Triclosan Lacks (Anti-)Estrogenic Effects in Zebrafish Cells but Modulates Estrogen Response in Zebrafish Embryos

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
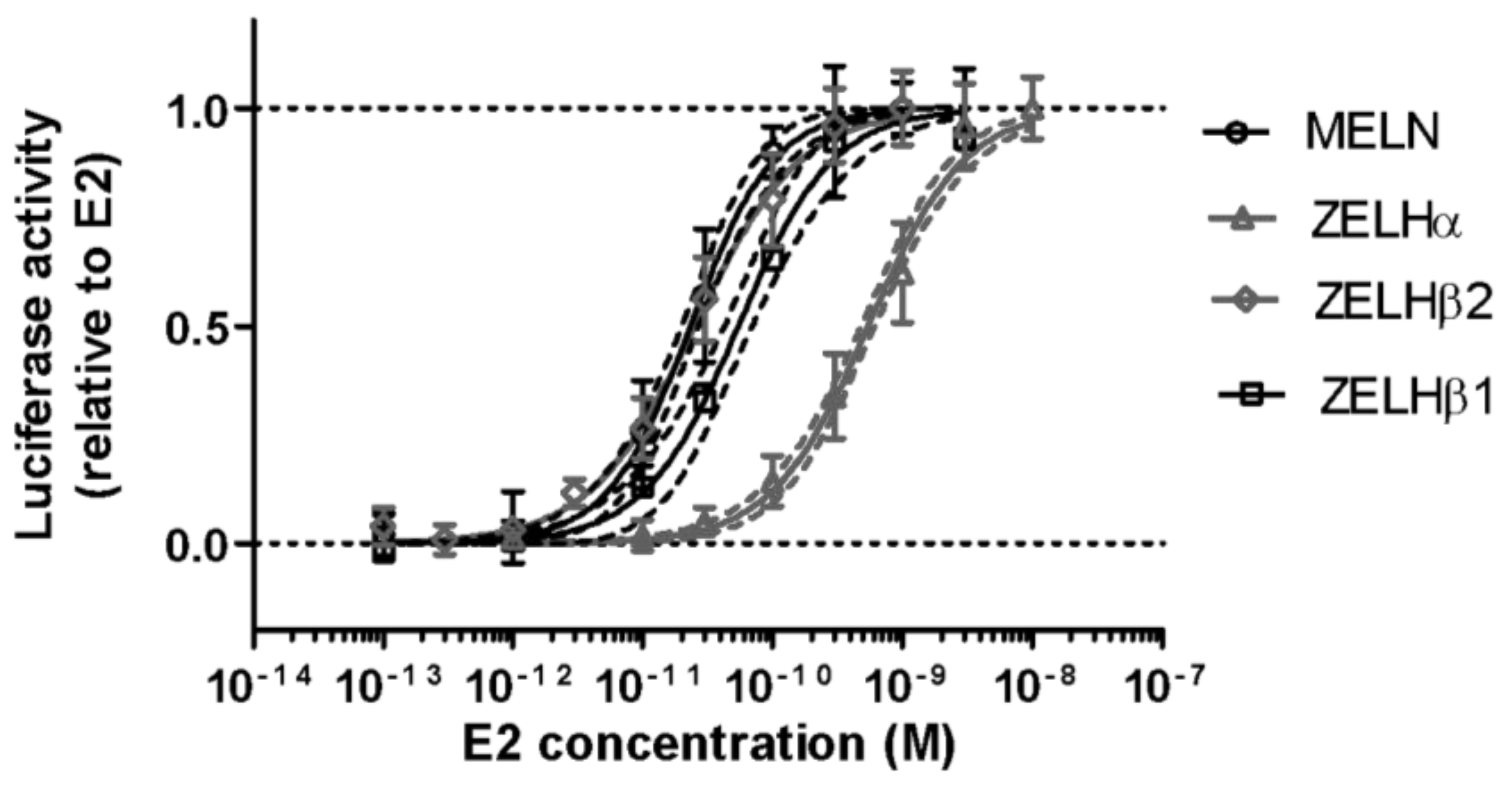
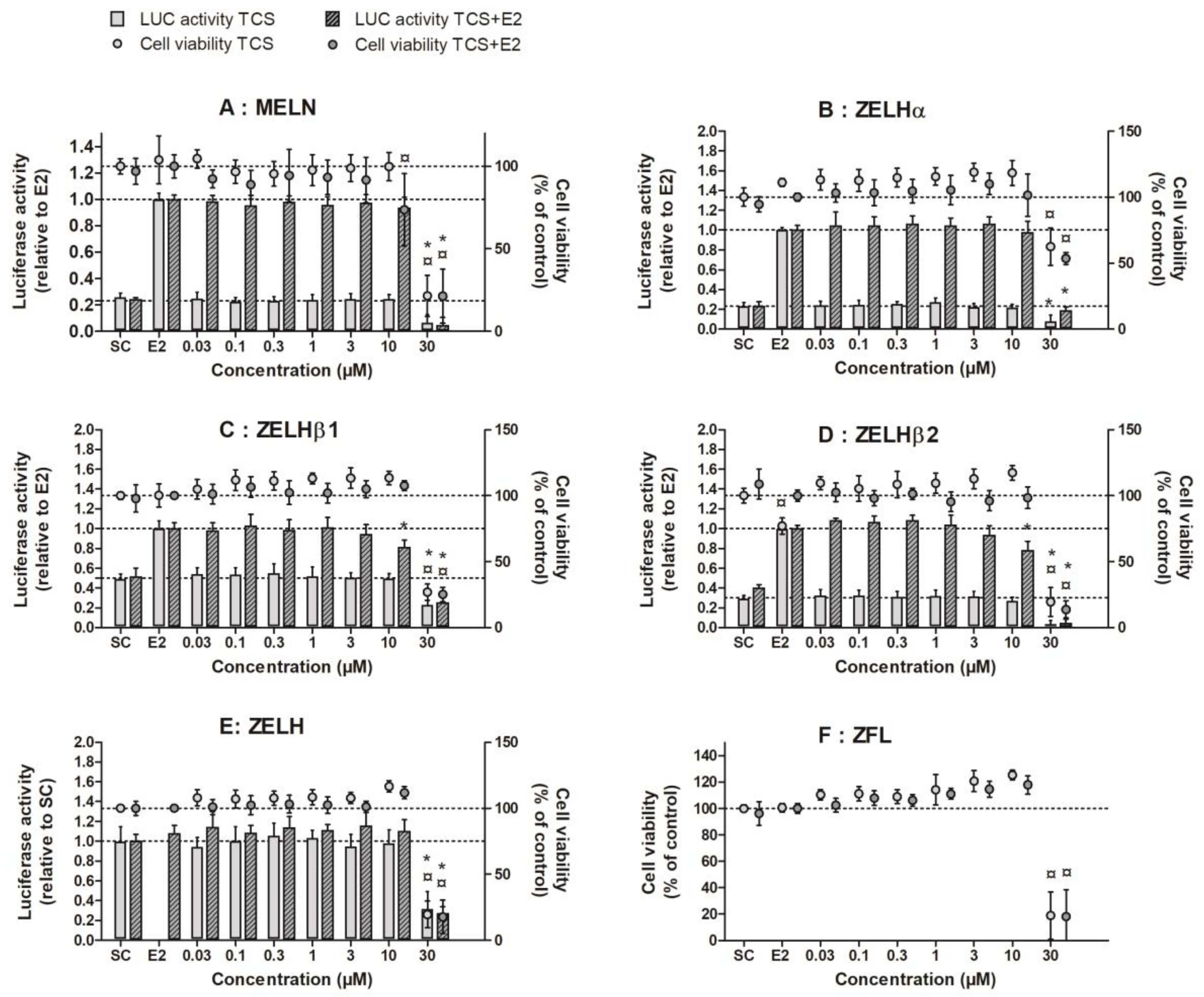
2.1. Triclosan Does Not Alter ZfER Transactivation In Vitro
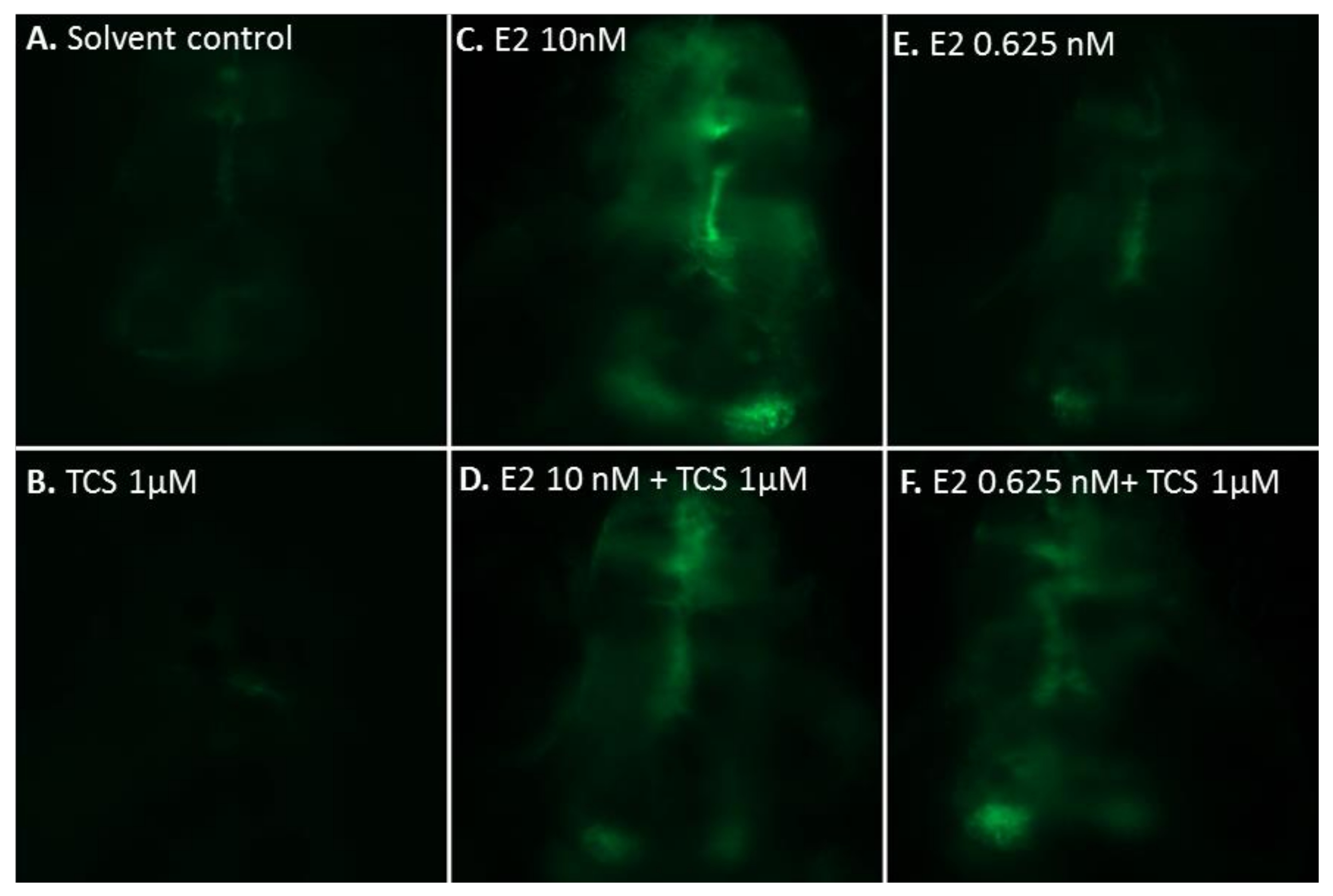
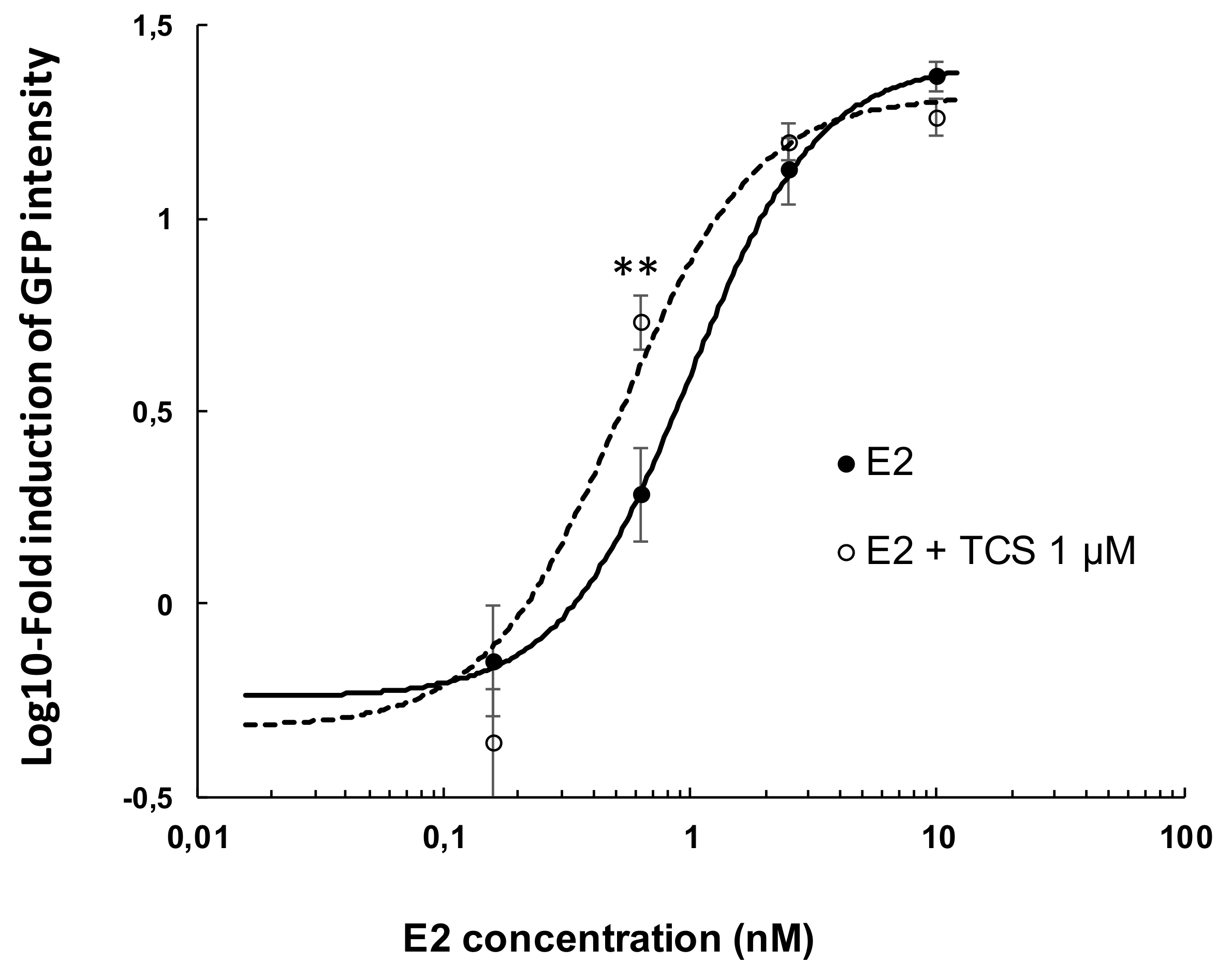
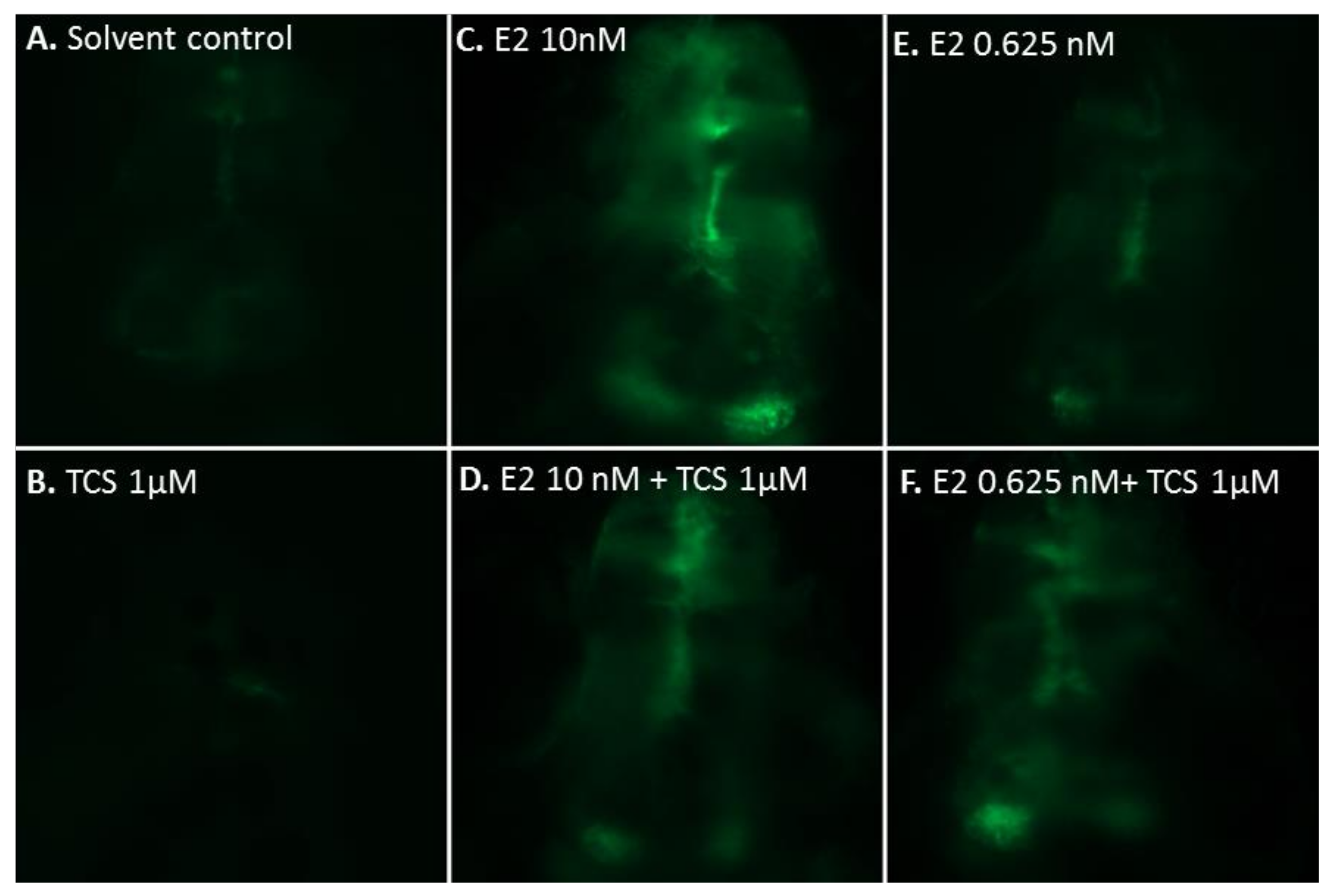
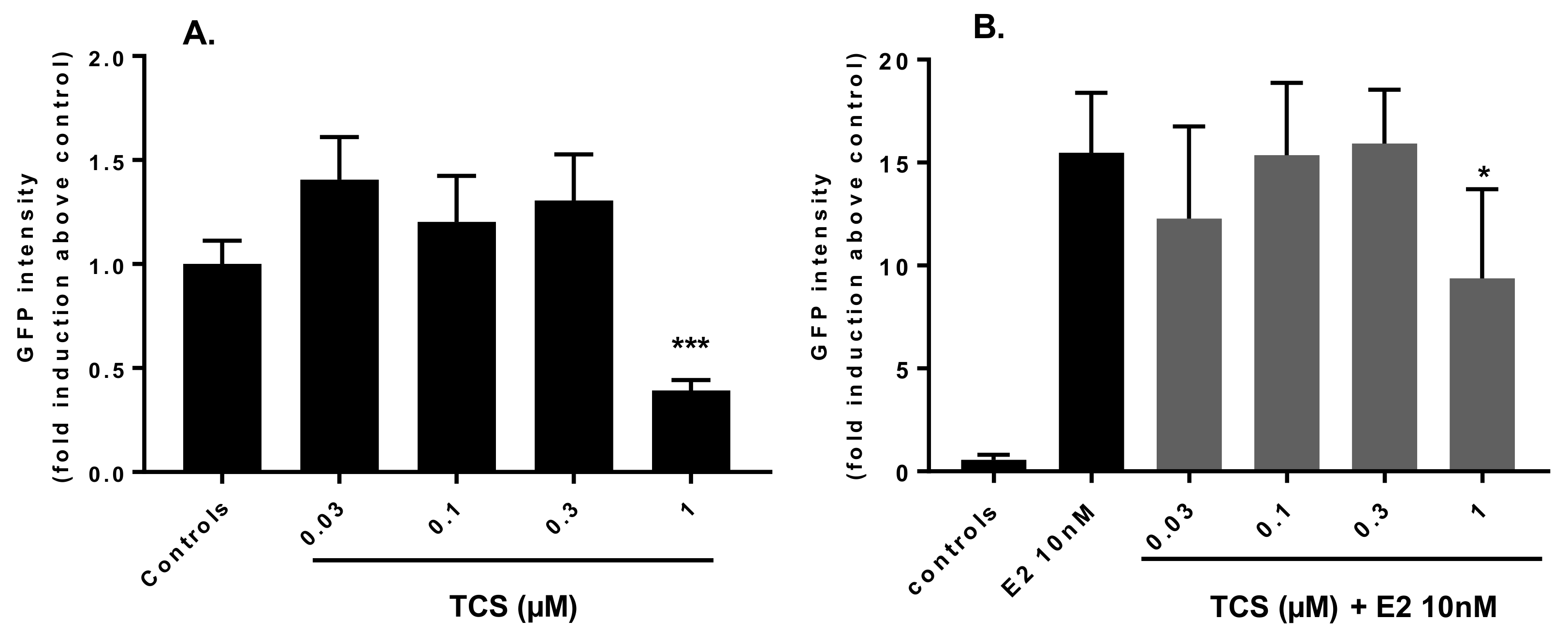
2.2. Effect of Triclosan on Brain Aromatase Expression Using the Cyp19a1b-GFP Transgenic Zebrafish Embryo Assay (EASZY Assay)
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. In Vitro Assays: Cell Culture, Luciferase and Cell Viability Assays
4.3. In Vivo Zebrafish Bioassays
4.4. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- SCCS (Scientific Committee on Consumer Safety). Opinion on Triclosan (Antimicrobial Resistance); SCCS: Brussels, Belgium, 2010. [Google Scholar]
- Dann, A.B.; Hontela, A. Triclosan: Environmental exposure, toxicity and mechanisms of action. J. Appl. Toxicol. 2011, 31, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Carvalho, R.; Antonio, D.C.; Cornero, S.; Locoro, G.; Tavazzi, S.; Paracchini, B.; Ghiani, M.; Lettieri, T.; Blaha, L.; et al. EU-wide monitoring survey on emerging polar organic contaminants in wastewater treatment plant effluents. Water Res. 2013, 47, 6475–6487. [Google Scholar] [CrossRef] [PubMed]
