Potential Roles of Dec and Bmal1 Genes in Interconnecting Circadian Clock and Energy Metabolism


1
Department of Pathology, Wakayama Medical University School of Medicine, 811-1 Kimiidera, Wakayama 641-8509, Japan
2
Department of Physiology, Wakayama Medical University School of Medicine, 811-1 Kimiidera, Wakayama 641-8509, Japan
3
Department of Biochemistry and Molecular Biology, Nihon University School of Dentistry at Matsudo, 2-870-1 Sakae-cho Nishi, Matsudo, Chiba 271-8587, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(3), 781; https://doi.org/10.3390/ijms19030781
Submission received: 11 February 2018
/
Revised: 4 March 2018
/
Accepted: 7 March 2018
/
Published: 8 March 2018
(This article belongs to the Special Issue Molecular and Cellular Mechanisms of Circadian Rhythms)
Abstract
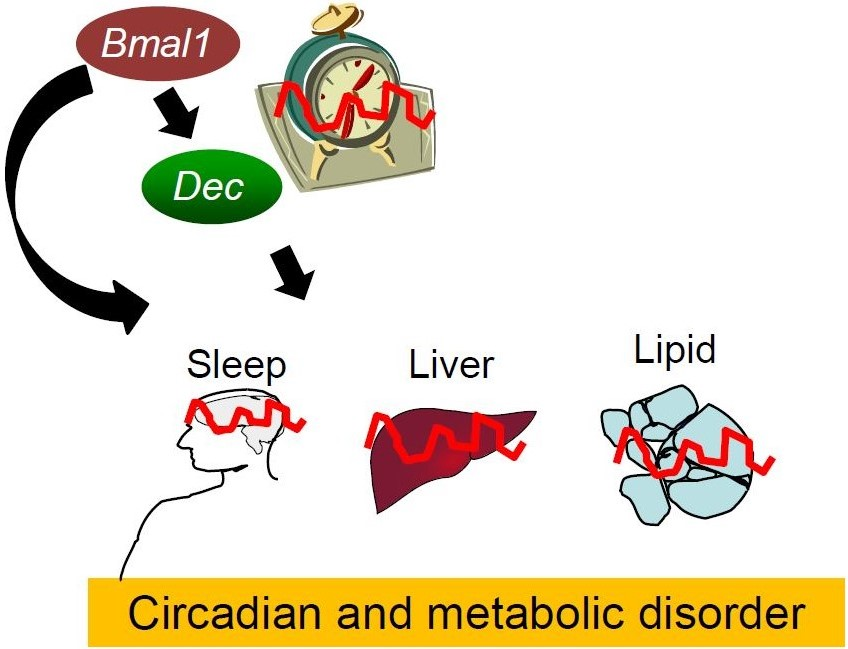
:The daily rhythm of mammalian energy metabolism is subject to the circadian clock system, which is made up of the molecular clock machinery residing in nearly all cells throughout the body. The clock genes have been revealed not only to form the molecular clock but also to function as a mediator that regulates both circadian and metabolic functions. While the circadian signals generated by clock genes produce metabolic rhythms, clock gene function is tightly coupled to fundamental metabolic processes such as glucose and lipid metabolism. Therefore, defects in the clock genes not only result in the dysregulation of physiological rhythms but also induce metabolic disorders including diabetes and obesity. Among the clock genes, Dec1 (Bhlhe40/Stra13/Sharp2), Dec2 (Bhlhe41/Sharp1), and Bmal1 (Mop3/Arntl) have been shown to be particularly relevant to the regulation of energy metabolism at the cellular, tissue, and organismal levels. This paper reviews our current knowledge of the roles of Dec1, Dec2, and Bmal1 in coordinating the circadian and metabolic pathways.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The supply and demand of cellular bioenergy vary across the 24 h light-dark cycle in mammals. This phenomenon is subject to the circadian clock system [1,2,3,4,5,6], which comprises the central clock in the suprachiasmatic nucleus (SCN) of the hypothalamus and peripheral clocks that reside in extra-SCN areas of the brain and in peripheral tissues. Both the central and peripheral clocks are made up of similar clock machinery, the so-called molecular clock, which consists of a series of clock genes [7,8,9,10]. Since their discovery as components of a circadian oscillator, the clock genes have been shown to regulate cellular energy metabolism and body energy homeostasis through the transcriptional control of metabolic genes, whose function has a great influence on cellular redox and bioenergy [1,3,4,5,6]. Of interest, it has also been shown that the function of clock genes is in turn altered by changes in energy balance [2,4,11,12], suggesting the mutual regulation of the circadian and metabolic systems. Here, we offer a description of the interplay between the circadian and metabolic systems, focusing especially on the key clock genes Dec1, Dec2, and Bmal1, which have been shown to be linked to energy metabolism, and insights into their emerging roles in the understanding of circadian and metabolic disorders.
2. The Circadian Clock Machinery
Circadian clock genes are defined as genes that are required to regulate circadian rhythms of physiology at the cellular, tissue, and organismal levels. Clock genes form the molecular clock which is composed of interconnected feedback loops of gene transcription and translation (Figure 1). In mammals, two core clock genes, Clock and Bmal1, encode proteins that are members of the basic helix-loop-helix (bHLH)-PAS transcription factor family [13,14,15,16]. Through protein-protein interactions involving their PAS domains, CLOCK and BMAL1 form a heterodimeric complex in the cytoplasm, which then translocates to the nucleus and binds to E-box enhancer sequences to activate the transcription of repressor clock genes [13,17]. These include the Period (Per1 and Per2) and Cryptochrome genes (Cry1 and Cry2), whose protein products lower their own transcription by repressing CLOCK:BMAL, thus composing an autoregulatory feedback loop. Since the discovery of this “core” feedback loop, additional feedback loops have also been described. By binding to E-box enhancers, CLOCK:BMAL1 activates the transcription of another core clock gene, Rev-erbα, which codes for an orphan nuclear receptor [18,19]. The REV-ERBα protein in turn inhibits Bmal1 transcription by direct binding to the ROREs (retinoic acid-related orphan receptor response elements) residing in the Bmal1 gene promoter, thereby forming another feedback loop [18,19].
The Dec1 and Dec2 genes, which encode bHLH transcription factors, have also been proposed as clock genes that comprise an additional autoregulatory feedback loop [20,21,22,23]. Dec1 and Dec2, similar to the Rev-erbα gene, are transactivated by CLOCK:BMAL1 through E-box elements in their promoters [21,22,23]. However, unlike the REV-ERBα protein, which inhibits its own expression by repressing the transcription of one of its activators, Bmal1, DECs repress their own transcription by directly binding to the BMAL1 protein and/or by competing with CLOCK:BMAL1 for the occupancy of E-box sequences in their promoters [20,21,22,23,24,25]. As repressors of CLOCK:BMAL1-induced transactivation, DECs also inhibit the transcription of other clock and clock-controlled genes, such as Per1 and Dbp (albumin d-element binding protein) [20,22,26]. Collectively, the molecular clock is composed of a complex network of autoregulatory feedback loops of gene transcription and translation, thus generating circadian rhythms at the molecular level.
3. The Diurnal Rhythm of Energy Metabolism and Its Significance
The function of a wide range of metabolic processes, such as the synthesis and breakdown of carbohydrates, lipids, and proteins varies across the sleep/wake cycle in mammals. For instance, glycogen synthesis in the muscle and liver occurs during the waking period, whereas glycogenolysis in the liver peaks during sleep [27,28]. An increase in lipoprotein lipase activity, which promotes lipid absorption from the blood-stream into tissues, also occurs during the fed (i.e., waking) state [29]. One benefit of these metabolic rhythms is that it enables metabolic tissues to anticipate supply and demand of energy in the body and optimize both energy storage and utilization. Therefore, it is conceivable that uncoupling the energy supply from its demand in each metabolic organ results in a malfunction of body energy metabolism in mammals. Indeed, both clinical and experimental studies have long suggested that disruption of behavioural rhythms such as sleep/wake and fasting/feeding cycles leads to defects in glucose and lipid metabolism. Best known are the epidemiological findings that sleep disturbances and shift work are highly associated with the development of obesity, dyslipidaemia, and type 2 diabetes in humans [30,31,32,33,34,35,36,37,38]. Similarly, in rodents, it has also been reported that a shift in meal time not only disturbs the behavioural rhythm but also induces insulin resistance [39,40]. Dec1 knockout mice showed behavioral rhythm with longer circadian period and affected the circadian phase of Per1 [26]. In addition, double mutant mice of Per1 and Dec revealed synergistic effects of circadian disturbance [41]. We demonstrated that DECs have the strongest suppressive effects on CLOCK:BMAL1-induced transactivation among negative regulators DECs, PERs and CRYs [20,21,22]. Recent our reports using Dec knockout mice showed regarding the role of Dec in metabolism. First, Dec1 knockout mice decreased lipid levels and oxidative stresses, increasing fibroblast growth factor 21 (FGF21) levels [42]. Second, we revealed dominant energy sensor phosphorylation AMP-activated protein kinase (pAMPK), which mention at later Section 6, is markedly increased in Dec2 knockout mice livers [43]. These evidences suggest that Dec play important roles in the regulation of circadian rhythm and metabolism.
It has been proven that Clock mutant mice disturbed diurnal feeding rhythm and developed a metabolic syndrome [44]. It has also been reported that DEC2 amino-acid mutation P385R is associated with short sleep [45]. On the other hand, Dec2 knockout mice did not reveal the sleep phenotype [46]. It seems that DEC2 P385R works as a dominant negative fashion. We also revealed that human DEC1 amino-acid mutation R65A worked as a dominant negative fashion against CLOCK:BMAL1 transactivation [21]. These dominant negative functions of DEC mutation may affect differences in sleep phenotype of Dec knockout mice. Collectively, these human and animal studies strongly suggest a significant role of metabolic and circadian rhythms in maintaining health and preventing metabolic and circadian disorders.
