The JAK2 GGCC (46/1) Haplotype in Myeloproliferative Neoplasms: Causal or Random?



Department of Emergency and Organ Transplantation (D.E.T.O.), Hematology Section, University of Bari, 70124 Bari, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(4), 1152; https://doi.org/10.3390/ijms19041152
Submission received: 19 March 2018
/
Revised: 5 April 2018
/
Accepted: 9 April 2018
/
Published: 11 April 2018
(This article belongs to the Section Biochemistry)
Abstract
:The germline JAK2 haplotype known as “GGCC or 46/1 haplotype” (haplotypeGGCC_46/1) consists of a combination of single nucleotide polymorphisms (SNPs) mapping in a region of about 250 kb, extending from the JAK2 intron 10 to the Insulin-like 4 (INLS4) gene. Four main SNPs (rs3780367, rs10974944, rs12343867, and rs1159782) generating a “GGCC” combination are more frequently indicated to represent the JAK2 haplotype. These SNPs are inherited together and are frequently associated with the onset of myeloproliferative neoplasms (MPN) positive for both JAK2 V617 and exon 12 mutations. The association between the JAK2 haplotypeGGCC_46/1 and mutations in other genes, such as thrombopoietin receptor (MPL) and calreticulin (CALR), or the association with triple negative MPN, is still controversial. This review provides an overview of the frequency and the role of the JAK2 haplotypeGGCC_46/1 in the pathogenesis of different myeloid neoplasms and describes the hypothetical mechanisms at the basis of the association with JAK2 gene mutations. Moreover, possible clinical implications are discussed, as different papers reported contrasting data about the correlation between the JAK2 haplotypeGGCC_46/1 and blood cell count, survival, or disease progression.
1. Introduction
Classic mutations of Janus Kinase 2 gene (JAK2), such as V617F in exon 14 and a large spectrum of mutations in exon 12, represent the molecular hallmark of polycythemia vera (PV), where they are found in more than 95% of patients [1,2,3,4]. The JAK2 V617F mutation is also detected in approximately 50–60% of patients with primary myelofibrosis (PMF) or essential thrombocythemia (ET) [5]. To investigate the possible interplay between somatically acquired gene mutations and inherited genetic variations in patients with myeloproliferative neoplasms (MPNs), Pardanani et al. in 2008 studied the role of single nucleotide polymorphisms (SNPs) within four candidate genes involved in the JAK-Signal transducer and activator of transcription (STAT) signaling pathway, including receptors for erythropoietin (EPOR), thrombopoietin (MPL), granulocyte colony stimulating factor (GCSFR), and JAK2 [6]. This study revealed for the first time a significant association between the occurrence of specific SNPs in JAK2 gene and the onset of different MPNs [6]. Subsequently, in 2009, three independent groups revealed an interesting and strong association between the risk of developing a JAK2 V617F positive MPN and a germline haplotype including the 3′ portion of JAK2, named GGCC or 46/1 haplotype (haplotypeGGCC_46/1) [7,8,9]. The term haplotype stands for “haploid genotype” and refers to a group of genetic markers, generally represented by different SNPs, mapping on the same chromosome that are inherited together, as they are not separated by meiotic or mitotic recombination; having one of these genetic markers usually implies having all the others as well.
2. Genomic Architecture of the JAK2 HaplotypeGGCC_46/1
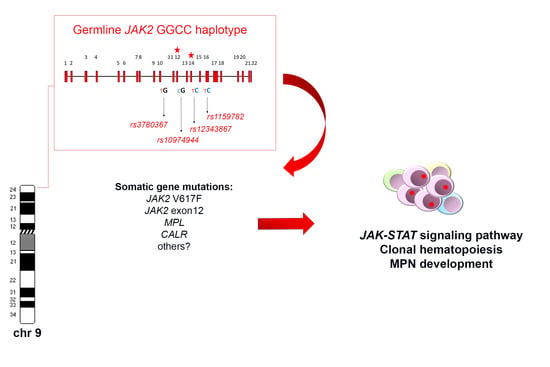
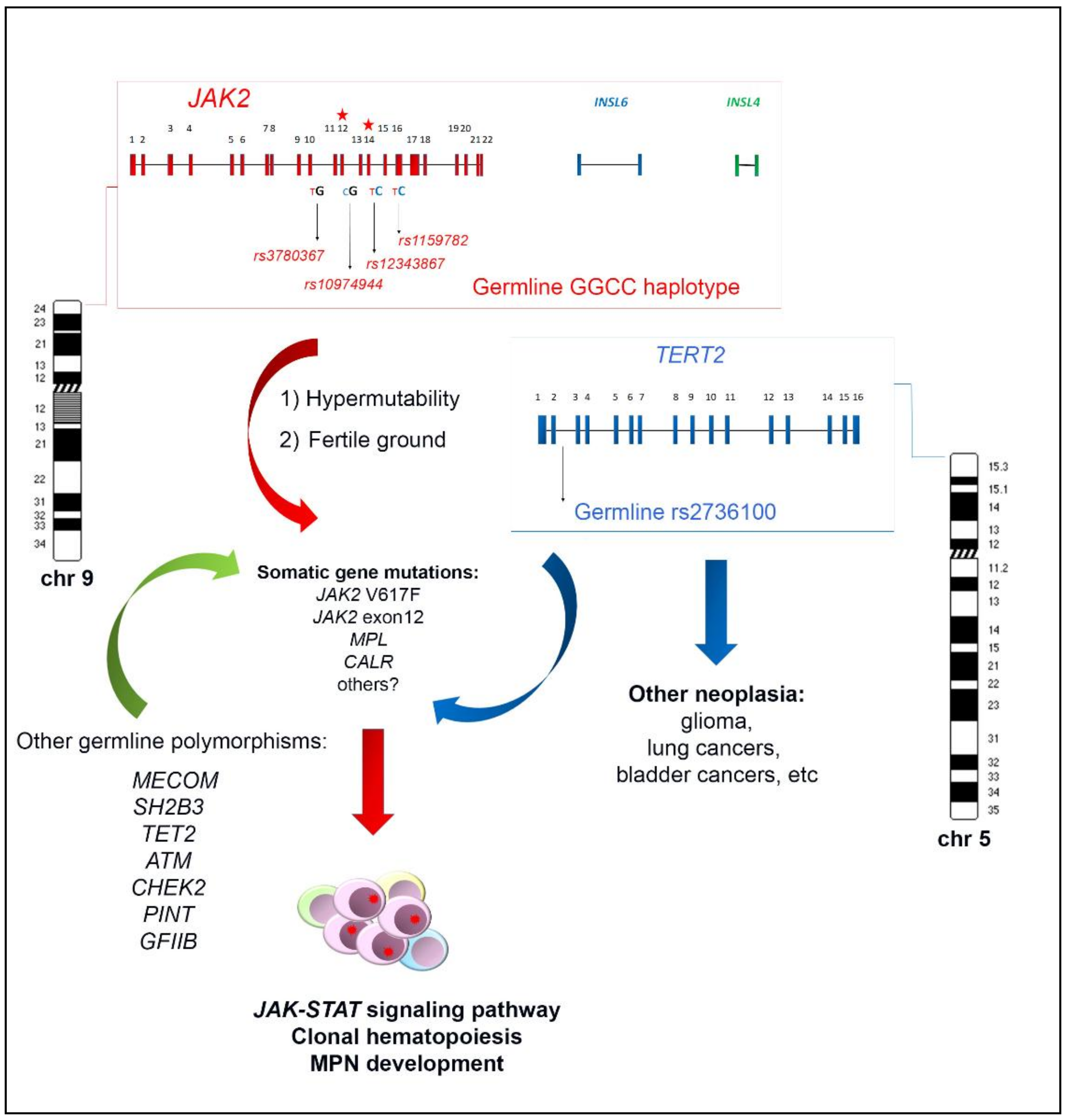