- Brausch, J.M.; Rand, G.M. A review of personal care products in the aquatic environment: Environmental concentrations and toxicity. Chemosphere 2011, 82, 1518–1532. [Google Scholar] [CrossRef] [PubMed]
- Kolpin, D.W.; Furlong, E.T.; Meyer, M.T.; Thurman, E.M.; Zaugg, S.D.; Barber, L.B.; Buxton, H.T. Pharmaceuticals, hormones, and other organic wastewater contaminants in US streams, 1999–2000: A national reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.M.; Bartell, S.E.; Schoenfuss, H.L. Effects of Triclosan and Triclocarban, Two Ubiquitous Environmental Contaminants, on Anatomy, Physiology, and Behavior of the Fathead Minnow (Pimephales promelas). Arch. Environ. Contam. Toxicol. 2012, 63, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Foran, C.M.; Bennett, E.R.; Benson, W.H. Developmental evaluation of a potential non-steroidal estrogen: Triclosan. Mar. Environ. Res. 2000, 50, 153–156. [Google Scholar] [CrossRef]
- Raut, S.A.; Angus, R.A. Triclosan has endocrine-disrupting effects in male western mosquitofish, Gambusia affinis. Environ. Toxicol. Chem. 2010, 29, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, H.; Matsumura, N.; Hirano, M.; Matsuoka, M.; Shiratsuchi, H.; Ishibashi, Y.; Takao, Y.; Arizono, K. Effects of triclosan on the early life stages and reproduction of medaka Oryzias latipes and induction of hepatic vitellogenin. Aquat. Toxicol. 