Molecular Characterization and the Function of Argonaute3 in RNAi Pathway of Plutella xylostella


1
State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, College of Life Science, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2
Joint International Research Laboratory of Ecological Pest Control, Ministry of Education, Fuzhou 350002, China
3
Key Laboratory of Integrated Pest Management for Fujian-Taiwan Crops, Ministry of Agriculture, Fuzhou 350002, China
4
Key Laboratory of Green Pest Control (Fujian Agriculture and Forestry University), Fujian Province University, Fuzhou 350002, China
5
Key Laboratory of Natural Pesticides and Chemical Biology of the Ministry of Education, South China Agricultural University, Guangzhou 510642, China
6
Department of Biological Sciences, Brock University, 500 Glenridge Avenue, St. Catharines, ON L2S 3A1, Canada
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(4), 1249; https://doi.org/10.3390/ijms19041249
Submission received: 9 March 2018
/
Revised: 14 April 2018
/
Accepted: 17 April 2018
/
Published: 20 April 2018
(This article belongs to the Special Issue Molecular Entomology of Insects of Economic Importance)
Abstract
:Argonaute (Ago) protein family plays a key role in the RNA interference (RNAi) process in different insects including Lepidopteran. However, the role of Ago proteins in the RNAi pathway of Plutella xylostella is still unknown. We cloned an Argonaute3 gene in P. xylostella (PxAgo3) with the complete coding sequence of 2832 bp. The encoded protein had 935 amino acids with an expected molecular weight of 108.9 kDa and an isoelectric point of 9.29. It contained a PAZ (PIWI/Argonaute/Zwile) domain and PIWI (P-element-induced whimpy testes) domain. PxAgo3 was classified into the Piwi subfamily of Ago proteins with a high similarity of 93.0% with Bombyx mori Ago3 (BmAgo3). The suppression of PxAgo3 by dsPxAgo3 was observed 3 h after treatment and was maintained until 24 h. Knockdown of PxAgo3 decreased the suppression level of PxActin by dsPxActin in P. xylostella cells, while overexpression of PxAgo3 increased the RNAi efficiency. Our results suggest that PxAgo3 play a key role in the double stranded RNA (dsRNA)-regulated RNAi pathway in P. xylostella.
1. Introduction
RNA interference (RNAi), also called double stranded RNA (dsRNA)-induced post-transcriptional gene silencing, is a cellular mechanism regulated by a unique group of RNA-binding proteins named Argonautes (Ago) [1,2]. Ago proteins bind to small administrative RNAs, such as small interfering RNA (siRNA), micro RNA (microRNA), and Piwi-interacting RNA (piRNA), to generate the RNA-induced silencing complex (RISC) [3]. Each Ago protein consists of two fundamental domains, PAZ (PIWI/Argonaute/Zwile) and PIWI (P-element-induced wimpy testes), in their structure [4]. Ago protein structures of eukaryotes and prokaryotes share a bilobed architectural structure containing four domains of N, PAZ, MID (middle) and, PIWI connected by two linkers [5]. The N domain is linked with a small RNA duplex loosening up during the RISC formation [6]. The PAZ domain contains an oligonucleotide-binding fold at the 3′-end of RNA and can bind to siRNAs [7]. The MID domain generates an associating pocket for fixing 5′-phosphate of the small RNAs [8], while PIWI domain is orthologous to ribonuclease H (RNase H) and encloses the slicing residues Asp-Asp-His (DDH), which is significant for the endonucleolytic “cleavage” action of the RISC [9].