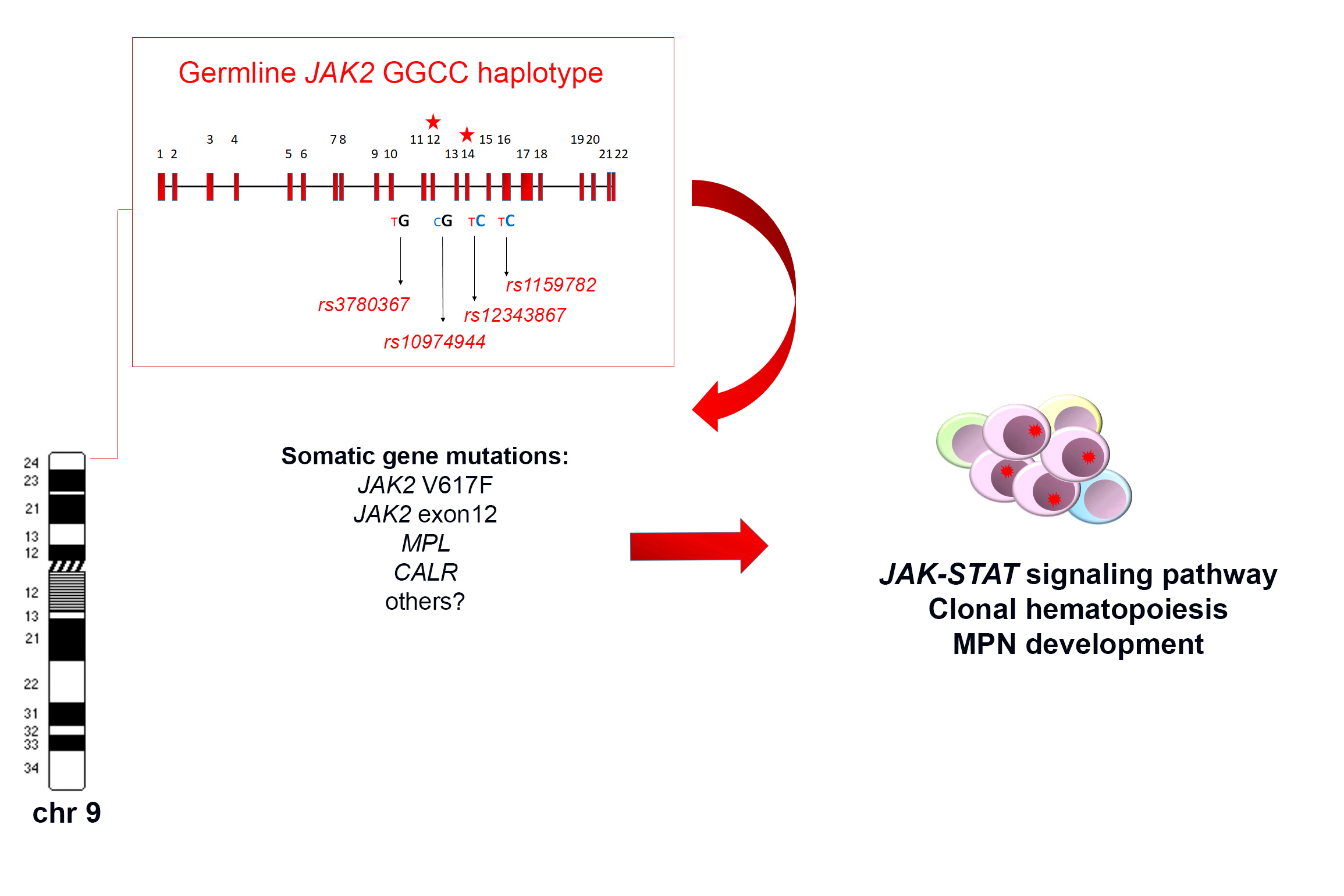
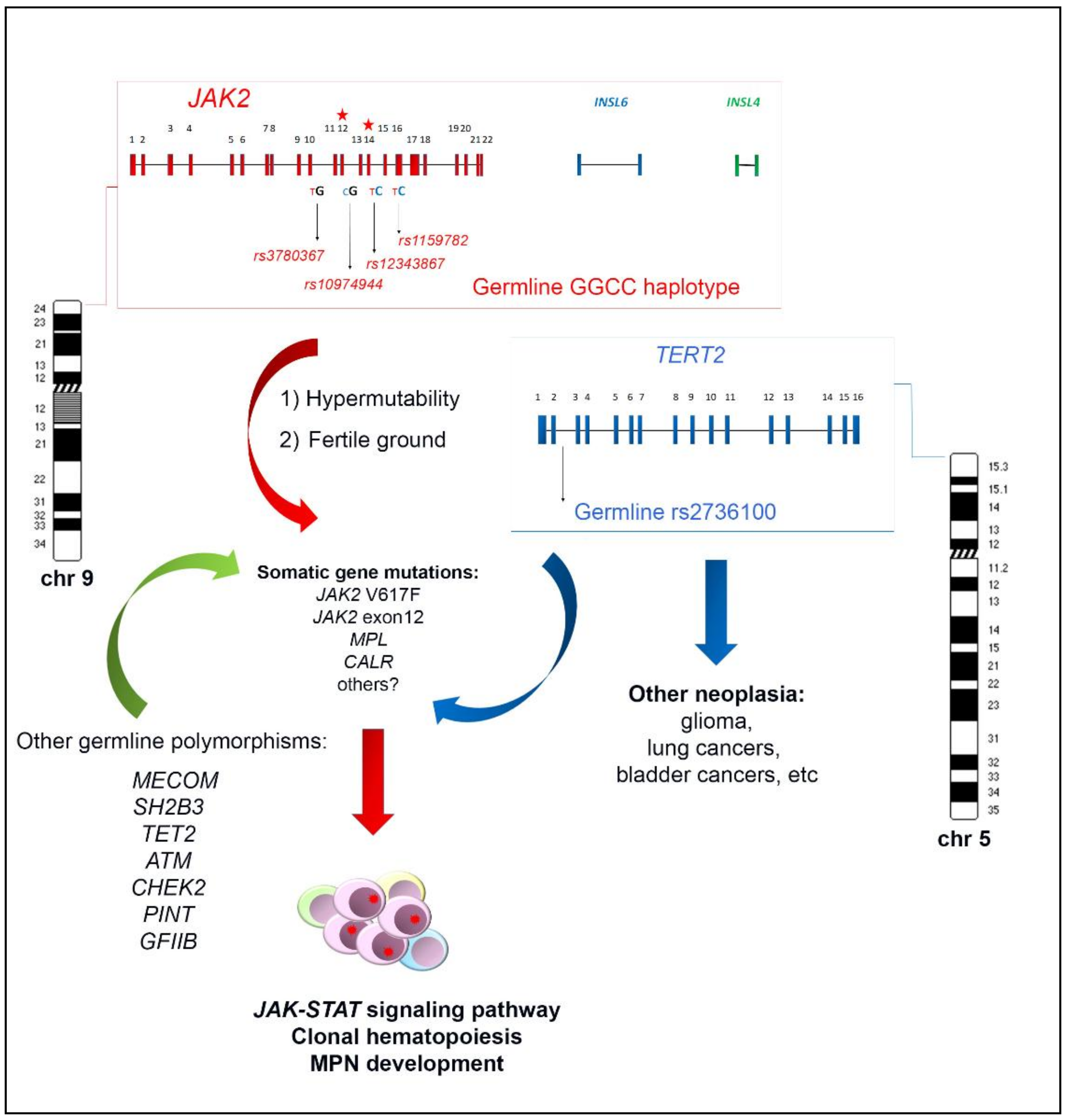
The haplotypeGGCC_46/1 consists of hundreds of SNPs mapping in a region of about 250–280 Kb on chromosome 9p and including the three genes JAK2, Insulin-like 6 (INSL6) and Insulin-like 4 (INSL4) [8,10,11] (Figure 1). Among these genes, only JAK2 is expressed in the hematopoietic tissue, whereas INSL6 is preferentially expressed in the testis where it plays a role in sperm development and fertilization, and INSL4 is highly expressed in the early placenta and influences trophoblast development and bone formation. The promoter region of JAK2 gene is not included in the haplotypeGGCC_46/1 as the genotyped SNPs in this region are not in linkage disequilibrium (LD) with the haplotype [8]. The JAK2 46/1 haplotype is also referred to as the “GGCC” haplotype, as can be represented by four main SNPs (rs3780367, rs10974944, rs12343867, and rs1159782) that replace three thymidines (T) and one cytosine (C) by two guanosines (G) and two cytosines, generating a “GGCC” combination (Figure 1) [8]. These four SNPs map on JAK2 introns 10, 12, 14, and 15, respectively, and are always inherited together, being in complete linkage disequilibrium (Figure 1). As regards the origin of the “46/1 haplotype” definition, it derives from the study by Jones et al. that determined the haplotype structure of the JAK2 gene using 14 SNPs genotyped by the Wellcome Trust Case Control Consortium (WTCCC) in 1500 healthy blood donors. Nine haplotypes were found to account for 94% of JAK2 alleles and, among these, two haplotypes (numbers 46 and 1) were found to have a combined frequency of 0.24 in healthy individuals and to be identical except for one SNP [7]. These two haplotypes were found to be more frequently associated with JAK2 V617F positive MPNs [7].
3. Frequency of the HaplotypeGGCC_46/1 in MPN and Other Myeloid Neoplasms
The frequency of the JAK2 haplotypeGGCC_46/1 in the healthy population is about 24%, whereas it was found in 40–80% of JAK2 V617F positive MPN [7,8,12,13,14,15], in about 64% of cases bearing JAK2 exon 12 mutations [10], and in approximately 36% of MPN cases bearing MPL mutations [16]. However, these last data were not confirmed by a subsequent study that found no association between the JAK2 haplotype and MPL mutations [17]. The possible role of the JAK2 haplotypeGGCC_46/1 in JAK2 V617F negative MPNs is still controversial, as some authors did not identify significant association in this group of patients [7,18]. On the other hand, other studies identified a weak association between the haplotypeGGCC_46/1 and JAK2 V617F negative MPN, suggesting that the presence of this germline condition confers a more generalized predisposition to MPN development, independently of the V617F mutation [7,12,16,19,20]. However, in 2010, calreticulin (CALR) gene mutations had not yet been identified, therefore the mutational status of V617F negative TE and MF patients could not be correctly assessed. After the identification of CALR mutations, further studies produced conflicting results regarding the frequency of the haplotypeGGCC_46/1 in this group of MPN patients, mostly suggesting a lack of association [14,21,22,23]. Therefore, the possible association between the occurrence of the haplotypeGGCC_46/1 and JAK2 V617F negative MPN cases warrants further investigation. The presence of the haplotypeGGCC_46/1 has also been investigated in chronic myeloid leukemia (CML) but no significantly increased frequency was revealed [24]. Analysis of the haplotypeGGCC_46/1 in acute myeloid leukemia (AML) patients showed that the allele frequency did not significantly differ as compared to normal controls; however, an altered haplotypeGGCC_46/1 frequency was observed in AML patients with and without normal karyotype (NK) (33–34% versus 12–25%) [13,25].