2004, 67, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y.; Yamagishi, T.; Takahashi, H.; Iguchi, T.; Tatarazako, N. Effects of triclosan on Japanese medaka (Oryzias latipes) during embryo development, early life stage and reproduction. J. Appl. Toxicol. 2018, 38, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Du, G.Z.; Zhang, W.; Hu, J.L.; Wu, D.; Song, L.; Xia, Y.K.; Wang, X.R. The in vitro estrogenic activities of triclosan and triclocarban. J. Appl. Toxicol. 2014, 34, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.D.; Fair, P.A. Comparison of in vitro cytotoxicity, estrogenicity and anti-estrogenicity of triclosan, perfluorooctane sulfonate and perfluorooctanoic acid. J. Appl. Toxicol. 2013, 33, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.C.; Zhao, B.; Chen, J.; Cherednichenko, G.; Sanmarti, E.; Denison, M.S.; Lasley, B.; Pessah, I.N.; Kultz, D.; Chang, D.P.Y.; et al. In vitro biologic activities of the antimicrobials triclocarban, its analogs, and triclosan in bioassay screens: Receptor-based bioassay screens. Environ. Health Perspect. 2008, 116, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Gee, R.H.; Charles, A.; Taylor, N.; Darbre, P.D. Oestrogenic and androgenic activity of triclosan in breast cancer cells. J. Appl. Toxicol. 2008, 28, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Tarnow, P.; Tralau, T.; Hunecke, D.; Luch, A. Effects of triclocarban on the transcription of estrogen, androgen and aryl hydrocarbon receptor responsive genes in human breast