The number of Ago proteins varies among different insect species. For example, two Ago proteins have been reported in Anopheles gambiae [10], three in Leptinotarsa decemlineata [11], four in Bombyx mori [12], and five in Drosophila melanogaster [13]. Ago proteins are divided into three groups of Piwi subfamily, Ago subfamily, and worm-specific Argonautes (WAGO) [14]. In Penaeus monodon, PmAgo3 is found within the Ago subfamily [9], while in D. melanogaster [15] and B. mori [16], the Ago3 protein is placed in the Piwi subfamily. Ago3 protein is related to the piRNA-mediated pathway and silences transposable elements (TEs) in D. melanogaster [17]. The sense piRNAs bind to Ago3 to cleave the primary piRNA precursors [18]. Ago3 protein is located in the cytoplasm of D. melanogaster germ cells, where it works in the Ping-Pong cycle to destroy transposon mRNAs [19]. In humans, Ago3 cleaves in a similar pattern as Ago2 protein [20], and up-regulates the ratio between passenger strand and guide strand of miRNA let-7a [21]. In Tribolium castaneum, Ago3 is homologous with DmAgo3 and is involved in transcriptional silencing mechanism [22]. In D. melanogaster ovary, transposon elements are suppressed by Ago3 protein [23]. DmAgo3 knockdown in embryos shows an increase in both telomeric transposition and telomere length [24]. Ago3 is also recognized as a slicer protein and, in insects, it is involved in piRNA-mediated gene silencing [25]. The overexpression of Ago3 increases the RNAi efficiency in Lepidopteran cell lines [26], Caenorhabditis elegans [14], and H. sapiens [21].
Plutella xylostella (diamondback moth) is a worldwide pest of cruciferous plants that has become a model species due to its high reproductive rate, fast dispersal, and capacity to develop resistance to most classes of pesticides [27,28]. Previous studies have shown the phenomenon of RNAi in P. xylostella larvae [29]. Here, we identified and described the Ago3 gene in P. xylostella (PxAgo3) and its function in the RNAi pathway of P. xylostella. The expression of PxAgo3 in response to dsPxAgo3 challenges were investigated in P. xylostella larvae. We then evaluated the effects of knockdown or overexpression of PxAgo3 on the suppression of targeting gene, PxActin by dsPxActin in P. xylostella cells. Our results suggest that PxAgo3 plays a significant role in the RNAi pathway of P. xylostella.
2. Results
2.1. Identification and Sequence Analysis of PxAgo3 Gene
Using Ago3 CDS sequence from B. mori (BmAgo3, GenBank: AB372007.1), we identified Px011435 as the Ago3 coding sequence in P. xylostella genome. Using the specific primers of PxAgo3-F and PxAgo3-R (Table 1) based on Px011435, PxAgo3 was amplified through PCR and sequenced. The sequence was deposited in the GenBank with the accession number of MG778697. The sequence of PxAgo3 was of 2832 bp with the predicted protein of 943 aa (Figure 1B). The cloned sequence contained two conserved regions with the PAZ domain of 420 bp in length and the Piwi domain of 1335 bp from 3′ end to 5′ end.
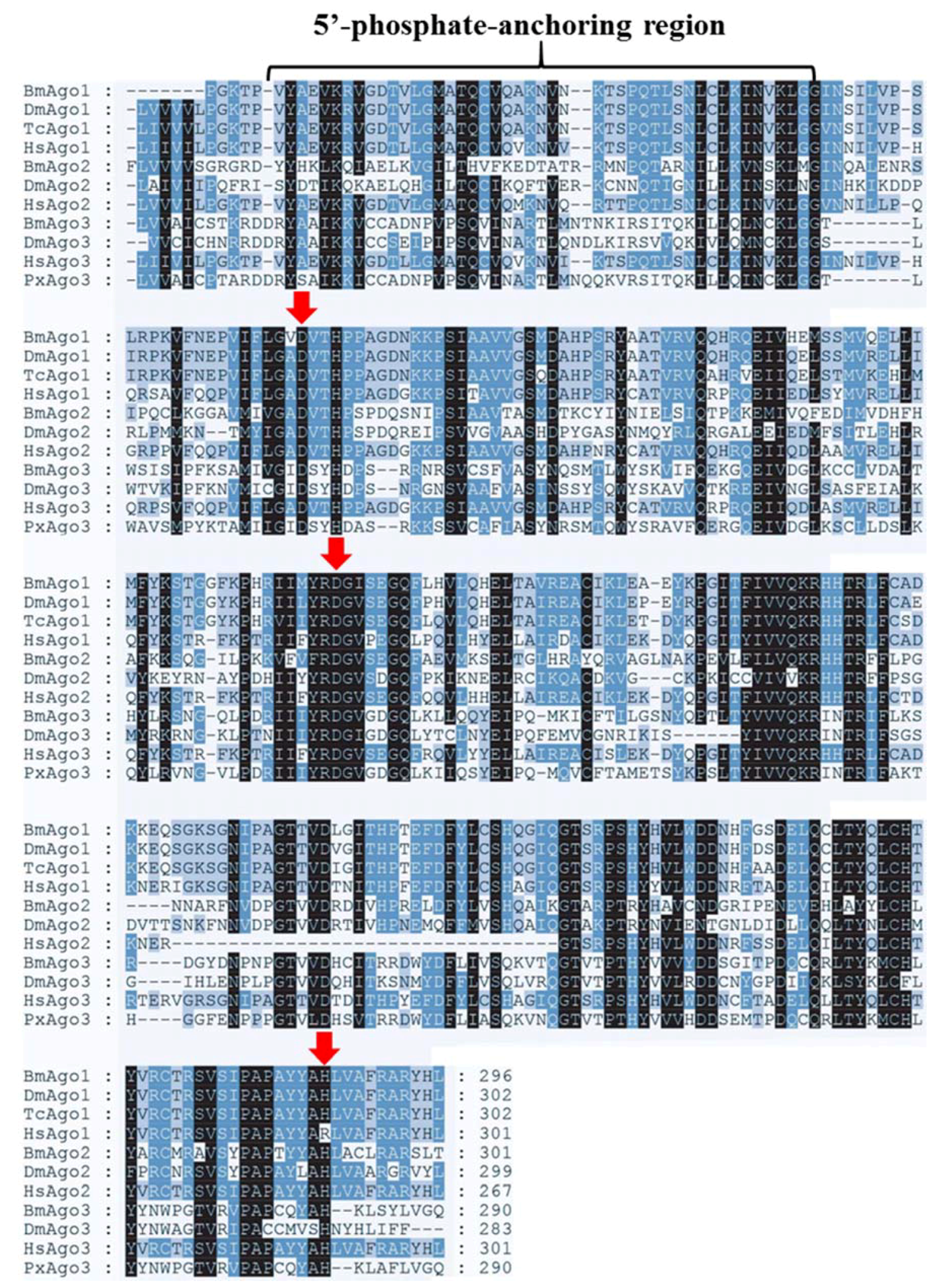
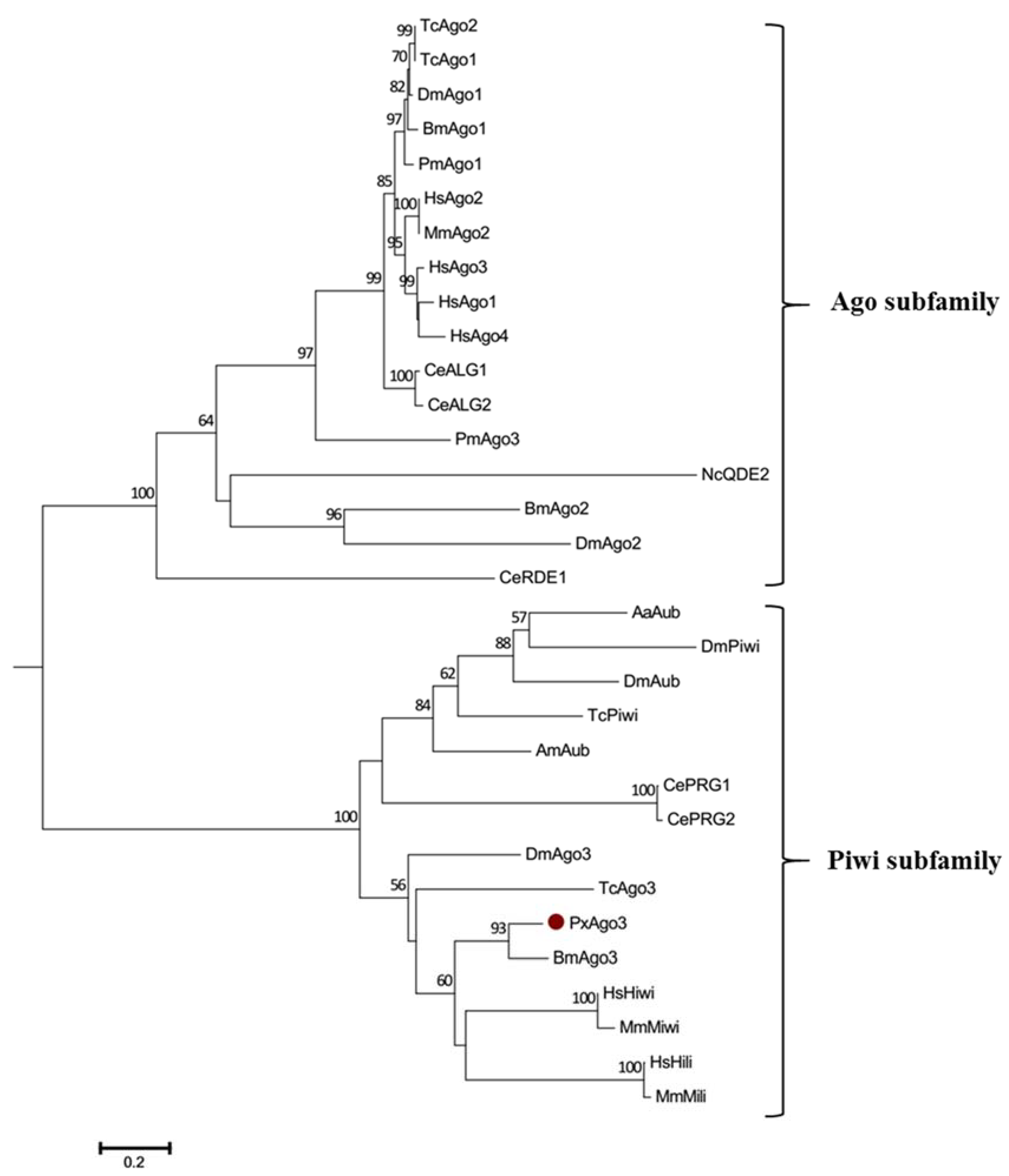
GenBank database sequence comparison showed that PxAgo3 was similar to B. mori Piwi-like Ago3 protein with the similarity of 49.3%. To analyze the functional sequence, the deduced amino acid sequence of PxAgo3 and some expressive Ago family protein sequences were aligned based on the conserved PIWI domain. The multiple-sequence alignment led to the identification of one 5′-phosphate-anchoring region and three catalytic residues (DDH positioned by red arrows, Figure 2) inside the PIWI domain. Thirty-one Ago proteins sequences from ten different species were collected through the NCBI website to construct the phylogenetic tree (Table 2). The phylogenetic tree showed that the Ago proteins were divided into two subfamilies, Ago- and Piwi, with PxAgo3 in the Piwi subfamily located with BmAgo3 (Figure 3).
2.2. PxAgo3 Expression in Response to dsRNA in P. xylostella Larvae
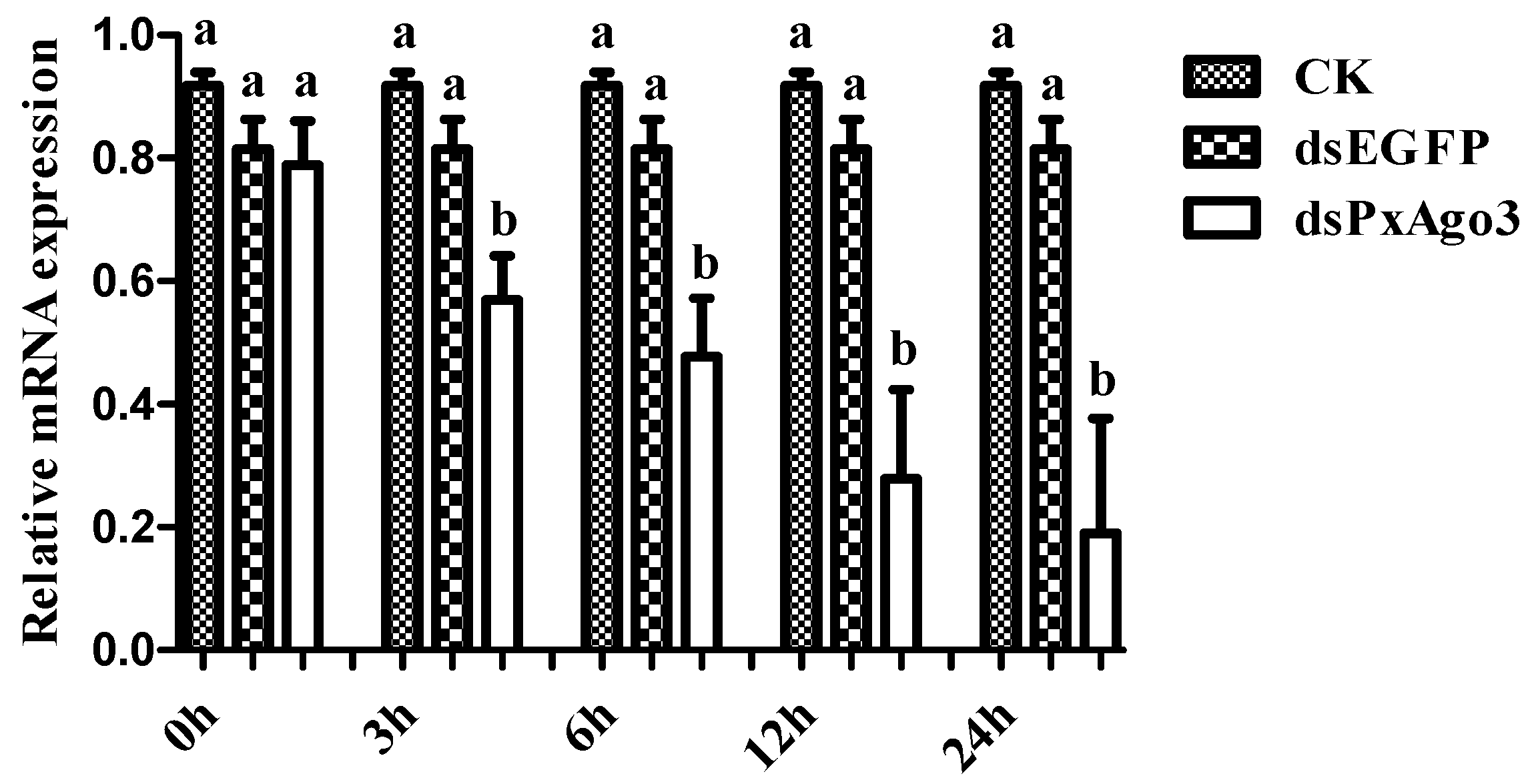
The expression of PxAgo3 was significantly more suppressed by dsPxAgo3 in the 3rd instar larvae of P. xylostella than by CK and dsEGFP. The suppression was first observed in 3 h after treatment and maintained up to 24 h (Figure 4).