4. Clock Genes Regulate Both Metabolic Rhythms and Processes
As the molecular mechanism of the mammalian circadian clock was revealed, it became clear that metabolic rhythms are under the control of the molecular clock [1,2,3,4,5,6]. For example, in studies of with tissue-specific knockout of the clock gene, the diurnal rhythm in hepatic glucose metabolism was found to be regulated by the liver clock [44,47]. Recent studies have also shown that the production of hepatic metabolites such as nucleotides and amino acids exhibits diurnal variation in mice and that BMAL1 plays an important role in the circadian oscillation of these metabolites [48]. In addition to the liver clock, the molecular clock in the pancreas was found to regulate the daily rhythm of insulin secretion [49]. While these local clocks play an important role in generating local metabolic rhythms, systemic signals produced by the fasting/feeding cycle, which is driven by the SCN clock, also affect certain cellular metabolic rhythms independently of the local clock [50].
Apart from its role in generating metabolic rhythms, the molecular clock also participates in fundamental metabolic processes such as the proliferation of islet cells in the pancreas, glucose transport by skeletal muscle, and adipogenesis in white adipose tissue. Both the BMAL1 and CLOCK proteins in the pancreas have been shown to regulate secretion of insulin and proliferation of pancreatic islets [51,52]. Insulin-dependent glucose uptake in skeletal muscle was also found to be associated with the function of the Bmal1 gene [53]. Furthermore, BMAL1 regulates adipocyte differentiation and lipogenesis in white adipose tissue and plays an important role in ectopic fat deposition in the liver and skeletal muscle [54,55]. Intriguingly, both genetic and epigenetic in silico studies support these experimental findings that the Bmal1 gene is linked to energy metabolism. Human genome-wide association studies have revealed that certain haplotypes of the Bmal1 gene are associated with the development of type 2 diabetes [56]. Moreover, using whole-genome chromatin immunoprecipitation-based analyses, a large number of BMAL1 target genes were found to be classified as genes that are related to cellular energy metabolism [57,58,59].
Given that BMAL1, together with CLOCK, directly activates the transcription of both Dec1 and Dec2 [20,21,22,23], it is possible that BMAL1 may control energy metabolism through the function of DEC1 and DEC2. Although this possibility remains to be examined, the DECs per se regulate lipid metabolism in metabolic tissues. DEC1 and DEC2 inhibit lipogenesis in the liver by repressing the transcription of Srebp-1c (sterol regulatory element-binding protein-1c), which codes for the master regulator of lipogenic genes, by binding to the E-box in its promoter region [60,61]. Therefore, changes in the DEC levels could affect the degree of lipid storage in the liver. Indeed, both leptin-deficient ob/ob and leptin receptor-deficient db/db mice show a decrease in Dec1 expression in the liver and a significant increase in hepatic triglycerides [61], whereas only a mild increase in hepatic triglyceride content was observed in mice with alcohol-induced fatty liver, in which Dec1 expression was increased [62]. White adipose tissue also requires the function of the Dec1 gene to regulate adipogenesis. DEC1 suppresses adipocyte differentiation by inhibiting the expression of peroxisome proliferator-activated receptor γ (Pparγ), a gene encoding a nuclear receptor critical for adipogenesis [63]. Collectively, these findings suggest that, in addition to its role as a circadian oscillator, the molecular clock plays an important role in regulating fundamental properties of energy metabolism in major metabolic organs.
5. The Clock Senses Energy Balance
While the molecular clock regulates energy metabolism, changes in energy balance, in turn, affect the function of the molecular clock [64]. For example, mice fed a high-fat diet exhibit an increase in the circadian period of locomotor activity under constant dark conditions, which indicates that high-fat feeding alters the function of the SCN clock [65]. Energy excess also attenuates the amplitude of rhythmic expression of clock genes in peripheral tissues [12,65]. In addition to excess energy intake, the timing of energy intake has been shown to impact the rhythmic expression of clock genes in metabolic tissues [66]. Specifically, in mice, when food availability is restricted only to the light (i.e., sleep) phase, the rhythmic expression of clock genes occurs in anti-phase to that observed in ad lib feeding. Although the molecular mechanism is not fully understood, this phenomenon is predominantly observed in peripheral metabolic tissues, such as the liver and white adipose tissues, and not in the SCN [39,67]. It should be noted that changes in the fasting/feeding cycle not only affect patterns of rhythmic expression of clock genes but also alter their expression levels. In the murine liver, the mRNA expression level of Dec1 is significantly decreased by fasting and increased by re-feeding [68,69,70], although Dec2 expression is not altered by feeding [68,69]. The expression level of Per2 in the liver, but not in the heart and lung, is also increased by re-feeding after fasting [68,70]. Although the effects of changes in the fasting/feeding cycle on clock gene expression are limited to certain clock genes and to specific tissues, these studies nevertheless suggest that the molecular clock is sensitive to alteration in the balance and the timing of energy intake.
6. Molecules at the Intersection of the Molecular Clock and Energy Metabolism
How does the molecular clock sense changes in energy balance? The fasting/feeding and activate/resting cycles induce a dynamic change in the levels of metabolites and fuels in the cells and interstitial spaces surrounding the cells. The molecules with altered levels include cellular redox components and energy resources, energy substrates and oxygen. Growing evidence now suggests the possibility that any of these molecules can be sensed by the molecular clock through molecules that not only function as key metabolic regulators but also modulate clock function.
6.1. Cellular Redox
Molecules associated with the cellular redox state were perhaps the first metabolic factors to be proposed to influence the function of the molecular clock (Figure 2). Specifically, cellular levels of the hydrogen carriers NAD+ (nicotinamide adenine dinucleotide) and FAD (flavin adenine dinucleotide) have been shown to either directly or indirectly affect the regulation of clock gene function. For example, the reduced forms of NAD+ and NADP+ (i.e., NADH and NADPH) increase the DNA-binding activity of the CLOCK:BMAL1 heterodimer in vitro [71]. Accordingly, inhibition of the pentose phosphate pathway, a major source of NADPH, prolongs the circadian period of clock gene expression in cultured cells [72]. Recently, it has also been reported that the circadian period of PER2 expression is lengthened by FAD through stabilization of CRY proteins [73]. Given that the levels of NAD+ and FAD oscillate in both tissues and cultured cells [73,74,75,76], the diurnal changes in these hydrogen carriers may play an important role in shaping the rhythmic expression of clock genes. The oscillation of hydrogen carriers seems to be under enzymatic control because both the mRNA and protein levels of genes encoding nicotinamide phosphoribosyltransferase (NAMPT) and riboflavin kinase (RFK), which are the rate-limiting enzymes of the NAD+ salvage pathway and FAD biosynthesis, respectively, vary dynamically in a circadian fashion in the murine liver [73,74,75]. Although the molecular mechanisms underlying the rhythmic expression of Rfk remain unclear, the transcription of Nampt has been reported to be directly regulated by the CLOCK:BMAL1 heterodimer through the E-box in its promoter [74,75]. Collectively, these findings reveal that cellular redox affects clock function which in turn participates in the circadian regulation of redox state, suggesting a mutual relationship between controls of cellular redox and clock function.
NAD+-consuming enzymes have also been shown to link energy metabolism with the molecular clock (Figure 2). Sirtuin 1 (SIRT1), whose activity relies on the NAD+ level, deacetylates histones and a wide variety of proteins, including those that regulate glucose and lipid metabolism [77,78,79,80,81]. SIRT1 forms a complex with CLOCK:BMAL1 and promotes the deacetylation of BMAL1, which then alters the transcriptional activity of CLOCK:BMAL1 [82,83]. As mentioned earlier, since DEC1 and DEC2 are transactivated by CLOCK:BMAL1, it is possible that SIRT1 indirectly regulates DEC1 and DEC2 levels through BMAL1 deacetylation. SIRT1 also deacetylates and degrades PER2 [83]. In addition, SIRT1 deacetylates and activates PGC-1α (PPARγ coactivator 1-alpha) [81,84], a transcriptional coactivator that promotes Bmal1 expression through coactivation of ROR family members, including RORα and RORγ [85]. Consistent with these findings, deletion of the Sirt1 gene leads to a significant reduction in the expression level of Bmal1 [83,86]. Interestingly, mice with brain-specific knockout of Sirt1 showed a lengthened circadian period of locomotor activity [86], which further indicates that SIRT1 is linked to clock function. In addition to SIRT1, poly (ADP-ribose) polymerase 1 (PARP-1), which exerts its physiological function by transferring ADP-ribose subunits from NAD+ to its target proteins, has also been reported to modulate the function of the molecular clock. PARP-1 binds and poly (ADP-ribosyl) ates the CLOCK protein, thereby reducing the DNA binding activity of CLOCK: BMAL1 [87]. Importantly, this phenomenon is observed to occur in a circadian fashion. It is possible that PARP-1 indirectly decreases DEC1 and DEC2 levels through the CLOCK:BMAL1 reduction.
Nocturnin which encodes circadian deadenylase is regulated by CLOCK:BMAL1, and crosstalk circadian and metabolism [88]. Nocturnin plays important roles in lipid metabolism, adipogenesis, and glucose homeostasis, involving circadian clock [88].
Taken together, these findings suggest that the clock can sense cellular redox through changes in the level of hydrogen carriers and/or the activity of NAD+-consuming enzymes.