cancer cells. Toxicol. Vitro 2013, 27, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, S.; Lange, A.; Hirakawa, I.; Tohyama, S.; Ogino, Y.; Mizutani, T.; Kagami, Y.; Kusano, T.; Ihara, M.; Tanaka, H.; et al. Differing Species Responsiveness of Estrogenic Contaminants in Fish Is Conferred by the Ligand Binding Domain of the Estrogen Receptor. Environ. Sci. Technol. 2014, 48, 5254–5263. [Google Scholar] [CrossRef] [PubMed]
- Le Fol, V.; Ait-Aissa, S.; Sonavane, M.; Porcher, J.M.; Balaguer, P.; Cravedi, J.P.; Zalko, D.; Brion, F. In vitro and in vivo estrogenic activity of BPA, BPF and BPS in zebrafish-specific assays. Ecotoxicol. Environ. Saf. 2017, 142, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Sonavane, M.; Creusot, N.; Maillot-Marechal, E.; Pery, A.; Brion, F.; Ait-Aissa, S. Zebrafish-based reporter gene assays reveal different estrogenic activities in river waters compared to a conventional human-derived assay. Sci. Total Environ. 2016, 550, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Cosnefroy, A.; Brion, F.; Maillot-Marechal, E.; Porcher, J.M.; Pakdel, F.; Balaguer, P.; Ait-Aissa, S. Selective Activation of Zebrafish Estrogen Receptor Subtypes by Chemicals by Using Stable Reporter Gene Assay Developed in a Zebrafish Liver Cell Line. Toxicol. Sci. 2012, 125, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Balaguer, P.; Francois, F.; Comunale, F.; Fenet, H.; Boussioux, A.M.; Pons, M.; Nicolas, J.C.; Casellas, C. Reporter cell lines to study the estrogenic effects of xenoestrogens. Sci. Total Environ. 1999, 233, 47–56. [Google Scholar] [CrossRef]
