Therapeutic Potential of Sclareol in Experimental Models of Rheumatoid Arthritis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
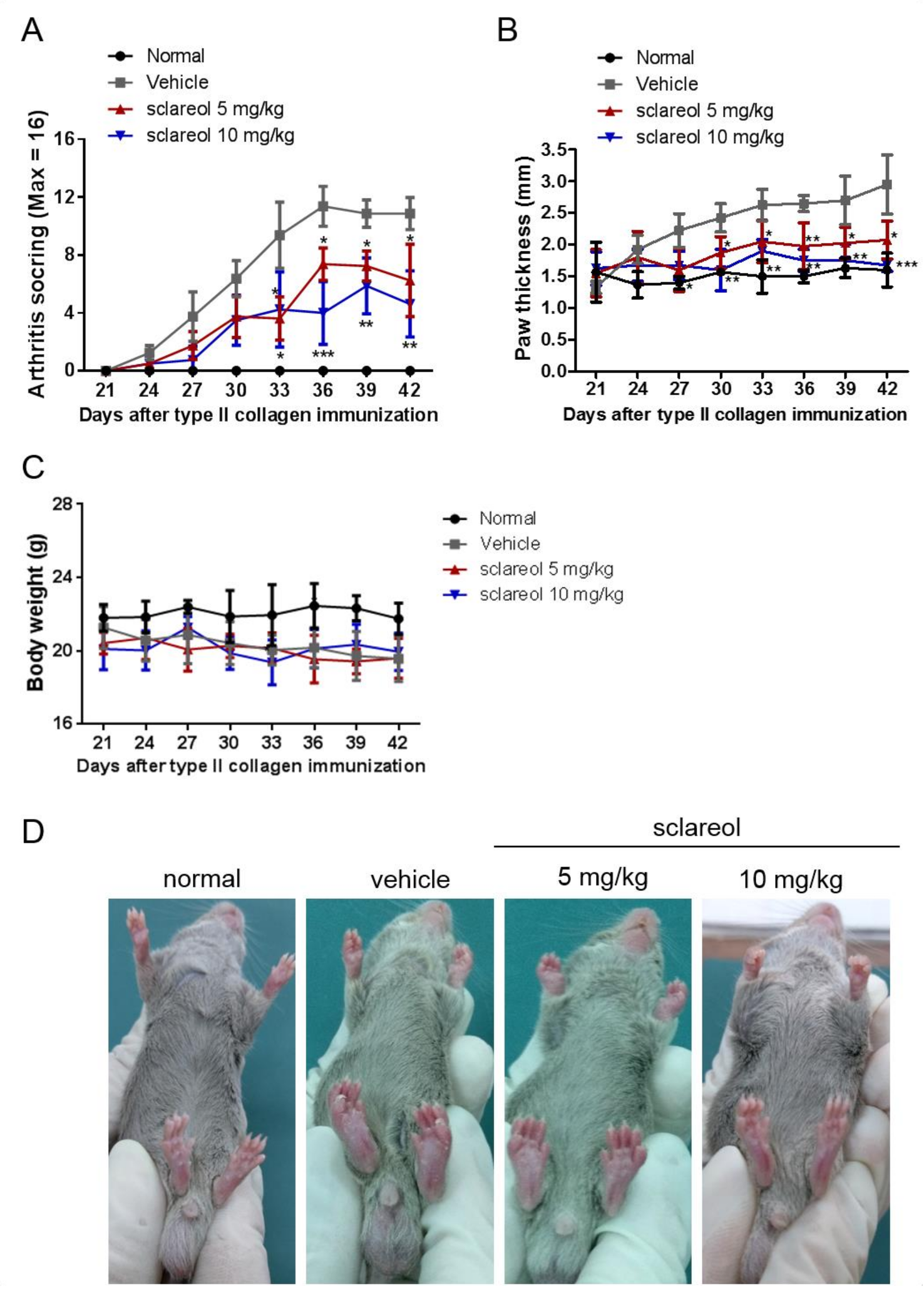
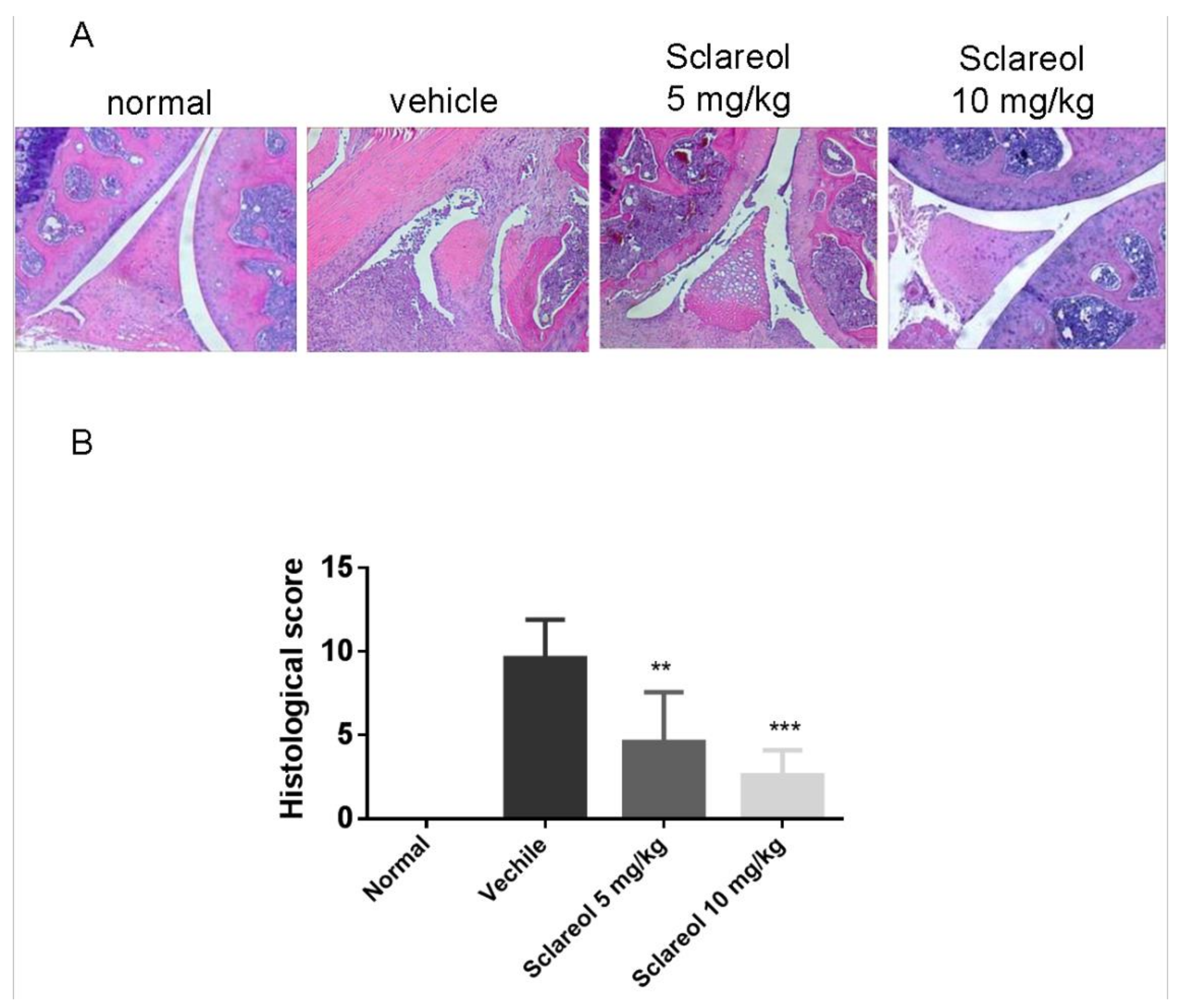
2.1. Amelioration of CIA by Sclareol Treatment
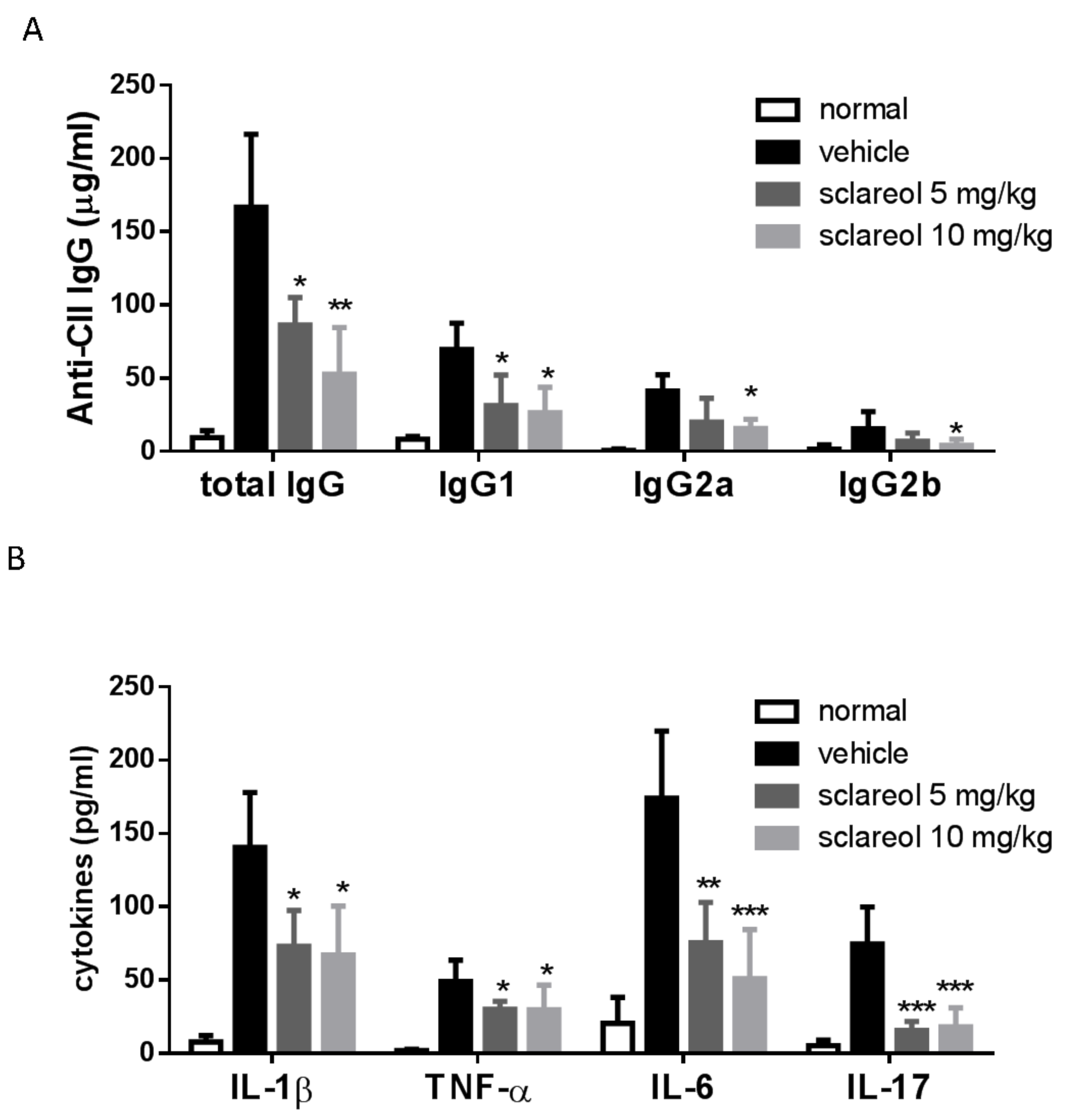
2.2. Decreased Levels of Circulating Anti-CII Abs and Cytokines in Sclareol-Treated CIA Mice
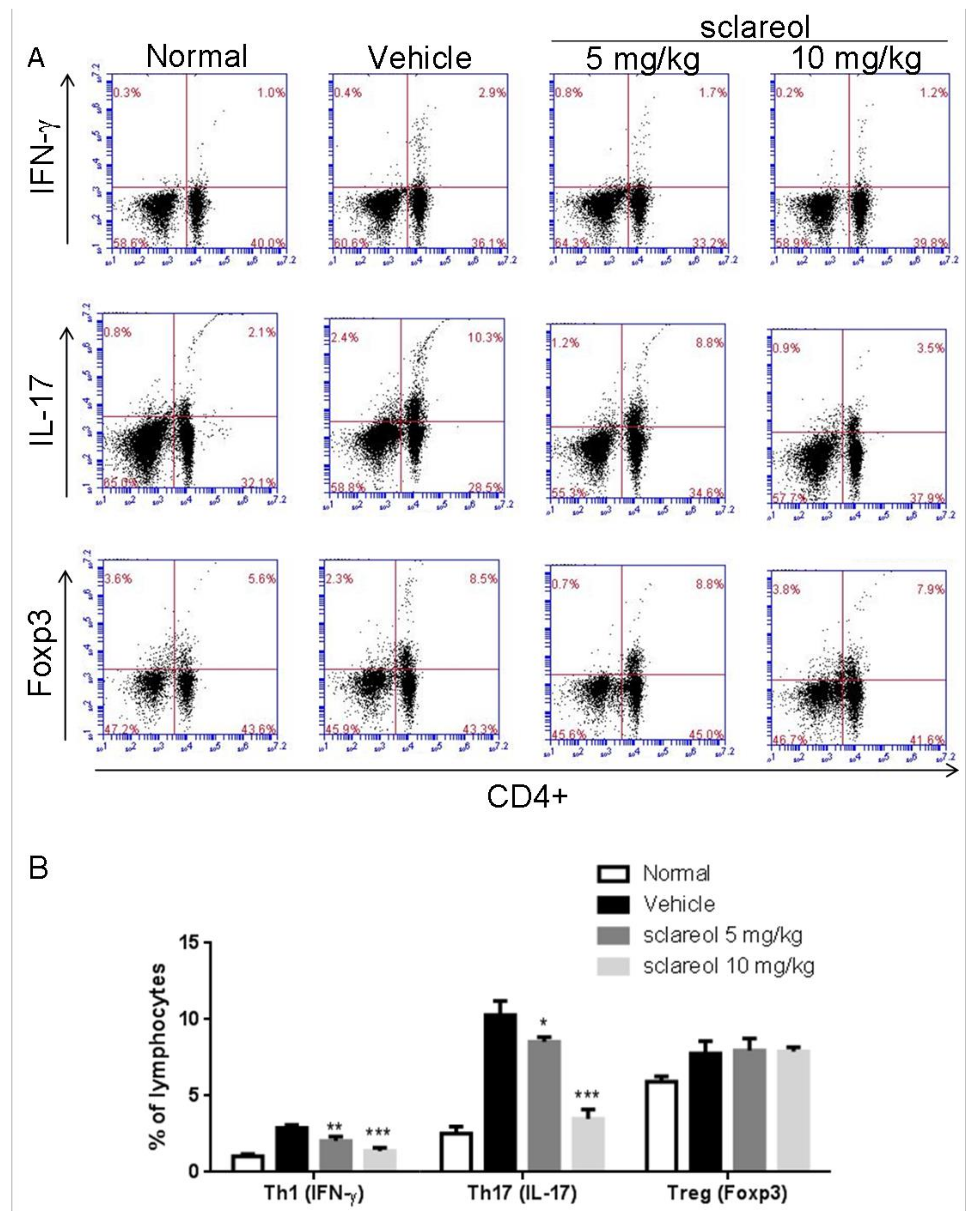
2.3. Altered Frequency of Th17 and Th1 Cells in Lymph Nodes by Sclareol Treatment
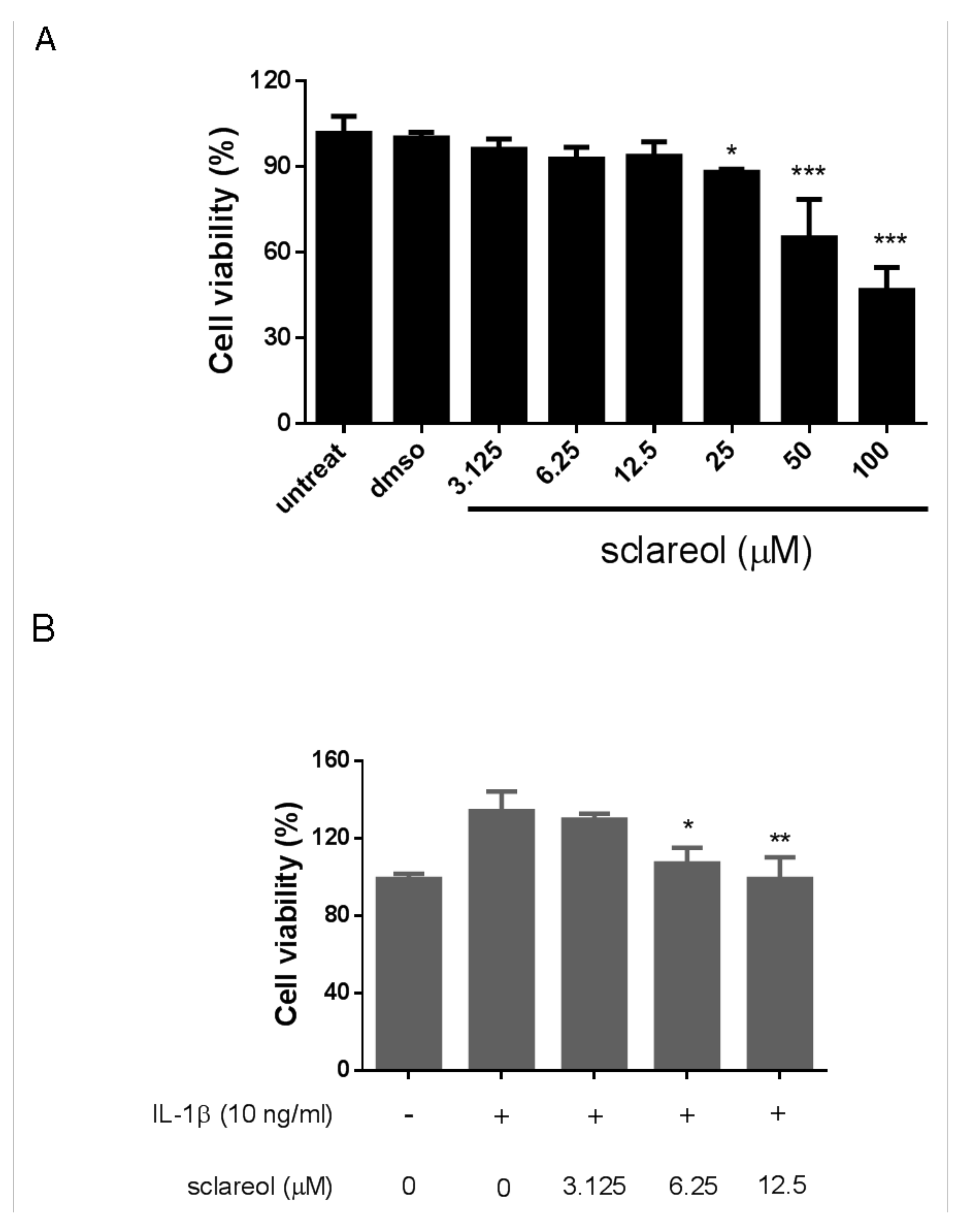
2.4. Cytotoxicity of Sclareol on Synovial Cells In Vitro
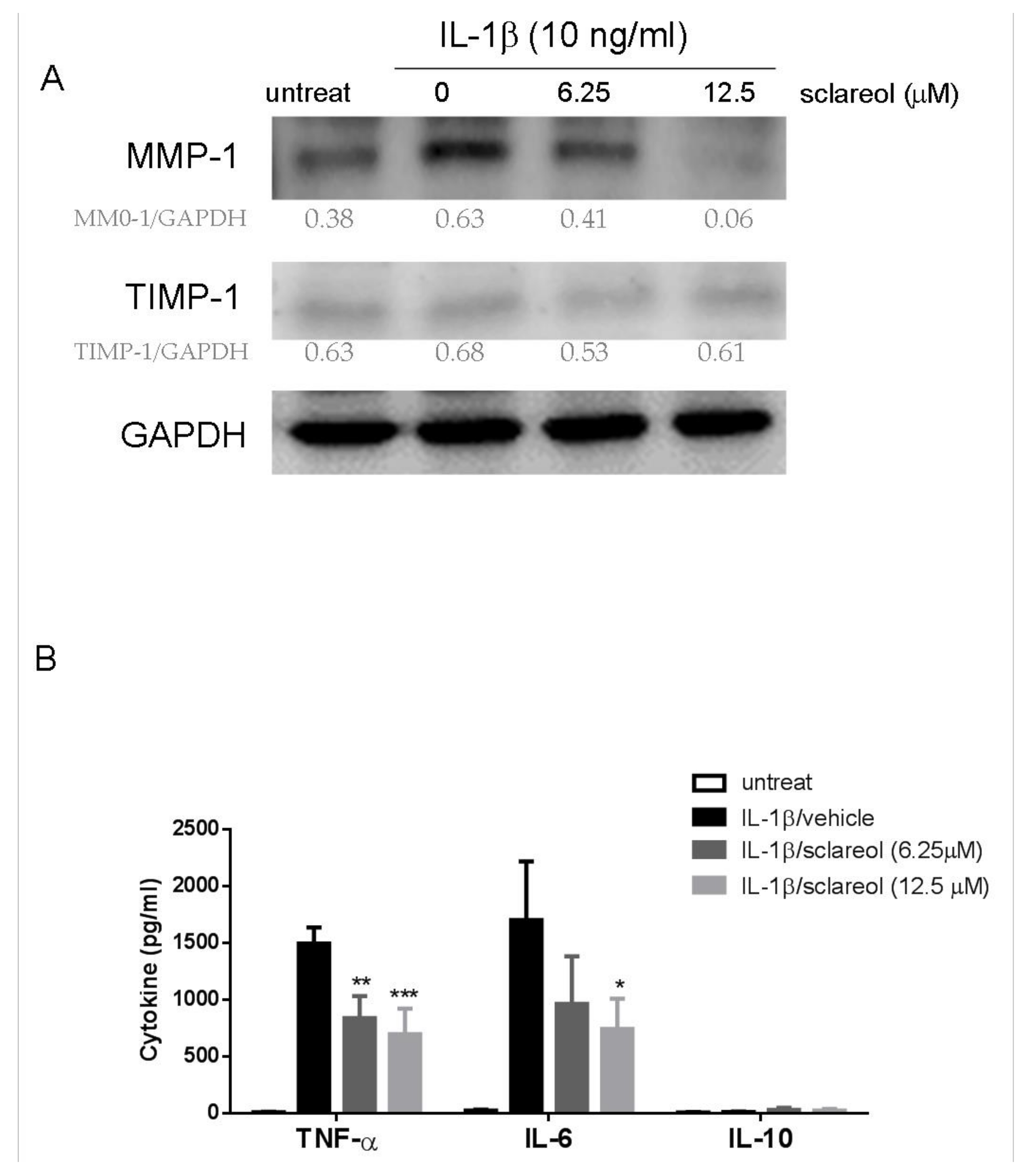
2.5. Sclareol Downregulated Interleukin-1β-Induced Expression of Matrix Metalloproteinases and Proinflammatory Cytokines in Synovial Cells
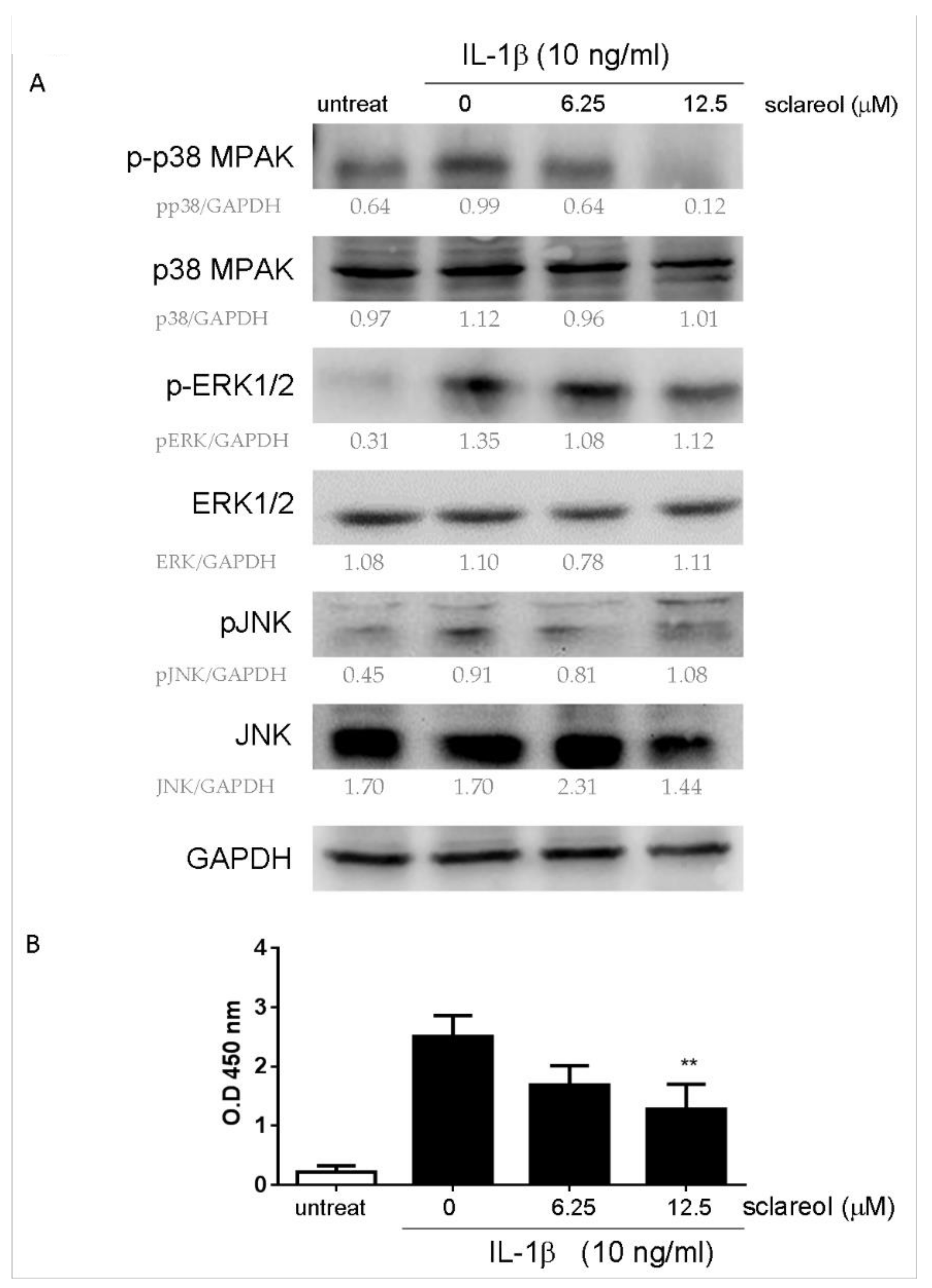
2.6. Sclareol Modulates IL-1β-Induced MAPK and NF-κB Pathways in SW982 Cells
3. Discussion
4. Material and Methods
4.1. Animal Experiments
4.2. Histological Analysis
4.3. ELISA for Serum Anti-Mouse Collagen II Antibodies (Anti-CII Abs)
4.4. Flow Cytometry
4.5. Cell Culture
4.6. Cell Counting Kit-8 (CCK8) Assay
