Bone Cell Activity in Clinical Prostate Cancer Bone Metastasis and Its Inverse Relation to Tumor Cell Androgen Receptor Activity


Abstract
:1. Introduction
2. Results
2.1. Parallel Activation of Osteoclasts and Osteoblasts in Prostate Cancer Bone Metastasis
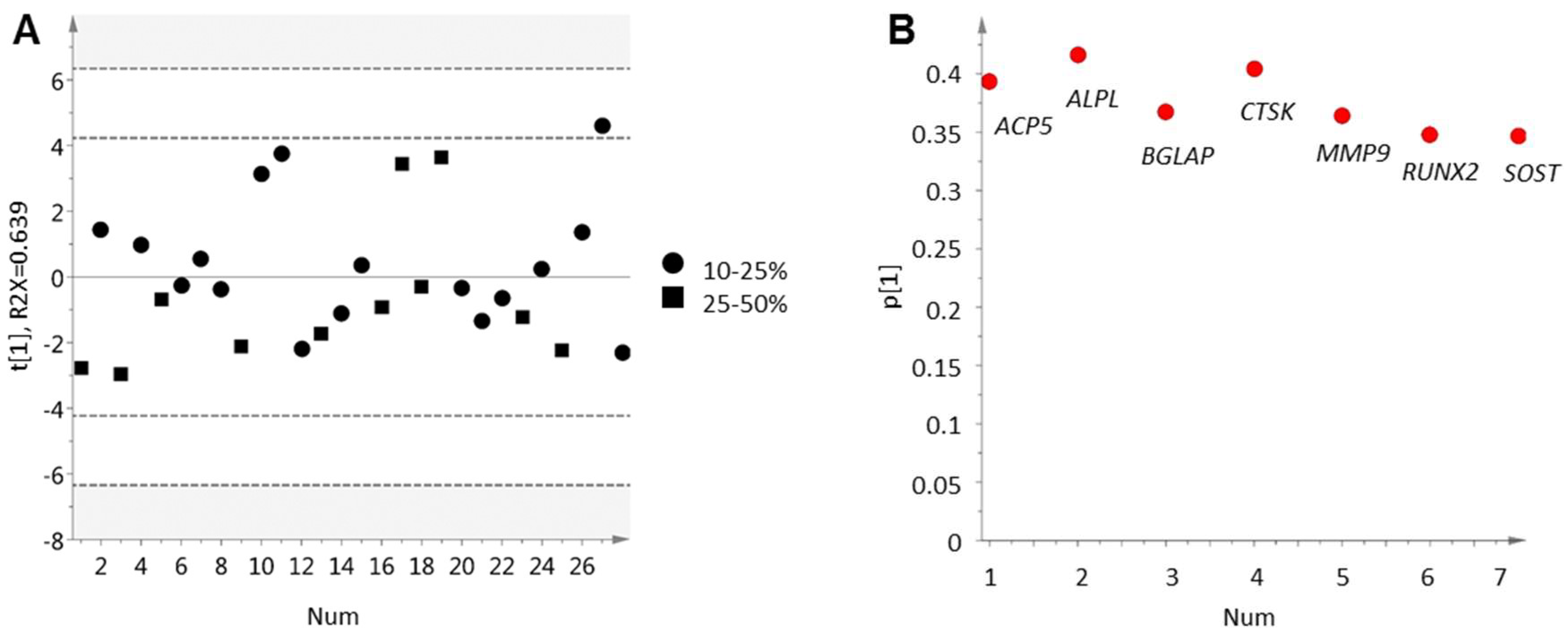
2.2. Inverse Relation between Bone Cell Activity and Tumor Cell AR Activity
2.3. Bone Formation in Prostate Cancer Metastasis Is Associated with Bone Morphogenetic Protein Signaling
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. RNA Extraction and Gene Expression Analysis
4.3. Univariate Statistics
4.4. Multivariate Data Analysis
4.5. Functional Enrichment Analysis
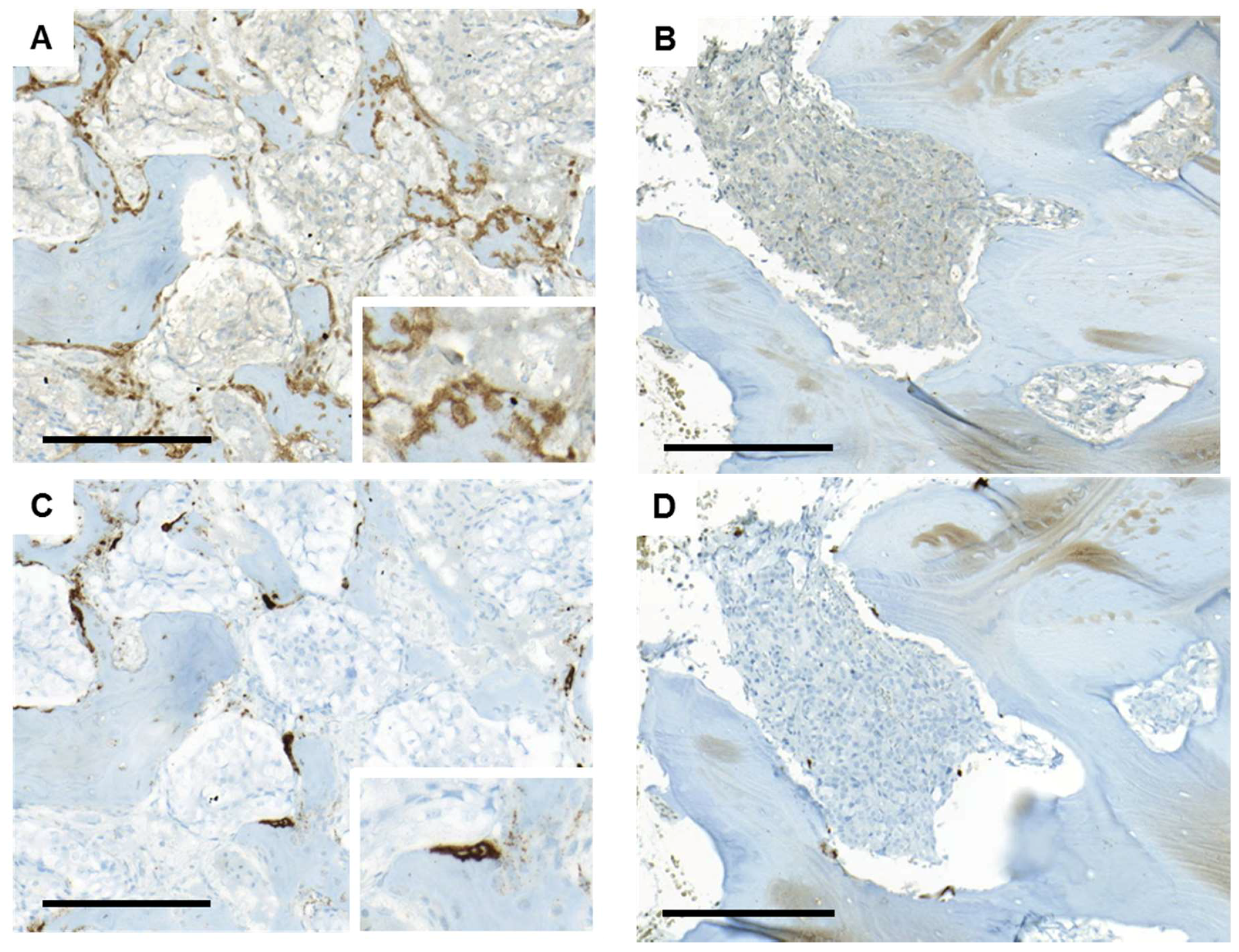
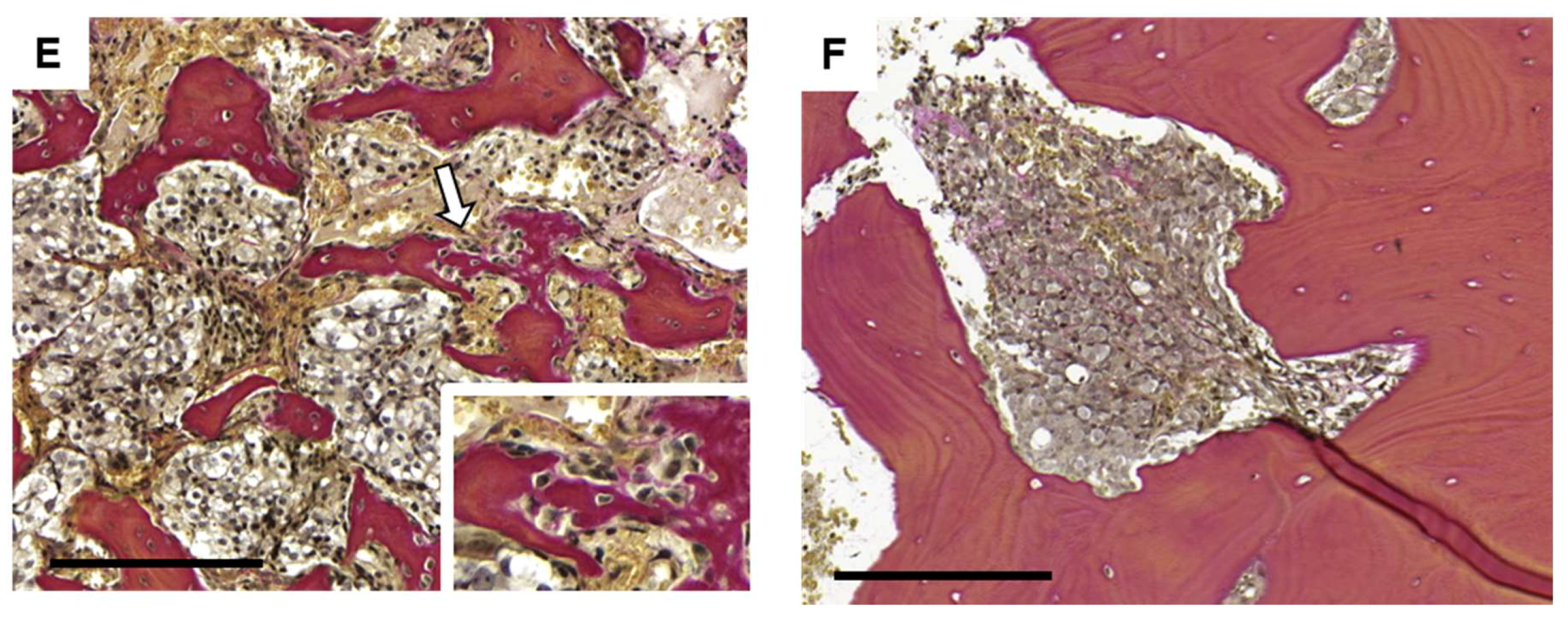
4.6. Immunohistochemistry and Bone Histomorphometry
5. Conclusions
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Bubendorf, L.; Schöpfer, A.; Wagner, U.; Sauter, G.; Moch, H.; Willi, N.; Gasser, T.C.; Mihatsch, M.J. Metastatic Patterns of Prostate Cancer: An Autopsy Study of 1,589 Patients. Hum. Pathol. 2000, 31, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Lorente, D.; Fizazi, K.; Sweeney, C.; de Bono, J.S. Optimal Treatment Sequence for Metastatic Castration-Resistant Prostate Cancer. Eur. Urol. Focus 2016, 2, 488–498. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Knudsen, B.S.; Erho, N.; Alshalalfa, M.; Takhar, M.; Ashab, H.A.D.; Davicioni, E.; Karnes, R.J.; Klein, E.A.; Den, R.B.; et al. Integrated Classification of Prostate Cancer Reveals a Novel Luminal Subtype with Poor Outcome. Cancer Res. 2016, 76, 4948–4958. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.G.; Chang, S.L.; Spratt, D.E.; Erho, N.; Yu, M.; Ashab, H.A.D.; Alshalalfa, M.; Speers, C.; Tomlins, S.A.; Davicioni, E.; et al. Development and Validation of a 24-Gene Predictor of Response to Postoperative Radiotherapy in Prostate Cancer: A Matched, Retrospective Analysis. Lancet Oncol. 2016, 17, 1612–1620. [Google Scholar] [CrossRef]
- Hörnberg, E.; Ylitalo, E.B.; Crnalic, S.; Antti, H.; Stattin, P.; Widmark, A.; Bergh, A.; Wikström, P. Expression of Androgen Receptor Splice Variants in Prostate Cancer Bone Metastases Is Associated with Castration-Resistance and Short Survival. PLoS ONE 2011, 6, e19059. [Google Scholar] [CrossRef] [PubMed]
- Jernberg, E.; Thysell, E.; Ylitalo, E.B.; Rudolfsson, S.; Crnalic, S.; Widmark, A.; Bergh, A.; Wikström, P. Characterization of Prostate Cancer Bone Metastases according to Expression Levels of Steroidogenic Enzymes and Androgen Receptor Splice Variants. PLoS ONE 2013, 8, e77407. [Google Scholar] [CrossRef] [PubMed]