- Brion, F.; Le Page, Y.; Piccini, B.; Cardoso, O.; Tong, S.K.; Chung, B.C.; Kah, O. Screening Estrogenic Activities of Chemicals or Mixtures In vivo Using Transgenic (cyp19a1b-GFP) Zebrafish Embryos. PLoS ONE 2012, 7, e36069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, G.W.; Hallinger, D.R.; Stoker, T.E. The effect of triclosan on the uterotrophic response to extended doses of ethinyl estradiol in the weanling rat. Reprod. Toxicol. 2013, 36, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Park, B.K.; Gonzales, E.L.T.; Yang, S.M.; Bang, M.J.; Choi, C.S.; Shin, C.Y. Effects of Triclosan on Neural Stem Cell Viability and Survival. Biomol. Ther. 2016, 24, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Szychowski, K.A.; Sitarz, A.M.; Wojtowicz, A.K. Triclosan induces fas receptor-dependent apoptosis in mouse neocrotical neurons in vitro. Neuroscience 2015, 284, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Xu, B.; Han, X.M.; Mao, Z.L.; Chen, M.J.; Du, G.Z.; Talbot, P.; Wang, X.R.; Xia, Y.K. The effects of triclosan on pluripotency factors and development of mouse embryonic stem cells and zebrafish. Arch. Toxicol. 2015, 89, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Q.; Wang, Y.Q.; Fillgrove, K.L.; Anderson, V.E. Triclosan inhibits enoyl-reductase of type I fatty acid synthase in vitro and is cytotoxic to MCF-7 and SKBr-3 breast cancer cells. Cancer Chemother. Pharmacol. 