4. The Role of the JAK2 HaplotypeGGCC_46/1 and Other Germ Line Variants in Familial and Sporadic MPNs
As reviewed, the JAK2 haplotypeGGCC_46/1 is a germline inherited condition which confers a predisposition and an increased risk of developing MPN, preferentially but not exclusively in association with the JAK2 V617F mutation (Figure 1). The identification of this germline variant raised the hypothesis that this inherited predisposition might explain familial MPN clustering. However, different studies investigated the haplotypeGGCC_46/1 allele frequency in both familial and sporadic MPN in comparison with normal controls, without finding a significant difference in these two groups [26,27]. These findings suggest that that the JAK2 haplotypeGGCC_46/1 does not explain familial MPNs, which account for 5–10% of all MPN cases [28,29,30,31]. At the same time, it was found that the rs2736100 SNP, located in the second intron of the telomerase reverse transcriptase (TERT) gene, had a different allele frequency in familial MPN compared to sporadic cases [27] and exhibited a strong cancer predisposition effect in all MPN subtypes, regardless of the JAK2 gene mutations occurrence [23,32]. The TERT gene at 5p15.33 encodes the catalytic subunit of the telomerase complex, playing an important role in maintaining telomere length [33]. Germline mutations in the TERT gene lead to dyskeratosis congenita, a disorder characterized by cancer susceptibility due to telomeres shortening [34]. The rs2736100_C allele of the TERT gene was previously shown to be associated with an elevated risk for several other neoplasia, such as glioma, lung and bladder cancers (Figure 1) [35,36,37,38]. A recent meta-analysis confirmed that the TERT rs2736100 polymorphism is associated with increased overall cancer risk, including solid cancers, myeloproliferative neoplasms, and acute myeloid leukemia [39]. The JAK2 haplotypeGGCC_46/1 and TERT rs2736100_C are independent factors predisposing to MPN and confering an additional disease risk (Figure 1) [22,27,40,41]. Moreover, it has been shown that MPN patients carrying TERT rs2736100_C have an increased risk of developing solid tumors, especially if treated with cytoreductive therapy [40]. A recent genome-wide association study was performed with a SNP array platform to identify novel predisposition alleles associated with the onset of MPNs and JAK2 V617F clonal hematopoiesis in the general population [42]. This study confirmed the role of the JAK2 haplotypeGGCC_46/1 and TERT SNPs as germline factors predisposing to MPN but also identified significant associations between the occurrence of polymorphisms near SH2B3, TET2, ATM, CHEK2, PINT, and GFI1B genes and JAK2 V617F clonal hematopoiesis and/or MPN development (Figure 1) [42]. Another recent study investigated the contribution of additional germline polymorphisms, such as MECOM rs2201862, HBS1L-MYB rs9376092 and THRB-RARB rs4858647, to the onset of MPN [23]. The SNP rs2201862, located downstream of MECOM gene, had the third strongest influence on the risk of developing MPN, after the JAK2 46/1 haplotype and TERT rs2736100 polymorphisms, MECOM rs2201862 was found to predispose especially to PV and to CALR mutated ET and PMF (Figure 1) [23].