6.2. Cellular Energy Status
In addition to cellular redox, changes in the intracellular level of energy resources (i.e., ATP) are sensed by the molecular clock. AMPK has been proposed as the key molecule that mediates the ATP-sensing property of the molecular clock [89,90]. AMPK is a heterotrimeric protein composed of a catalytic α and regulatory β and γ subunits, and each of these subunits is encoded by either 2 (α1 and α2 or β1 and β2) or 3 genes (γ1, γ2, and γ3) [91,92,93]. AMPK is activated when a threonine residue (Thr172) within the α subunit is phosphorylated by upstream kinases. These include liver kinase B1 (LKB1) and calmodulin-dependent protein kinase kinase (CaMKK). LKB1 is thought to be constitutively active and promotes phosphorylation of Thr172 when AMP binds to the AMPK-γ subunit, while CaMKK activates (i.e., phosphorylates) AMPK in response to a rise in intracellular Ca2+, independent of changes in the AMP/ATP ratio. Due to the energy-dependent phosphorylation of AMPK by LKB1, AMPK has been considered a cellular energy sensor. Importantly, it has been reported that activated AMPK directly or indirectly interacts with the molecular clock (Figure 3) [89,90]. For instance, AMPK directly phosphorylates CRY1 and indirectly phosphorylates PER2 by activating (i.e., phosphorylating) CKIε (casein kinase Iε) [94,95], which leads to the degradation of both CRY1 and PER2. Therefore, it could be speculated that a defect in the function of AMPK causes dysregulation of the circadian clock. Indeed, mice deficient in the Ampkα1 gene show a shorter circadian period of locomotor activity [96]. Moreover, the rhythmic expression of clock genes in peripheral tissues is altered in mice lacking either Ampkα1 or Ampkα2 [96].
While the regulation of the molecular clock is modulated by AMPK, the molecular clock in turn regulates AMPK activity [43,97]. We have recently reported that DEC1 directly binds to the E-box box of the Lkb1 promoter and suppresses the expression of the LKB1 protein, thereby reducing the phosphorylation of AMPK [97]. We also reported that the phosphorylation of AMPK is increased in the liver and lung in Dec2 knockout mice, indicating that DEC2 may also suppress AMPK activity [43], although the precise mechanism remains to be elucidated. Of note, phosphorylation of AMPK is rhythmic and is inversely correlated with the rhythmic expression of DECs [20,96,97], further indicating that AMPK is a downstream target of DECs. Taken together, these findings suggest a mutual relationship between the function of the molecular clock and AMPK activity. It will be important to identify other molecules that link clock function and changes in the cellular energy balance.
6.3. Glucose Metabolism
In vitro studies have shown that the change in the glucose level also affects the function of the molecular clock. By using rat-1 fibroblasts, Hirota et al. demonstrated that glucose down-regulates the transcription of the Per1 and Per2 genes [98]. We and others have also reported that in cultured cells, Dec1 expression is increased by glucose supplementation, while glucose depletion decreases both DEC1 and DEC2 protein levels [43,99]. These findings are consistent with in vivo studies that fasting decreases and re-feeding increases Dec1 expression in the murine liver [68,69,70]. The concept that glucose is a key nutrient cue that alters clock gene function has been further supported by a series of animal studies. Among food macronutrients, glucose has been shown to alter the phase of the rhythm of locomotor activity in mice [100,101]. In addition, Oike et al. showed that intraperitoneal administration of glucose with amino acids increases the mRNA expression levels of Per1, Per2, Dec1, and Dec2 in the liver in fasted mice [70]. While the precise mechanisms underlying the glucose-induced alteration in clock function remain unknown, transcriptional regulators have been proposed to mediate the effect of glucose on clock genes [98]. Liver X receptor (LXR), a glucose-activated transcription factor [102], may be an attractive candidate transmitter of the glucose signal to the molecular clock because LXR was found to induce Dec1 expression by binding its promoter [69].
In addition to glucose, insulin was also found to affect the function of the molecular clock. Recent studies have shown that insulin promotes the phosphorylation of the BMAL1 protein by Akt, thereby suppressing its transcriptional activity [103]. Furthermore, several studies demonstrated that insulin increases the expression levels of Per1, Per2, Dec1, and Dec2 in cultured cells [104,105,106,107]. While the molecular mechanisms underlying insulin-induced expression of these clock genes remain to be elucidated, insulin signalling pathways may play an important role in the regulation of the molecular clock because insulin-induced expression of Dec1 and Dec2 was found to be inhibited by the blockade of phosphoinositide 3-kinase, protein kinase C, or mammalian target of rapamycin [104,105,106].
We showed that high-fat diet induced the insulin and glucose levels [65]. As described above, insulin and glucose induced Dec1 and Dec2 expression, respectively. We speculate that high-fat diet induces Dec1 and Dec2 expression through insulin and glucose supplement.
6.4. Lipid Metabolism
The observation that the regulation of PPARs is under the control of clock proteins has led to the idea that the function of the molecular clock is linked to lipid metabolism [107]. Both DEC1 and DEC2 were found to regulate adipogenesis by repressing the transcription of a gene encoding PPARγ, the master transcriptional regulator of adipogenesis [63,108,109]. Accordingly, Dec1 overexpression has been shown to suppress adipocyte differentiation [110]. DEC1 does not directly bind to the Pparγ promoter, but exerts its repressive effect on Pparγ expression by interacting with DNA-bound CCAAT/enhancer binding protein [63,108]. In addition to DEC proteins, other clock proteins have also been shown to regulate adipocyte differentiation [111]. PER2 inhibits the recruitment of PPARγ to its target gene promoters, and a defect in the Per2 gene increases adipogenesis in cultured cells [112]. In contrast, knockdown of the Bmal1 gene was found to suppress adipocyte differentiation in 3T3-L1 cells [55], although in vivo studies have shown contradictory results, specifically, that mice with adipocyte-specific knockout of the Bmal1 gene develop obesity [113]. It should be noted that the function of PPARγ could in turn affect the molecular clock. The transcription of a gene coding for REV-ERBα, a circadian transcriptional repressor, was found to be induced by PPARγ through the direct repeat-2 response element in the Rev-erbα gene [114].
In addition to PPARγ, PPARα, the major regulator of fatty acid oxidation in the liver, has also been reported to interact with the molecular clock. The interconnection between PPARα and the circadian clock was first indicated in studies showing that the expression of PPARα is rhythmic at both the mRNA and protein levels [115]. Subsequently, PPARα was found to be a direct target of clock genes [116,117]. The CLOCK:BMAL1 heterodimer activates the transcription of the Pparα gene by binding to the E-box within the gene promoter [116]. PER2 also interacts with the PPARα protein through its LXXLL motifs [117]. Similar to the case of PPARγ, PPARα in turn regulates the transcription of clock genes. PPARα transactivates the Bmal1 and Rev-erbα genes via PPAR response elements located in the promoters of these genes [118,119].
6.5. Hypoxia
Changes in tissue oxygen levels have also been shown to alter clock gene function. Both Dec1 and Dec2 expression were found to be induced by hypoxic conditions in cultured cells [120,121,122,123]. Promoter analyses have revealed that hypoxia-inducible factor-1α (HIF-1α), a master regulator of oxygen homeostasis, activates the transcription of the Dec1 and Dec2 genes by binding to the hypoxia-response elements (HREs) present within the promoters of these genes [121]. More recently, an additional interrelationship between hypoxia and clock gene function has emerged from studies by Peek et al. [124], who demonstrated that HIF-1α binds to the E-box within the promoters of Per2 and Cry1. While HIF-1α regulates the transcription of core clock genes, it has also been shown that the clock proteins in turn interact with the HIF-1α protein. For instance, DEC2 binds to HIF-1α and decreases the binding of HIF-1α to HREs to activate the transcription of the HIF target genes [120]. Co-expression of HIF-1α and BMAL1 has also been shown to significantly transactivate HRE [124], further indicating the interaction between the clock proteins and HIF-1α. These findings are consistent with the observation that HIF-1α target genes show diurnal rhythms in their expression [120,125]. Because the alternation of HIF-1α and DEC expression is closely associated with tumor progression [120,126], it would be of interest to examine whether conditional Hif-1α and Dec knockout mice implanted tumor cells affect the circadian rhythm and metabolism. Collectively, these studies suggest that cellular oxygen homeostasis is tightly coupled to the regulation of daily rhythms in various physiological processes.
7. Conclusions
The clock genes Dec1, Dec2, and Bmal1 are now recognized as important mediators that integrate the function of the circadian and metabolic systems. Dysfunction of the clock genes can therefore provoke both circadian and metabolic disorders. Further efforts to understand how Dec1, Dec2, and Bmal1 sense energy balance and how energy imbalance impacts the function of these clock genes will provide insight into pathophysiologic interactions between circadian disorders such as sleep disturbances and metabolic disorders such as diabetes and obesity in humans.
Acknowledgments
This work was supported by the Japan Society for the Promotion of Science Grants-in-Aid for Scientific Research (KAKENHI; 16K09624 of FS).