2.3. PxAgo3 Suppression Decreased the RNAi Efficiency in P. xylostella Cells
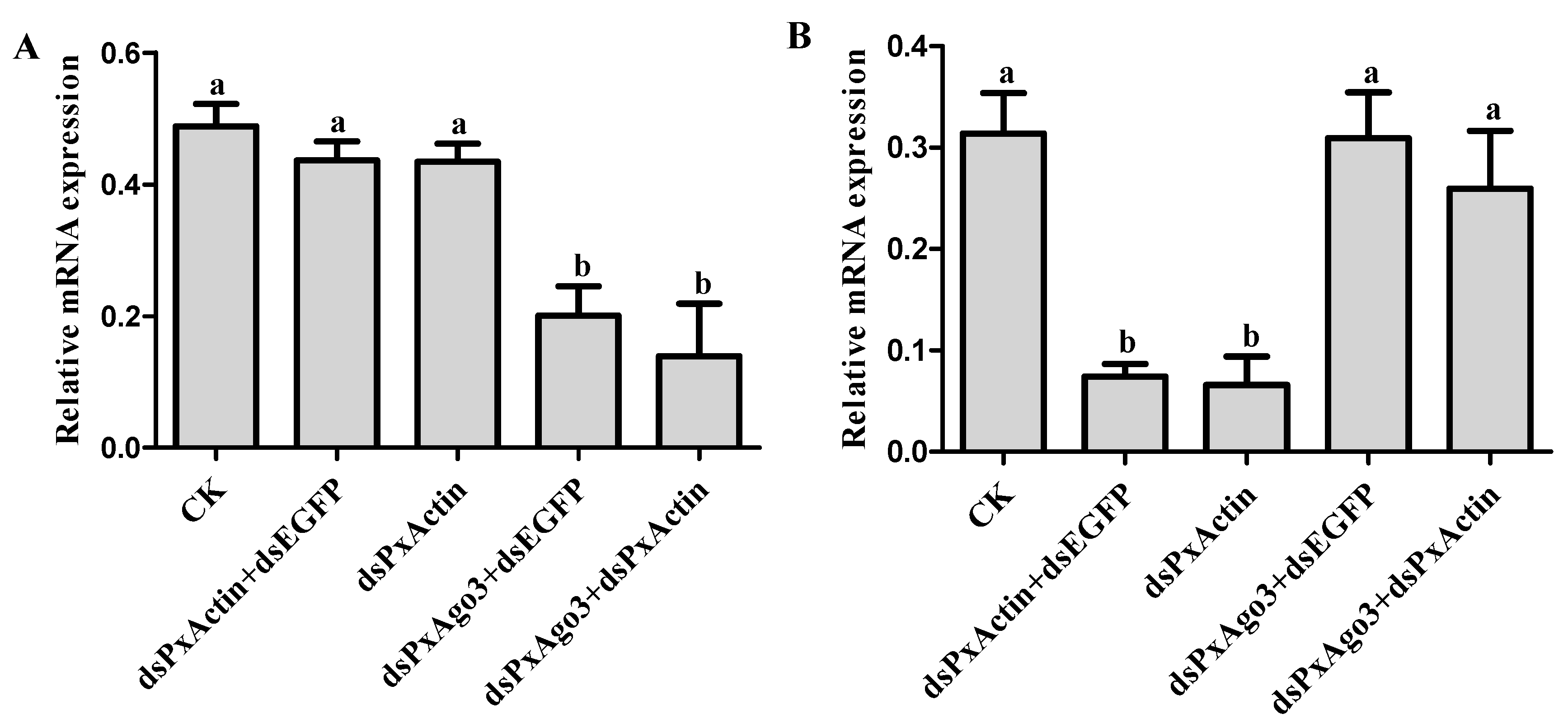
We hypothesized that the knockdown of PxAgo3 by dsPxAgo3 would affect the suppression of endogenous gene PxActin by dsPxActin. Silencing of PxAgo3 by dsPxAgo3 led to a significant down-regulation (F = 11.34, df1 = 4, df2 = 10, p = 0.0010) (Figure 5A). For RNAi of RNAi, P. xylostella cells were exposed to dsPxAgo3 for 24 h, followed by exposure of these cells to dsPxActin for another 24 h. PxActin expression was significantly suppressed by dsPxActin and dsPxActin+dsEGFP, but not by dsPxAgo3+dsPxActin (F = 9.91, df1 = 4, df2 = 10, p = 0.0017) (Figure 5B). These results demonstrated that the dsRNA-mediated suppression of PxAgo3 reduced the RNAi efficiency in P. xylostella cells.
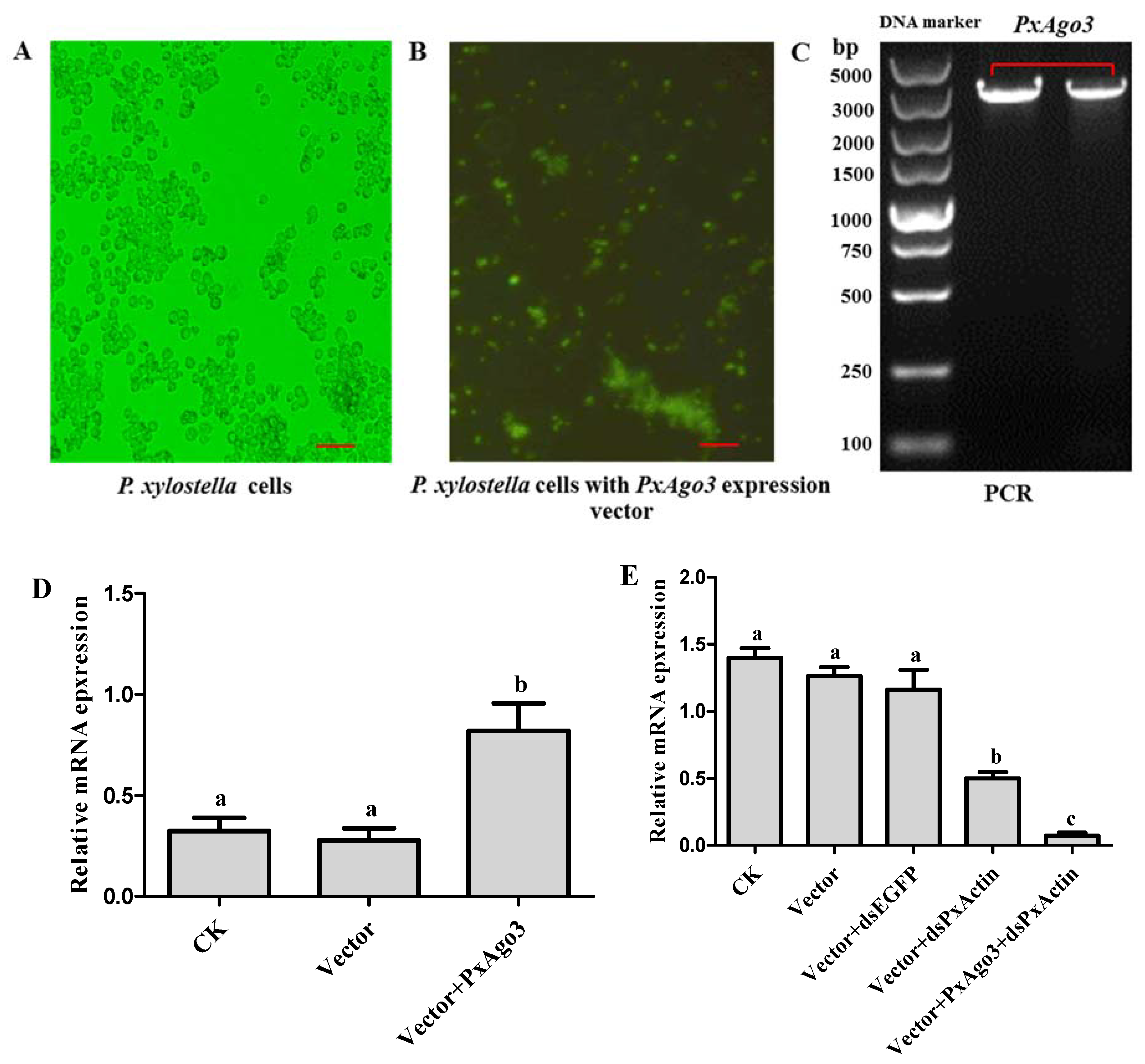
2.4. PxAgo3 Overexpression Increased the RNAi Efficiency in P. xylostella Cells
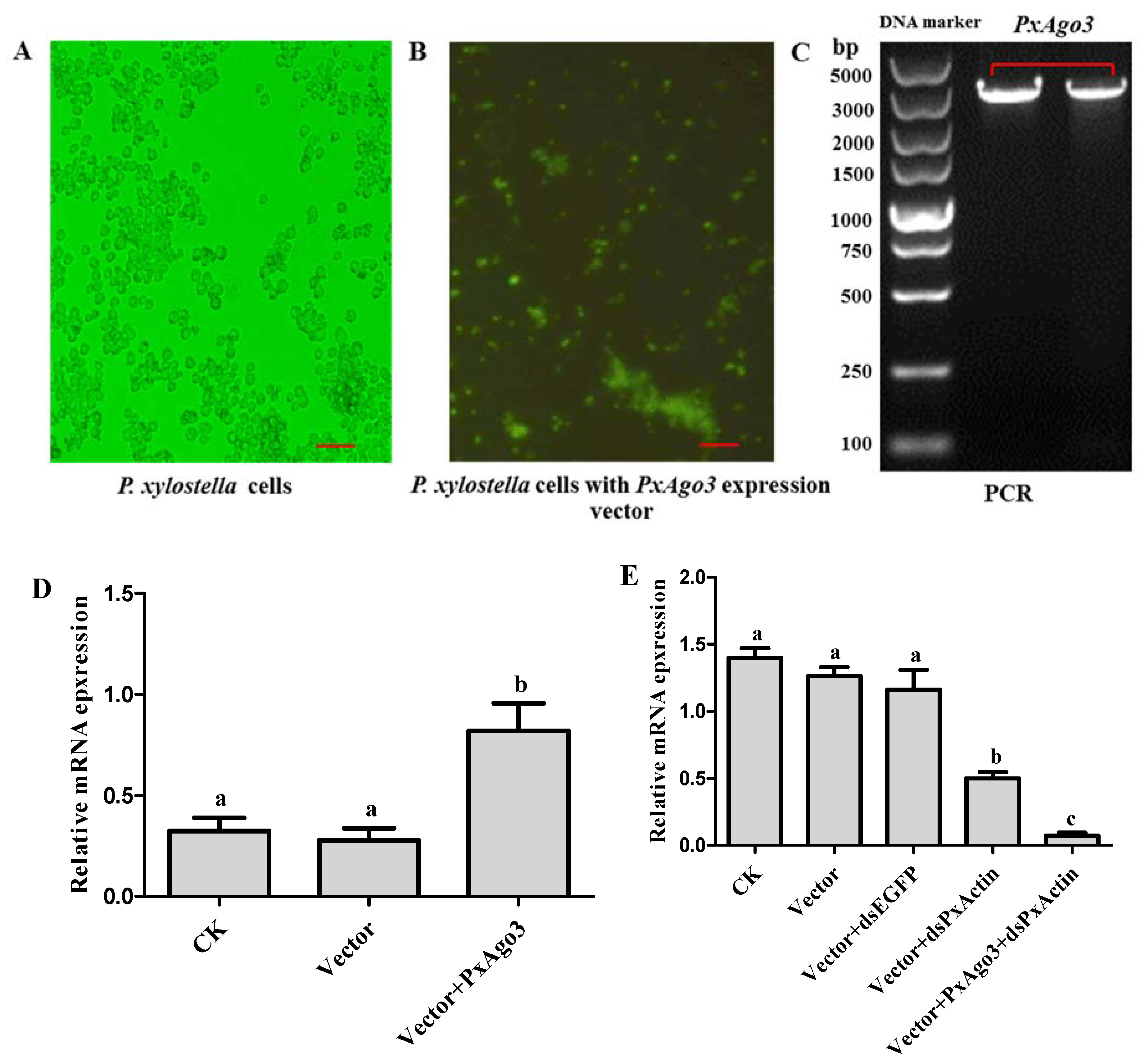
To overexpress PxAgo3 in P. xylostella cells, the expression construct was inserted into the PIZT/V5 vector and transfected into P. xylostella cells. The success of transfection was verified by the enhanced green fluorescence protein (EGFP) expression with the green fluorescence observed under the florescence microscope (Figure 6A,B) and by PCR using vector-based primers (Figure 6C). The expression level of PxAgo3 in P. xylostella cells significantly increased in transfected P. xylostella cells (F = 10.45, df1 = 2, df2 = 6, p = 0.0111) (Figure 6D). To observe the RNAi efficiency in PxAgo3-overexpressed cells, PxActin was targeted to suppress in PxAgo3-overexpressed cells. The PxActin expression was affected by the treatments (F = 45.95, df1 = 4, df2 = 10, p < 0.0001). PxActin expression was reduced by dsPxActin in normal cells and the suppression level was increased in PxAgo3-overexpressing cells (Figure 6E). These results suggested that overexpression of PxAgo3 increased the dsRNA-regulated RNAi efficiency in P. xylostella cells.
3. Discussion
In this study, we have cloned and identified a new Ago gene from P. xylostella, PxAgo3. The amino acid sequence of PxAgo3 included a PAZ domain and a PIWI domain. PAZ and PIWI are similar in different species, such as P. monodon [9], B. mori [16], D. melanogaster [15]. The PAZ domain is a particular nucleic acid binding site, which plays a significant role in RNAi mechanism [7]. Ago3 is physically bound with nucleic acid through the PAZ domain, while the PIWI domain interacts with the 5′ end of the guide RNA in Archaeoglobus fulgidus [30]. The three catalytic residues (DDH) within the PIWI domain of Ago3 form a cleavage triad in DmAgo3 protein [31], in BmAgo3 protein [32], and in PmAgo3 protein [9], and was also observed in PxAgo3.
The phylogenetic tree showed that PxAgo3 protein belongs to the Piwi subfamily. Ago3 has been reported to belong to the Piwi subfamily in B. mori [16,32,33], D. melanogaster [22], and Aedes aegypti [34]. however, Ago3 belongs to the Ago subfamily in C. elegans [14,35] and H. sapiens [36,37].
The silencing of PxAgo3 occurred 3 h after dsRNA treatment and was maintained for 24 h. Ago3 knockdown is observed 24 h after introduction of dsRNA in mammalian cells [38] as well as A. aegypti, and A. albopictus cells [39]. In Culex quinquefasciatus, Ago3 suppression is only observed 48 h after treatment [40]. Therefore, it is suggested that Ago3 can be effectively suppressed by dsRNA in animals.
PxAgo3 knockdown reduced RNAi efficiency in P. xylostella cells. Ago3 is involved in dsRNA-regulated RNAi phenomenon [22] and reduces RNAi response in D. melanogaster [41] and B. mori [42]. Ago2 has been considered the only cleavage component in siRNA-regulated RNAi process in insects [42,43]. Our results, however, suggest that Ago3 may be involved in the siRNA-regulated RNAi pathway, which is known to be associated with the piRNA pathway. The overexpression of PxAgo3 also enhanced the RNAi response. The overexpression of Ago3 in Lepidopteran cell lines [26], C. elegans [14], as well as HEK293 and HeLa cells [21] can increase the RNAi efficiency. It is possible that PxAgo3 overexpression may facilitate the dsRNA-mediated RNAi in P. xylostella.
