1. Introduction
Neurodegenerative diseases account for a significant proportion of morbidity and mortality in the developed countries. Moreover, these disorders are becoming more frequent due to the increased life expectancy [
1]. Alzheimer’s disease (AD), the most common type of senile dementia, is a neurodegenerative disorder with enormous social and economic impact [
2]. Although the cause(s) of AD is still controversial, it is accepted that the accumulation of amyloid β peptide (Aβ) in brain plaques triggers downstream neurotoxic events, leading to neuronal dysfunction, cell death, and neurodegeneration [
3]. Aβ is a 38–43 amino acid peptide which derives from the β-amyloid precursor protein (βAPP) through proteolytic processing by β- and γ-secretases. Aβ 1–42 (Aβ
1–42) appears to be the species that first forms deposits in AD [
2], and its role in the oxidative damage in AD brain has also been established [
4]. These findings suggest amyloid plaques or Aβ production as targets for drug development [
1]. In addition, compounds that exhibit antioxidant activity can be considered potential therapeutic agents for AD [
5]. In this context, the nematode
Caenorhabditis elegans engineered to express human Aβ
1–42 is a convenient in vivo model that has been used in drug screening for potential AD therapeutics. The resulting transgenic strains develop a concomitant progressive paralysis phenotype, being a well-suited model for correlating Aβ expression and toxicity [
6,
7,
8].
Prolyl endopeptidase (PEP; E.C. 3.4.21.26) also called prolyl oligopeptidase (POP) is a highly conserved serine protease enzyme that cleaves peptide bonds at the carboxyl side of proline residues (P) in peptides up to 30 amino acids long. It has been found in a wide range of species and tissues, especially in the human brain. PEP can degrade biologically active P-containing peptides, including peptide-like hormones, neuroactive peptides, and various cellular factors [
9]. Moreover, levels of PEP activity are altered in many degenerative conditions and psychiatric disorders, such as AD, amnesia, depression, and schizophrenia, and therefore, the enzyme is a potential therapeutic target for these diseases [
10]. PEP was suggested to be a putative γ-secretase and accordingly PEP inhibitors abolished the formation of Aβ in neuroblastoma cells and prevented amyloid deposition in a mouse model of accelerated senescence [
11]. However, PEP is not only involved in cleaving off physiologically active peptides, and it has been speculated that its physiological role results from its direct interaction with partner proteins [
12].
The potential therapeutic use of peptides derived from natural sources has been extensively discussed during the last decades [
13,
14,
15]. Dietary peptides have been claimed to have positive effects on weight loss and glycemia management [
16] and also in the prevention and treatment of cancer, cardiovascular and infective diseases, and mental health disorders [
17,
18]. Moreover, many peptides seem to act through more than a single mechanism of action and, hence, they can be considered multifunctional sequences [
19,
20]. Natural peptides also represent an excellent starting point as leading candidates for the rational design of synthetic sequences with improved biological activity, specificity, and stability [
21].
Natural sources of PEP-inhibitory peptides mainly include meat and fish by-products [
22,
23,
24,
25,
26], cereals [
27], and milk proteins [
25,
28,
29,
30]. By contrast, peptides showing protection against the toxicity caused by the accumulation of Aβ are scarce. These potential neuroprotective peptides were isolated from a cocoa by-product [
31], maize [
32], and scorpion venom [
33]. Furthermore, small rationally-designed peptides based on specific Aβ motifs able to interact with Aβ, modifying its kinetics of aggregation, and reducing its toxicity have been described as an attractive pharmacological strategy [
34,
35].
Lactoferrin (LF), a well-characterized component of milk whey, is a multifunctional iron glycoprotein that exhibits a diverse range of biological activities including antimicrobial, antiviral, antioxidant, and immunomodulatory activities [
36]. LF-derived peptides share some biological activities with the intact protein and possess antihypertensive properties [
20]. Recently, we have shown the inhibitory effect of an LF-based product on Aβ toxicity [
37], but there is no information about the neuroprotective effects of LF-derived peptides.
In the present study, we investigated the PEP-inhibitory activity of LF-derived peptides and of a set of sequence-related synthetic heptapeptides. Peptides exhibiting PEP-inhibitory activity in vitro were examined in a transgenic C. elegans model of AD to evaluate their in vivo protective effects against Aβ toxicity. With the aim of characterizing the mechanism(s) mediating C. elegans protection, we further examined their in vivo antioxidant effect and their in silico molecular interactions with Aβ. Finally, the role of tryptophan residues (W) on the neuroprotective activity of peptides was investigated.
3. Discussion
Currently, peptides have a wide range of applications in medicine and biotechnology. Moreover, multifunctionality is a common trait of many peptides, which might function as polypharmacological sequences. The present study characterizes new dual neuroprotective peptides that show in vitro PEP-inhibitory properties and C. elegans protection against Aβ1–42-associated toxicity in vivo. The neuroprotective peptides include natural sequences derived from the milk protein LF and sequence-related rationally-designed synthetic heptapeptides.