Author Contributions
Fuyuki Sato made draft and figures and completed manuscript. Akira Kohsaka helped making draft and figures. Ujjal K. Bhawal and Yasuteru Muragaki corrected draft.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gerhart-Hines, Z.; Lazar, M.A. Circadian metabolism in the light of evolution. Endocr. Rev. 2015, 36, 289–304. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, G.; Young, M. Circadian regulation of metabolic homeostasis: Causes and consequences. Nat. Sci. Sleep 2016, 8, 163–180. [Google Scholar] [PubMed]
- Bass, J.; Takahashi, J.S. Circadian integration of metabolism and energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Schibler, U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell Metab. 2011, 13, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.; Lazar, M.A. Circadian time signatures of fitness and disease. Science 2016, 354, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Sahar, S.; Sassone-Corsi, P. Regulation of metabolism: The circadian clock dictates the time. Trends Endocrinol. Metab. 2012, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [PubMed]
- Schibler, U.; Gotic, I.; Saini, C.; Gos, P.; Curie, T.; Emmenegger, Y.; Sinturel, F.; Gosselin, P.; Gerber, A.; Fleury-Olela, F.; et al. Clock-Talk: Interactions between Central and Peripheral Circadian Oscillators in Mammals. Cold Spring Harb. Symp. Quant. Biol. 2015, 80, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Barnea, M.; Madar, Z.; Froy, O. High-fat diet delays and fasting advances the circadian expression of adiponectin signaling components in mouse liver. Endocrinology 2009, 150, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Yanagihara, H.; Hayashi, Y.; Obi, Y.; Tsuruoka, S.; Takamura, T.; Kaneko, S.; Fujimura, A. Rhythmic messenger ribonucleic acid expression of clock genes and adipocytokines in mouse visceral adipose tissue. Endocrinology 2005, 146, 5631–5636. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Hogenesch, J.B.; Gu, Y.Z.; Jain, S.; Bradfield, C.A. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef] [PubMed]
- Bunger, M.K.; Wilsbacher, L.D.; Moran, S.M.; Clendenin, C.; Radcliffe, L.A.; Hogenesch, J.B.; Simon, M.C.; Takahashi, J.S.; Bradfield, C.A. Mop3 is an essential component of the master circadian pacemaker in mammals. Cell 2000, 103, 1009–1017. [Google Scholar] [CrossRef]
- King, D.P.; Zhao, Y.; Sangoram, A.M.; Wilsbacher, L.D.; Tanaka, M.; Antoch, M.P.; Steeves, T.D.L.; Vitaterna, M.H.; Kornhauser, J.M.; Lowrey, P.L.; et al. Positional cloning of the mouse circadian Clock gene. Cell 1997, 89, 641–653. [Google Scholar] [CrossRef]
- Kume, K.; Zylka, M.J.; Sriram, S.; Shearman, L.P.; Weaver, D.R.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 1999, 98, 193–205. [Google Scholar] [CrossRef]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Ueda, H.R.; Chen, W.; Adachi, A.; Wakamatsu, H.; Hayashi, S.; Takasugi, T.; Nagano, M.; Nakahama, K.; Suzuki, Y.; Sugano, S.; et al. A transcription factor response element for gene expression during circadian night. Nature 2002, 418, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Honma, S.; Kawamoto, T.; Takagi, Y.; Fujimoto, K.; Sato, F.; Noshiro, M.; Kato, Y.; Honma, K. Dec1 and Dec2 are regulators of the mammalian molecular clock. Nature 2002, 419, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Kawamoto, T.; Fujimoto, K.; Noshiro, M.; Honda, K.K.; Honma, S.; Honma, K.; Kato, Y. Functional analysis of the basic helix-loop-helix transcription factor DEC1 in circadian regulation. Interaction with BMAL1. Eur. J. Biochem. 2004, 271, 4409–4419. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, T.; Noshiro, M.; Sato, F.; Maemura, K.; Takeda, N.; Nagai, R.; Iwata, T.; Fujimoto, K.; Furukawa, M.; Miyazaki, K.; et al. A novel autofeedback loop of Dec1 transcription involved in circadian rhythm regulation. Biochem. Biophys. Res. Commun. 2004, 313, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, H.; Fujimoto, K.; Kawamoto, T.; Noshiro, M.; Maemura, K.; Takeda, N.; Nagai, R.; Furukawa, M.; Honma, S.; Honma, K.; et al. Expression of the gene for Dec2, a basic helix-loop-helix transcription factor, is regulated by a molecular clock system. Biochem. J. 2004, 382, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, M.; Song, X.; Gragen, S.; Sachdeva, K.; Wan, Y.; Yan, B. DEC1 negatively regulates the expression of DEC2 through binding to the E-box in the proximal promoter. J. Biol. Chem. 2003, 278, 16899–16907. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, B.; Flock, G.; Zacksenhaus, E.; Egan, S.E. Stra13 homodimers repress transcription through class B E-box elements. J. Biol. Chem. 2002, 277, 46544–46551. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Kawamoto, T.; Honda, K.K.; Ueshima, T.; Noshiro, M.; Iwata, T.; Fujimoto, K.; Kubo, H.; Honma, S.; Yorioka, N.; et al. DEC1 modulates the circadian phase of clock gene expression. Mol. Cell. Biol. 2008, 28, 4080–4092. [Google Scholar] [CrossRef] [PubMed]
- Roesler, W.J.; Khandelwal, R.L. Diurnal variations in the activities of the glycogen metabolizing enzymes in mouse liver. Int. J. Biochem. 1985, 17, 81–85. [Google Scholar] [CrossRef]
- Conlee, R.K.; Rennie, M.J.; Winder, W.W. Skeletal muscle glycogen content: Diurnal variation and effects of fasting. Am. J. Physiol. 1976, 231, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Benavides, A.; Siches, M.; Llobera, M. Circadian rhythms of lipoprotein lipase and hepatic lipase activities in intermediate metabolism of adult rat. Am. J. Physiol. 1998, 275, R811–R817. [Google Scholar] [CrossRef] [PubMed]
- Tasali, E.; Leproult, R.; Ehrmann, D.A.; Van Cauter, E. Slow-wave sleep and the risk of type 2 diabetes in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, K.; Leproult, R.; Van Cauter, E. Impact of sleep debt on metabolic and endocrine function. Lancet 1999, 354, 1435–1439. [Google Scholar] [CrossRef]
- Lumeng, J.C.; Somashekar, D.; Appugliese, D.; Kaciroti, N.; Corwyn, R.F.; Bradley, R.H. Shorter sleep duration is associated with increased risk for being overweight at ages 9 to 12 years. Pediatrics 2007, 120, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, B.; Knutsson, A.; Lindahl, B. Is there an association between shift work and having a metabolic syndrome? Results from a population based study of 27,485 people. Occup. Environ. Med. 2001, 58, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Suwazono, Y.; Dochi, M.; Sakata, K.; Okubo, Y.; Oishi, M.; Tanaka, K.; Kobayashi, E.; Kido, T.; Nogawa, K. A longitudinal study on the effect of shift work on weight gain in male Japanese workers. Obesity 2008, 16, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Biggi, N.; Consonni, D.; Galluzzo, V.; Sogliani, M.; Costa, G. Metabolic syndrome in permanent night workers. Chronobiol. Int. 2008, 25, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.J.L.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Schernhammer, E.S.; Sun, Q.; Hu, F.B. Rotating night shift work and risk of type 2 diabetes: Two prospective cohort studies in women. PLoS Med. 2011, 8, e1001141. [Google Scholar] [CrossRef] [PubMed]