5. Clinical Implications of the JAK246/1_GGCC Haplotype
The majority of studies failed to detect any association of the JAK2 haplotypeGGCC_46/1 with the distribution of age, sex or clinical parameters, including hemoglobin level, leukocyte count, rate of thrombosis/disease transformation and survival, suggesting that the JAK2 haplotypeGGCC_46/1 does not seem to affect the clinical phenotype or prognosis (Table 1) [12,19,20,43]. Preliminary data supporting a functional difference between alleles bearing or not the JAK2 haplotypeGGCC_46/1 were provided by Jones’ study in 2009, which investigated whether the haplotypeGGCC_46/1 influences myeloid colony formation in healthy individuals showing that the presence of at least one 46/1 allele is associated with fewer circulating granulocyte-macrophage progenitor cells than are present without the 46/1 allele [7]. Another study showed that normal individuals bearing the JAK2 haplotypeGGCC_46/1 had slightly increased erythrocyte and decreased platelet count as compared to non-carriers [44]. However, these findings were not confirmed by further genome-wide association studies [45,46,47]. Tefferi et al., in 2010, found that MF patients negative for the JAK2 haplotypeGGCC_46/1 had a significantly shorter survival as compared to MF cases bearing the haplotype [12]; these data, however, were not confirmed by a subsequent analysis [20], whereas the study by Martínez-Trillos et al. in 2014 showed that MF patients with a homozygous 46/1 haplotype had significantly higher hemoglobin values and higher leukocyte counts but no association with other clinical characteristics [48]. As regards the prognostic significance of the JAK2 haplotypeGGCC_46/1 in AML cases, it was demonstrated that cases with a normal karyotype bearing the haplotypeGGCC_46/1 showed a trend towards myelomonocytic proliferation and shorter disease-free survival and overall survival compared to GGCC_46/1 non carriers; on the other hand, the haplotypeGGCC_46/1 had no impact on prognosis in the subgroup of AML with an abnormal karyotype [13,25]. Moreover, interesting evidence reported an increased frequency of the JAK2 haplotypeGGCC_46/1 in patients with MPN characterized by splanchnic vein thrombosis (SVT), both in the presence and absence of the JAK2 V617F mutation [49,50,51,52,53]. A meta-analysis was performed on 26 observational studies involving 8561 cases, which showed that the JAK2 haplotypeGGCC_46/1 significantly raised the risk of development of MPNs and SVT [54]. Other evidence suggested a significant association between some SNPs included in the JAK2 haplotypeGGCC_46/1 (e.g., rs10758669) and inflammatory disorders such as ulcerative colitis and Crohn's disease [55,56,57]. In this regard, it is known that the JAK2-STAT3 pathway is one of the most important cell signaling pathways in Crohn’s disease, being activated by the production of IL-6 and is responsible for the production of pro-inflammatory proteins [58,59]. A possible explanation is that the haplotypeGGCC_46/1 may cause an excessive production of cytokines with pro-inflammatory action which would further impair immune responses [11,60]. A recent study investigated the association of recipient and donor JAK2 haplotypeGGCC_46/1 and the outcome of allogeneic hematopoietic stem cell transplantation (allo-HSCT) in a series of 124 AML patients [60]. The findings from this study suggest that the occurrence of the JAK2 haplotypeGGCC_46/1 in both recipients and donors significantly affected the development of acute graft-versus-host disease, confirming that JAK2 polymorphisms may have an influence on cytokines signaling pathways [61].
6. Correlation between the JAK2 HaplotypeGGCC_46/1 and JAK2 V617F Allele Burden
Several reports showed that the haplotypeGGCC_46/1 seems to be associated with a high mutant allele burden in JAK2 V617F positive MPN patients, being significantly enriched in patients with higher V617F allele burden [12,18,19,20,21]. These data suggest that the JAK2 haplotypeGGCC_46/1 could confer a possible selective advantage to the V617F mutant clone as well as promoting the acquisition of JAK2 V617F mutation. Moreover, the JAK2 haplotypeGGCC_46/1 might have an influence on the occurrence of the 9p mitotic recombination, causing homozygosity of the JAK2 V617F mutation [18]. Alvarez-Larrán et al. investigated the influence of the JAK2 haplotypeGGCC_46/1 during disease follow-up revealing that PV patients homozygous for the haplotype show a progressive and higher increase in the JAK2 V617F allele burden during the disease evolution without cytoreductive therapy, as compared to patients with a negative or heterozygous haplotype [43]. However, a recent report found no significant difference in the JAK2 haplotypeGGCC_46/1 frequency between groups of MPN patients with different JAK2 V617F allele burdens [15], therefore this aspect remains to be further clarified.
7. Potential Mechanisms Explaining the Association between the JAK2 HaplotypeGGCC_46/1 and JAK2 V617F Mutation
There are two principal hypotheses that aim to explain the association between the germline haplotypeGGCC_46/1 and the JAK2 V617F somatic mutation (Figure 1). The “hypermutability hypothesis” derives from the observation that MPN patients heterozygous for the haplotypeGGCC_46/1 preferentially acquire the V617F mutation in cis with the GGCC predisposition allele [7,8,9]; this germline haplotype may somehow determine an increase in the mutation rate at the JAK2 locus and those mutations that confer a selective advantage, such as JAK2 V617F, would cause a clonal myeloproliferative disorder [9,62]. JAK2 exon 12 mutations are also preferentially acquired in cis with this haplotype, supporting the hypermutability hypothesis. It is possible that a “regulatory environment” of an unknown nature is present on the haplotypeGGCC_46/1 and renders DNA more susceptible to damage or replication errors; this could promote the acquisition of gene mutations in cis and induce clonal expansion and the onset of MPNs. It is also plausible that this cis regulatory environment could promote alterations of gene expression, although the haplotypeGGCC_46/1 does not include the promoter region of the JAK2 gene. However, no genotype-specific differences in JAK2 gene expression were observed [7,9,63]. Moreover, it was shown that the SNPs within the haplotypeGGCC_46/1 are not in LD with nonsynonymous SNPs that might alter protein function and structure [8]. It cannot be excluded that the haplotypeGGCC_46/1 is associated with an altered expression of INSL6 or INSL4 genes, which are not normally expressed in hematopoietic cells; their abnormal activation could eventually lead to an altered cytokine production, concurring in stimulation of inflammation pathways [11], but further studies are needed to clarify this matter. The second possible explanation, named the “fertile ground hypothesis”, suggests that JAK2 mutations arise on all haplotypes at the same rate, but the GGCC_46/1 allele confers a selective advantage to the JAK2 V617F positive clone. In this hypothesis, the haplotypeGGCC_46/1 could provide a global DNA propensity for gene mutations and MPN development, as some conflicting evidence indicates that this haplotype could also be associated to MPL or CALR gene mutations [16,17,22,62]. Moreover, this second hypothesis could explain the unclear observations that the haplotype may also be enriched in individuals with JAK2 V617F negative MPNs [7,12,13,19,20,39]. The “fertile ground hypothesis” could also explain the acquisition of mutations in other genes that are critical for expansion and differentiation of myeloid cells, possibly causing malignant transformation to MPN or AML [13].
8. Conclusions
About ten years after its discovery, the possible pathogenic role of the JAK2 haplotypeGGCC_46/1 in MPN patients, as well as in other myeloid malignancies, is not yet understood. The association with JAK2 mutations has been largely confirmed, whereas there is no agreement about the frequency in JAK2 V617F negative or MPL and CALR mutated MPN patients. The most probable explanation is that the JAK2 haplotypeGGCC_46/1 could influence the activation of the constitutive JAK-STAT signaling pathway rather than a specific mutation, supporting clonal hematopoietic proliferation. The mechanism of this possible activation, however, needs further investigations.