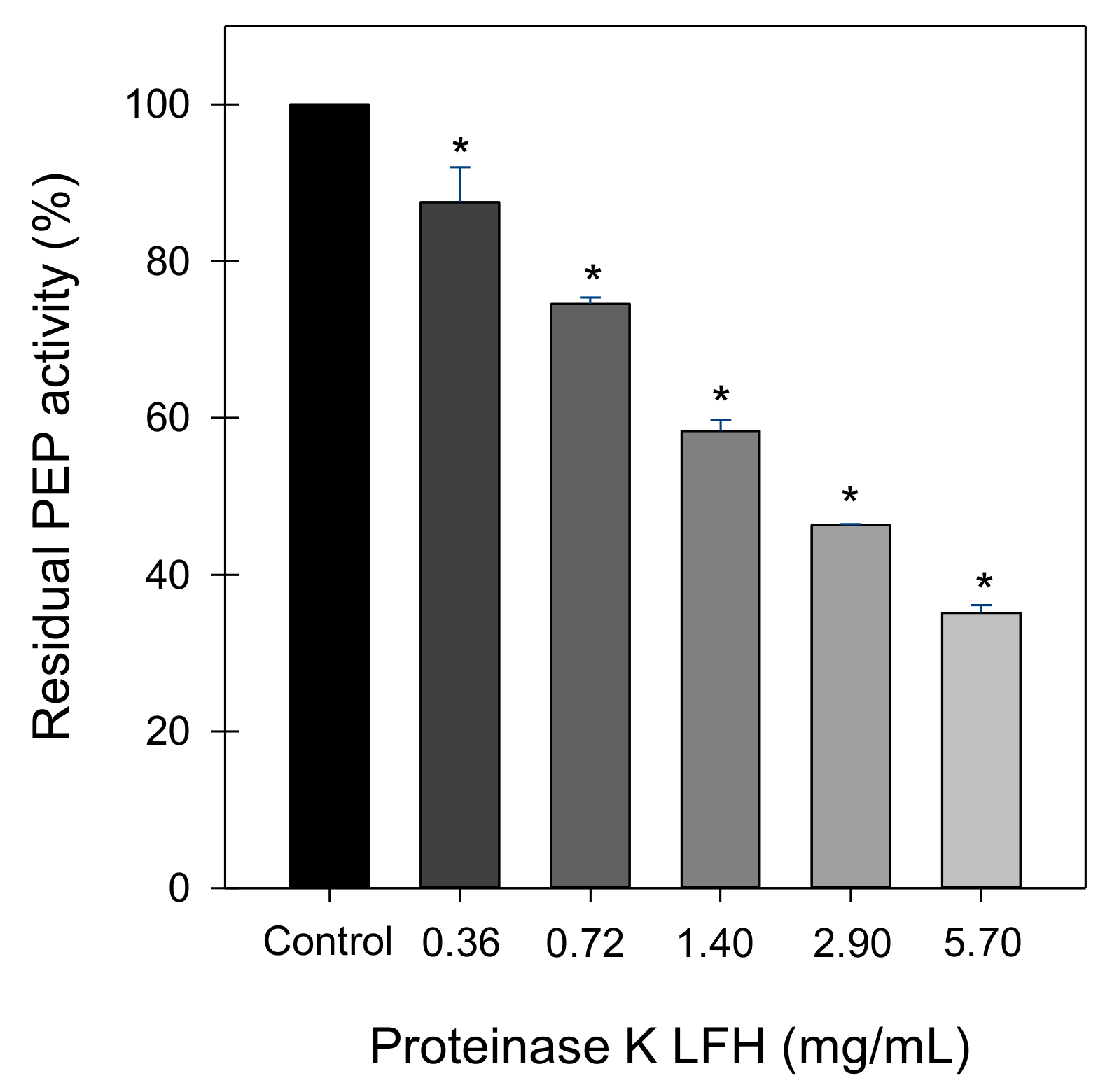
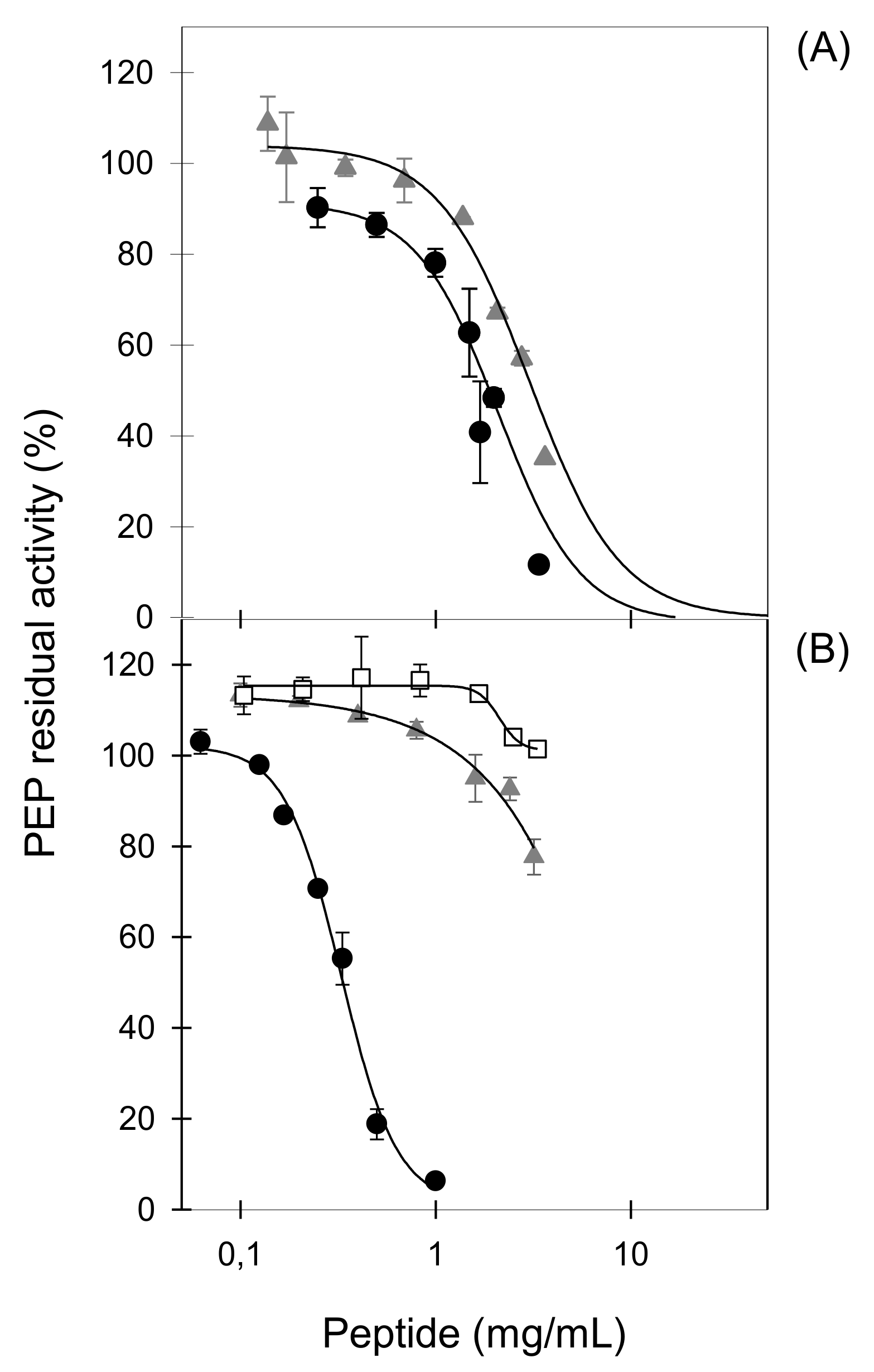
LFHs obtained with three different proteases showed a moderate ability to inhibit PEP that was not shown by non-hydrolysed LF, suggesting that LF-derived peptides possess the inhibitory activity. The hydrolysates generated with three different proteases showed different PEP-inhibitory activities pointing out the importance of peptide sequences for enzyme inhibiting activity. In our study, the most potent LFH was generated using proteinase K, which exhibits specificity for peptide bonds adjacent to the carboxylic group of aliphatic and aromatic amino acids [
40]. Moreover, the inhibitory activity of the hydrolysate was comparable to the previously reported IC
50 values of a sodium caseinate hydrolysate (0.77 mg/mL) [
29] and different fish protein hydrolysates (1.10–4.21 mg/mL) [
23]. LFHs obtained with proteinase K were previously described as in vitro inhibitors of the endothelin-converting enzyme (ECE) and ex vivo as inhibitors of ECE-dependent vasoconstriction [
38]. Our results describe a new biological activity related to neuroprotection for LF-derived peptides.
PEP inhibitors developed as potential therapeutic drugs are substrate-like compounds containing one P or one P-analogue residue [
10]. Most of the PEP-inhibitory peptides derived from natural sources described to date contain at least one and up to six P residues in their sequences, and from 3 to 18 amino acid residues in length [
22,
27,
29]. In spite of the fact that six of the eleven LF-derived peptides evaluated here contain one P residue in their sequences, none of them displayed PEP-inhibitory activity, as shown for some P-containing peptides derived from collagen or corn γ-zein [
27]. By contrast, the only two LF-derived peptides that provoked a modest in vitro enzyme inhibition (PKH8, NEGLTW; PKH11, SVDGKEDLIW) do not contain any P residue in their sequence, suggesting that both LF-derived peptides do not act as substrate-type inhibitors. Non-P-containing peptides derived from α-casein were also described as PEP inhibitors [
30].