In conclusion, we sequenced PxAgo3 and identified PxAgo3 as a Piwi subfamily member containing the cleavage motif of DDH in the PIWI domain. We discovered that Ago3 is involved in the siRNA-mediated pathway in P. xylostella by decreasing PxActin suppression by dsPxActin through knockdown of PxAgo3 and enhancing PxActin suppression by dsPxActin through overexpression of PxAgo3 in P. xylostella cells.
4. Materials and Methods
4.1. P. xylostella Fuzhou-S Rearing and Cell Culture
The P. xylostella individuals used in this study came from an insecticide-susceptible strain previously used for P. xylostella genomic sequencing [44]. P. xylostella larvae were reared on radish seedlings at 25 ± 1 °C, 65 ± 5% RH and 16 h L: 8 h D. A stable P. xylostella cell line was used, which was established from embryonic tissues of P. xylostella [45]. Several generations of P. xylostella cells were maintained at 27 °C in Grace’s medium (Invitrogen, Waltham, MA, USA) supplemented with 10% of fetal bovine serum (FBS).
4.2. Total RNAs Purification and cDNA Preparation
Total RNAs were purified from four 3rd instar larvae of P. xylostella using the TRIzol-Reagent (Invitrogen). The quantity of RNAs were measured using a Nanodrop ND-1000 spectrophotometer (Invitrogen), and the RNA quality was verified through 1% gel electrophoresis. Total RNAs were diluted with nuclease-free water (Promega, Madison, WI, USA) and maintained at a concentration of 500 ng/μL. Furthermore, 1 μL of purified RNA was used as the template for cDNA preparation by using the GoTaq® 2-Step RT-qPCR System (Promega).
4.3. Molecular Cloning and Expression Vector Construction of PxAgo3
For PxAgo3 cloning, a specific pair of primers (PxAgo3-F and PxAgo3-R) (Table 1) were designed using the Primer Premier 5 according to the gene sequence of PxAgo3 (gene ID: Px011435) from P. xylostella genome (Available online: http://iae.fafu.edu.cn/DBM/index.php). The Phanta Max Super-Fidelity DNA Polymerase (Vazyme, Nanjing, China) was used to conduct PCR following the manufacture protocol and conditions: denaturation at 95 °C for 3 min, amplification by 30 cycles at 95 °C for 15 s, 56 °C for 15 s, and 72 °C for 3 min, the last expansion at 72 °C for 10 min and then storage at 4 °C. The PCR product was extracted from the gel by using the QIAquick Gel extraction kit (Qiagen, Venlo, The Netherlands) according to the manufacturer protocol. The extracted PCR product and PJET vector were ligated using T4 DNA ligase enzyme (Promega) according to the ClonJET PCR Cloning kit (Invitrogen) and incubated at 25 °C for 1 h. The ligated sample was mixed with 50 μL high efficiency competent cells of DH-5α (Tiangen, Beijing, China) and incubated on ice for 30 min. The sample was cultured in the selection media to obtain white colonies. The colony PCR was done using the Go Taq® Green Master Mix enzyme (Promega) with the vector-based primers (Table 1). The PCR product was verified through 1% gel electrophoresis following the procedure previously described. Eight white colonies were checked and one verified positive white colony was cultured. An amount of 500 µL of the cultured solution was sent to General Biosystems (China) for sequencing. The protein sequence of PxAgo3 was blasted in the National Center for Biotechnology Information (NCBI) (Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGETYPE=BlastSearch&LINK_LOC=blasthome) to verify its identity and submitted to GenBank (Available online: https://submit.ncbi.nlm.nih.gov/subs/genbank/) for accession number.