4.7. Western Blot Analysis
4.8. NF-κB Activity Assay
4.9. Measurement of the Cytokine Concentrations in Cell Culture
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bustamante, M.F.; Garcia-Carbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 110. [Google Scholar] [CrossRef] [PubMed]
- Bartok, B.; Firestein, G.S. Fibroblast-like synoviocytes: Key effector cells in rheumatoid arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef] [PubMed]
- Bresnihan, B. Pathogenesis of joint damage in rheumatoid arthritis. J. Rheumatol. 1999, 26, 717–719. [Google Scholar] [PubMed]
- Chang, S.K.; Gu, Z.; Brenner, M.B. Fibroblast-like synoviocytes in inflammatory arthritis pathology: The emerging role of cadherin-11. Immunol. Rev. 2010, 233, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, R.; Rasool, M. Fibroblast-like synoviocytes-dependent effector molecules as a critical mediator for rheumatoid arthritis: Current status and future directions. Int. Rev. Immunol. 2017, 36, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hashimoto, T.; Sakai, Y.; Hashiramoto, A. Involvement of the circadian rhythm and inflammatory cytokines in the pathogenesis of rheumatoid arthritis. J. Immunol. Res. 2014, 2014, 282495. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.S.; Nielsen, M.T.; Severson, R.F. Biorationals from Nicotiana protect cucumbers against Colletotrichum lagenarium (Pass.) ell. & halst disease development. J. Chem. Ecol. 1995, 21, 221–231. [Google Scholar] [PubMed]
- Sun, Y.J.; Gao, M.L.; Zhang, Y.L.; Wang, J.M.; Wu, Y.; Wang, Y.; Liu, T. Labdane Diterpenes from the Fruits of Sinopodophyllum emodi. Molecules 2016, 21, 434. [Google Scholar] [CrossRef] [PubMed]
- Tran, Q.T.N.; Wong, W.S.F.; Chai, C.L.L. Labdane diterpenoids as potential anti-inflammatory agents. Pharmacol. Res. 2017, 124, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Pal, M.; Sharma, R.P. Biological activity of the labdane diterpenes. Planta Med. 1999, 65, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.J.; Pan, C.H.; Wu, C.H. Sclareol exhibits anti-inflammatory activity in both lipopolysaccharide-stimulated macrophages and the lambda-carrageenan-induced paw edema model. J. Nat. Prod. 2012, 75, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.H.; Deng, J.S.; Pan, H.P.; Liao, J.C.; Huang, S.S.; Huang, G.J. Sclareol ameliorate lipopolysaccharide-induced acute lung injury through inhibition of MAPK and induction of HO-1 signaling. Int. Immunopharmacol. 