- Ylitalo, E.B.; Thysell, E.; Jernberg, E.; Lundholm, M.; Crnalic, S.; Egevad, L.; Stattin, P.P.; Widmark, A.; Bergh, A.; Wikström, P.; et al. Subgroups of Castration-Resistant Prostate Cancer Bone Metastases Defined through an Inverse Relationship between Androgen Receptor Activity and Immune Response. Eur. Urol. 2017, 71, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Thysell, E.; Ylitalo, E.B.; Jernberg, E.; Bergh, A.; Wikström, P. A Systems Approach to Prostate Cancer Classification-Letter. Cancer Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Roudier, M.P.; Vesselle, H.; True, L.D.; Higano, C.S.; Ott, S.M.; King, S.H.; Vessella, R.L. Bone Histology at Autopsy and Matched Bone Scintigraphy Findings in Patients with Hormone Refractory Prostate Cancer: The Effect of Bisphosphonate Therapy on Bone Scintigraphy Results. Clin. Exp. Metastasis 2003, 20, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Keller, E.T.; Brown, J. Prostate Cancer Bone Metastases Promote both Osteolytic and Osteoblastic Activity. J. Cell. Biochem. 2004, 91, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, T.; Flamini, E.; Mercatali, L.; Sacanna, E.; Serra, P.; Amadori, D. Pathogenesis of Osteoblastic Bone Metastases from Prostate Cancer. Cancer 2010, 116, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Berruti, A.; Dogliotti, L.; Bitossi, R.; Fasolis, G.; Gorzegno, G.; Bellina, M.; Torta, M.; Porpiglia, F.; Fontana, D.; Angeli, A. Incidence of Skeletal Complications in Patients with Bone Metastatic Prostate Cancer and Hormone Refractory Disease: Predictive Role of Bone Resorption and Formation Markers Evaluated at Baseline. J. Urol. 2000, 164, 1248–1253. [Google Scholar] [CrossRef]
- Roudier, M.P.; Morrissey, C.; True, L.D.; Higano, C.S.; Vessella, R.L.; Ott, S.M. Histopathological Assessment of Prostate Cancer Bone Osteoblastic Metastases. J. Urol. 2008, 180, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B. Normal Bone Anatomy and Physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. S3), 131–139. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.E. Clinical Features of Metastatic Bone Disease and Risk of Skeletal Morbidity. Clin. Cancer Res. 2006, 2 Pt 12, 6243s–6249s. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Chiu, M.; Moore, A.J.; Anderson, P.H.; Ghasem-Zadeh, A.; McManus, J.F.; Ma, C.; Seeman, E.; Clemens, T.L.; Morris, H.A.; et al. Mineralization and Bone Resorption Are Regulated by the Androgen Receptor in Male Mice. J. Bone Miner. Res. 2009, 24, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C.; O’Brien, C.A.; Almeida, M. The Role of Estrogen and Androgen Receptors in Bone Health and Disease. Nat. Rev. Endocrinol. 