To construct the PxAgo3 expression vector (PIZT/V5-His+PxAgo3), the PCR product and PIZT/V5-His were individually double-digested at 37 °C using two restriction enzymes of FastDigest KpnI and EcoRI (Invitrogen). Both double digested products (PCR product and PIZT/V5-His) were ligated and transfected into competent cells and checked by colony PCR and 1% gel electrophoresis (as previously described). A positive white colony cultured solution of PxAgo3 expression vector was extracted using the Hispeed Plasmid Midi kit (Qiagen).
4.4. Phylogenetic Tree and Sequence Analysis
Ago protein sequences were downloaded from the GenBank database of NCBI (Available online: https://www.ncbi.nlm.nih.gov/) and are listed in Table 2. A multiple-sequence alignment was performed based on the PIWI domain using the ClustalW software. The phylogenetic tree was constructed using MEGA 6.0 based on the neighbor-joining method with a bootstrap value of 1000 replicates. The isoelectric point and molecular weight of PxAgo3 were identified using the Expasy website (Available online: http://www.expasy.org).
4.5. Double-Stranded RNA (dsRNA) Preparation
The following gene-specific sequences were used as the dsRNA templates: 389-bp fragment targeting PxAgo3, 461-bp fragment targeting PxActin, and 457-bp fragment targeting EGFP. DsRNAs were prepared using the Ambion MEGAscript RNAi kit (Invitrogen). The primers listed in Table 1 were used to amplify these fragments with the T7-polymerase promoter (TAATACGACTCACTATAGGG) sequence. The reaction mixture was incubated at 37 °C for 3 h, heat shocked at 75 °C for 5 min and then kept at room temperature for 1 h for the annealing reaction. Subsequently, dsRNAs were treated with DNase I to remove the template DNA, purified with the phenol/chloroform method and finally diluted in nuclease-free water. The dsRNA quality and quantity were measured following the procedure previously described.
4.6. DsRNA Injection and Transfection into P. xylostella Larvae and Cells
For dsRNA injection, 3rd instar larvae of P. xylostella were selected and borosilicate micro-capillaries (Glass Replacement 1.1) were pulled using heater setting 64 in PC-10 (Narishige, Tokyo, Japan). We injected 10 μL dsRNAs (1 μg) of PxAgo3, PxActin and EGFP (control) by using micro-injection world precision instruments (Sarasota, FL, USA) into the hemolymph of 3rd instar larvae (n = 60) individually. After injection, larvae were transferred into separate plastic rearing box and fed with fresh radish leaves. Purified dsRNAs were transfected into the P. xylostella cells using the Cellfectin® II reagent (Promega) according to the manufacturer’s instructions.
4.7. RNAi of RNAi Assay in P. xylostella Cells
For RNAi of RNAi assay, one day before transfection of dsRNAs into cells, P. xylostella cells were cultured in 2 mL of Sf-900 III serum free media (SFM) (Promega) with 10% fetal bovine serum (FBS) (HyClone, Utah, United States) and containing 1% anti-toxin antimycotic (pH 6.2) at a concentration of 6 × 105 cells/mL, in 6-well plates (Corning Incorporated, New York, NY, USA). Cells were incubated at 27 °C overnight or until 50–60% of the cells converged. We transfected two types of dsRNA into P. xylostella cells in order. Firstly, 1 μg of dsPxAgo3 was transfected into P. xylostella cells by following the Cellfectin® (Gibco Invitrogen, Waltham, MA, USA) manufacturer’s instructions. The treated P. xylostella cells were incubated at 27 °C for 24 h in a closed chamber to prevent environmental contamination. Secondly, 1 μg of dsPxActin was transfected using the Cellfectin reagent (described as before) into the same P. xylostella cells 24 h after the first transfection of dsPxAgo3. The transfection of dsEGFP was used as a control. The PxAgo3 expression level was analyzed by using RT-qPCR. Similarly, the overexpression of PxAgo3 was measured after transfection of PIZT/V5-His+PxAgo3 into P. xylostella cells by RT-qPCR. The presence of vectors was examined through green fluorescence by using a fluorescence microscope Olympus IX51 (Olympus Corporation, Tokyo, Japan).
The cells were collected 48 h post transfection in 1.5 mL tubes (Axygen Incorporation, Tewksbury, MA, USA) and centrifuged at 1500× g for 5 min at 4 °C. Total RNA extraction, quality and quantity verifications, and cDNA preparation were performed as previously described. PxAgo3 and PxActin expression levels were monitored through RT-qPCR using the primers listed in Table 1. All procedures were done with three biological replicates.
4.8. RT-qPCR Analysis
RT-qPCR was performed using the GoTaq® qPCR master mix (Promega). RT-qPCR was performed according to the following conditions: at 95 °C for 3 min for denaturation, followed by 40 cycles at 95 °C for 5 s and 60 °C for 20 s. The degree of amplification was determined at the end of each cycle by fluorescence intensity measurements. Quantitative mRNA measurements were done in triplicate and standardized to an internal control of ribosomal protein 32 (RL-32) (reference gene). All of the RT-qPCR primers are listed in Table 1.
4.9. Statistical Analysis
One-way analysis of variance (ANOVA) was done using the GraphPad (version 5.01) (GraphPad Software, La Jolla, San Diego, CA, USA), and then differences between treatments were determined using Tukey test at p < 0.05.
Acknowledgments
The authors would like to thank Wei Chen for excellent technical assistance in RNA extraction and gene cloning. We are grateful to Xiao-Li Ma for providing P. xylsotella cells for the experiments. This work was supported by the Natural Science Foundation of China project 31772237, Fujian Science and Technology Major Projects 2015NZ01010012 and 2018NZ01010013.
Author Contributions
Guang Yang and Muhammad Salman Hameed conceived the study. Muhammad Salman Hameed, Guang Yang and Liette Vasseur wrote the manuscript; Muhammad Salman Hameed performed experiments; Muhammad Salman Hameed and Zhengbing Wang performed statistical analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Dueck, A.; Ziegler, C.; Eichner, A.; Berezikov, E.; Meister, G. microRNAs associated with the different human Argonaute proteins. Nucleic Acids Res. 2012, 40, 9850–9862. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-J.; Liu, J.; Tolia, N.H.; Schneiderman, J.; Smith, S.K.; Martienssen, R.A.; Hannon, G.J.; Joshua-Tor, L. The crystal structure of the Argonaute2 PAZ domain reveals an RNA binding motif in RNAi effector complexes. Nat. Struct. Mol. Biol. 2003, 10, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-J.; Smith, S.K.; Hannon, G.J.; Joshua-Tor, L. Crystal structure of Argonaute and its implications for RISC slicer activity. Science 2004, 305, 1434–1437. [Google Scholar] [CrossRef] [PubMed]
