Main Olfactory and Vomeronasal Epithelium Are Differently Affected in Niemann-Pick Disease Type C1


Abstract
:1. Introduction
2. Results
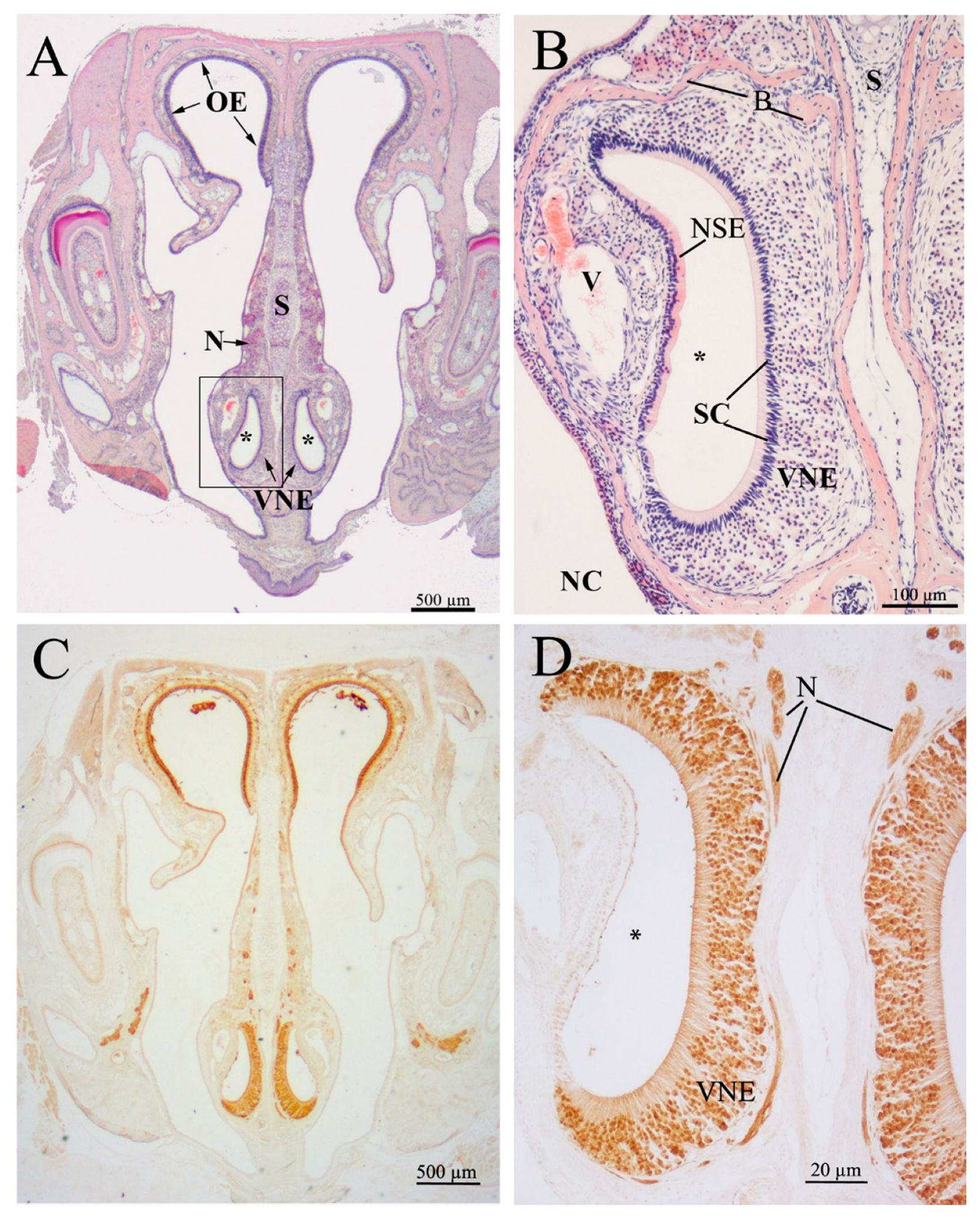
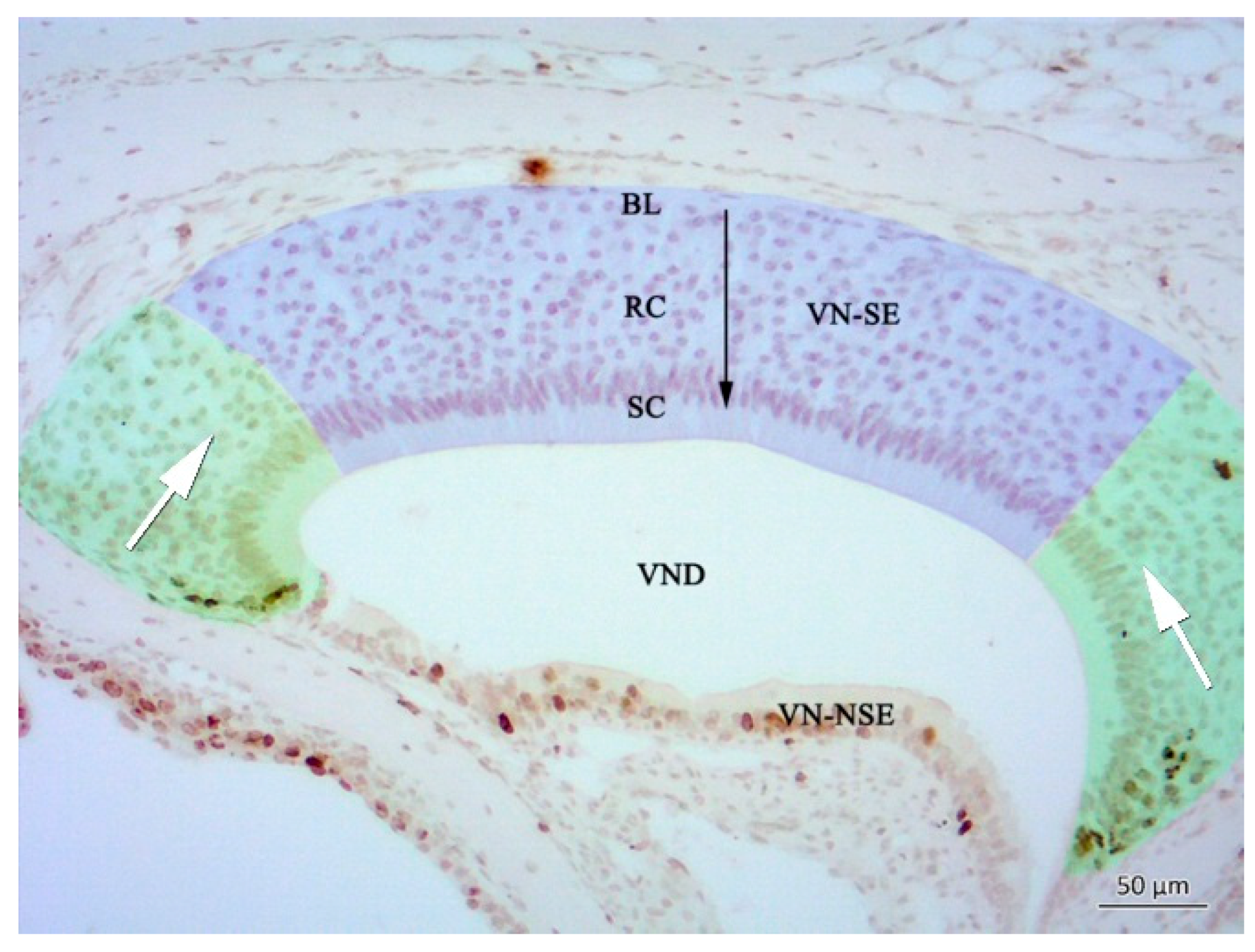
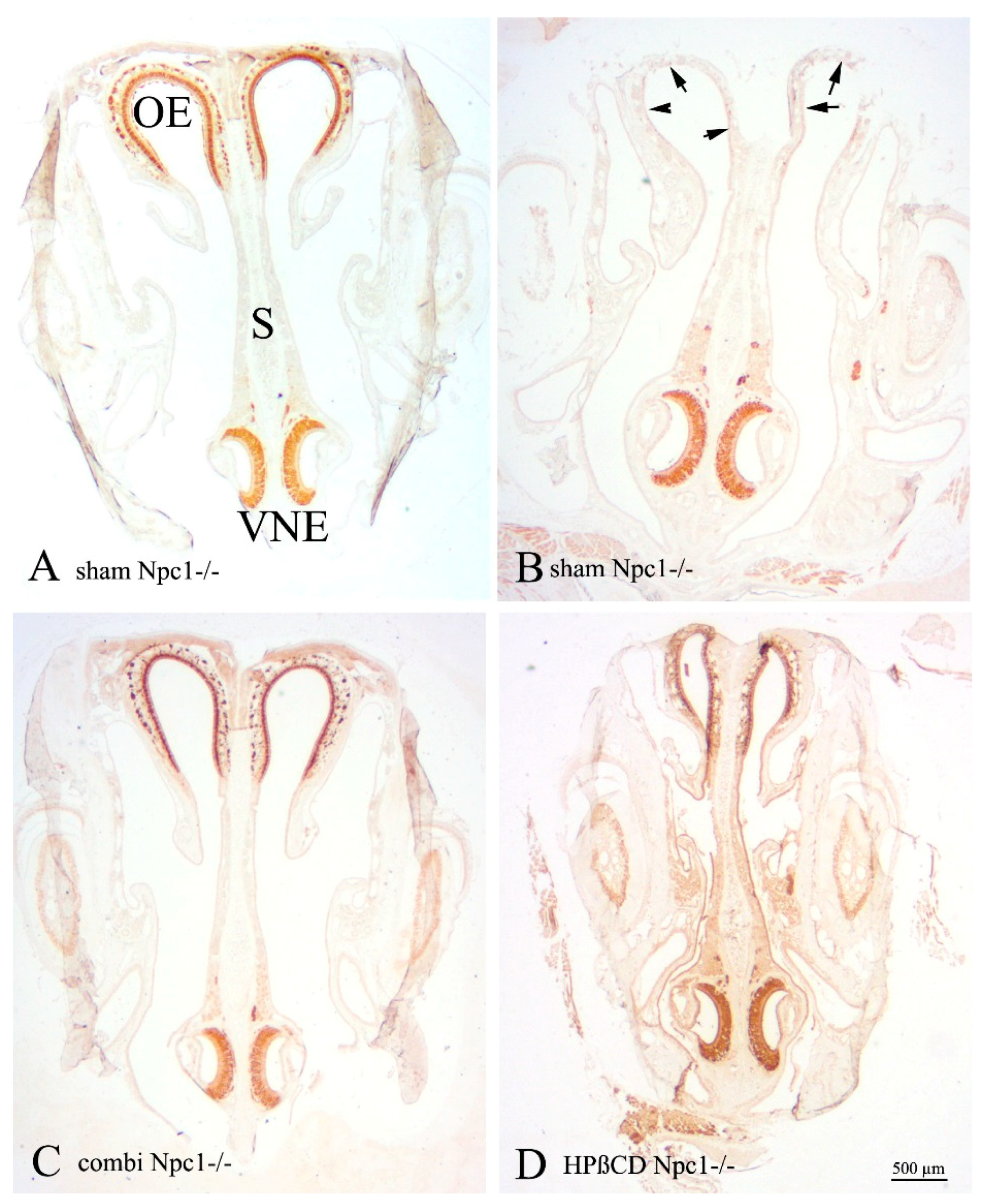
2.1. Histology of the Vomeronasal Epithelium (VNE)
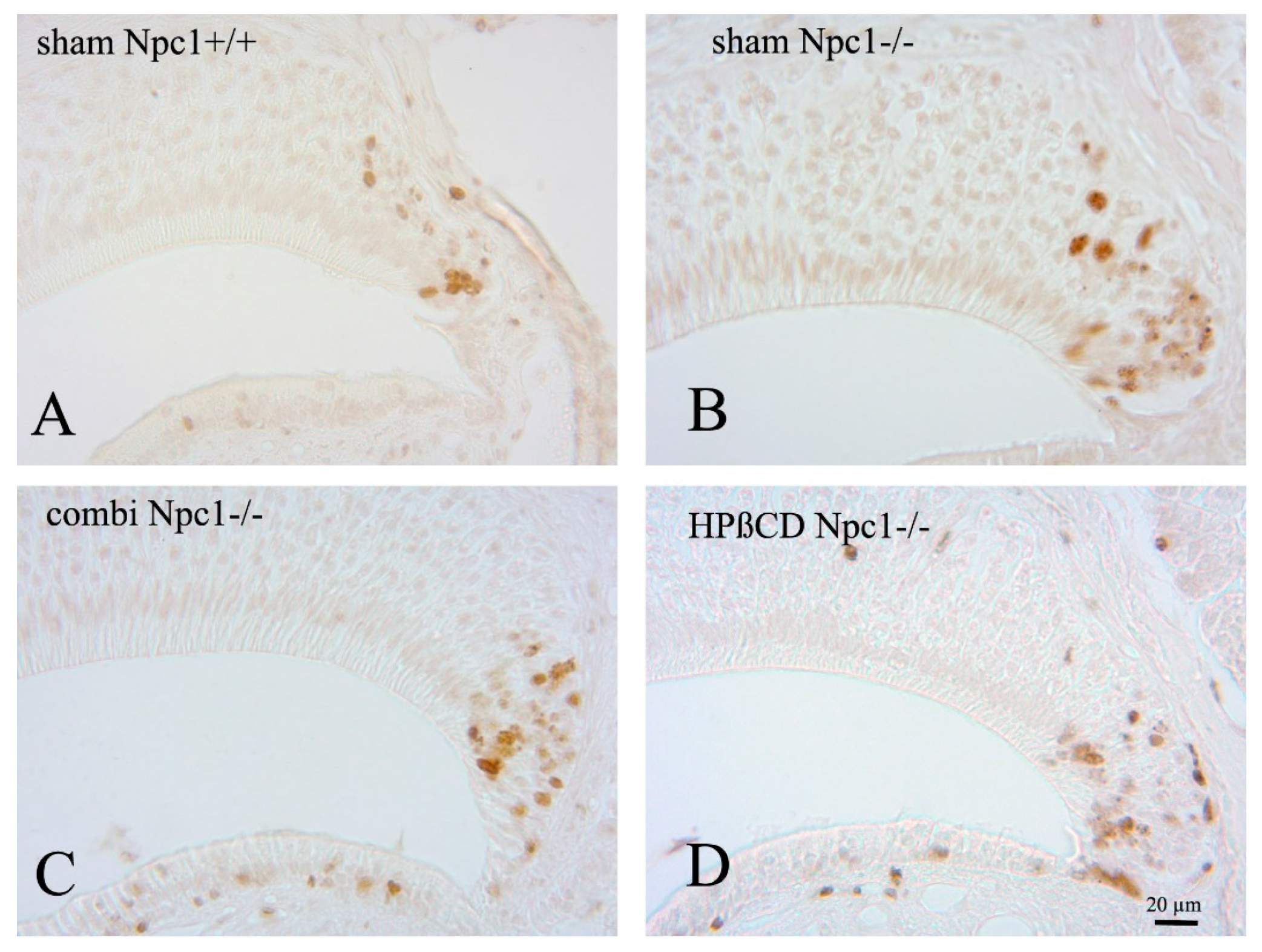
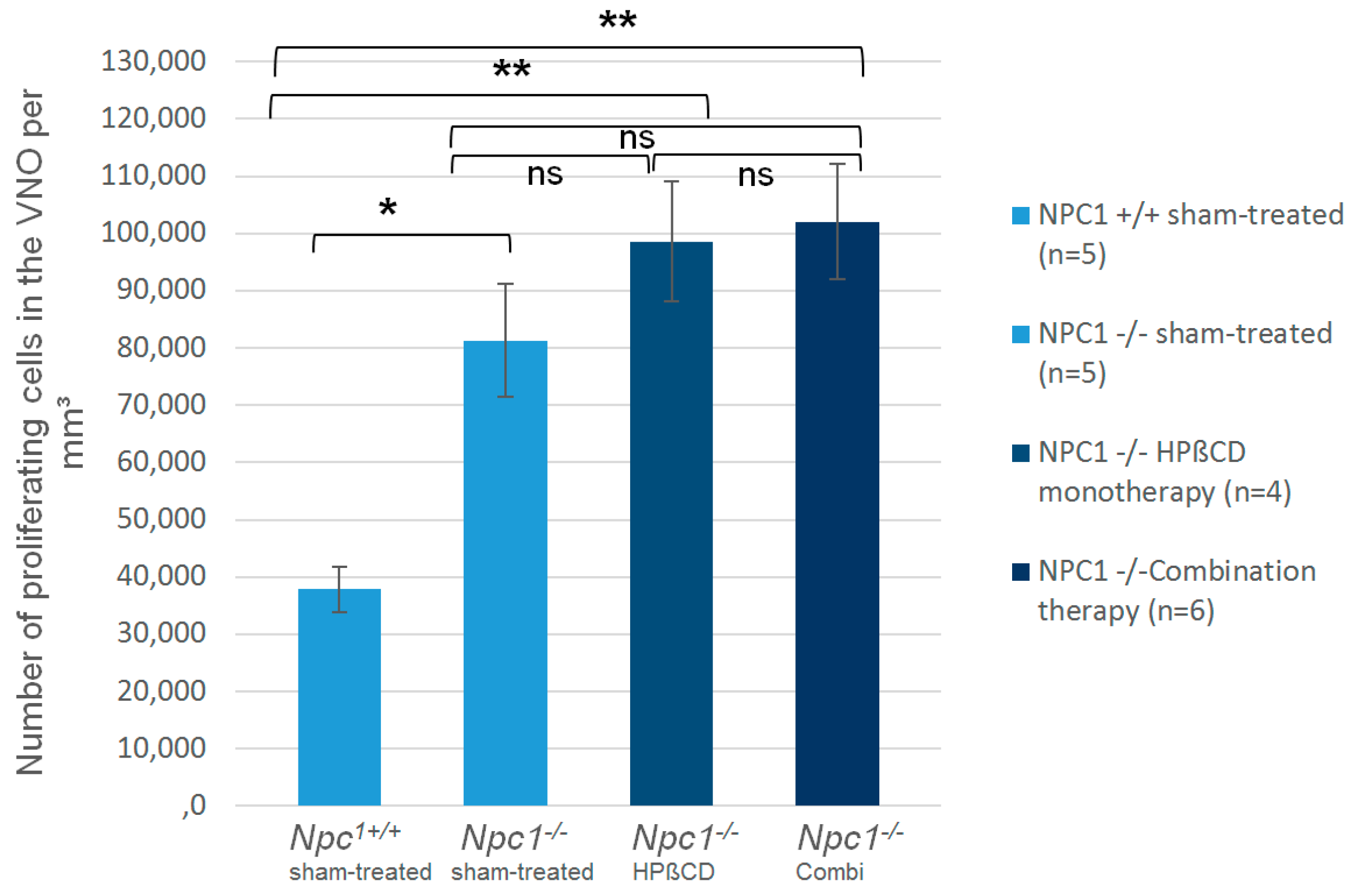