- Suwazono, Y.; Dochi, M.; Oishi, M.; Tanaka, K.; Kobayashi, E.; Sakata, K. Shiftwork and impaired glucose metabolism: A 14-year cohort study on 7104 male workers. Chronobiol. Int. 2009, 26, 926–941. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, A.; Kobiita, A.; Damara, M.; Misra, N.; Meziane, H.; Champy, M.-F.; Chambon, P. Shifting eating to the circadian rest phase misaligns the peripheral clocks with the master SCN clock and leads to a metabolic syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, E6691–E6698. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.A.; Han, D.H.; Noh, J.Y.; Kim, M.H.; Son, G.H.; Kim, K.; Kim, C.J.; Pak, Y.K.; Cho, S. Meal time shift disturbs circadian rhythmicity along with metabolic and behavioral alterations in mice. PLoS ONE 2012, 7, e44053. [Google Scholar] [CrossRef] [PubMed]
- Bode, B.; Shahmoradi, A.; Rossner, M.J.; Oster, H. Genetic Interaction of Per1 and Dec1/2 in the Regulation of Circadian Locomotor Activity. J. Biol. Rhythms 2011, 26, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Makishima, M.; Bhawal, U.K. Differentiated embryo chondrocyte 1 (DEC1) is a novel negative regulator of hepatic fibroblast growth factor 21 (FGF21) in aging mice. Biochem. Biophys. Res. Commun. 2016, 469, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Muragaki, Y.; Kawamoto, T.; Fujimoto, K.; Kato, Y.; Zhang, Y. Rhythmic expression of DEC2 protein in vitro and in vivo. Biomed. Rep. 2016, 4, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Lamia, K.A.; Storch, K.-F.; Weitz, C.J. Physiological significance of a peripheral tissue circadian clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jones, C.R.; Fujiki, N.; Xu, Y.; Guo, B.; Holder, J.L.; Rossner, M.J.; Nishino, S.; Fu, Y.H. The transcriptional repressor DEC2 regulates sleep length in mammals. Science 2009, 325, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Rossner, M.J.; Oster, H.; Wichert, S.P.; Reinecke, L.; Wehr, M.C.; Reinecke, J.; Eichele, G.; Taneja, R.; Nave, K.A. Disturbed clockwork resetting in sharp-1 and sharp-2 single and double mutant mice. PLoS ONE 2008, 3, e2762. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Ushijima, K.; Shimba, S.; Fujimura, A. Daily fasting blood glucose rhythm in male mice: A role of the circadian clock in the liver. Endocrinology 2016, 157, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, S.Y.; Wu, G.; Altman, B.J.; Growe, J.; Rhoades, S.D.; Coldren, F.; Venkataraman, A.; Olarerin-George, A.O.; Francey, L.J.; Mukherjee, S.; et al. Clock Regulation of Metabolites Reveals Coupling between Transcription and Metabolism. Cell Metab. 2017, 25, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Perelis, M.; Marcheva, B.; Ramsey, K.M.; Schipma, M.J.; Hutchison, A.L.; Taguchi, A.; Peek, C.B.; Hong, H.; Huang, W.; Omura, C.; et al. Pancreatic β cell enhancers regulate rhythmic transcription of genes controlling insulin secretion. Science 2015, 350, aac4250. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, B.; Schaad, O.; Bujard, H.; Takahashi, J.S.; Schibler, U. System-driven and oscillator-dependent circadian transcription in mice with a conditionally active liver clock. PLoS Biol. 2007, 5, 0179–0189. [Google Scholar] [CrossRef] [PubMed]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Rakshit, K.; Hsu, T.W.; Matveyenko, A.V. Bmal1 is required for beta cell compensatory expansion, survival and metabolic adaptation to diet-induced obesity in mice. Diabetologia 2016, 59, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Dyar, K.A.; Ciciliot, S.; Wright, L.E.; Biensø, R.S.; Tagliazucchi, G.M.; Patel, V.R.; Forcato, M.; Paz, M.I.P.; Gudiksen, A.; Solagna, F.; et al. Muscle insulin sensitivity and glucose metabolism are controlled by the intrinsic muscle clock. Mol. Metab. 2014, 3, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Shimba, S.; Ogawa, T.; Hitosugi, S.; Ichihashi, Y.; Nakadaira, Y.; Kobayashi, M.; Tezuka, M.; Kosuge, Y.; Ishige, K.; Ito, Y.; et al. Deficient of a clock gene, brain and muscle arnt-like protein-1 (BMAL1), induces dyslipidemia and ectopic fat formation. PLoS ONE 2011, 6, e25231. [Google Scholar] [CrossRef] [PubMed]
- Shimba, S.; Ishii, N.; Ohta, Y.; Ohno, T.; Watabe, Y.; Hayashi, M.; Wada, T.; Aoyagi, T.; Tezuka, M. Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 12071–12076. [Google Scholar] [CrossRef] [PubMed]
- Woon, P.Y.; Kaisaki, P.J.; Braganca, J.; Bihoreau, M.T.; Levy, J.C.; Farrall, M.; Gauguier, D. Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, F.; Matsubara, C.; Myung, J.; Yoritaka, T.; Kamimura, N.; Tsutsumi, S.; Kanai, A.; Suzuki, Y.; Sassone-Corsi, P.; Aburatani, H.; et al. Genome-Wide Profiling of the Core Clock Protein BMAL1 Targets Reveals a Strict Relationship with Metabolism. Mol. Cell. Biol. 2010, 30, 5636–5648. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tang, D.; Liu, N.; Xiong, W.; Huang, H.; Li, Y.; Ma, Z.; Zhao, H.; Chen, P.; Qi, X.; et al. Reciprocal Regulation between the Circadian Clock and Hypoxia Signaling at the Genome Level in Mammals. Cell Metab. 2017, 25, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Trott, A.J.; Menet, J.S. Regulation of circadian clock transcriptional output by CLOCK:BMAL1. PLoS Genet. 2018, 14, e1007156. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Cho, H.J.; Cho, H.; Kim, K.H.; Kim, J.B.; Park, H. Stra13/DEC1 and DEC2 inhibit sterol regulatory element binding protein-1c in a hypoxia-inducible factor-dependent mechanism. Nucleic Acids Res. 2008, 36, 6372–6385. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Cui, A.; Xue, Y.; Cui, Y.; Dong, X.; Gao, Y.; Yang, H.; Fang, F.; Chang, Y. Hepatic differentiated embryo-chondrocyte-expressed gene 1 (Dec1) inhibits sterol regulatory element-binding protein-1c (Srebp-1c) expression and alleviates fatty liver phenotype. J. Biol. Chem. 2014, 289, 23332–23342. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Goda, N.; Kanai, M.; Niwa, D.; Osanai, K.; Yamamoto, Y.; Senoo-Matsuda, N.; Johnson, R.S.; Miura, S.; Kabe, Y.; et al. HIF-1alpha induction suppresses excessive lipid accumulation in alcoholic fatty liver in mice. J. Hepatol. 2012, 56, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Yun, Z.; Maecker, H.L.; Johnson, R.S.; Giaccia, A.J. Inhibition of PPARgamma2 gene expression by the HIF-1-regulated gene DEC1/Stra13: A mechanism for regulation of adipogenesis by hypoxia. Dev. Cell 2002, 2, 331–341. [Google Scholar] [CrossRef]
- Peek, C.B.; Ramsey, K.M.; Marcheva, B.; Bass, J. Nutrient sensing and the circadian clock. Trends Endocrinol. Metab. 2012, 23, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Panda, S. Fasting, Circadian Rhythms, and Time-Restricted Feeding in Healthy Lifespan. Cell Metab. 2016, 23, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Damiola, F.; Le Minli, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, T.; Noshiro, M.; Furukawa, M.; Honda, K.K.; Nakashima, A.; Ueshima, T.; Usui, E.; Katsura, Y.; Fujimoto, K.; Honma, S.; et al. Effects of fasting and re-feeding on the expression of Dec, Per1, and other clock-related genes. J. Biochem. 2006, 140, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Noshiro, M.; Usui, E.; Kawamoto, T.; Sato, F.; Nakashima, A.; Ueshima, T.; Honda, K.; Fujimoto, K.; Honma, S.; Honma, K.I.; et al. Liver X receptors (LXRalpha and LXRbeta) are potent regulators for hepatic Dec1 expression. Genes Cells 2009, 14, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Oike, H.; Nagai, K.; Fukushima, T.; Ishida, N.; Kobori, M. Feeding cues and injected nutrients induce acute expression of multiple clock genes in the mouse liver. PLoS ONE 2011, 6, e23709. [Google Scholar] [CrossRef] [PubMed]