2013, 9, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Shahinian, V.B.; Kuo, Y.-F.; Freeman, J.L.; Goodwin, J.S. Risk of Fracture after Androgen Deprivation for Prostate Cancer. N. Engl. J. Med. 2005, 352, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.L.; Alibhai, S.M.H.; Basaria, S.; D’Amico, A.V.; Kantoff, P.W.; Keating, N.L.; Penson, D.F.; Rosario, D.J.; Tombal, B.; Smith, M.R. Adverse Effects of Androgen Deprivation Therapy and Strategies to Mitigate Them. Eur. Urol. 2015, 67, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Mohler, J.L.; Gregory, C.W.; Ford, O.H., 3rd; Kim, D.; Weaver, C.M.; Petrusz, P.; Wilson, E.M.; French, F.S. The Androgen Axis in Recurrent Prostate Cancer. Clin. Cancer Res. 2004, 10, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.B.; Mostaghel, E.A.; Vessella, R.; Hess, D.L.; Kalhorn, T.F.; Higano, C.S.; True, L.D.; Nelson, P.S. Maintenance of Intratumoral Androgens in Metastatic Prostate Cancer: A Mechanism for Castration-Resistant Tumor Growth. Cancer Res. 2008, 68, 4447–4454. [Google Scholar] [CrossRef] [PubMed]
- Crnalic, S.; Hörnberg, E.; Wikström, P.; Lerner, U.H.; Tieva, Å.; Svensson, O.; Widmark, A.; Bergh, A. Nuclear Androgen Receptor Staining in Bone Metastases Is Related to a Poor Outcome in Prostate Cancer Patients. Endocr. Relat. Cancer 2010, 17, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Fizazi, K.; Carducci, M.; Smith, M.; Damião, R.; Brown, J.; Karsh, L.; Milecki, P.; Shore, N.; Rader, M.; Wang, H.; et al. Denosumab versus Zoledronic Acid for Treatment of Bone Metastases in Men with Castration-Resistant Prostate Cancer: A Randomised, Double-Blind Study. Lancet 2011, 377, 813–822. [Google Scholar] [CrossRef]
- Parker, C.; Nilsson, S.; Heinrich, D.; Helle, S.I.; O’Sullivan, J.M.; Fosså, S.D.; Chodacki, A.; Wiechno, P.; Logue, J.; Seke, M.; et al. Alpha Emitter Radium-223 and Survival in Metastatic Prostate Cancer. N. Engl. J. Med. 2013, 369, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, R.; Hata, K.; Matsubara, T.; Wakabayashi, M.; Yoneda, T. Regulation of Bone and Cartilage Development by Network between BMP Signalling and Transcription Factors. J. Biochem. 2012, 151, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Van Beuningen, H.M.; Glansbeek, H.L.; van der Kraan, P.M.; van den Berg, W.B. Differential Effects of Local Application of BMP-2 or TGF-β 1 on Both Articular Cartilage Composition and Osteophyte Formation. Osteoarthr. Cartil. 