2017, 44, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Huang, Y.; Santoso, M.B.; Wu, L.D. Sclareol exerts anti-osteoarthritic activities in interleukin-1beta-induced rabbit chondrocytes and a rabbit osteoarthritis model. Int. J. Clin. Exp. Pathol. 2015, 8, 2365–2374. [Google Scholar] [PubMed]
- Cho, Y.G.; Cho, M.L.; Min, S.Y.; Kim, H.Y. Type II collagen autoimmunity in a mouse model of human rheumatoid arthritis. Autoimmun. Rev. 2007, 7, 65–70. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Astry, B.; Venkatesha, S.H.; Moudgil, K.D. Involvement of the IL-23/IL-17 axis and the Th17/Treg balance in the pathogenesis and control of autoimmune arthritis. Cytokine 2015, 74, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Mauri, C.; Williams, R.O.; Walmsley, M.; Feldmann, M. Relationship between Th1/Th2 cytokine patterns and the arthritogenic response in collagen-induced arthritis. Eur. J. Immunol. 1996, 26, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lee, W.S.; Lee, E.G.; Sung, M.S.; Yoo, W.H. Sulforaphane inhibits IL-1beta-induced proliferation of rheumatoid arthritis synovial fibroblasts and the production of MMPs, COX-2, and PGE2. Inflammation 2014, 37, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yokoyama, T.; Akatsu, H.; Tukiyama, T.; Tokiwa, T. Phenotypic characterization of a human synovial sarcoma cell line, SW982, and its response to dexamethasone. In Vitro Cell. Dev. Biol. Anim. 2003, 39, 337–339. [Google Scholar] [CrossRef]
- Ou, M.C.; Hsu, T.F.; Lai, A.C.; Lin, Y.T.; Lin, C.C. Pain relief assessment by aromatic essential oil massage on outpatients with primary dysmenorrhea: A randomized, double-blind clinical trial. J. Obstet. Gynaecol. Res. 2012, 38, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Caissard, J.C.; Olivier, T.; Delbecque, C.; Palle, S.; Garry, P.P.; Audran, A.; Valot, N.; Moja, S.; Nicole, F.; Magnard, J.L.; et al. Extracellular localization of the diterpene sclareol in clary sage (Salvia sclarea L., Lamiaceae). PLoS ONE 2012, 7, e48253. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Kwon, J.E.; Lee, S.Y.; Lee, E.J.; Kim, D.S.; Moon, S.J.; Lee, J.; Kwok, S.K.; Park, S.H.; Cho, M.L. IL-17-mediated mitochondrial dysfunction impairs apoptosis in rheumatoid arthritis synovial fibroblasts through activation of autophagy. Cell Death Dis. 2017, 8, e2565. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Kim, H.R.; Kim, B.M.; Cho, M.L.; Lee, S.H. Th17 cytokines regulate osteoclastogenesis in rheumatoid arthritis. Am. J. Pathol. 2015, 185, 3011–3024. [Google Scholar] [CrossRef] [PubMed]
- Fazaa, A.; Ben Abdelghani, K.; Abdeladhim, M.; Laatar, A.; Ben Ahmed, M.; Zakraoui, L. The level of interleukin-17 in serum is linked to synovial hypervascularisation in rheumatoid arthritis. Jt. Bone Spine 2014, 81, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.; Hassan, Z.M.; Salehian, O. Sclareol reduces CD4+ CD25+ FoxP3+ Treg cells in a breast cancer model in vivo. Iran. J. Immunol. 2013, 10, 10–21. [Google Scholar] [PubMed]
- Jeong, J.W.; Lee, H.H.; Lee, K.W.; Kim, K.Y.; Kim, S.G.; Hong, S.H.; Kim, G.Y.; Park, C.; Kim, H.K.; Choi, Y.W.; et al. Mori folium inhibits interleukin-1beta-induced expression of matrix metalloproteinases and inflammatory mediators by suppressing the activation of NF-kappaB