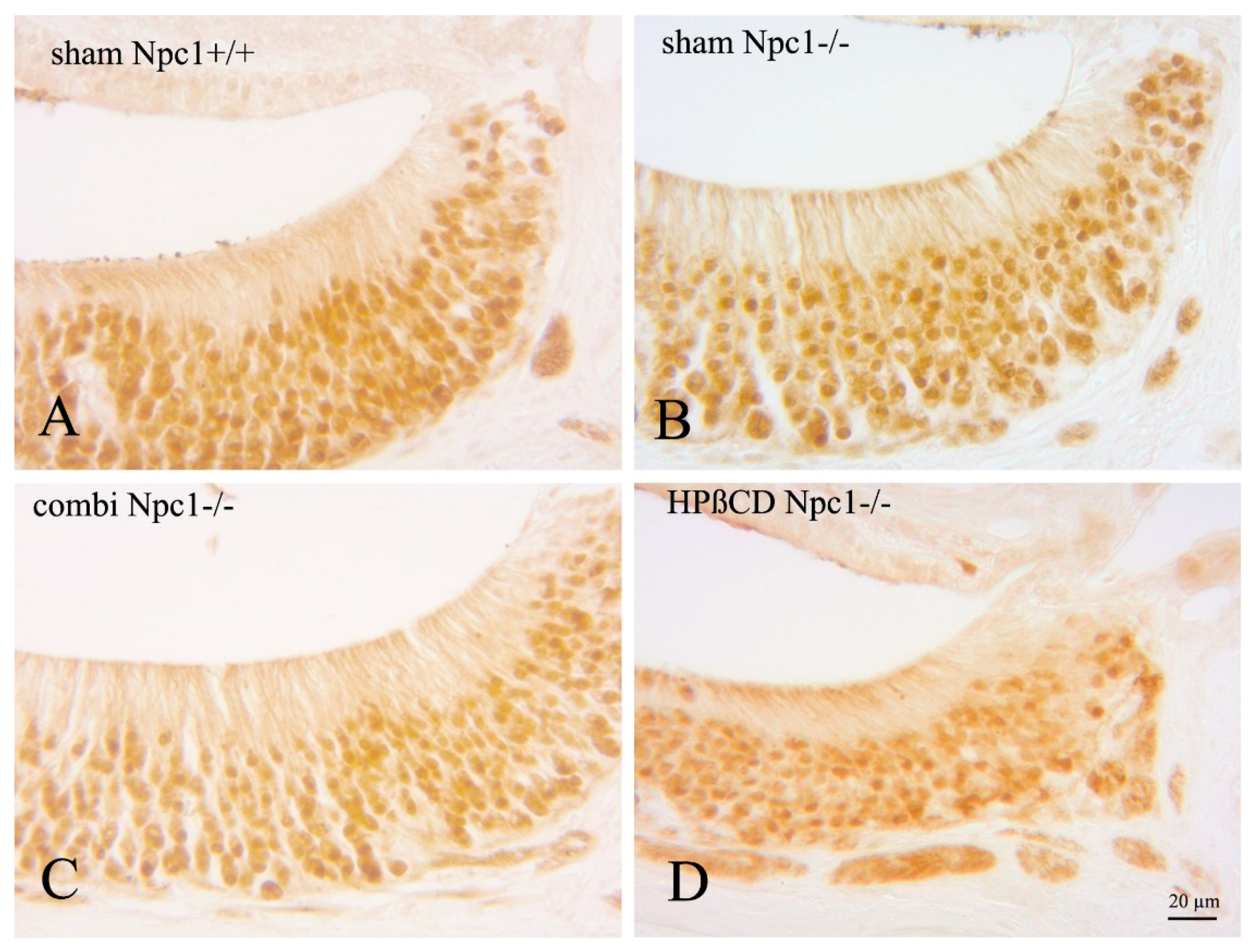
2.2. Quantification of BrdU(+) Proliferating Cells
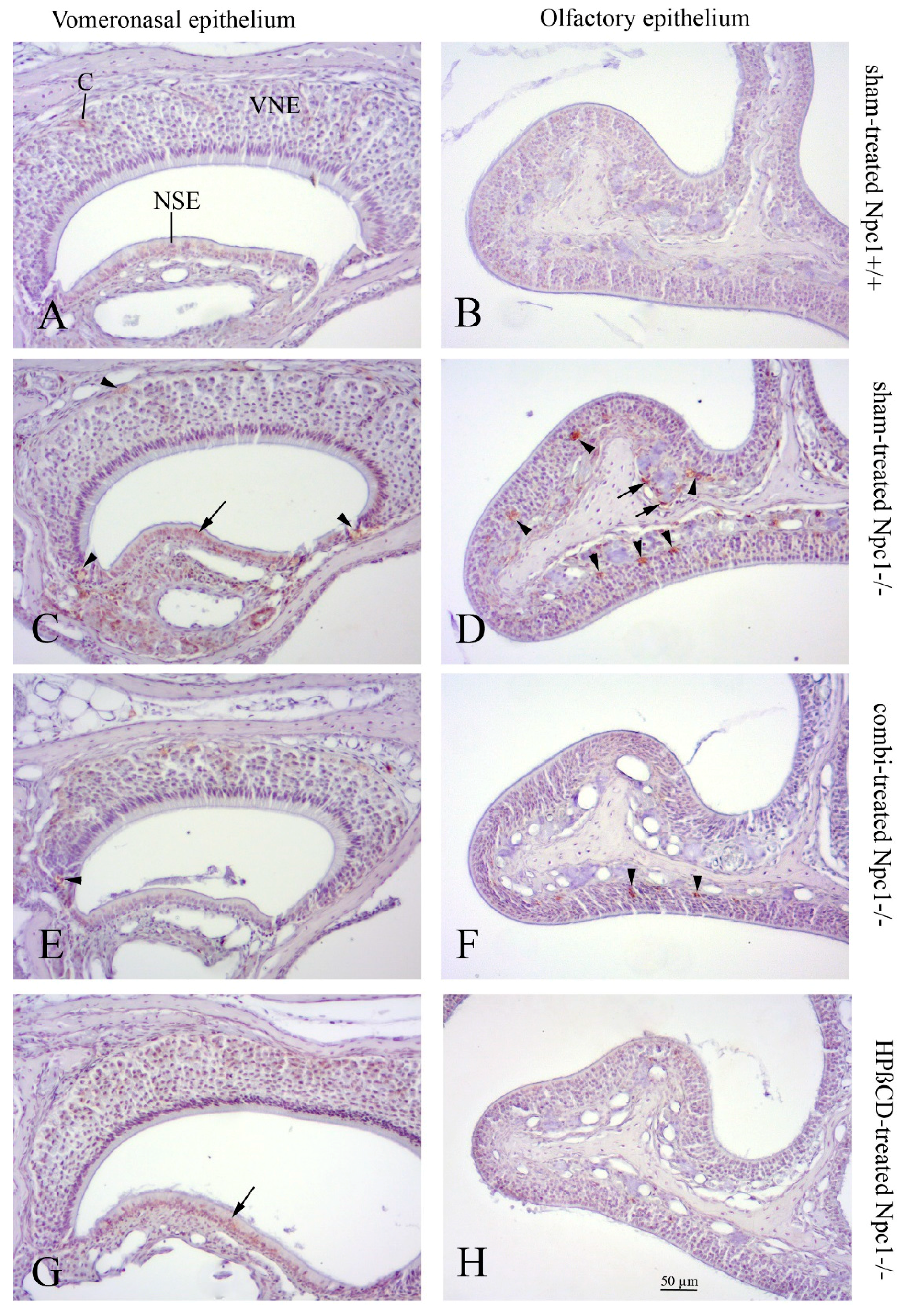
2.3. Caspase-3(+) Immunohistochemistry
2.4. OMP(+) Mature ORNs and Vomeronasal Receptor Neurons
2.5. Cathepsin D Immunohistochemistry
3. Discussion
3.1. Vomeronasal vs. Main Olfactory Epithelium
3.2. VNE Proliferation is Enhanced in Npc1−/− Mice, But Without Signs of Degeneration