- Kwak, P.B.; Tomari, Y. The N domain of Argonaute drives duplex unwinding during RISC assembly. Nat. Struct. Mol. Biol. 2012, 19, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Lingel, A.; Simon, B.; Izaurralde, E.; Sattler, M. Structure and nucleic-acid binding of the Drosophila Argonaute 2 PAZ domain. Nature 2003, 426, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Boland, A.; Tritschler, F.; Heimstädt, S.; Izaurralde, E.; Weichenrieder, O. Crystal structure and ligand binding of the MID domain of a eukaryotic Argonaute protein. EMBO Rep. 2010, 11, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Phetrungnapha, A.; Ho, T.; Udomkit, A.; Panyim, S.; Ongvarrasopone, C. Molecular cloning and functional characterization of Argonaute-3 gene from Penaeus monodon. Fish Shellfish Immunol. 2013, 35, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Dimopoulos, G.; Zhu, J. Association of microRNAs with Argonaute proteins in the malaria mosquito Anopheles gambiae after blood ingestion. Sci. Rep. 2017, 7, 6493–6506. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-S.; Shukla, J.N.; Gong, Z.J.; Mogilicherla, K.; Palli, S.R. RNA interference in the Colorado potato beetle, Leptinotarsa decemlineata: Identification of key contributors. Insect Biochem. Mol. Biol. 2016, 78, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Masaki, Y.; Tatsuke, T.; Li, Z.; Mon, H.; Xu, J.; Lee, J.; Kusakabe, T. A MC motif in silkworm Argonaute 1 is indispensible for translation repression. Insect Mol. Biol. 2013, 22, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Ishizuka, A.; Siomi, H.; Siomi, M.C. Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev. 2004, 18, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Yigit, E.; Batista, P.J.; Bei, Y.; Pang, K.M.; Chen, C.-C.G.; Tolia, N.H.; Joshua-Tor, L.; Mitani, S.; Simard, M.J.; Mello, C.C. Analysis of the Caenorhabditis elegans Argonaute family reveals that distinct Argonautes act sequentially during RNAi. Cell 2006, 127, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.W.; Rubin, G.M. ARGONAUTE1 is required for efficient RNA interference in Drosophila embryos. Proc. Natl. Acad. Sci. USA 2002, 99, 6889–6894. [Google Scholar] [CrossRef] [PubMed]
- Kawaoka, S.; Minami, K.; Katsuma, S.; Mita, K.; Shimada, T. Developmentally synchronized expression of two Bombyx mori Piwi subfamily genes, SIWI and BmAGO3 in germ-line cells. Biochem. Biophys. Res. Commun. 2008, 367, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Mituyama, T.; Huang, H.; Chen, D.; Siomi, M.C.; Siomi, H. Biogenesis pathways of piRNAs loaded onto AGO3 in the Drosophila testis. RNA 2010, 16, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Gunawardane, L.S.; Saito, K.; Nishida, K.M.; Miyoshi, K.; Kawamura, Y.; Nagami, T.; Siomi, H.; Siomi, M.C. A slicer-mediated mechanism for repeat-associated siRNA 5′end formation in Drosophila. Science 2007, 315, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Rozhkov, N.V.; Hammell, M.; Hannon, G.J. Multiple roles for Piwi in silencing Drosophila transposons. Genes Dev. 2013, 27, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Phan, H.-D.; Busch, F.; Hinckley, S.H.; Brackbill, J.A.; Wysocki, V.H.; Nakanishi, K. Human Argonaute3 has slicer activity. Nucleic Acids Res. 2017, 45, 11867–11877. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Diederichs, S. Argonaute-3 activates the let-7a passenger strand microRNA. RNA Biol. 2013, 10, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Ishizu, H.; Siomi, H.; Siomi, M.C. Biology of PIWI-interacting RNAs: New insights into biogenesis and function inside and outside of germlines. Genes Dev. 2012, 26, 2361–2373. [Google Scholar] [CrossRef] [PubMed]
- Khurana, J.S.; Xu, J.; Weng, Z.; Theurkauf, W.E. Distinct functions for the Drosophila piRNA pathway in genome maintenance and telomere protection. PLoS Genet. 2010, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Vagin, V.V.; Lee, S.; Xu, J.; Ma, S.; Xi, H.; Seitz, H.; Horwich, M.D.; Syrzycka, M.; Honda, B.M. Collapse of germline piRNAs in the absence of Argonaute3 reveals somatic piRNAs in flies. Cell 2009, 137, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Mingels, L.; Cordeiro dos Santos, D.; Wynant, N.; Vanden Broeck, J.; Swevers, L. RNAi-based interactions: A latent viral infection in a lepidopteran cell line. In Proceeding of the 28th Conference of European Comparative Endocrinologists, Leuven, Belgium, 21–25 August 2016. [Google Scholar]
- Phillips, A. Diamondback Moth: A Key Pest of Cruciferous Crops; Cooperative Extension Report; Cornell University Publishers: Ithaca, NY, USA, 1983; pp. 4–12. [Google Scholar]
- Wang, Y.; Xu, T.; He, W.; Shen, X.; Zhao, Q.; Bai, J.; You, M. Genome-wide identification and characterization of putative lncRNAs in the diamondback moth, Plutella xylostella (L.). Genomics 2018, 110, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gong, L.; Qi, J.; Hu, M.; Zhong, G. Molecular cloning and characterization of a SID-1-like gene in Plutella xylostella. Arch. Insect Biochem. Physiol. 2014, 87, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Roe, S.M.; Barford, D. Structural insights into mRNA recognition from a PIWI domain–siRNA guide complex. Nature 2005, 434, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, Y.; Szulwach, K.E.; Zhang, G.; Jin, P.; Chen, D. AGO3 Slicer activity regulates mitochondria–nuage localization of Armitage and piRNA amplification. J. Cell Biol. 2014, 206, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.H.; Jiang, L.; Zhu, L.; Cheng, T.C.; Niu, W.H.; Yan, Y.F.; Xia, Q.Y. Characterization of Argonaute family members in the silkworm, Bombyx mori. Insect Sci. 2013, 20, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Tatsuke, T.; Sakashita, K.; Masaki, Y.; Lee, J.M.; Kawaguchi, Y.; Kusakabe, T. The telomere-specific non-LTR retrotransposons SART1 and TRAS1 are suppressed by Piwi subfamily proteins in the silkworm, Bombyx mori. Cell. Mol. Biol. Lett. 2010, 15, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Black, W.C.; Hess, A.M.; Foy, B.D. Comparative genomics of small RNA regulatory pathway components in vector mosquitoes. BMC Genom. 2008, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Rifo, A.; Jannot, G.; Armisen, J.; Labouesse, M.; Bukhari, S.I.A.; Rondeau, E.L.; Miska, E.A.; Simard, M.J. Developmental characterization of the microRNA-specific Caeborhabditis elegans Argonautes alg-1 and alg-2. PLoS ONE 2012, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Seto, A.G.; Kingston, R.E.; Lau, N.C. The coming of age for Piwi proteins. Mol. Cell 2007, 26, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Landthaler, M.; Patkaniowska, A.; Dorsett, Y.; Teng, G.; Tuschl, T. Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell 2004, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Ruda, V.M.; Chandwani, R.; Sehgal, A.; Bogorad, R.L.; Akinc, A.; Charisse, K.; Tarakhovsky, A.; Novobrantseva, T.I.; Koteliansky, V. The roles of individual mammalian argonautes in RNA interference in vivo. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schnettler, E.; Donald, C.L.; Human, S.; Watson, M.; Siu, R.W.; McFarlane, M.; Fazakerley, J.K.; Kohl, A.; Fragkoudis, R. Knockdown of piRNA pathway proteins results in enhanced Semliki Forest virus production in mosquito cells. J. Gen. Virol. 2013, 94, 1680–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, I.; Jansen, S.; Fall, G.; Lorenzen, S.; Rudolf, M.; Huber, K.; Heitmann, A.; Schicht, S.; Watson, M.; Castelli, I. RNA interference restricts Rift Valley Fever virus in multiple insect systems. mSphere 2017, 2, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, D.; Sykora, M.M.; Sachidanandam, R.; Mechtler, K.; Brennecke, J. An in vivo RNAi assay identifies major genetic and cellular requirements for primary piRNA biogenesis in Drosophila. EMBO J. 2010, 29, 3301–3317. [Google Scholar] [CrossRef] [PubMed]
- Kolliopoulou, A.; Swevers, L. Functional analysis of the RNAi response in ovary-derived silkmoth Bm5 cells. Insect Biochem. Mol. Biol. 2013, 43, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zeng, B.; Ling, L.; Xu, J.; You, L.; Aslam, A.F.; Tan, A.; Huang, Y. Enhancement of larval RNAi efficiency by over-expressing Argonaute2 in Bombyx mori. Int. J. Biol. Sci. 2015, 11, 176–185. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.L.; He, W.Y.; Wang, P.; You, M.S. Cell lines from diamondback moth exhibiting differential susceptibility to baculovirus infection and expressing midgut genes. Insect Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
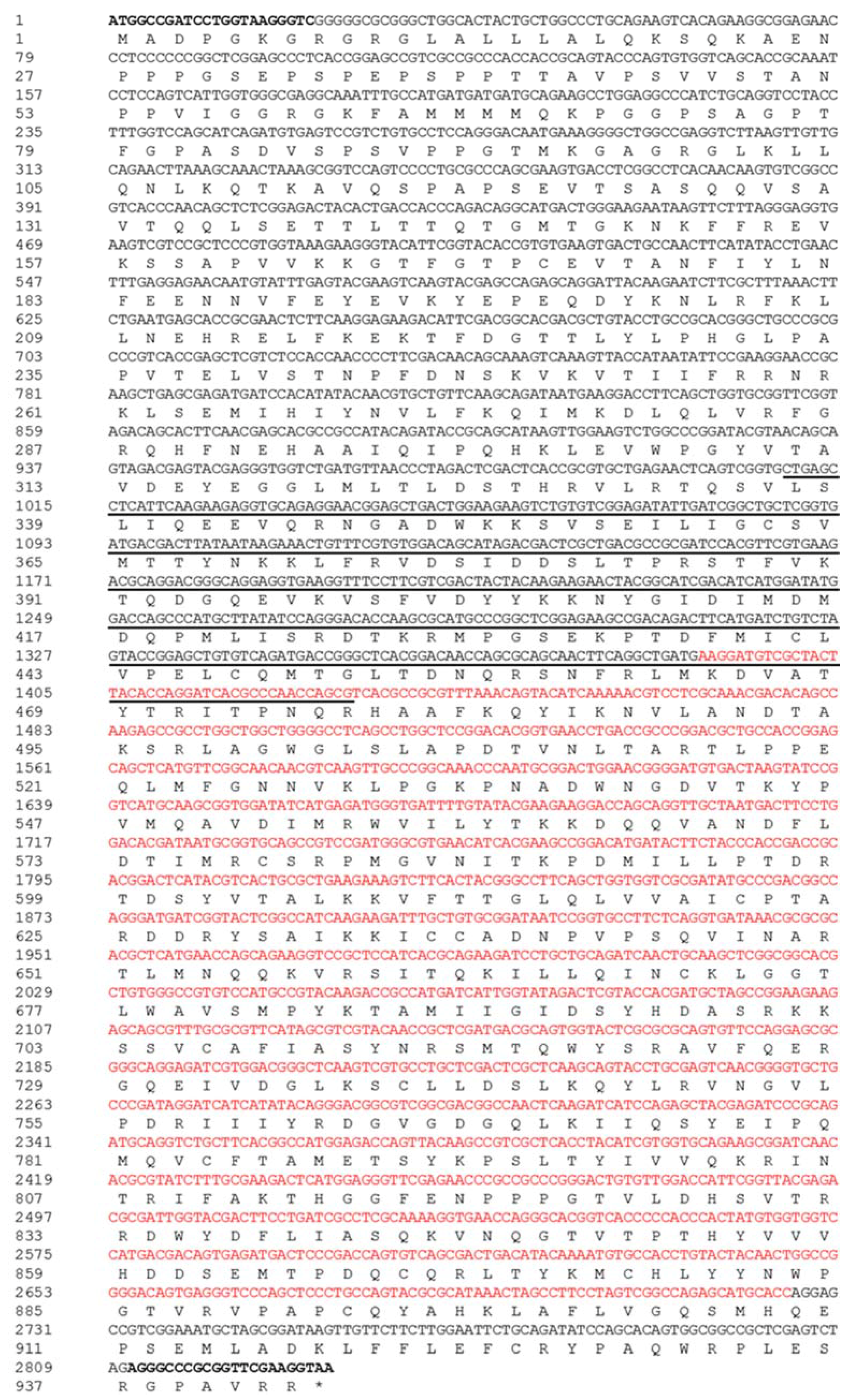
Figure 1.