1998, 6, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yuan, G.; Li, X.; Liu, P.; Chen, Z.; Fan, M. BMP-2 Induction of Dlx3 Expression Is Mediated by p38/Smad5 Signaling Pathway in Osteoblastic MC3T3-E1 Cells. J. Cell. Physiol. 2014, 229, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-D.; Fisher, L.W.; Robey, P.G.; Young, M.F. The Small Leucine-Rich Proteoglycan Biglycan Modulates BMP-4-Induced Osteoblast Differentiation. FASEB J. 2004, 18, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Mochida, Y.; Parisuthiman, D.; Yamauchi, M. Biglycan Is a Positive Modulator of BMP-2 Induced Osteoblast Differentiation. Adv. Exp. Med. Biol. 2006, 585, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-O.; Kim, E.-J.; Cho, K.-W.; Nakagawa, E.; Kwon, H.-J.; Cho, S.-W.; Jung, H.-S. BMP4 Signaling Mediates Zeb Family in Developing Mouse Tooth. Histochem. Cell Biol. 2012, 137, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Inoue, Y.; Tobimatsu, T.; Hendy, G.; Canaff, L.; Sugimoto, T.; Seino, S.; Kaji, H. Smad7 Inhibits Differentiation and Mineralization of Mouse Osteoblastic Cells. Endocr. J. 2012, 59, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Abe, T.; Aida, Y.; Hara, Y.; Maeda, K. Follistatin Restricts Bone Morphogenetic Protein (BMP)-2 Action on the Differentiation of Osteoblasts in Fetal Rat Mandibular Cells. J. Bone Miner. Res. 2004, 19, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Florencio-Silva, R.; Sasso, G.R.D.S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. Biomed Res. Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Kim, H.J.H.-H.; Chang, E.-J.; Huang, H.; Banno, Y.; Kim, H.J.H.-H. Sphingosine 1-Phosphate as a Regulator of Osteoclast Differentiation and Osteoclast-Osteoblast Coupling. EMBO J. 2006, 25, 5840–5851. [Google Scholar] [CrossRef] [PubMed]
- Ecarot, B.; Desbarats, M. 1,25-(OH)2D3 Down-Regulates Expression of Phex, a Marker of the Mature Osteoblast 1. Endocrinology 1999, 140, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Koizumi, M.; Yoshimura, K.; Yamauchi, T.; Kawai, T.; Ogata, E. Correlation between Bone Metabolic Markers and Bone Scan in Prostatic Cancer. J. Urol. 1997, 157, 539–543. [Google Scholar] [CrossRef]