- Rutter, J.; Reick, M.; Wu, L.C.; McKnight, S.L. Regulation of crock and NPAS2 DNA binding by the redox state of NAD cofactors. Science 2001, 293, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Rey, G.; Valekunja, U.K.; Feeney, K.A.; Wulund, L.; Milev, N.B.; Stangherlin, A.; Ansel-Bollepalli, L.; Velagapudi, V.; O’Neill, J.S.; Reddy, A.B. The Pentose Phosphate Pathway Regulates the Circadian Clock. Cell Metab. 2016, 24, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Hirano, A.; Braas, D.; Fu, Y.H.; Ptáček, L.J. FAD Regulates CRYPTOCHROME Protein Stability and Circadian Clock in Mice. Cell Rep. 2017, 19, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, K.M.; Yoshino, J.; Brace, C.S.; Abrassart, D.; Kobayashi, Y.; Marcheva, B.; Hong, H.K.; Chong, J.L.; Buhr, E.D.; Lee, C.; et al. Circadian clock feedback cycle through NAMPT-Mediated NAD+ biosynthesis. Science 2009, 324, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.A.; Yu, Y.V.; Govindaiah, G.; Ye, X.; Artinian, L.; Coleman, T.P.; Sweedler, J.V.; Cox, C.L.; Gillette, M.U. Circadian rhythm of redox state regulates excitability in suprachiasmatic nucleus neurons. Science 2012, 337, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Bordone, L.; Motta, M.C.; Picard, F.; Robinson, A.; Jhala, U.S.; Apfeld, J.; McDonagh, T.; Lemieux, M.; McBurney, M.; Szilvasi, A.; et al. Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic β cells. PLoS Biol. 2006, 4, e31. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsy, R.; Cohen, H.Y.; et al. Stress-Dependent Regulation of FOXO Transcription Factors by the SIRT1 Deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Motta, M.C.; Divecha, N.; Lemieux, M.; Kamel, C.; Chen, D.; Gu, W.; Bultsma, Y.; McBurney, M.; Guarente, L. Mammalian SIRT1 Represses Forkhead Transcription Factors. Cell 2004, 116, 551–563. [Google Scholar] [CrossRef]
- Li, X.; Zhang, S.; Blander, G.; Tse, J.G.; Krieger, M.; Guarente, L. SIRT1 Deacetylates and Positively Regulates the Nuclear Receptor LXR. Mol. Cell 2007, 28, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-Dependent Deacetylase SIRT1 Modulates CLOCK-Mediated Chromatin Remodeling and Circadian Control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 Regulates Circadian Clock Gene Expression through PER2 Deacetylation. Cell 2008, 134, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, S.; Fergusson, M.M.; Finkel, T. SIRT1 functionally interacts with the metabolic regulator and transcriptional coactivator PGC-1α. J. Biol. Chem. 2005, 280, 16456–16460. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, S.; Liu, T.; Borjigin, J.; Lin, J.D. Transcriptional coactivator PGC-1alpha integrates the mammalian clock and energy metabolism. Nature 2007, 447, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Guarente, L. SIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging. Cell 2013, 153, 1448–1460. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Reinke, H.; Altmeyer, M.; Gutierrez-Arcelus, M.; Hottiger, M.O.; Schibler, U. Poly (ADP-Ribose) Polymerase 1 Participates in the Phase Entrainment of Circadian Clocks to Feeding. Cell 2010, 142, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Stubblefield, J.J.; Terrien, J.; Green, C.B. Nocturnin: At the crossroads of clocks and metabolism. Trends Endocrinol. Metab. 2012, 23, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, E.-K. AMP-activated protein kinase as a key molecular link between metabolism and clockwork. Exp. Mol. Med. 2013, 45, e33. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.D.; Lamia, K.A. AMPK at the crossroads of circadian clocks and metabolism. Mol. Cell. Endocrinol. 2013, 366, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, D.G. AMPK: Positive and negative regulation, and its role in whole-body energy homeostasis. Curr. Opin. Cell Biol. 2015, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.C.; Zierath, J.R. AMP-activated protein kinase signaling in metabolic regulation. J. Clin. Investig. 2006, 116, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Lamia, K.A.; Sachdeva, U.M.; DiTacchio, L.; Williams, E.C.; Alvarez, J.G.; Egan, D.F.; Vasquez, D.S.; Juguilon, H.; Panda, S.; Shaw, R.J.; et al. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science 2009, 326, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Jee, H.U.; Yang, S.; Yamazaki, S.; Kang, H.; Viollet, B.; Foretz, M.; Chung, J.H. Activation of 5′-AMP-activated kinase with diabetes drug metformin induces casein kinase Iε (CKIε)-dependent degradation of clock protein mPer2. J. Biol. Chem. 2007, 282, 20794–20798. [Google Scholar]
- Um, J.H.; Pendergast, J.S.; Springer, D.A.; Foretz, M.; Viollet, B.; Brown, A.; Kim, M.K.; Yamazaki, S.; Chung, J.H. AMPK regulates circadian rhythms in a tissue- and isoform-specific manner. PLoS ONE 2011, 6, e18450. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Muragaki, Y.; Zhang, Y. DEC1 negatively regulates AMPK activity via LKB1. Biochem. Biophys. Res. Commun. 2015, 467, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Okano, T.; Kokame, K.; Shirotani-Ikejima, H.; Miyata, T.; Fukada, Y. Glucose down-regulates Per1 and Per2 mRNA levels and induces circadian gene expression in cultured rat-1 fibroblasts. J. Biol. Chem. 2002, 277, 44244–44251. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Huang, Z.; Lin, L.; Fu, M.; Song, Y.; Shen, Y.; Ren, D.; Gao, Y.; Su, Y.; Zou, Y.; et al. miRNA-130b is required for the ERK/FOXM1 pathway activation-mediated protective effects of isosorbide dinitrate against mesenchymal stem cell senescence induced by high glucose. Int. J. Mol. Med. 2015, 35, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Stephan, F.K.; Davidson, A.J. Glucose, but not fat, phase shifts the feeding-entrained circadian clock. Physiol. Behav. 1998, 65, 277–288. [Google Scholar] [CrossRef]
- Hirao, A.; Tahara, Y.; Kimura, I.; Shibata, S. A balanced diet is necessary for proper entrainment signals of the mouse liver clock. PLoS ONE 2009, 4, e6909. [Google Scholar] [CrossRef] [PubMed]
- Mitro, N.; Mak, P.A.; Vargas, L.; Godio, C.; Hampton, E.; Molteni, V.; Kreusch, A.; Saez, E. The nuclear receptor LXR is a glucose sensor. Nature 2007, 445, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Sun, X.; Ma, X.; Wu, R.; Zhang, D.; Chen, Y.; Xu, Q.; Wu, Y.; Liu, Y. Insulin post-transcriptionally modulates Bmal1 protein to affect the hepatic circadian clock. Nat. Commun. 2016, 7, 12696. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Asano, K.; Komatsu, Y.; Takagi, K.; Ono, M.; Tanaka, T.; Tomita, K.; Haneishi, A.; Tsukada, A.; Yamada, K. Induction of the SHARP-2 mRNA level by insulin is mediated by multiple signaling pathways. Biosci. Biotechnol. Biochem. 2017, 81, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Asano, K.; Haneishi, A.; Ono, M.; Komatsu, Y.; Yamamoto, T.; Tanaka, T.; Ueno, H.; Ogawa, W.; Tomita, K.; et al. Insulin stimulates the expression of the SHARP-1 gene via multiple signaling pathways. Horm. Metab. Res. 2014, 46, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Kawata, H.; Shou, Z.; Mizutani, T.; Noguchi, T.; Miyamoto, K. Insulin induces the expression of the SHARP-2/Stra13/DEC1 gene via a phosphoinositide 3-kinase pathway. J. Biol. Chem. 2003, 278, 30719–30724. [Google Scholar] [CrossRef] [PubMed]