The complete coding sequence and the assumed amino acid sequence of PxAgo3. The coding sequence is of 2832 bp, starting from ATG marked in boldface and ending with TAA denoted with asterisk (*). The protein sequence is of 943 aa under the nucleotide sequence, with two motifs of the PAZ domain (underline, position 337–477) and the PIWI domain (red in color, position 464–909). Besides this, PCR primers (sense and antisense) are highlighted in bold.
Figure 1.
The complete coding sequence and the assumed amino acid sequence of PxAgo3. The coding sequence is of 2832 bp, starting from ATG marked in boldface and ending with TAA denoted with asterisk (*). The protein sequence is of 943 aa under the nucleotide sequence, with two motifs of the PAZ domain (underline, position 337–477) and the PIWI domain (red in color, position 464–909). Besides this, PCR primers (sense and antisense) are highlighted in bold.
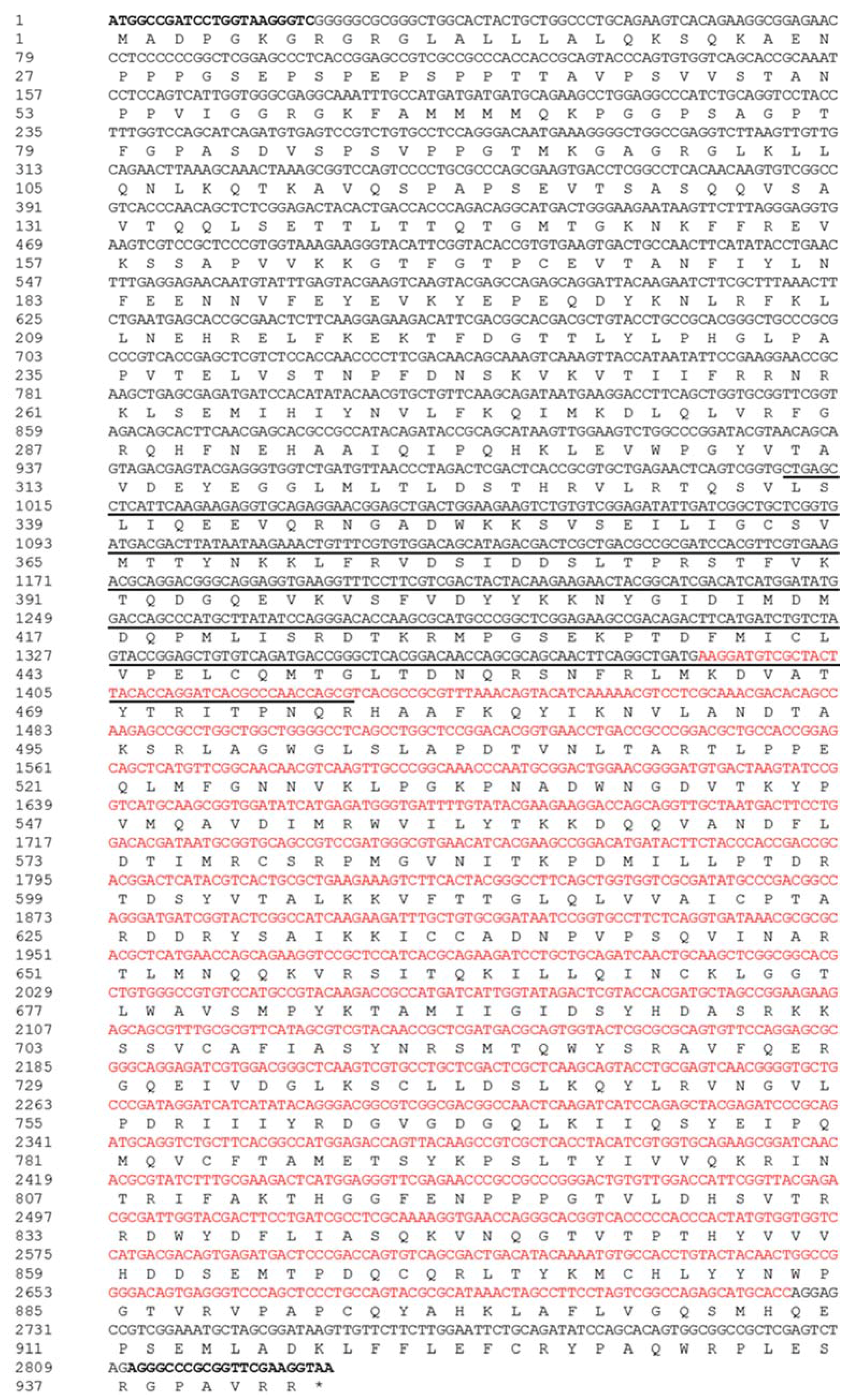
Figure 2.
Multiple-sequence alignment of the PIWI domain of PxAgo3 with those of other species. Multiple-sequence alignment of the PIWI domains of PxAgo3 and descriptive Ago proteins by using the ClustalW software (Omega, Bienne, Switzerland). The undistinguishable amino acids are marked in black shadow, whereas alike amino acids are denoted in blue and grey shadow. The location of catalytic residues of DDH motif are specified with red arrows. Bm means Bombyx mori; Dm, Drosophila melanogaster; Tc, Tribolium castaneum; Hs, Homo sapiens; Px, Plutella xylostella.
Figure 2.
Multiple-sequence alignment of the PIWI domain of PxAgo3 with those of other species. Multiple-sequence alignment of the PIWI domains of PxAgo3 and descriptive Ago proteins by using the ClustalW software (Omega, Bienne, Switzerland). The undistinguishable amino acids are marked in black shadow, whereas alike amino acids are denoted in blue and grey shadow. The location of catalytic residues of DDH motif are specified with red arrows. Bm means Bombyx mori; Dm, Drosophila melanogaster; Tc, Tribolium castaneum; Hs, Homo sapiens; Px, Plutella xylostella.

Figure 3.
Phylogenetic tree of Ago proteins. The neighbor-joining tree was developed by using the MEGA 6.0 software (Molecular Evolutionary Genetics Analysis, Tokyo, Japan). The abbreviations are listed in Table 2. The bootstrap values of 1000 repeats are shown at the nodes.
Figure 3.
Phylogenetic tree of Ago proteins. The neighbor-joining tree was developed by using the MEGA 6.0 software (Molecular Evolutionary Genetics Analysis, Tokyo, Japan). The abbreviations are listed in Table 2. The bootstrap values of 1000 repeats are shown at the nodes.
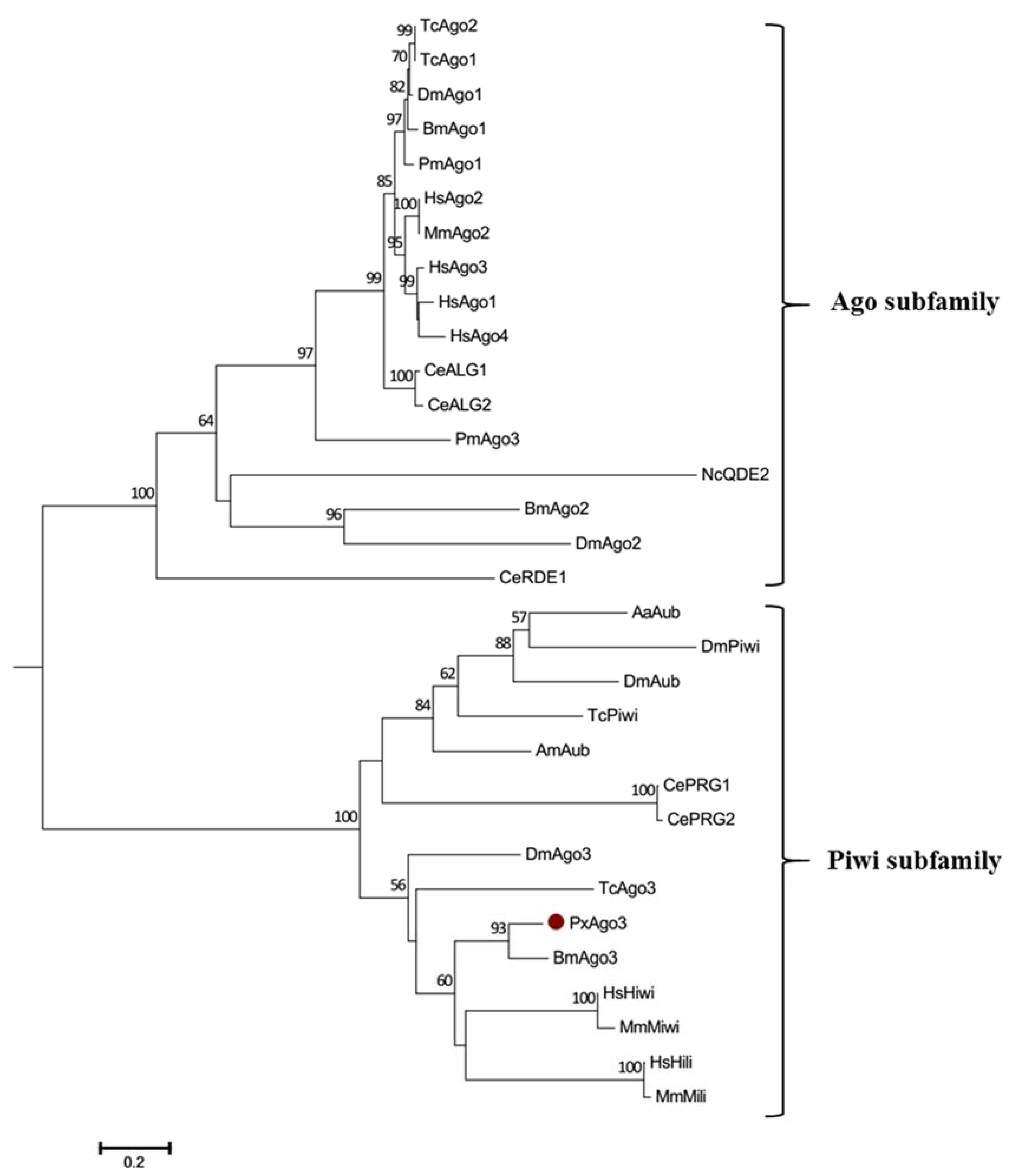
Figure 4.