2002, 49, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Le Page, Y.; Menuet, A.; Kah, O.; Pakdel, F. Characterization of a cis-acting element involved in cell-specific expression of the zebrafish brain aromatase gene. Mol. Reprod. Dev. 2008, 75, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Menuet, A.; Pellegrini, E.; Brion, F.; Gueguen, M.M.; Anglade, I.; Pakdel, F.; Kah, O. Expression and estrogen-dependent regulation of the zebrafish brain aromatase gene. J. Comp. Neurol. 2005, 485, 304–320. [Google Scholar] [CrossRef] [PubMed]
- Cano-Nicolau, J.; Garoche, C.; Hinfray, N.; Pellegrini, E.; Boujrad, N.; Pakdel, F.; Kah, O.; Brion, F. Several synthetic progestins disrupt the glial cell specific-brain aromatase expression in developing zebra fish. Toxicol. Appl. Pharmacol. 2016, 305, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Neale, P.A.; Altenburger, R.; Ait-Aissa, S.; Brion, F.; Busch, W.; Umbuzeiro, G.d.A.; Denison, M.S.; Du Pasquier, D.; Hilscherova, K.; Hollert, H.; et al. Development of a bioanalytical test battery for water quality monitoring: Fingerprinting identified micropollutants and their Contribution to effects in surface water. Water Res. 2017, 123, 734–750. [Google Scholar] [CrossRef] [PubMed]