- Costa, L.; Demers, L.M.; Gouveia-Oliveira, A.; Schaller, J.; Costa, E.B.; De Moura, M.C.; Lipton, A. Prospective Evaluation of the Peptide-Bound Collagen Type I Cross-Links N-Telopeptide and C-Telopeptide in Predicting Bone Metastases Status. J. Clin. Oncol. 2002, 20, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Weilbaecher, K.N.; Guise, T.A.; McCauley, L.K. Cancer to Bone: A Fatal Attraction. Nat. Rev. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K.; Kamiya, Y.; Morikawa, M. Bone Morphogenetic Protein Receptors and Signal Transduction. J. Biochem. 2010, 147, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.; Bae, J.S.; Afza, F.; Gutierrez, S.; Pratap, J.; Zaidi, S.K.; Lou, Y.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Structural Coupling of Smad and Runx2 for Execution of the BMP2 Osteogenic Signal. J. Biol. Chem. 2008, 283, 8412–8422. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Lein, M. Bone Turnover Markers in Serum and Urine as Diagnostic, Prognostic and Monitoring Biomarkers of Bone Metastasis. Biochim. Biophys. Acta Rev. Cancer 2014, 1846, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.L.; Khosla, S. Androgens and Bone. Steroids 2009, 74, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, D.; Laurent, M.R.; Claessens, F.; Gielen, E.; Lagerquist, M.K.; Vandenput, L.; Börjesson, A.E.; Ohlsson, C. Sex Steroid Actions in Male Bone. Endocr. Rev. 2014, 35, 906–960. [Google Scholar] [CrossRef] [PubMed]
- Russell, P.K.; Clarke, M.V.; Cheong, K.; Anderson, P.H.; Morris, H.A.; Wiren, K.M.; Zajac, J.D.; Davey, R.A. Androgen Receptor Action in Osteoblasts in Male Mice Is Dependent on Their Stage of Maturation. J. Bone Miner. Res. 2015, 30, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Inoue, T.; Narita, S.; Maita, S.; Huang, M.; Numakura, K.; Tsuruta, H.; Saito, M.; Maeno, A.; Satoh, S.; et al. Inhibition of the RANK/RANKL Signaling with Osteoprotegerin Prevents Castration-Induced Acceleration of Bone Metastasis in Castration-Insensitive Prostate Cancer. Cancer Lett. 2017, 397, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Proell, V.; Xu, H.; Schüler, C.; Weber, K.; Hofbauer, L.C.; Erben, R.G. Orchiectomy Upregulates Free Soluble RANKL in Bone Marrow of Aged Rats. Bone 2009, 45, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.; Raja, E.; Miyazono, K.; Tsubakihara, Y.; Moustakas, A. Mechanisms of Action of Bone Morphogenetic Proteins in Cancer. Cytokine Growth Factor Rev. 2016, 27, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Jiang, W.G. Bone Morphogenetic Proteins in Tumour Associated Angiogenesis and Implication in Cancer Therapies. Cancer Lett. 2016, 380, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhong, C.; Frenkel, B.; Reddi, A.H.; Roy-Burman, P. Diverse Biological Effect and Smad Signaling of Bone Morphogenetic Protein 7 in Prostate Tumor Cells. Cancer Res. 2005, 65, 5769–5777. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Cheng, C.J.; Bilen, M.A.; Lu, J.F.; Satcher, R.L.; Yu-Lee, L.Y.; Gallick, G.E.; Maity, S.N.; Lin, S.H. BMP4 Promotes Prostate Tumor Growth in Bone through Osteogenesis. Cancer Res. 2011, 71, 5194–5203. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lee, Y.C.; Yu, G.; Cheng, C.J.; Zhou, X.; Chu, K.; Murshed, M.; Le, N.T.; Baseler, L.; Abe, J.-I.; et al. Endothelial-to-Osteoblast Conversion Generates Osteoblastic Metastasis of Prostate Cancer. Dev. Cell 2017, 41, 467–480.e3. [Google Scholar] [CrossRef] [PubMed]
- Kruger, T.E.; Miller, A.H.; Godwin, A.K.; Wang, J. Bneo Sialoprotein and Osteopontin in Bone Metastasis of Osteotropic Cancers. Crit. Rev. Oncol. Hematol. 2014, 89, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Reinholt, F.P.; Hultenby, K.; Oldberg, A.; Heinegård, D. Osteopontin—A Possible Anchor of Osteoclasts to Bone. Proc. Natl. Acad. Sci. USA 1990, 87, 4473–4475. [Google Scholar] [CrossRef] [PubMed]
- Khodavirdi, A.C.; Song, Z.; Yang, S.; Wu, H.; Pritchard, C.; Nelson, P.S.; Roy-burman, P. Increased Expression of Osteopontin Contributes to the Progression of Prostate Cancer. Cancer Res. 2006, 66, 883–888. [Google Scholar] [CrossRef] [PubMed]
- McAllister, S.S.; Gifford, A.M.; Greiner, A.L.; Kelleher, S.P.; Saelzler, M.P.; Ince, T.A.; Reinhardt, F.; Harris, L.N.; Hylander, B.L.; Repasky, E.A.; et al. Systemic Endocrine Instigation of Indolent Tumor Growth Requires Osteopontin. Cell 2008, 133, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Liu, X.; Wang, J.; Chen, X.; Zhang, H.; Kim, S.H.; Cui, J.; Li, R.; Zhang, W.; Kong, Y.; et al. Wnt Signaling in Bone Formation and Its Therapeutic Potential for Bone Diseases. Ther. Adv. Musculoskelet. Dis. 2013, 5, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Guise, T.A.; Yin, J.J.; Taylor, S.D.; Kumagai, Y.; Dallas, M.; Boyce, B.F.; Yoneda, T.; Mundy, G.R. Evidence for a Causal Role of Parathyroid Hormone-Related Protein in the Pathogenesis of Human Breast Cancer-Mediated Osteolysis. J. Clin. Investig. 1996, 98, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Djusberg, E.; Jernberg, E.; Thysell, E.; Golovleva, I.; Lundberg, P.; Crnalic, S.; Widmark, A.; Bergh, A.; Brattsand, M.; Wikström, P. High Levels of the AR-V7 Splice Variant and Co-Amplification of the Golgi Protein Coding YIPF6 in AR Amplified Prostate Cancer Bone Metastases. Prostate 2017, 77, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Gato, D.; Wikström, P.; Tyanova, S.; Lavallee, C.; Thysell, E.; Carlsson, J.; Hägglöf, C.; Cox, J.; Andrén, O.; Stattin, P.; et al. The Proteome of Primary Prostate Cancer. Eur. Urol. 2016, 69, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Thysell, E.; Surowiec, I.; Hörnberg, E.; Crnalic, S.; Widmark, A.; Johansson, A.I.; Stattin, P.; Bergh, A.; Moritz, T.; Antti, H.; et al. Metabolomic Characterization of Human Prostate Cancer Bone Metastases Reveals Increased Levels of Cholesterol. PLoS ONE 2010, 5, e14175. [Google Scholar] [CrossRef] [PubMed]
- Nordstrand, A.; Lundholm, M.; Larsson, A.; Lerner, U.H.; Widmark, A.; Wikström, P. Inhibition of the Insulin-like Growth Factor-1 Receptor Enhances Effects of Simvastatin on Prostate Cancer Cells in Co-Culture with Bone. Cancer Microenviron. 2013, 6, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Feeley, B.T.; Krenek, L.; Liu, N.; Hsu, W.K.; Gamradt, S.C.; Schwarz, E.M.; Huard, J.; Lieberman, J.R. Overexpression of Noggin Inhibits BMP-Mediated Growth of Osteolytic Prostate Cancer Lesions. Bone 2006, 38, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yoshioka, Y.; Mandai, M.; Matsumura, N.; Baba, T.; Yamaguchi, K.; Hamanishi, J.; Kharma, B.; Murakami, R.; Abiko, K.; et al. The BMP Signaling Pathway Leads to Enhanced Proliferation in Serous Ovarian Cancer-a Potential Therapeutic Target. Mol. Carcinog. 2015, 55, 335–345. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristic | Non-Treated n = 11 | Castration-Resistant a n = 28 |
|---|---|---|
| Age at diagnosis (years) | 77 (74–82) | 67 (63–74) |
| Age at metastasis surgery (years) | 77 (74–82) | 71 (68–79) |
| Serum PSA at diagnosis (ng/mL) | 170 (60–980) | 97 (37–260) |
| Serum PSA at metastasis surgery (ng/mL) | 170 (60–980) | 260 (61–490) |
| Gleason score at diagnosis | ||
| 7 | 1 (9) | 9 (32) |
| 8–10 | 1 (9) | 14 (50) |
| Not available | 9 (82) | 5 (18) |
| Bicalutamide prior to surgery | ||
| Yes | 15 (54) | |
| No | 12 (43) | |
| Not available | 1 (3.6) | |
| Chemotherapy prior to surgery b | ||
| Yes | 4 (14) | |
| No | 24 (86) | |
| Ra-223 prior to surgery | ||
| Yes | 2 (7.1) | |
| No | 26 (93) | |
| Radiation prior to surgery c | ||
| Yes | 7 (25) | |
| No | 21 (75) | |
| Follow up after first ADT (months) | 37 (24–72) | 43 (25–70) |
| Follow up after metastasis surgery (months) | 37 (24–72) | 8.5 (2.2–18) |
| Network | Total Gene Products | p (FDR b) | Gene Products in Data |
|---|---|---|---|
| Development: Ossification and bone remodeling | 157 | 3.4 × 10−11 | ALPL, FST, FOXC2, TEAD4, LEF1, PHEX, MEPE, OMD, DLX5, FOXC1, SP7, SOST, COL1A1, COL1A2, BGLAP, DLX3, CEBPB, SMAD7, RUNX2, CEBPD, SPP1, IBSP, BMP4 |