- Yamajuku, D.; Inagaki, T.; Haruma, T.; Okubo, S.; Kataoka, Y.; Kobayashi, S.; Ikegami, K.; Laurent, T.; Kojima, T.; Noutomi, K.; et al. Real-time monitoring in three-dimensional hepatocytes reveals that insulin acts as a synchronizer for liver clock. Sci. Rep. 2012, 2, srep00439. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, G. PPARs integrate the mammalian clock and energy metabolism. PPAR Res. 2014, 2014, 653017. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Park, H. Differentiated embryo chondrocyte 1 (DEC1) represses PPARgamma2 gene through interacting with CCAAT/enhancer binding protein beta (C/EBPbeta). Mol. Cells 2012, 33, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, T.; Mei, W.; Li, D.; Cai, R.; Zuo, Y.; Cheng, J. Small ubiquitin-like modifier (SUMO) protein-specific protease 1 de-SUMOylates sharp-1 protein and controls adipocyte differentiation. J. Biol. Chem. 2014, 289, 22358–22364. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Kawamoto, T.; Sasabe, E.; Miyazaki, K.; Fujimoto, K.; Noshiro, M.; Kurihara, H.; Kato, Y. Effects of overexpression of basic helix-loop-helix transcription factor Dec1 on osteogenic and adipogenic differentiation of mesenchymal stem cells. Eur. J. Cell Biol. 2006, 85, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 controls lipid metabolism by direct regulation of PPARγ. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Paschos, G.K.; Ibrahim, S.; Song, W.L.; Kunieda, T.; Grant, G.; Reyes, T.M.; Bradfield, C.A.; Vaughan, C.H.; Eiden, M.; Masoodi, M.; et al. Obesity in mice with adipocyte-specific deletion of clock component Arntl. Nat. Med. 2012, 18, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, C.; Dubois, G.; Duguay, Y.; Helledie, T.; Vu-Dac, N.; Gervois, P.; Soncin, F.; Mandrup, S.; Fruchart, J.C.; Fruchart-Najib, J.; et al. The orphan nuclear receptor Rev-Erbalpha is a peroxisome proliferator-activated receptor (PPAR) gamma target gene and promotes PPARgamma-induced adipocyte differentiation. J. Biol. Chem. 2003, 278, 37672–37680. [Google Scholar] [CrossRef] [PubMed]
- Lemberger, T.; Saladin, R.; Vázquez, M.; Assimacopoulos, F.; Staels, B.; Desvergne, B.; Wahli, W.; Auwerx, J. Expression of the peroxisome proliferator-activated receptor alpha gene is stimulated by stress and follows a diurnal rhythm. J. Biol. Chem. 1996, 271, 1764–1769. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Shirai, H.; Ishida, N. CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated receptor alpha (PPARalpha) in mice. Biochem. J. 2005, 386, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, I.; Ripperger, J.A.; Baeriswyl-Aebischer, S.; Albrecht, U. The mammalian clock component PERIOD2 coordinates circadian output by interaction with nuclear receptors. Genes Dev. 2010, 24, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Canaple, L.; Rambaud, J.; Dkhissi-Benyahya, O.; Rayet, B.; Tan, N.S.; Michalik, L.; Delaunay, F.; Wahli, W.; Laudet, V. Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol. Endocrinol. 2006, 20, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervois, P.; Chopin-Delannoy, S.; Fadel, A.; Dubois, G.; Kosykh, V.; Fruchart, J.C.; Najib, J.; Laudet, V.; Staels, B. Fibrates increase human REV-ERBalpha expression in liver via a novel peroxisome proliferator-activated receptor response element. Mol. Endocrinol. 1999, 13, 400–409. [Google Scholar] [PubMed]
- Sato, F.; Bhawal, U.K.; Kawamoto, T.; Fujimoto, K.; Imaizumi, T.; Imanaka, T.; Kondo, J.; Koyanagi, S.; Noshiro, M.; Yoshida, H.; et al. Basic-helix-loop-helix (bHLH) transcription factor DEC2 negatively regulates vascular endothelial growth factor expression. Genes Cells 2008, 13, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.; Kawamoto, T.; Tanimoto, K.; Nishiyama, M.; Honda, H.; Kato, Y. Identification of functional hypoxia response elements in the promoter region of the DEC1 and DEC2 genes. J. Biol. Chem. 2002, 277, 47014–47021. [Google Scholar] [CrossRef] [PubMed]
- Wykoff, C.C.; Pugh, C.W.; Maxwell, P.H.; Harris, A.L.; Ratcliffe, P.J. Identification of novel hypoxia dependent and independent target genes of the von Hippel-Lindau (VHL) tumour suppressor by mRNA differential expression profiling. Oncogene 2000, 19, 6297–6305. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.V.; Ivanov, S.V.; Danilkovitch-Miagkova, A.; Lerman, M.I. Regulation of STRA13 by the von Hippel-Lindau Tumor Suppressor Protein, Hypoxia, and the UBC9/Ubiquitin Proteasome Degradation Pathway. J. Biol. Chem. 2001, 276, 15306–15315. [Google Scholar] [CrossRef] [PubMed]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; McNulty, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1alpha Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.; Kumagai, M.; Nakajima, Y.; Shirotake, S.; Kodaira, K.; Oyama, M.; Ueno, M.; Ikeda, M. The impact of HIF1α on the Per2 circadian rhythm in renal cancer cell lines. PLoS ONE 2014, 9, e109693. [Google Scholar] [CrossRef] [PubMed]
- Sato, F.; Bhawal, U.K.; Yoshimura, T.; Muragaki, Y. DEC1 and DEC2 crosstalk between circadian rhythm and tumor progression. J. Cancer 2016, 7, 153–159. [Google Scholar] [CrossRef] [PubMed]
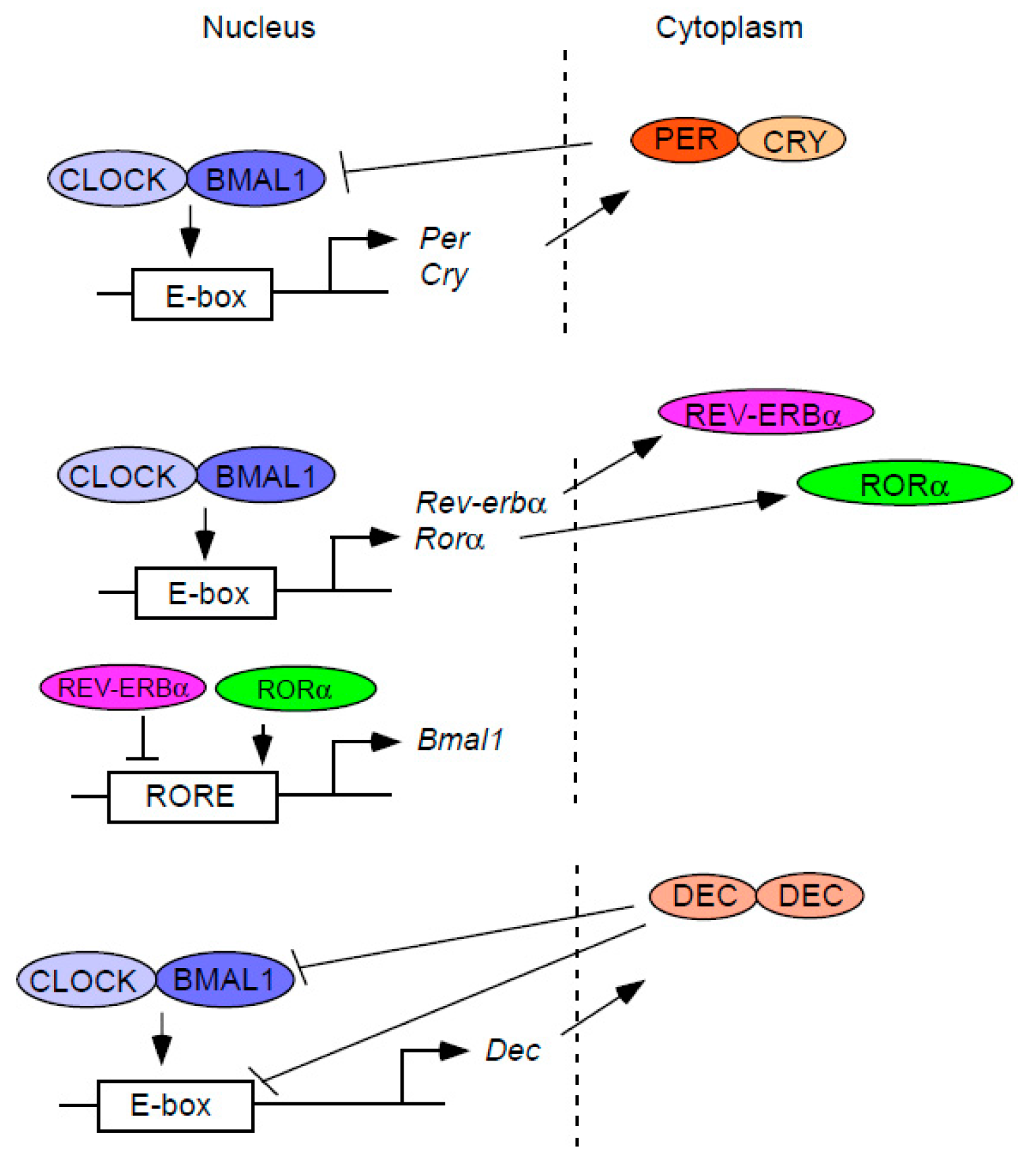
Figure 1.
The core molecular mechanism of the circadian clock. The molecular clock is composed of diverse autoregulatory feedback loops. The core components of the molecular clock, CLOCK and BMAL1, activate the transcription of repressor clock genes such as Per, Cry, Rev-erbα, and Dec. CLOCK and BMAL1 also activate the transcription of inducer Rorα. The protein products of these repressor or inducer genes inhibit or promote their own transcription in distinct ways, thus forming various feedback loops that create a coordinated circadian rhythm at the gene expression level.
Figure 1.
The core molecular mechanism of the circadian clock. The molecular clock is composed of diverse autoregulatory feedback loops. The core components of the molecular clock, CLOCK and BMAL1, activate the transcription of repressor clock genes such as Per, Cry, Rev-erbα, and Dec. CLOCK and BMAL1 also activate the transcription of inducer Rorα. The protein products of these repressor or inducer genes inhibit or promote their own transcription in distinct ways, thus forming various feedback loops that create a coordinated circadian rhythm at the gene expression level.

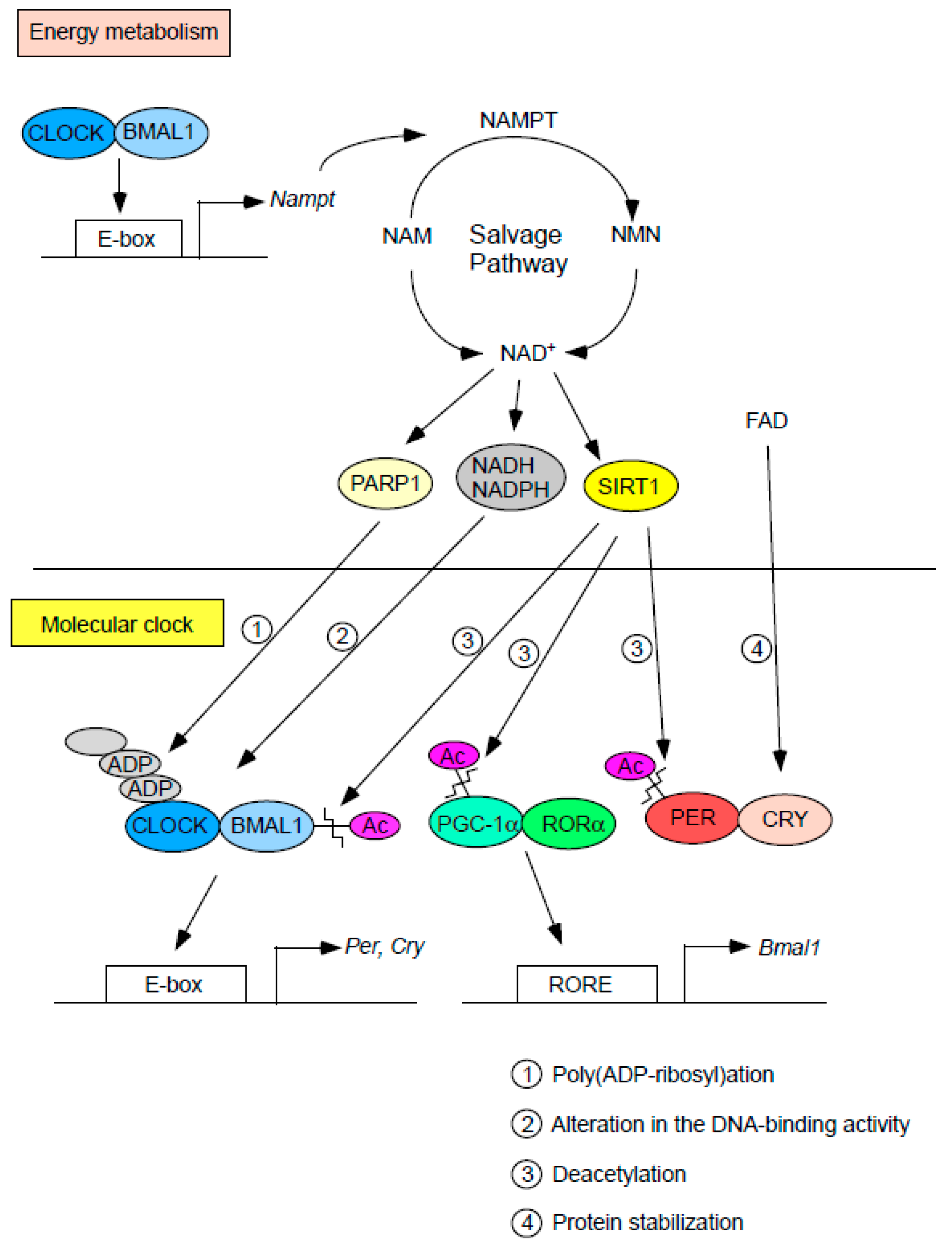
Figure 2.
Interplay between cellular redox factors and the molecular clock. Cellular redox status directly and indirectly affects the function of the molecular clock through changes in the levels of cofactors (i.e., NADH, NADPH, and FAD) and/or the activity of NAD+-consuming enzymes (SIRT1 and PARP1). The molecular clock in turn regulates the transcription of the Nampt gene, which encodes the rate-limiting enzyme in the NAD+ salvage pathway.
Figure 2.
Interplay between cellular redox factors and the molecular clock. Cellular redox status directly and indirectly affects the function of the molecular clock through changes in the levels of cofactors (i.e., NADH, NADPH, and FAD) and/or the activity of NAD+-consuming enzymes (SIRT1 and PARP1). The molecular clock in turn regulates the transcription of the Nampt gene, which encodes the rate-limiting enzyme in the NAD+ salvage pathway.