Acknowledgments
The authors would like to thank Mary Victoria Pragnell, B.A. for language revision of the manuscript. This work was supported by “Associazione Italiana contro le Leucemie (AIL)-BARI”.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| PV | polycythemia vera |
| PMF | primary myelofibrosis |
| ET | essential thrombocythemia |
| MPN | myeloproliferative neoplasms |
| SNP | single nucleotide polymorphisms |
| EPOR | receptor for erythropoietin |
| MPL | receptors for thrombopoietin |
| GCSFR | colony stimulating factor |
| INSL4 | insulin-like 4 |
| INSL6 | insulin-like 6 |
| WTCCC | Wellcome Trust Case Control Consortium |
| CALR | calreticulin |
| CML | chronic myeloid leukemia |
| AML | acute myeloid leukemia |
| NK | normal karyotype |
| TERT | telomerase reverse transcriptase |
| allo-HSCT | allogeneic hematopoietic stem cell transplantation |
References
- Baxter, E.J.; Scott, L.M.; Campbell, P.J.; East, C.; Fourouclas, N.; Swanton, S.; Vassiliou, G.S.; Bench, A.J.; Boyd, E.M.; Curtin, N.; et al. Cancer Genome Project Acquired mutation of the tyrosine kinase JAK2 in human myeloproliferative disorders. Lancet 2005, 365, 1054–1061. [Google Scholar] [CrossRef]
- Kralovics, R.; Passamonti, F.; Buser, A.S.; Teo, S.-S.; Tiedt, R.; Passweg, J.R.; Tichelli, A.; Cazzola, M.; Skoda, R.C. A Gain-of-function mutation of JAK2 in myeloproliferative disorders. N. Engl. J. Med. 2005, 352, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Tefferi, A.; Gilliland, D.G. Oncogenes in myeloproliferative disorders. Cell Cycle 2007, 6, 550–566. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.M.; Tong, W.; Levine, R.L.; Scott, M.A.; Beer, P.A.; Stratton, M.R.; Futreal, P.A.; Erber, W.N.; McMullin, M.F.; Harrison, C.N.; et al. JAK2 exon 12 mutations in polycythemia vera and idiopathic erythrocytosis. N. Engl. J. Med. 2007, 356, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L. X-inactivation-based clonality analysis and quantitative JAK2V617F assessment reveal a strong association between clonality and JAK2V617F in PV but not ET/MMM, and identifies a subset of JAK2V617F-negative ET and MMM patients with clonal hematopoiesis. Blood 2006, 107, 4139–4141. [Google Scholar] [CrossRef] [PubMed]
- Pardanani, A.; Fridley, B.L.; Lasho, T.L.; Gilliland, D.G.; Tefferi, A. Host genetic variation contributes to phenotypic diversity in myeloproliferative disorders. Blood 2008, 111, 2785–2789. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.V.; Chase, A.; Silver, R.T.; Oscier, D.; Zoi, K.; Wang, Y.L.; Cario, H.; Pahl, H.L.; Collins, A.; Reiter, A.; et al. JAK2 haplotype is a major risk factor for the development of myeloproliferative neoplasms. Nat. Genet. 2009, 41, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Olcaydu, D.; Harutyunyan, A.; Jäger, R.; Berg, T.; Gisslinger, B.; Pabinger, I.; Gisslinger, H.; Kralovics, R. A common JAK2 haplotype confers susceptibility to myeloproliferative neoplasms. Nat. Genet. 2009, 41, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Kilpivaara, O.; Mukherjee, S.; Schram, A.M.; Wadleigh, M.; Mullally, A.; Ebert, B.L.; Bass, A.; Marubayashi, S.; Heguy, A.; Garcia-Manero, G.; et al. A germline JAK2 SNP is associated with predisposition to the development of JAK2(V617F)-positive myeloproliferative neoplasms. Nat. Genet. 2009, 41, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Olcaydu, D.; Skoda, R.C.; Looser, R.; Li, S.; Cazzola, M.; Pietra, D.; Passamonti, F.; Lippert, E.; Carillo, S.; Girodon, F.; et al. The “GGCC” haplotype of JAK2 confers susceptibility to JAK2 exon 12 mutation-positive polycythemia vera. Leukemia 2009, 23, 1924–1926. [Google Scholar] [CrossRef] [PubMed]
- Hermouet, S.; Vilaine, M. The JAK2 46/1 haplotype: A marker of inappropriate myelomonocytic response to cytokine stimulation, leading to increased risk of inflammation, myeloid neoplasm, and impaired defense against infection? Haematologica 2011, 96, 1575–1579. [Google Scholar] [CrossRef] [PubMed]
- Tefferi, A.; Lasho, T.L.; Patnaik, M.M.; Finke, C.M.; Hussein, K.; Hogan, W.J.; Elliott, M.A.; Litzow, M.R.; Hanson, C.A.; Pardanani, A. JAK2 germline genetic variation affects disease susceptibility in primary myelofibrosis regardless of V617F mutational status: Nullizygosity for the JAK2 46/1 haplotype is associated with inferior survival. Leukemia 2010, 24, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Andrikovics, H.; Nahajevszky, S.; Koszarska, M.; Meggyesi, N.; Bors, A.; Halm, G.; Lueff, S.; Lovas, N.; Matrai, Z.; Csomor, J.; et al. JAK2 46/1 haplotype analysis in myeloproliferative neoplasms and acute myeloid leukemia. Leukemia 2010, 24, 1809–1813. [Google Scholar] [CrossRef] [PubMed]
- Soler, G.; Bernal-Vicente, A.; Antón, A.I.; Torregrosa, J.M.; Caparrós-Pérez, E.; Sánchez-Serrano, I.; Martínez-Pérez, A.; Sánchez-Vega, B.; Vicente, V.; Ferrer-Marin, F. The JAK2 46/1 haplotype does not predispose to CALR-mutated myeloproliferative neoplasms. Ann. Hematol. 2015, 94, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Stolyar, M.A.; Klimova, O.A.; Gorbenko, A.S.; Brenner, E.V.; Titov, S.E.; Ivanov, M.K.; Olkhovskiy, I.A. JAK2 haplotype 46/1 and JAK2 V617F allele burden in MPN: New evidence against the “hypermutability” hypothesis? Int. J. Lab. Hematol. 2018, 40, e8–e10. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.V.; Campbell, P.J.; Beer, P.A.; Schnittger, S.; Vannucchi, A.M.; Zoi, K.; Percy, M.J.; McMullin, M.F.; Scott, L.M.; Tapper, W.; et al. The JAK2 46/1 haplotype predisposes to MPL-mutated myeloproliferative neoplasms. Blood 2010, 115, 4517–4523. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, M.M.; Lasho, T.L.; Finke, C.M.; Gangat, N.; Caramazza, D.; Siragusa, S.; Hanson, C.A.; Pardanani, A.; Tefferi, A. MPL mutation effect on JAK2 46/1 haplotype frequency in JAK2V617F-negative myeloproliferative neoplasms. Leukemia 2010, 24, 859–860. [Google Scholar] [CrossRef] [PubMed]
- Trifa, A.P.; Cucuianu, A.; Petrov, L.; Urian, L.; Militaru, M.S.; Dima, D.; Pop, I.V.; Popp, R.A. The G allele of the JAK2 rs10974944 SNP, part of JAK2 46/1 haplotype, is strongly associated with JAK2 V617F-positive myeloproliferative neoplasms. Ann. Hematol. 2010, 89, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Pardanani, A.; Lasho, T.L.; Finke, C.M.; Gangat, N.; Wolanskyj, A.P.; Hanson, C.A.; Tefferi, A. The JAK2 46/1 haplotype confers susceptibility to essential thrombocythemia regardless of JAK2V617F mutational status-clinical correlates in a study of 226 consecutive patients. Leukemia 2010, 24, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Guglielmelli, P.; Biamonte, F.; Spolverini, A.; Pieri, L.; Isgrò, A.; Antonioli, E.; Pancrazzi, A.; Bosi, A.; Barosi, G.; Vannucchi, A.M. Frequency and clinical correlates of JAK2 46/1 (GGCC) haplotype in primary myelofibrosis. Leukemia 2010, 24, 1533–1537. [Google Scholar] [CrossRef] [PubMed]