Since the two LF-derived peptides with PEP-inhibitory activity described here shared a C-terminal W, we hypothesized its potential key contribution to enzyme inhibition. Our hypothesis was confirmed by the ability of the six sequence-related synthetic heptapeptides (PACEI48L to PACEI53L) to inhibit PEP. PACEI heptapeptides are the second generation of angiotensin converting enzyme inhibitory peptides based on two hexapeptides leads, PACEI32L (RKWHFW) and PACEI34L (RKWLFW) [
41]. Heptapeptides were designed by combinations of F, H, and L residues in positions 4–6, all of them share R, K, and W residues at the N-terminus and W residue at the C-terminus of a given heptapeptide (
Table 3) [
39]. Among these peptides, the highest inhibitory activity was recorded for PACEI50L with H, F, and L residues at positions 4, 5, and 6, respectively. The activity of this heptapeptide is sequence-specific, as demonstrated by the swapping of H and F residues (sequences PACEI50L and PACEI48L) which drastically reduced the PEP-inhibitory activity. These results confirm and extend previously reported data on how minor amino acid exchanges affect biological properties [
41,
42].
Further confirmation of the key role of W residue at the C-terminus was provided by the evaluation of PACEI50L and PKH11 variants that contain W to A substitutions. Our results showed that the effect of W residue on PEP-inhibitory activity is dependent on the peptide sequence since a severe reduction of the biological activity was observed for the two variants of PACEI50L while