3.3. Distinct Differences of Regeneration Mechanisms in OE and VNE
3.4. Treatment: Both Strategies Do Increase Proliferation, But Do Not Change the VNE Phenotype
4. Methods
4.1. Animals
4.2. Genotyping
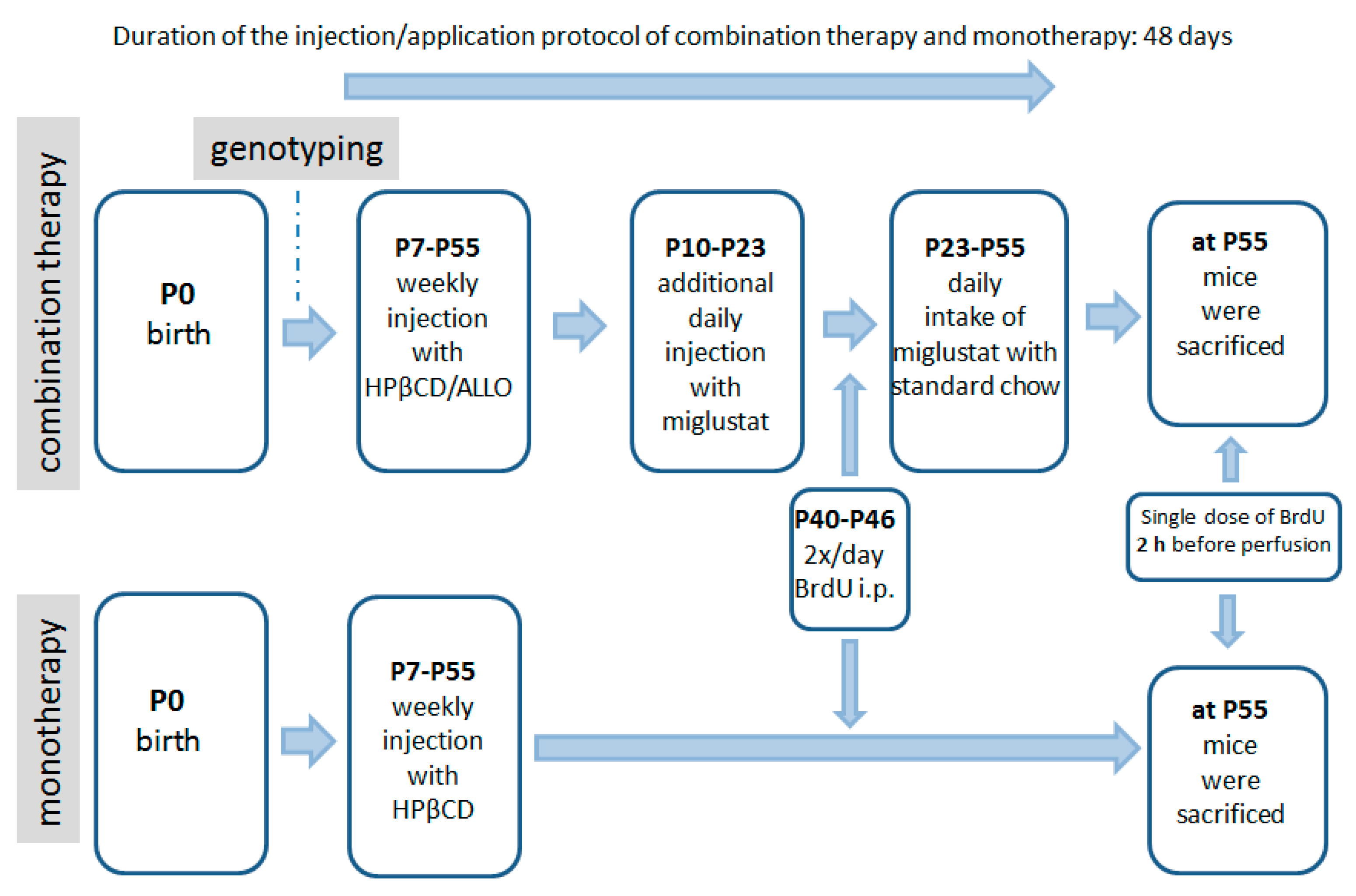
4.3. Pharmacologic Treatment
4.4. BrdU Injections
4.5. Sample Preparation
4.6. Histology and Immunohistochemistry
4.7. Stereology and Statistic Evaluation