- Gau, J.-P.; Chen, C.-C.; Chou, Y.-S.; Liu, C.-J.; Yu, Y.-B.; Hsiao, L.-T.; Liu, J.-H.; Hsu, H.-C.; Chiou, T.-J.; Chen, P.-M.; et al. No increase of JAK2 46/1 haplotype frequency in essential thrombocythemia with CALR mutations: Functional effect of the haplotype limited to allele with JAK2V617F mutation but not CALR mutation. Blood Cells Mol. Dis. 2015, 55, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Trifa, A.P.; Bănescu, C.; Tevet, M.; Bojan, A.; Dima, D.; Urian, L.; Török-Vistai, T.; Popov, V.M.; Zdrenghea, M.; Petrov, L.; et al. TERT rs2736100 A>C SNP and JAK2 46/1 haplotype significantly contribute to the occurrence of JAK2 V617F and CALR mutated myeloproliferative neoplasms—A multicentric study on 529 patients. Br. J. Haematol. 2016, 174, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Trifa, A.P.; Bănescu, C.; Bojan, A.S.; Voina, C.M.; Ștefana, P.; Vișan, S.; Ciubean, A.D.; Tripon, F.; Dima, D.; Popov, V.M.; et al. MECOM, HBS1L-MYB, THRB-RARB, JAK2, and TERT polymorphisms defining the genetic predisposition to myeloproliferative neoplasms: A study on 939 patients. Am. J. Hematol. 2018, 93, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Spolverini, A.; Jones, A.V.; Hochhaus, A.; Pieri, L.; Cross, N.C.P.; Vannucchi, A.M. The myeloproliferative neoplasm-associated JAK2 46/1 haplotype is not overrepresented in chronic myelogenous leukemia. Ann. Hematol. 2011, 90, 365–366. [Google Scholar] [CrossRef] [PubMed]
- Nahajevszky, S.; Andrikovics, H.; Batai, A.; Adam, E.; Bors, A.; Csomor, J.; Gopcsa, L.; Koszarska, M.; Kozma, A.; Lovas, N.; et al. The prognostic impact of germline 46/1 haplotype of Janus kinase 2 in cytogenetically normal acute myeloid leukemia. Haematologica 2011, 96, 1613–1618. [Google Scholar] [CrossRef] [PubMed]
- Olcaydu, D.; Rumi, E.; Harutyunyan, A.; Passamonti, F.; Pietra, D.; Pascutto, C.; Berg, T.; Jäger, R.; Hammond, E.; Cazzola, M.; et al. The role of the JAK2 GGCC haplotype and the TET2 gene in familial myeloproliferative neoplasms. Haematologica 2011, 96, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Harutyunyan, A.S.; Rumi, E.; Pietra, D.; Berg, T.; Olcaydu, D.; Houlston, R.S.; Cazzola, M.; Kralovics, R. Common germline variation at the TERT locus contributes to familial clustering of myeloproliferative neoplasms. Am. J. Hematol. 2014, 89, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Rumi, E.; Passamonti, F.; della Porta, M.G.; Elena, C.; Arcaini, L.; Vanelli, L.; del Curto, C.; Pietra, D.; Boveri, E.; Pascutto, C.; et al. Familial chronic myeloproliferative disorders: Clinical phenotype and evidence of disease anticipation. J. Clin. Oncol. 2007, 25, 5630–5635. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, A.S.; Kralovics, R. Role of germline genetic factors in MPN pathogenesis. Hematol. Oncol. Clin. N. Am. 2012, 26, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.V.; Cross, N.C.P. Inherited predisposition to myeloproliferative neoplasms. Ther. Adv. Hematol. 2013, 4, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Rumi, E.; Cazzola, M. Advances in understanding the pathogenesis of familial myeloproliferative neoplasms. Br. J. Haematol. 2017, 178, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Oddsson, A.; Kristinsson, S.Y.; Helgason, H.; Gudbjartsson, D.F.; Masson, G.; Sigurdsson, A.; Jonasdottir, A.; Jonasdottir, A.; Steingrimsdottir, H.; Vidarsson, B.; et al. The germline sequence variant rs2736100_C in TERT associates with myeloproliferative neoplasms. Leukemia 2014, 28, 1371–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojesen, S.E. Telomeres and human health. J. Intern. Med. 2013, 274, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Alter, B.P.; Rosenberg, P.S.; Giri, N.; Baerlocher, G.M.; Lansdorp, P.M.; Savage, S.A. Telomere length is associated with disease severity and declines with age in dyskeratosis congenita. Haematologica 2012, 97, 353–359. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.D.; Hung, R.J.; Gaborieau, V.; Boffetta, P.; Chabrier, A.; Byrnes, G.; Zaridze, D.; Mukeria, A.; Szeszenia-Dabrowska, N.; Lissowska, J.; et al. Lung cancer susceptibility locus at 5p15.33. Nat. Genet. 2008, 40, 1404–1406. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Broderick, P.; Webb, E.; Wu, X.; Vijayakrishnan, J.; Matakidou, A.; Qureshi, M.; Dong, Q.; Gu, X.; Chen, W.V.; et al. Common 5p15.33 and 6p21.33 variants influence lung cancer risk. Nat. Genet. 2008, 40, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Gago-Dominguez, M.; Jiang, X.; Conti, D.V.; Castelao, J.E.; Stern, M.C.; Cortessis, V.K.; Pike, M.C.; Xiang, Y.-B.; Gao, Y.-T.; Yuan, J.-M.; et al. Genetic variations on chromosomes 5p15 and 15q25 and bladder cancer risk: Findings from the Los Angeles-Shanghai bladder case-control study. Carcinogenesis 2011, 32, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Shete, S.; Hosking, F.J.; Robertson, L.B.; Dobbins, S.E.; Sanson, M.; Malmer, B.; Simon, M.; Marie, Y.; Boisselier, B.; Delattre, J.-Y.; et al. Genome-wide association study identifies five susceptibility loci for glioma. Nat. Genet. 2009, 41, 899–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xu, Y.; Mei, H.; Peng, L.; Li, X.; Tang, J. The TERT rs2736100 polymorphism increases cancer risk: A meta-analysis. Oncotarget 2017, 8, 38693–38705. [Google Scholar] [CrossRef] [PubMed]