only a 40% reduction of the inhibitory potency was recorded for the PKH11 variant. Our results also pointed to the relevance of both W residues in the PACEI50L sequence since the total loss of PEP inhibition was achieved only with the double substitution of W residues at positions 3 and 7. In agreement with these results, the most potent PEP-inhibitory peptides named 13L and 9L identified from a cocoa hydrolysate contained W in their sequence (13L, DNYDNSAGKWWVT; 9L, NYDNSAGKW) and the lower IC
50 value (0.19 mg/mL) corresponded to the sequence 13L [
31]. Additional studies are required to explain the role of W residues in PEP-inhibitory peptides.
C. elegans is a suitable in vivo model for research on the molecular biology and genetics of different diseases as well as for drug and bioactive compound screenings [
43].
C. elegans models present alternative approaches to understanding neurodegenerative diseases for which there are currently few effective therapies [
6]. Here the transgenic
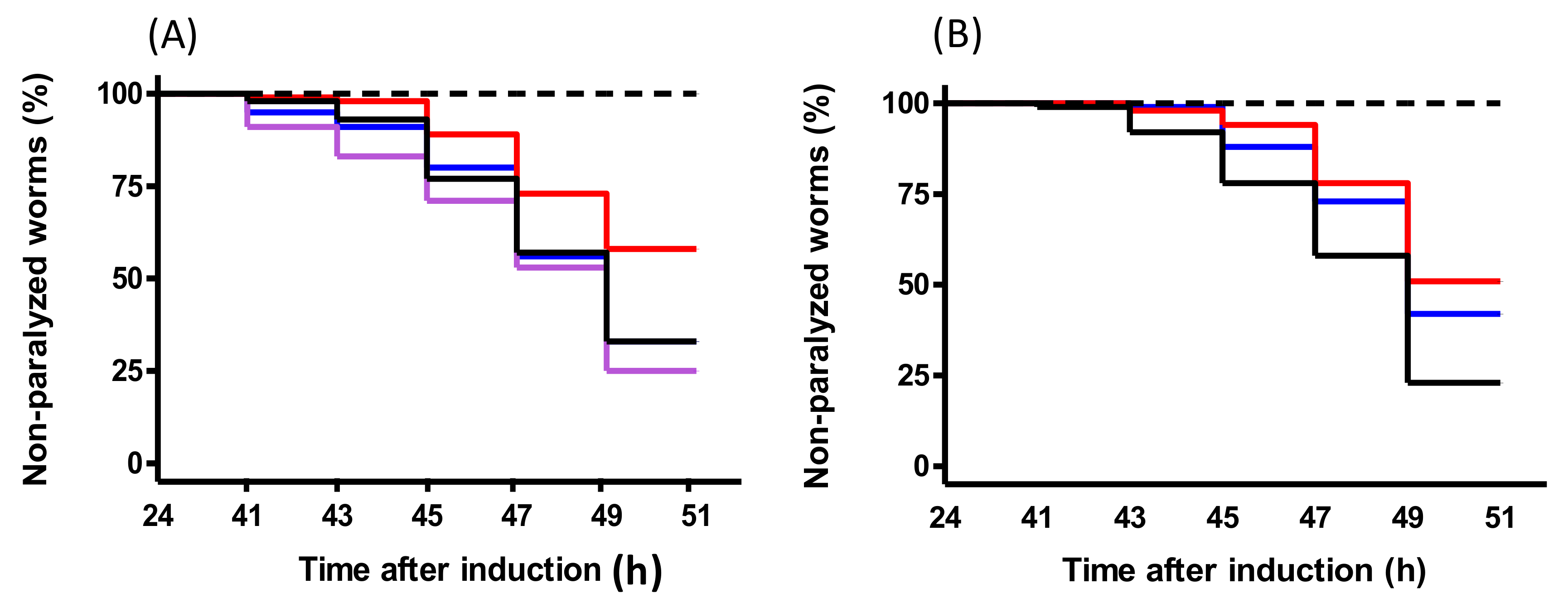
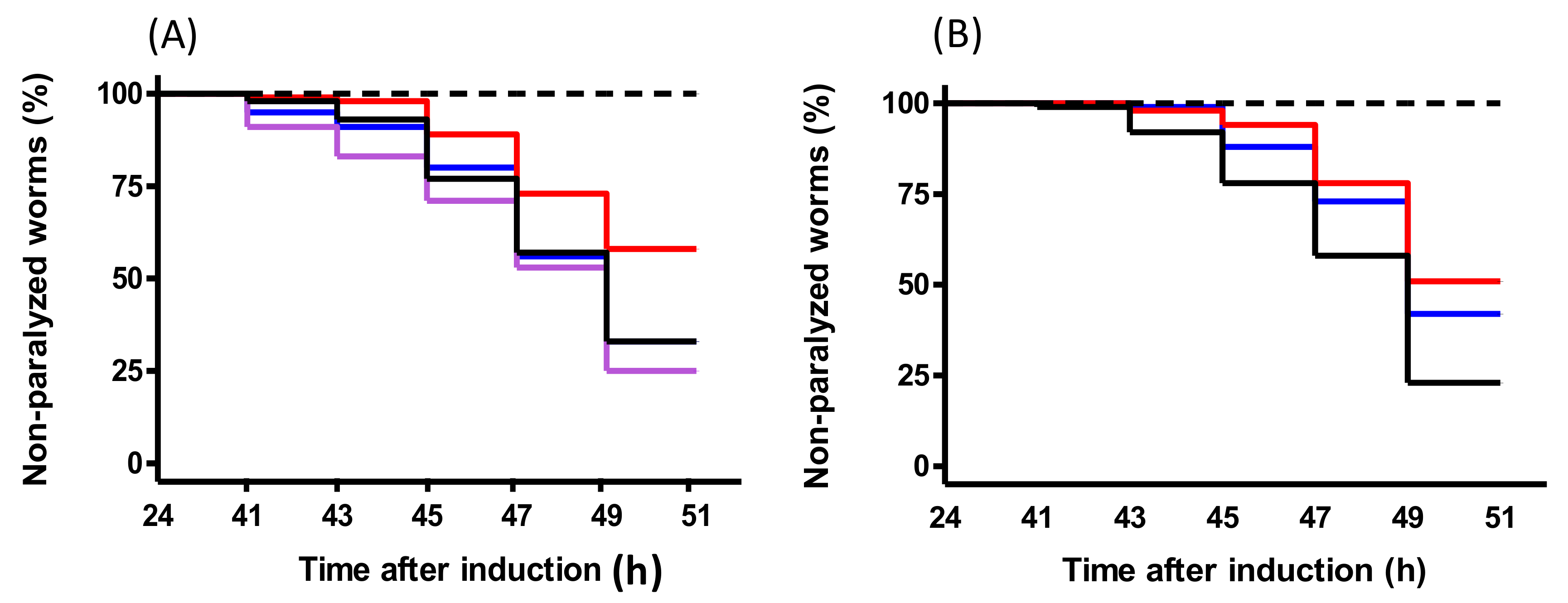
C. elegans model of AD, which develops a paralysis phenotype, was used to study the effects of peptides. PACEI50L and PKH11 treatments ameliorated Aβ-induced paralysis suggesting a potential in vivo protection from Aβ
1–42 toxicity, as described for cocoa and maize peptides [
31,
32] and also for a sequence purified from scorpion venom [
33]. Remarkably, and as observed for PEP-inhibitory activity, peptide variants of PACEI50L completely lost paralysis suppression in