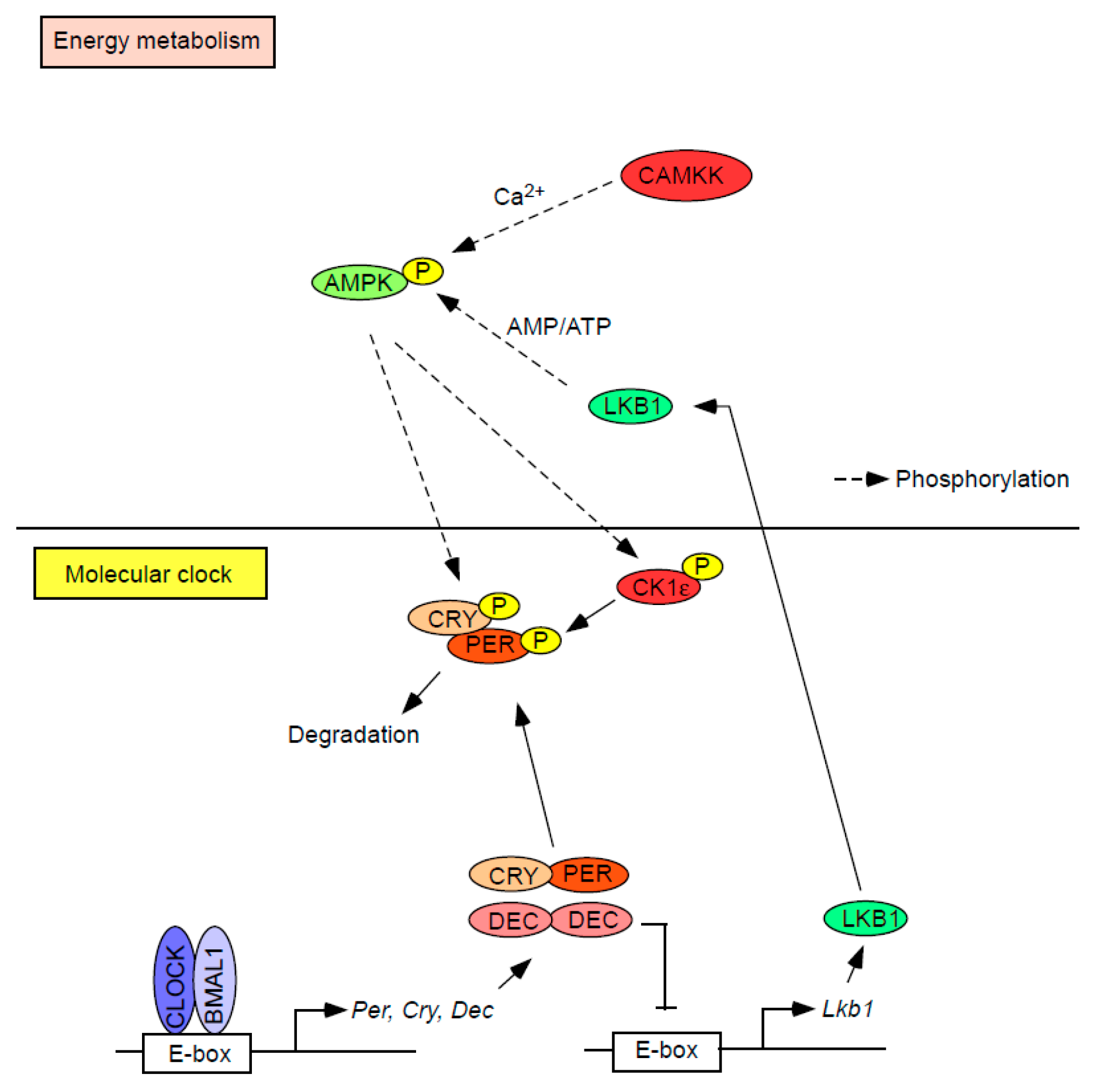
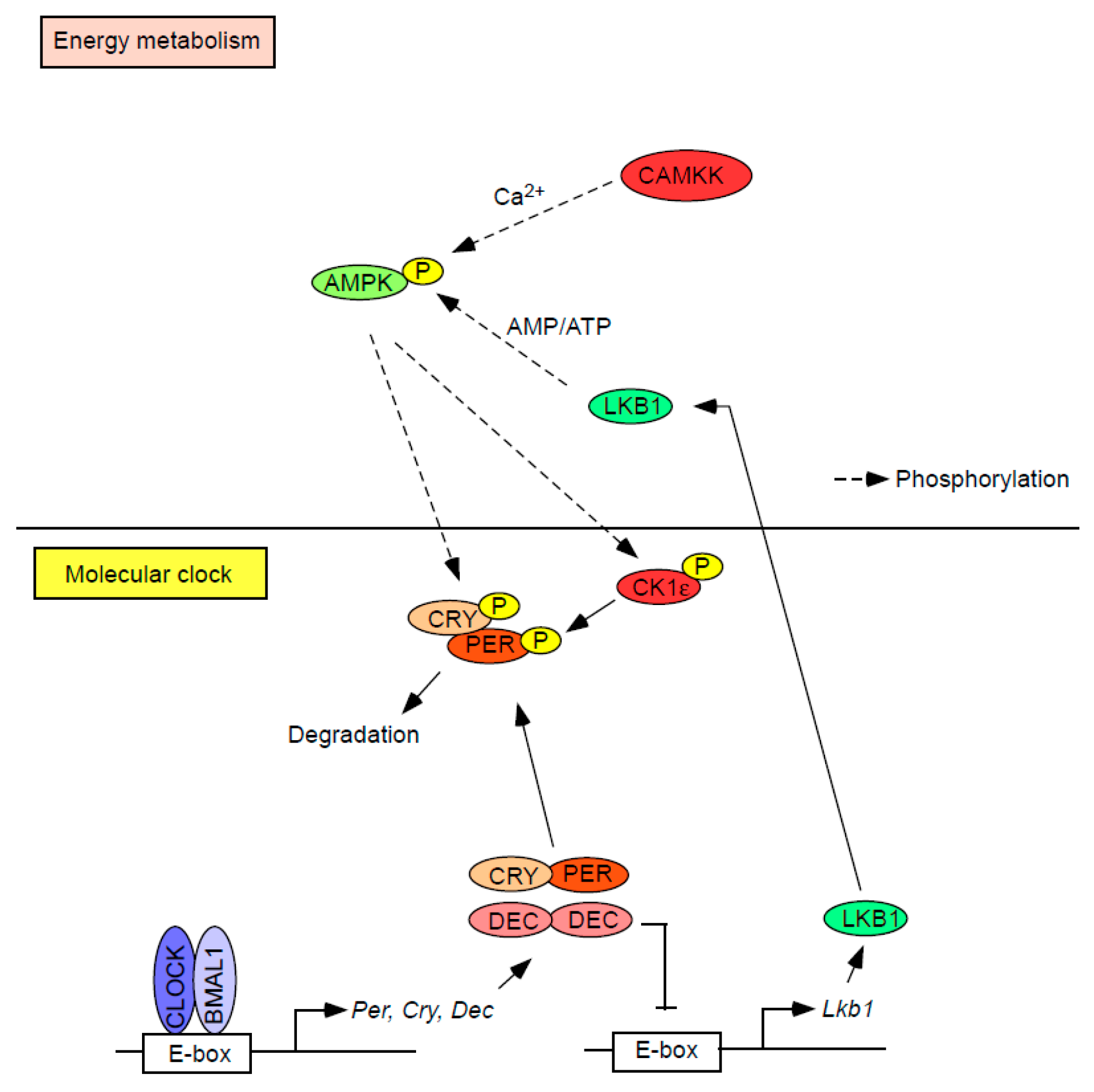
Figure 3.
Interplay between the regulation of bioenergy and the molecular clock. Activated AMPK directly or indirectly phosphorylates core circadian repressors (i.e., PER/CRY). DEC, another circadian repressor, suppresses the transcription of Lkb1 which codes for a serine-threonine kinase that directly phosphorylates and activates AMPK, thus forming a reciprocal relationship between regulators of bioenergy and the circadian clock.
Figure 3.
Interplay between the regulation of bioenergy and the molecular clock. Activated AMPK directly or indirectly phosphorylates core circadian repressors (i.e., PER/CRY). DEC, another circadian repressor, suppresses the transcription of Lkb1 which codes for a serine-threonine kinase that directly phosphorylates and activates AMPK, thus forming a reciprocal relationship between regulators of bioenergy and the circadian clock.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sato, F.; Kohsaka, A.; Bhawal, U.K.; Muragaki, Y. Potential Roles of Dec and Bmal1 Genes in Interconnecting Circadian Clock and Energy Metabolism. Int. J. Mol. Sci. 2018, 19, 781. https://doi.org/10.3390/ijms19030781
AMA Style
Sato F, Kohsaka A, Bhawal UK, Muragaki Y. Potential Roles of Dec and Bmal1 Genes in Interconnecting Circadian Clock and Energy Metabolism. International Journal of Molecular Sciences. 2018; 19(3):781. https://doi.org/10.3390/ijms19030781
Chicago/Turabian StyleSato, Fuyuki, Akira Kohsaka, Ujjal K. Bhawal, and Yasuteru Muragaki. 2018. "Potential Roles of Dec and Bmal1 Genes in Interconnecting Circadian Clock and Energy Metabolism" International Journal of Molecular Sciences 19, no. 3: 781. https://doi.org/10.3390/ijms19030781
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.