- Krahling, T.; Balassa, K.; Kiss, K.P.; Bors, A.; Batai, A.; Halm, G.; Egyed, M.; Fekete, S.; Remenyi, P.; Masszi, T.; et al. Co-occurrence of myeloproliferative neoplasms and solid tumors is attributed to a synergism between cytoreductive therapy and the common TERT polymorphism rs2736100. Cancer Epidemiol. Biomark. Prev. 2016, 25, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Tapper, W.; Jones, A.V.; Kralovics, R.; Harutyunyan, A.S.; Zoi, K.; Leung, W.; Godfrey, A.L.; Guglielmelli, P.; Callaway, A.; Ward, D.; et al. Genetic variation at MECOM, TERT, JAK2 and HBS1L-MYB predisposes to myeloproliferative neoplasms. Nat. Commun. 2015, 6, 6691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinds, D.A.; Barnholt, K.E.; Mesa, R.A.; Kiefer, A.K.; Do, C.B.; Eriksson, N.; Mountain, J.L.; Francke, U.; Tung, J.Y.; Nguyen, H.M.; et al. Germ line variants predispose to both JAK2 V617F clonal hematopoiesis and myeloproliferative neoplasms. Blood 2016, 128, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Larrán, A.; Angona, A.; Martínez-Avilés, L.; Bellosillo, B.; Besses, C. Influence of JAK2 46/1 haplotype in the natural evolution of JAK2V617F allele burden in patients with myeloproliferative neoplasms. Leuk. Res. 2012, 36, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.; Birgens, H.S.; Nordestgaard, B.G.; Bojesen, S.E. Diagnostic value of JAK2 V617F somatic mutation for myeloproliferative cancer in 49,488 individuals from the general population. Br. J. Haematol. 2013, 160, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Gieger, C.; Radhakrishnan, A.; Cvejic, A.; Tang, W.; Porcu, E.; Pistis, G.; Serbanovic-Canic, J.; Elling, U.; Goodall, A.H.; Labrune, Y.; et al. New gene functions in megakaryopoiesis and platelet formation. Nature 2011, 480, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Newton-Cheh, C.; Johnson, T.; Gateva, V.; Tobin, M.D.; Bochud, M.; Coin, L.; Najjar, S.S.; Zhao, J.H.; Heath, S.C.; Eyheramendy, S.; et al. Genome-wide association study identifies eight loci associated with blood pressure. Nat. Genet. 2009, 41, 666–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Harst, P.; Zhang, W.; Mateo Leach, I.; Rendon, A.; Verweij, N.; Sehmi, J.; Paul, D.S.; Elling, U.; Allayee, H.; Li, X.; et al. Seventy-five genetic loci influencing the human red blood cell. Nature 2012, 492, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Trillos, A.; Maffioli, M.; Colomer, D.; Alvarez-Larrán, A.; Pereira, A.; Angona, A.; Bellosillo, B.; Cervantes, F. Relationship between the 46/1 haplotype of the JAK2 gene and the JAK2 mutational status and allele burden, the initial findings, and the survival of patients with myelofibrosis. Ann. Hematol. 2014, 93, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Colaizzo, D.; Tiscia, G.L.; Bafunno, V.; Amitrano, L.; Vergura, P.; Grandone, E.; Guardascione, M.A.; Margaglione, M. The JAK2 rs12343867 CC genotype frequently occurs in patients with splanchnic venous thrombosis without the JAK2V617F mutation: A retrospective study. J. Thromb. Haemost. 2010, 8, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Smalberg, J.H.; Koehler, E.; Darwish Murad, S.; Plessier, A.; Seijo, S.; Trebicka, J.; Primignani, M.; de Maat, M.P.M.; Garcia-Pagan, J.-C.; Valla, D.C.; et al. The JAK2 46/1 haplotype in Budd-Chiari syndrome and portal vein thrombosis. Blood 2011, 117, 3968–3973. [Google Scholar] [CrossRef] [PubMed]
- Kouroupi, E.; Kiladjian, J.-J.; Chomienne, C.; Dosquet, C.; Bellucci, S.; Valla, D.; Cassinat, B. The JAK2 46/1 haplotype in splanchnic vein thrombosis. Blood 2011, 117, 5777–5778. [Google Scholar] [CrossRef] [PubMed]
- Villani, L.; Bergamaschi, G.; Primignani, M.; Rosti, V.; Carolei, A.; Poletto, V.; Catarsi, P.; Spolverini, A.; Vannucchi, A.M.; Barosi, G. JAK2 46/1 haplotype predisposes to splanchnic vein thrombosis-associated BCR-ABL negative classic myeloproliferative neoplasms. Leuk. Res. 2012, 36, e7–e9. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, R.H.; Lea, N.C.; Mohamedali, A.M.; Smith, A.E.; Orr, D.W.; Roberts, L.N.; Heaton, N.D.; Wendon, J.A.; O’Grady, J.G.; Heneghan, M.A.; et al. Prevalence and clinical outcomes of the 46/1 haplotype, Janus kinase 2 mutations, and ten-eleven translocation 2 mutations in Budd-Chiari syndrome and their impact on thrombotic complications post liver transplantation. Liver Transplant. 2012, 18, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-L.; Zhang, P.-J.; Sun, G.-X.; Lu, Z.-J. The JAK2 46/1 haplotype (GGCC) in myeloproliferative neoplasms and splanchnic vein thrombosis: A pooled analysis of 26 observational studies. Ann. Hematol. 2014, 93, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Han, D.Y.; Fraser, A.G.; Huebner, C.; Lam, W.J.; Morgan, A.R.; Duan, H.; Karunasinghe, N. Genetic factors in chronic inflammation: Single nucleotide polymorphisms in the STAT-JAK pathway, susceptibility to DNA damage and Crohn’s disease in a New Zealand population. Mutat. Res. 2010, 690, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-X.; Song, J.; Wang, J.; Dong, W.-G. JAK2 rs10758669 polymorphisms and susceptibility to ulcerative colitis and Crohn’s disease: A meta-analysis. Inflammation 2014, 37, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Can, G.; Tezel, A.; Gürkan, H.; Tozkır, H.; Ünsal, G.; Soylu, A.R.; Ümit, H.C. Investigation of IL23R, JAK2, and STAT3 gene polymorphisms and gene-gene interactions in Crohn’s disease and ulcerative colitis in a Turkish population. Turk. J. Gastroenterol. 2016, 27, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Hansoul, S.; Nicolae, D.L.; Cho, J.H.; Duerr, R.H.; Rioux, J.D.; Brant, S.R.; Silverberg, M.S.; Taylor, K.D.; Barmada, M.M.; et al. Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat. Genet. 2008, 40, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandzar, S.; Gupta, S.; Platt, M.O. Crohn’s disease: A review of treatment options and current research. Cell. Immunol. 2013, 286, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Hasselbalch, H.C. Perspectives on chronic inflammation in essential thrombocythemia, polycythemia vera, and myelofibrosis: Is chronic inflammation a trigger and driver of clonal evolution and development of accelerated atherosclerosis and second cancer? Blood 2012, 119, 3219–3225. [Google Scholar] [CrossRef] [PubMed]
- Balassa, K.; Krahling, T.; Remenyi, P.; Batai, A.; Bors, A.; Kiss, K.P.; Torbagyi, E.; Gopcsa, L.; Lengyel, L.; Barta, A.; et al. Recipient and donor JAK2 46/1 haplotypes are associated with acute graft-versus-host disease following allogeneic hematopoietic stem cell transplantation. Leuk. Lymphoma 2017, 58, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.J. Somatic and germline genetics at the JAK2 locus. Nat. Genet. 2009, 41, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Spasovski, V.; Tosic, N.; Nikcevic, G.; Stojiljkovic, M.; Zukic, B.; Radmilovic, M.; Karan-Djurasevic, T.; Srzentic, S.; Colovic, M.; Pavlovic, S. The influence of novel transcriptional regulatory element in intron 14 on the expression of Janus kinase 2 gene in myeloproliferative neoplasms. J. Appl. Genet. 2013, 54, 21–26. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the JAK2 haplotypeGGCC_46/1 genomic architecture and its possible interplay with the germline TERT rs2736100 in inducing somatic gene mutations, clonal expansion and MPN onset. Red, blu and green arrows refer to the possible induction of somatic gene mutations by the JAK2 haplotypeGGCC_46/1, TERT rs2736100, and other germline polymorphisms, respectively.