C. elegans demonstrating the positive role of W residues in the context of the heptapeptide sequence evaluated.
The effect of different compounds on Aβ
1–42–mediated paralysis in transgenic
C. elegans has also been associated with antioxidant effects [
31,
32,
33,
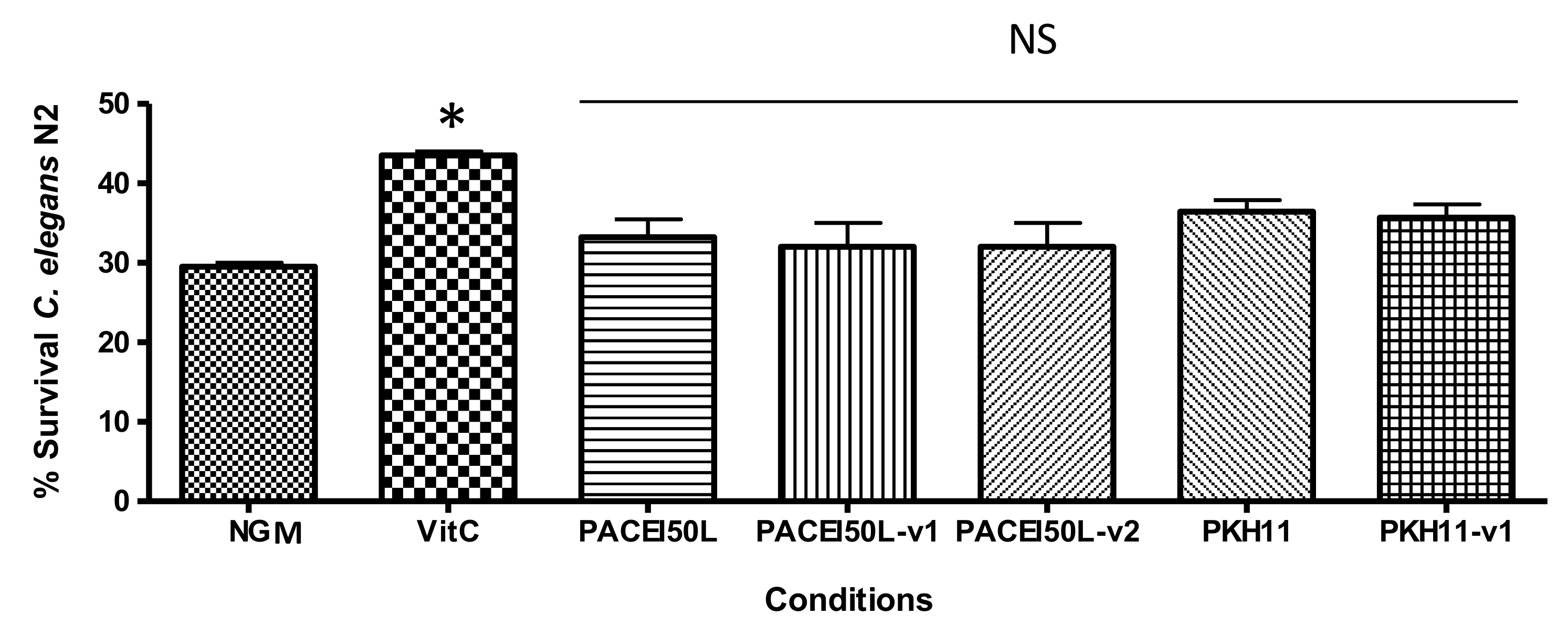
44]. In contrast to other neuroprotective peptides, the reduction of Aβ-toxicity by PACEI50L and PKH11 does not seem to operate through an antioxidant mechanism since both peptides did not produce significant
C. elegans protection upon oxidative stress under the conditions tested. Conversely, the antioxidant properties of maize and scorpion venom peptides might underline, at least in part, their protective effect observed in
C. elegans, since they inhibited the production of reactive oxygen species [
32,
33]. Additionally, in a similar experiment to the one described here, the peptide 13L from cocoa provided protection against oxidative stress, suggesting that the antioxidant activity might contribute to the protection against Aβ
1–42-induced damage [
31]. Therefore, our results suggest alternative mechanisms for PACEI50L and PKH11.
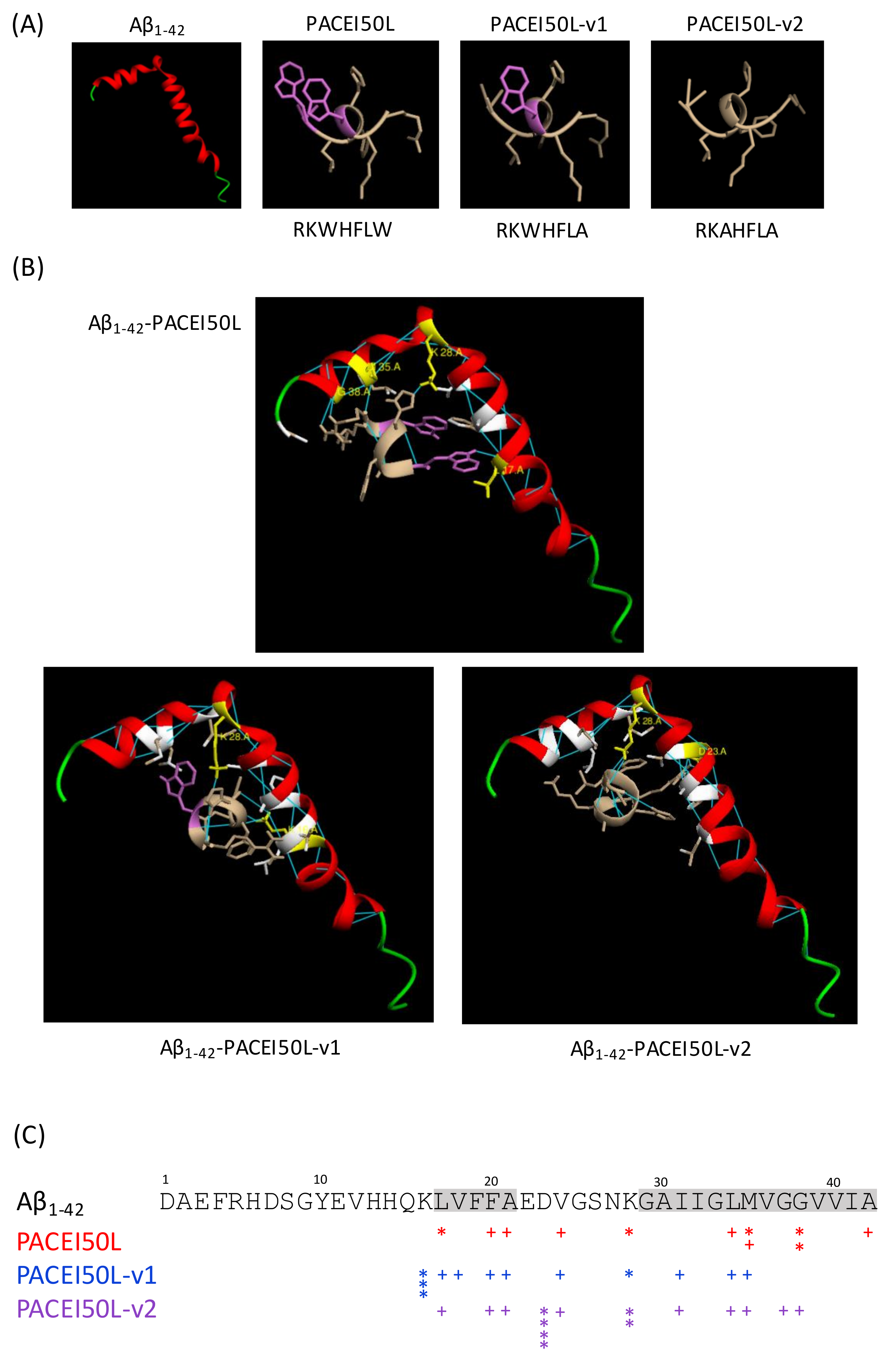
Aβ
1–42 contains hydrophobic motifs in the central (residues 17–21) and carboxy-terminal (residues 29–42) regions of the peptide [
45]. Amyloidogenic peptide aggregation seems to be primarily driven by these hydrophobic domains [
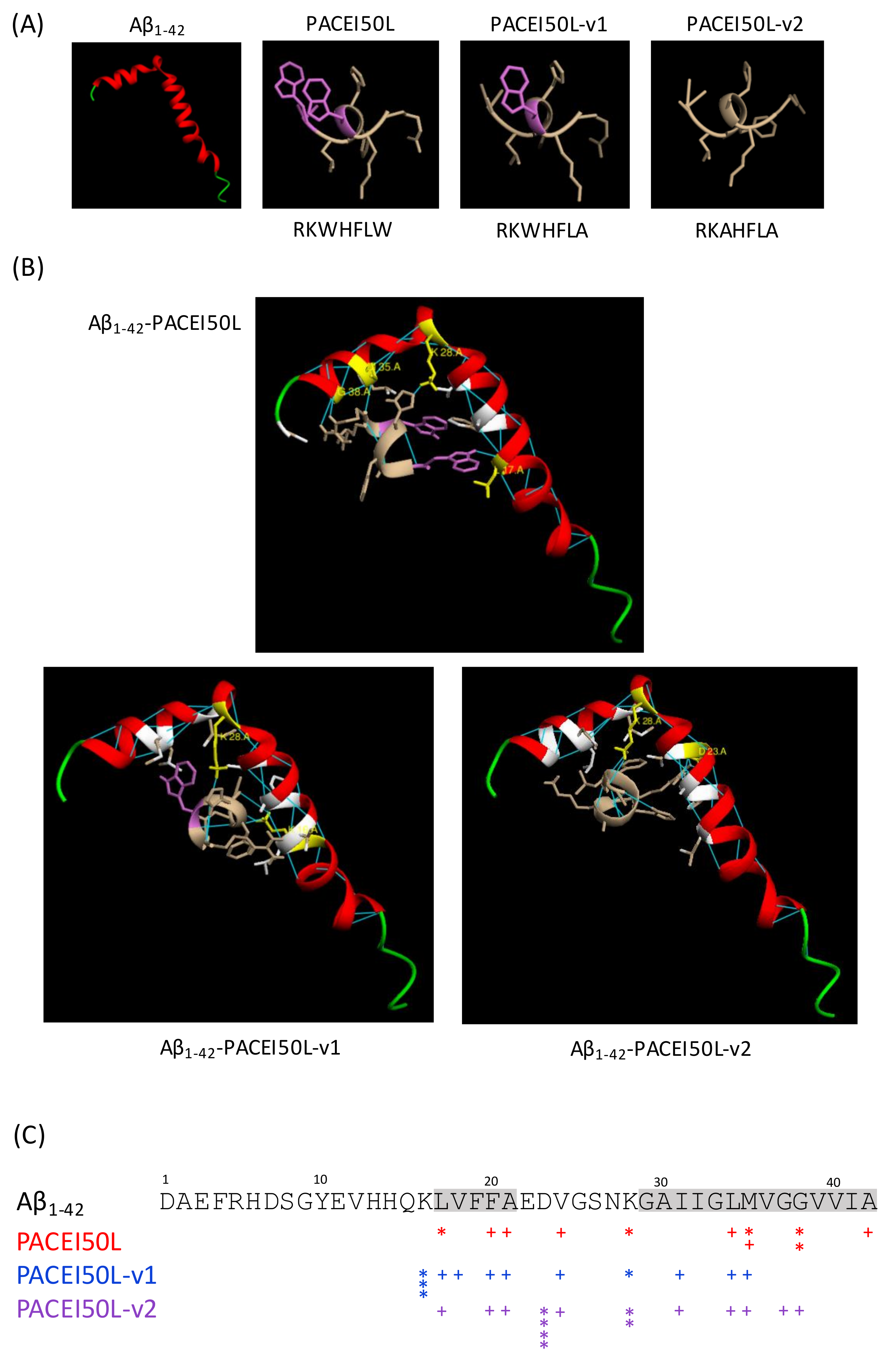
46]. Here, docking analysis showed that the potential hydrogen bond interactions between the active PACEI50L and Aβ
1–42 are focused on amino acids within both hydrophobic motifs (L17, M35, and G38) or close to them (K28). Particularly M35 and its hydrophobic surroundings seem to be important for the oxidative, neurotoxic, and aggregation properties of the peptide [
