cascade [13]. The overexpression of MKK4 in Arabidopsis led to increased resistance against beet cyst nematode as compared to wild type, suggesting an important role of MKK4 in signal transduction in plant immunity [45]. MAPK phosphatase AP2C1 may be involved in the putative dephosphorylation of MKK (MAP kinase kinase) into the MPK3 and MPK6 kinases to further transduce the signal for nematode resistance [38]. We have experimentally demonstrated that the overexpression of WRKY33 and the knocking-out of PAD3 resulted in enhanced nematode resistance, which highly support our hypothesis that the phosphorylation cascade involving WRKY33 leads to camalexin biosynthesis which in turn leads to NTI [45]. Although some steps, such as the phosphorylation of MKK4 and MSK1 and the activation of WRKY33, in this cascade have not yet been demonstrated in plant–nematode interactions, the proposed NTI model indicates the importance of WRKY33-dependent camalexin production in Arabidopsis.
Figure 1.
Hypothetical overview of nematode-associated molecular pattern (NAMP)-triggered immunity (NTI) in the Arabidopsis-beet cyst nematode H. schachtii model system. NAMPs are specific molecular patterns released by plant parasitic nematodes which are recognized by transmembrane receptors such as kinases on the host to induce a signaling cascade [13]. The overexpression of MKK4 in Arabidopsis led to increased resistance against beet cyst nematode as compared to wild type, suggesting an important role of MKK4 in signal transduction in plant immunity [45]. MAPK phosphatase AP2C1 may be involved in the putative dephosphorylation of MKK (MAP kinase kinase) into the MPK3 and MPK6 kinases to further transduce the signal for nematode resistance [38]. We have experimentally demonstrated that the overexpression of WRKY33 and the knocking-out of PAD3 resulted in enhanced nematode resistance, which highly support our hypothesis that the phosphorylation cascade involving WRKY33 leads to camalexin biosynthesis which in turn leads to NTI [45]. Although some steps, such as the phosphorylation of MKK4 and MSK1 and the activation of WRKY33, in this cascade have not yet been demonstrated in plant–nematode interactions, the proposed NTI model indicates the importance of WRKY33-dependent camalexin production in Arabidopsis.
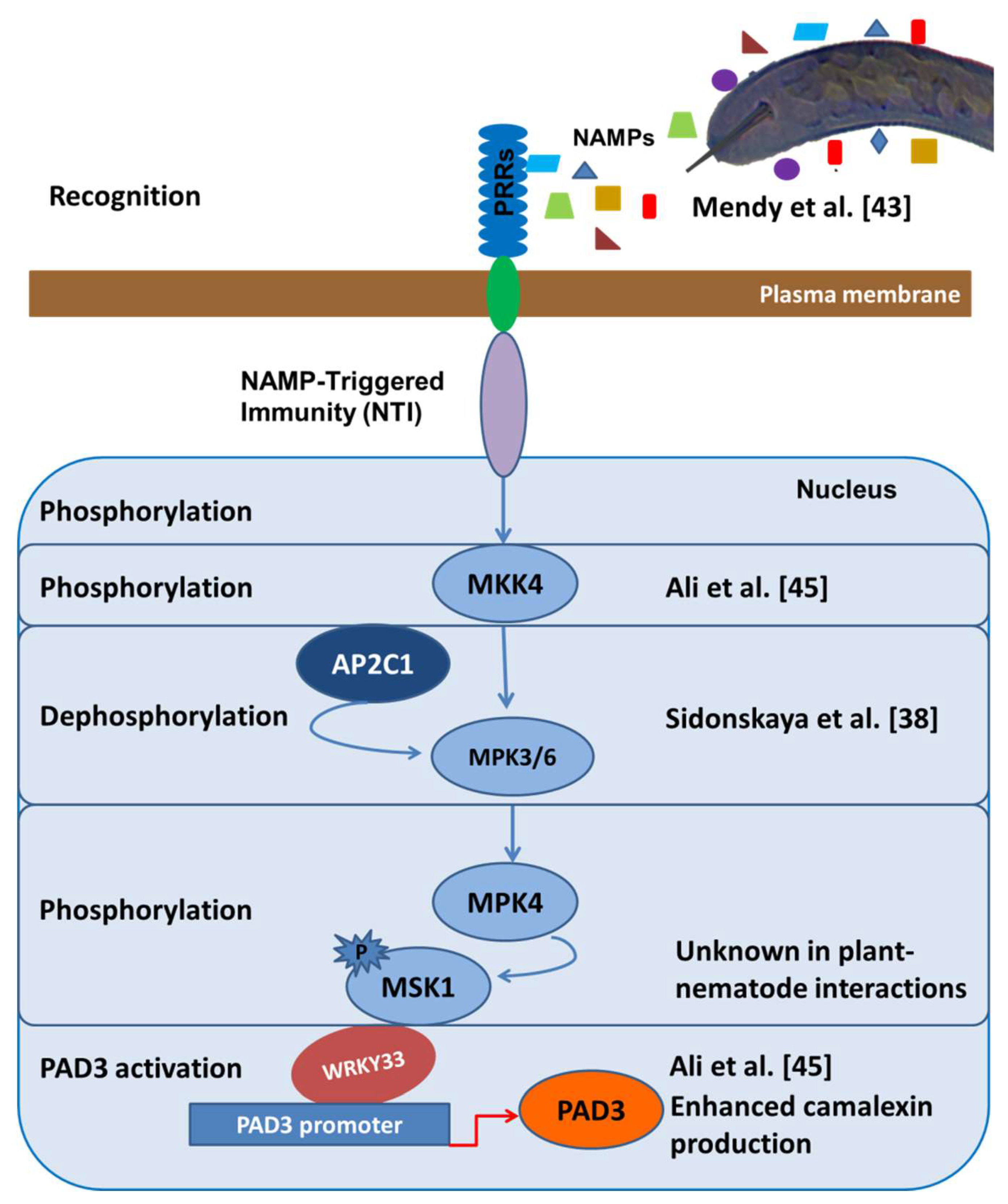
Figure 2.
Conservation of the CLE motif (xRxxPxGPDPxHx) in CLAVATA/ESR (CLE) peptides in different plant parasitic nematodes (PPNs) and plant species [4]. This multiple alignment demonstrates the highly conserved amino acid residues of CLE motif between PPNs and their corresponding host plants.
Figure 2.
Conservation of the CLE motif (xRxxPxGPDPxHx) in CLAVATA/ESR (CLE) peptides in different plant parasitic nematodes (PPNs) and plant species [4]. This multiple alignment demonstrates the highly conserved amino acid residues of CLE motif between PPNs and their corresponding host plants.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ali, M.A.; Anjam, M.S.; Nawaz, M.A.; Lam, H.-M.; Chung, G. Signal Transduction in Plant–Nematode Interactions. Int. J. Mol. Sci. 2018, 19, 1648. https://doi.org/10.3390/ijms19061648
AMA Style
Ali MA, Anjam MS, Nawaz MA, Lam H-M, Chung G. Signal Transduction in Plant–Nematode Interactions. International Journal of Molecular Sciences. 2018; 19(6):1648. https://doi.org/10.3390/ijms19061648
Chicago/Turabian StyleAli, Muhammad Amjad, Muhammad Shahzad Anjam, Muhammad Amjad Nawaz, Hon-Ming Lam, and Gyuhwa Chung. 2018. "Signal Transduction in Plant–Nematode Interactions" International Journal of Molecular Sciences 19, no. 6: 1648. https://doi.org/10.3390/ijms19061648
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.