Figure 1.
Schematic representation of the JAK2 haplotypeGGCC_46/1 genomic architecture and its possible interplay with the germline TERT rs2736100 in inducing somatic gene mutations, clonal expansion and MPN onset. Red, blu and green arrows refer to the possible induction of somatic gene mutations by the JAK2 haplotypeGGCC_46/1, TERT rs2736100, and other germline polymorphisms, respectively.

{kind=link}
{kind=link}
Table 1.
Main literature studies describing the frequency of the JAK2 haplotypeGGCC_46/1 in myeloid malignancies and possible clinical correlations.
Table 1.
Main literature studies describing the frequency of the JAK2 haplotypeGGCC_46/1 in myeloid malignancies and possible clinical correlations.
| Reference | Main Findings | Association between the GGCC Haplotype and Somatic Mutations | Association between the GGCC Haplotype and JAK2 V617F Neg MPN | Tagging SNPs | Series of Analyzed Patients | Clinical Findings |
|---|---|---|---|---|---|---|
| [6] | First identification of JAK2 SNPs significantly associated with PV or ET | - | - | rs7046736, rs10815148, rs12342421 | 84 PV, 58 PMF, and 37 ET | Association between JAK2 SNPs and leucocytosis, higher hemoglobin level, lower platelet count |
| [7] | Association between JAK2 46/1 haplotype and JAK2 V617F positive MPN | JAK2 V617F (48–56%) | Weak association | rs12340895 | 88 PMF | Hematologically normal individuals that carried at least one 46/1 allele grew significantly fewer CFU-GM |
| [19] | The incidence of the 46/1-linked C allele was significantly higher in ET than in population controls | JAK2 V617F (44%) | Significant association | rs12343867 | 226 ET | The clinical features of 46/1 positive and negative ET were indistinguishable, including blood counts, rate of thrombosis/disease transformation and survival |
| [12] | JAK2 germline genetic variation affects disease susceptibility in PMF regardless of VF mutational status | JAK2 V617F (50%) | Significant association | rs12343867 | 130 MF | Association between nullizygosity for the JAK2 46/1 haplotype SNP allele and shortened survival |
| [55] | The SNP rs10758669_C allele increase the risk of having Crohn’s disease | - | - | rs10758669 | 302 Crohn’s disease | This JAK2 variant strongly enhanced the risk of ileocolonic disease, with stricturing or ileal/stricturing behavior, requiring a bowel resection |
| [16] | The frequency of 46/1 was higher in MPL mutated cases compared with controls | MPL W515K/L mutations (36%) | The 46/1 haplotype was also overrepresented in cases without V617F mutation | rs12340895 | 176 MPL pos/V617F neg ET, and 212 V617F pos ET | No association between 46/1 and clinical or laboratory features |
| [20] | The frequency of the 46/1 haplotype, was significantly higher in PMF patients showing the highest V617F allele burden | JAK2 V617F (38.6%) | No statistical significant association | rs12343867 | 202 PMF | No statistically significant correlations between any of the possible rs12343867 genotypes and hematological or clinical variables |
| [13] | The 46/1 haplotype is a predisposition factor for JAK2 V617F positive MPN, and is also significantly associated with AML patients with normal karyotype | JAK2 V617F (85%) | Significant association | rs12343867 | 312 MPN, 339 AML | The 46/1 haplotype is not associated with MPN manifestations, like disease type, splenomegaly, signs of increased erythropoiesis or myelopoiesis and vascular complication, except the increased risk of the development of myelofibrosis in homozygous cases |
| [50] | The 46/1 haplotype was overrepresented in JAK2V617F positive SVT patients compared with controls | JAK2 V617F (43%) | JAK2V617F negative SVT patients with a proven MPN also exhibited an increased frequency of the 46/1 haplotype | rs12343867 | 199 SVT | The 46/1 haplotype was associated with increased erythropoiesis (higher hemoglobin levels, hematocrit, and red blood cell count) in JAK2 V617F negative SVT patients |
| [25] | Association of the JAK2 46/1 haplotype with disease characteristics and treatment outcome in AML patients | - | - | rs12343867 | 176 AML | The 46/1 haplotype was found to be a factor predisposing to the development of acute myelomonocytic leukemia. In NK-AML, the carriers of 46/1 haplotype are characterized by shorter disease-free survival and overall survival |
| [43] | Untreated PV patients with homozygous JAK2 46/1 haplotype experienced a progressive increase in the JAK2 V617F allele burden | JAK2 V617F (68%) | - | rs12340895, rs12343867 | 26 PV, 36 ET | The 46/1 JAK2 haplotype status was not statistically different according to age, gender, type of diagnosis (PV or ET) or baseline hematological values |
| [48] | Among JAK2 V617F positive patients, those who were homozygous for the 46/1 haplotype had a higher allele burden | JAK2 V617F (40%) | - | rs12340895 | 132 MF | Patients with homozygous 46/1 haplotype showed significantly higher hemoglobin values and leukocyte counts, but no association was seen with other clinic hematologic features |
| [21] | The frequency of 46/1 haplotype was significantly higher in JAK2 V617F positive PV/ET but not in ET patients with CALR mutations | JAK2 V617F (40–50%) | No statistical significant association | rs12340895 | 72 PV, 115 ET | The presence of 46/1 haplotype had a trend to have higher white blood cell count in JAK2 V617F mutated PV and ET patients but not in CALR mutated ET |
| [61] | Both, recipient and donor 46/1 haplotypes significantly affected aGvHD grades II–IV development | - | - | rs12343867 | 124 AML | The recipient haplotype remained independently related to aGvHD, while the donor not. Significantly less relapses were observed among haplotype carriers, but overall survival did not differ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Anelli, L.; Zagaria, A.; Specchia, G.; Albano, F. The JAK2 GGCC (46/1) Haplotype in Myeloproliferative Neoplasms: Causal or Random? Int. J. Mol. Sci. 2018, 19, 1152. https://doi.org/10.3390/ijms19041152
AMA Style
Anelli L, Zagaria A, Specchia G, Albano F. The JAK2 GGCC (46/1) Haplotype in Myeloproliferative Neoplasms: Causal or Random? International Journal of Molecular Sciences. 2018; 19(4):1152. https://doi.org/10.3390/ijms19041152
Chicago/Turabian StyleAnelli, Luisa, Antonella Zagaria, Giorgina Specchia, and Francesco Albano. 2018. "The JAK2 GGCC (46/1) Haplotype in Myeloproliferative Neoplasms: Causal or Random?" International Journal of Molecular Sciences 19, no. 4: 1152. https://doi.org/10.3390/ijms19041152
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.