47,
48] whereas G residues can stabilize amyloidogenic structures by means of hydrogen bonds [
49]. The conformation adopted by Aβ
1–42 seems to be an important factor in amyloid formation since the Aβ
1–42 peptide with α-helical or random coil structure aggregates slowly while Aβ with β-sheet conformation aggregates rapidly [
45]. L17 included in the central hydrophobic stretch 17–21 contributes to adopt a β-sheet conformation that facilitates monomeric interaction, β-sheet oligomers, and amyloid fibrils [
50]. Moreover, amino acids 17–20 served as a template for designing synthetic peptides able to inhibit fibrillogenesis in a rat brain model of amyloidosis [
35]. Our docking results suggest that molecular interactions through hydrogen bonding between PACEI50L and residues L17, M35, and G38 from Aβ
1–42 might impede monomeric interactions and thus, Aβ oligomerization. Docking analysis revealed the loss of hydrogen bonds between non-active PACEI50L variants and these three residues in Aβ
1–42. Whether PACEI50L might interfere with the folding of Aβ
1–42 to form aggregates as suggested by in silico analysis requires further in vitro and in vivo research.
Our study underscores the important role of W for the bioactivity of neuroprotective peptides. W-containing peptides as those described here display several biological activities including antihypertensive, antioxidant, antidiabetic, and satiating properties [
51]. Besides W is the sole precursor of serotonin, which has been reported to have an effect on the psychological/cognitive function in humans. Remarkably, increased dietary W intake reduced intra-neuronal Aβ accumulation in a mouse model of AD, suggesting a neuroprotective role of W through the increase of serotonin levels [