Comparison of PxAgo3 suppression levels in P. xylostella larvae by dsPxAgo3 at different times after treatment. Different lowercase letters represent significant differences among treatments at each time interval (Tukey’s post hoc test, p < 0.05).
Figure 4.
Comparison of PxAgo3 suppression levels in P. xylostella larvae by dsPxAgo3 at different times after treatment. Different lowercase letters represent significant differences among treatments at each time interval (Tukey’s post hoc test, p < 0.05).

Figure 5.
Effects of PxAgo3 knockdown on PxActin suppression in P. xylostella cells. (A) PxAgo3 expression; (B) PxActin expression. Different lowercase letters represent significant differences among treatments at each time interval (Tukey’s post hoc test, p < 0.05).
Figure 5.
Effects of PxAgo3 knockdown on PxActin suppression in P. xylostella cells. (A) PxAgo3 expression; (B) PxActin expression. Different lowercase letters represent significant differences among treatments at each time interval (Tukey’s post hoc test, p < 0.05).

Figure 6.
Effects of PxAgo3 overexpression on PxActin suppression in P. xylostella cells. (A) P. xylostella cells; (B) the phenotypical expression of PxAgo3 expression vector into P. xylostella cells. Scale Bar: 100 µm; (C) PCR product of PxAgo3 using P. xylostella cells with PxAgo3 expression vector as the template; (D) the expression level of PxAgo3 in P. xylostella cells; (E) the expression level of PxActin in P. xylostella cells. Different lowercase letters represent significant differences among treatments (Tukey’s post hoc test, p < 0.05)
Figure 6.
Effects of PxAgo3 overexpression on PxActin suppression in P. xylostella cells. (A) P. xylostella cells; (B) the phenotypical expression of PxAgo3 expression vector into P. xylostella cells. Scale Bar: 100 µm; (C) PCR product of PxAgo3 using P. xylostella cells with PxAgo3 expression vector as the template; (D) the expression level of PxAgo3 in P. xylostella cells; (E) the expression level of PxActin in P. xylostella cells. Different lowercase letters represent significant differences among treatments (Tukey’s post hoc test, p < 0.05)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in the three experiments of vector construction, expression and suppression for PxAgo3.
Table 1.
Primers used in the three experiments of vector construction, expression and suppression for PxAgo3.
| Primer | Sequence | Use |
|---|---|---|
| PxAgo3-F | 5′ ATGGCCGATCCTGGTAAGGGTC 3′ | PCR |
| PxAgo3-R | 5′ TTACCTTCGAACCGCGGGCCCT 3′ | PCR |
| PxAgo3 KpnI-F1 | 5′ CGGGGTACCATGGCCGATCCTGGTAAGG 3′ | PCR |
| PxAgo3 EcoRI-R1 | 5′ CCGGAATTCCAAGAAGAACAACTTATCCGC 3′ | PCR |
| PIZT-F | 5′ CGCAACGATCTGGTAAACAC 3′ | PCR |
| PIZT-R | 5′ CTGACTAAATCTTAGTTTGTATTGTC 3′ | PCR |
| PxAgo3-F1 | 5′ GCCGCCATACAGATACCG 3′ | RT-qPCR |
| PxAgo3-R1 | 5′ AACATCAGACCACCCTCG 3′ | RT-qPCR |
| PxActin-F2 | 5′ GGAGTGATGGTCGGTATGGGA 3′ | RT-qPCR |
| PxActin-R2 | 5′ CTCGATGGGGTACTTCAGGGT 3′ | RT-qPCR |
| RL32-F | 5′ CAATCAGGCCAATTTACCGC 3′ | RT-qPCR |
| RL32-R | 5′ CTGGGTTTACGCCAGTTACG 3′ | RT-qPCR |
| dsAgo3-F | 5′ GTCCGCTCCATCACGCAGAAA 3′ | dsRNA Preparation |
| dsAgo3-R | 5′ GCGGGATCTCGTAGCTCTGGA 3′ | dsRNA Preparation |
| dsActin-F | 5′ GCCAACCGTGAAAAGATGA 3′ | dsRNA Preparation |
| dsActin-R | 5′ AGGAATGAGGGCTGGAACA 3′ | dsRNA Preparation |
| dsEGFP-F | 5′ GTGTTCAATGCTTTTCCCGTTATCC 3′ | dsRNA Preparation |
| dsEGFP-R | 5′ ACCATGTGGTCACGCTTTTCG 3′ | dsRNA Preparation |
Note: PCR, polymerase chain reaction, RT-qPCR, quantitative reverse transcription PCR; dsRNA, double stranded RNA.
Table 2.
Ago proteins with the accession number for multiple-alignment comparison and phylogenetic tree analyses.
Table 2.
Ago proteins with the accession number for multiple-alignment comparison and phylogenetic tree analyses.
| Organism | Protein | Abbreviation | Acc. Number |
|---|---|---|---|
| Tribolium castaneum | Argonaute-1 | TcAgo1 | KYB26000.1 |
| Argonaute-2 | TcAgo2 | XP_015837988.1 | |
| Argonaute 3 | TcAgo3 | EFA02921.1 | |
| PIWI | TcPIWI | EFA07425.1 | |
| Drosophila melanogaster | Argonaute-1 | DmAgo1 | AAF58313.1 |
| Argonaute-2 | DmAgo2 | NP_730054.1 | |
| Argonaute-3 | DmAgo3 | EAA45981.3 | |
| Aubergine | DmAub | CAA64320.1 | |
| PIWI | DmPIWI | AGL81541.1 | |
| Bombyx mori | Argonaute-1 | BmAgo1 | NP_001095931.1 |
| Argonaute-2 | BmAgo2 | BAD91160.2 | |
| Argonaute-3 | BmAgo3 | A9ZSZ2 | |
| Caenorhabditis elegans | CeALG-1 | CeALG1 | CAR97837.1 |
| CeALG-2 | CeALG2 | CCD73271.1 | |
| CePRG-1 | CePRG1 | CAA98113.1 | |
| CePRG-2 | CePRG2 | CCD62443.1 | |
| CeRDE-1 | CeRDE1 | CAB05546 | |
| Homo sapiens | Argonaute-1 | HsAgo1 | NP_036331.1 |
| Argonaute-2 | HsAgo2 | NP_036286.2 | |
| Argonaute-3 | HsAgo3 | NP_079128.2 | |
| Argonaute-4 | HsAgo4 | NP_060099.2 | |
| HsHiwi | HsHiwi | AAC97371.2 | |
| HsHili | HsHili | NP060538.2 | |
| Penaeus monodon | Argonaute-1 | PmAgo1 | ABG66641.1 |
| Argonaute-3 | PmAgo3 | AGC95229.1 | |
| Apis mellifera | Aubergine | AmAub | NP_001159378.1 |
| Aedes aegypti | Aubergine | AaAub | XP_021707306.1 |
| Neurospora crassa | NcQDE-2 | NcQDE2 | AAF43641 |
| Mus musculus | Argonaute-2 | MmAgo2 | NP_694818 |
| MmMiwi | MmMiwi | BAM37459 | |
| MmMili | MmMili | BAA93706 | |
| Plutella xylostella | Argonaute-3 | PxAgo3 | MG778697 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hameed, M.S.; Wang, Z.; Vasseur, L.; Yang, G. Molecular Characterization and the Function of Argonaute3 in RNAi Pathway of Plutella xylostella. Int. J. Mol. Sci. 2018, 19, 1249. https://doi.org/10.3390/ijms19041249
AMA Style
Hameed MS, Wang Z, Vasseur L, Yang G. Molecular Characterization and the Function of Argonaute3 in RNAi Pathway of Plutella xylostella. International Journal of Molecular Sciences. 2018; 19(4):1249. https://doi.org/10.3390/ijms19041249
Chicago/Turabian StyleHameed, Muhammad Salman, Zhengbing Wang, Liette Vasseur, and Guang Yang. 2018. "Molecular Characterization and the Function of Argonaute3 in RNAi Pathway of Plutella xylostella" International Journal of Molecular Sciences 19, no. 4: 1249. https://doi.org/10.3390/ijms19041249
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.