and p38 MAPK in SW1353 human chondrocytes. Int. J. Mol. Med. 2016, 37, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Luo, J.; Wen, H.; Zhang, T.; Zuo, X.; Li, X. MDM2 promotes rheumatoid arthritis via activation of MAPK and NF-kappaB. Int. Immunopharmacol. 2016, 30, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Herrington, F.D.; Carmody, R.J.; Goodyear, C.S. Modulation of NF-kappaB Signaling as a Therapeutic Target in Autoimmunity. J. Biomol. Screen 2016, 21, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, R.K.; El-Ansary, A.K.; El-Eishi, H.H.; Kamal, H.M.; El-Saeed, N.H. Matrix metalloproteinases MMP-3 and MMP-1 levels in sera and synovial fluids in patients with rheumatoid arthritis and osteoarthritis. Ital. J. Biochem. 2005, 54, 248–257. [Google Scholar] [PubMed]
- Li, Y.R.; Chen, D.Y.; Chu, C.L.; Li, S.; Chen, Y.K.; Wu, C.L.; Lin, C.C. Naringenin inhibits dendritic cell maturation and has therapeutic effects in a murine model of collagen-induced arthritis. J. Nutr. Biochem. 2015, 26, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- McCann, F.E.; Perocheau, D.P.; Ruspi, G.; Blazek, K.; Davies, M.L.; Feldmann, M.; Dean, J.L.; Stoop, A.A.; Williams, R.O. Selective tumor necrosis factor receptor I blockade is antiinflammatory and reveals immunoregulatory role of tumor necrosis factor receptor II in collagen-induced arthritis. Arthritis Rheumatol. 2014, 66, 2728–2738. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, S.-W.; Hsieh, M.-C.; Li, S.; Lin, S.-C.; Wang, S.-P.; Lehman, C.W.; Lien, C.Z.; Lin, C.-C. Therapeutic Potential of Sclareol in Experimental Models of Rheumatoid Arthritis. Int. J. Mol. Sci. 2018, 19, 1351. https://doi.org/10.3390/ijms19051351
Tsai S-W, Hsieh M-C, Li S, Lin S-C, Wang S-P, Lehman CW, Lien CZ, Lin C-C. Therapeutic Potential of Sclareol in Experimental Models of Rheumatoid Arthritis. International Journal of Molecular Sciences. 2018; 19(5):1351. https://doi.org/10.3390/ijms19051351
Chicago/Turabian StyleTsai, Sen-Wei, Ming-Chia Hsieh, Shiming Li, Shih-Chao Lin, Shun-Ping Wang, Caitlin W. Lehman, Christopher Z. Lien, and Chi-Chien Lin. 2018. "Therapeutic Potential of Sclareol in Experimental Models of Rheumatoid Arthritis" International Journal of Molecular Sciences 19, no. 5: 1351. https://doi.org/10.3390/ijms19051351