52]. Increased W levels also extend longevity in
C. elegans and protect from alpha-synuclein and polyglutamine toxicity [
53,
54], suggesting a protective role against proteotoxicity in aging and age-related diseases. Additionally, in the
C. elegans model, supplementation with the W-containing 13L peptide upregulated the W metabolism, including genes involved in the synthesis of serotonin and other neurotransmitters [
31]. It is worthwhile to note that milk proteins and, among them, LF, are particularly rich in W in comparison with other dietary proteins opening the way to the future inclusion of LF in dietary recommendations for preventing or postponing AD.
In conclusion, we have identified novel natural and rationally-designed W-containing peptides showing in vitro PEP inhibition and in vivo protection from Aβ1–42 toxicity, although further research needs to be conducted in murine models to analyze the effectiveness of the peptides. These results add a new application to the antihypertensive proteinase K LFH and the synthetic heptapeptides confirming their multifunctionality. The effect of PACEI50L on delayed paralysis in C. elegans might be mediated, at least in part by the inhibition of Aβ1–42 oligomerization and thus, amyloid deposition, while the peptide antioxidant activity does not seem to be involved in the protective effect. Our results suggest that W-containing peptides are suitable starting leads to design effective neuroprotective agents. Further improvements to increase in vivo peptide stability based on the use of D-amino acid sequences are in progress. Future efforts are currently directed to clarify the mechanisms underlying the in vivo protective effects of W-containing peptides.

 and
and 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}