- Pinto, P.I.S.; Guerreiro, E.M.; Power, D.M. Triclosan interferes with the thyroid axis in the zebrafish (Danio rerio). Toxicol. Res. 2013, 2, 60–69. [Google Scholar] [CrossRef]
- Schnitzler, J.G.; Frederich, B.; Dussenne, M.; Klaren, P.H.M.; Silvestre, F.; Das, K. Triclosan exposure results in alterations of thyroid hormone status and retarded early development and metamorphosis in Cyprinodon variegatus. Aquat. Toxicol. 2016, 181, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Di Liegro, I. Thyroid hormones and the central nervous system of mammals (Review). Mol. Med. Rep. 2008, 1, 279–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, M. Possible role of glial cells in the relationship between thyroid dysfunction and mental disorders. Front. Cell. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Stoker, T.E.; Gibson, E.K.; Zorrilla, L.M. Triclosan Exposure Modulates Estrogen-Dependent Responses in the Female Wistar Rat. Toxicol. Sci. 2010, 117, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Guo, X.; Chen, W.; Sun, Y.; Fan, C. Effects of triclosan on hormones and reproductive axis in female Yellow River carp (Cyprinus carpio): Potential mechanisms underlying estrogen effect. Toxicol. Appl. Pharmacol. 2017, 336, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, F.; Chen, W.; Xu, R.; Wang, W. Effects of triclosan (TCS) on hormonal balance and genes of hypothalamus-pituitary-gonad axis of juvenile male Yellow River carp (Cyprinus carpio). Chemosphere 2018, 193, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival —Application to proliferation and cyto-toxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Tong, S.K.; Mouriec, K.; Kuo, M.W.; Pellegrini, E.; Gueguen, M.M.; Brion, F.; Kah, O.; Chung, B.C. A cyp19a1b-GFP (Aromatase B) Transgenic Zebrafish Line That Expresses GFP in Radial Glial Cells. Genesis 2009, 47, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Vindimian, E. REGTOX: Macro ExcelTM Pour Dose-Réponse. Available online: http://www.normalesup.org/~vindimian/fr_index.html (accessed on 29 January 2018).

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, H.; Brion, F.; Porcher, J.-M.; Budzinski, H.; Aït-Aïssa, S. Triclosan Lacks (Anti-)Estrogenic Effects in Zebrafish Cells but Modulates Estrogen Response in Zebrafish Embryos. Int. J. Mol. Sci. 2018, 19, 1175. https://doi.org/10.3390/ijms19041175
Serra H, Brion F, Porcher J-M, Budzinski H, Aït-Aïssa S. Triclosan Lacks (Anti-)Estrogenic Effects in Zebrafish Cells but Modulates Estrogen Response in Zebrafish Embryos. International Journal of Molecular Sciences. 2018; 19(4):1175. https://doi.org/10.3390/ijms19041175
Chicago/Turabian StyleSerra, Hélène, François Brion, Jean-Marc Porcher, Hélène Budzinski, and Selim Aït-Aïssa. 2018. "Triclosan Lacks (Anti-)Estrogenic Effects in Zebrafish Cells but Modulates Estrogen Response in Zebrafish Embryos" International Journal of Molecular Sciences 19, no. 4: 1175. https://doi.org/10.3390/ijms19041175