| Development: Regulation of epithelial-to-mesenchymal transition (EMT) | 225 | 5.4 × 10−6 | SNAI2, MMP9, FOXC2, LEF1, PDGFD, COL1A2, ILK, NFKBIA, EDNRA, COL1A1, COL1A2, ACTB, JAK2, BCL2, VIM, ZEB2, SMAD7, PIK3R1, BMP4 |
| Cell adhesion: Integrin mediated cell-matrix adhesion | 214 | 7.9 × 10−6 | ACTN1, ITGA10, RAPH1, OMD, ARHGEF6, ILK, TNC, COL1A1, COL1A2, ACTB, ITGA2, LEF1, PIK3R1, RND3, SPP1, IBSP, TNS3 |
| Cell adhesion: Cadherins | 180 | 2.8 × 10−5 | ACTN1, FXYD5, ILK, MTSS1, WISP1, SWAP70, LEF1, DKK1, ACTB, CDH15, PIK3R1, PCDH18, WIF1 |
| Proliferation: Positive regulation cell proliferation | 221 | 6.2 × 10−5 | RUNX3, TCIRG1, ZFP36L2, CSF1R, PLGF, GPC4, ILK, KIT, SCGF, CSPG4, EDNRA, JAK2, SIPR3, EMP1, GPC4, PIK3R1 |
| Signal Transduction: Bone morphogenic protein (BMP) and growth differentiation factor (GDF) signaling | 91 | 6.2 × 10−5 | FST, ID1, SOST, BGLAP, PPP2R2B, SMAD7, RUNX2, SPP1, IBSP, BMP4 |
| Cell adhesion: Cell junctions | 162 | 9.3 × 10−5 | ACTN1, ACTB, VIM, CLDN11, LEF1, ZEB2, CDH15, PIK3R1, YWHAH |
| Inflammation: IL-6 signaling | 119 | 9.3 × 10−5 | PIK3R1, NFKBIA, JAK2, BCL2, CEBPB, YWHAH |
| Development: Cartilage development | 66 | 1.3 × 10−4 | COL1A1, RUNX3, CHAD, COL1A2, SMAD7, RUNX2, BMP4 |
| Inflammation: Protein C signaling | 108 | 1.6 × 10−4 | ACTN1, SPHK1, NFKBIA, ACTB, S1PR3, PROS1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nordstrand, A.; Bovinder Ylitalo, E.; Thysell, E.; Jernberg, E.; Crnalic, S.; Widmark, A.; Bergh, A.; Lerner, U.H.; Wikström, P. Bone Cell Activity in Clinical Prostate Cancer Bone Metastasis and Its Inverse Relation to Tumor Cell Androgen Receptor Activity. Int. J. Mol. Sci. 2018, 19, 1223. https://doi.org/10.3390/ijms19041223
Nordstrand A, Bovinder Ylitalo E, Thysell E, Jernberg E, Crnalic S, Widmark A, Bergh A, Lerner UH, Wikström P. Bone Cell Activity in Clinical Prostate Cancer Bone Metastasis and Its Inverse Relation to Tumor Cell Androgen Receptor Activity. International Journal of Molecular Sciences. 2018; 19(4):1223. https://doi.org/10.3390/ijms19041223
Chicago/Turabian StyleNordstrand, Annika, Erik Bovinder Ylitalo, Elin Thysell, Emma Jernberg, Sead Crnalic, Anders Widmark, Anders Bergh, Ulf H. Lerner, and Pernilla Wikström. 2018. "Bone Cell Activity in Clinical Prostate Cancer Bone Metastasis and Its Inverse Relation to Tumor Cell Androgen Receptor Activity" International Journal of Molecular Sciences 19, no. 4: 1223. https://doi.org/10.3390/ijms19041223