5. Conclusions and Hypotheses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BrdU | 5-bromo-2′-deoxyuridine |
| cas-3 | cleaved caspase-3 |
| Cath D | cathepsin D |
| HPβCD | 2-hydroxypropyl-β-cyclodextrin |
| Npc1 | NPC1 gene |
| NPC1 | NPC protein 1 |
| OE | olfactory epithelium |
| OMP | olfactory marker protein |
| ORN | olfactory receptor neuron |
| VNE | vomeronasal epithelium |
| VNO | vomeronasal organ |
References
- Vanier, M.T.; Rodriguez-Lafrasse, C.; Rousson, R.; Gazzah, N.; Juge, M.C.; Pentchev, P.G.; Revol, A.; Louisot, P. Type C Niemann-Pick disease: Spectrum of phenotypic variation in disruption of intracellular LDL-derived cholesterol processing. Biochim. Biophys. Acta 1991, 1096, 328–337. [Google Scholar] [CrossRef]
- Higgins, J.J.; Patterson, M.C.; Dambrosia, J.M.; Pikus, A.T.; Pentchev, P.G.; Sato, S.; Brady, R.O.; Barton, N.W. A clinical staging classification for type C Niemann-Pick disease. Neurology 1992, 42, 2286–2290. [Google Scholar] [CrossRef] [PubMed]
- Carstea, E.D.; Morris, J.A.; Coleman, K.G.; Loftus, S.K.; Zhang, D.; Cummings, C.; Gu, J.; Rosenfeld, M.A.; Pavan, W.J.; Krizman, D.B.; et al. Niemann-Pick C1 disease gene: Homology to mediators of cholesterol homeostasis. Science 1997, 277, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Pentchev, P.G.; Comly, M.E.; Kruth, H.S.; Tokoro, T.; Butler, J.; Sokol, J.; Filling-Katz, M.; Quirk, J.M.; Marshall, D.C.; Patel, S.; et al. Group C Niemann-Pick disease: Faulty regulation of low-density lipoprotein uptake and cholesterol storage in cultured fibroblasts. FASEB J. 1987, 1, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Vanier, M.T. Niemann-Pick disease type C. Orphanet J. Rare Dis. 2010, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebner, L.; Gläser, A.; Bräuer, A.; Witt, M.; Wree, A.; Rolfs, A.; Frank, M.; Vollmar, B.; Kuhla, A. Evaluation of Two Liver Treatment Strategies in a Mouse Model of Niemann-Pick-Disease Type C1. Int. J. Mol. Sci. 2018, 19, 972. [Google Scholar] [CrossRef] [PubMed]
- Garver, W.S.; Francis, G.A.; Jelinek, D.; Shepherd, G.; Flynn, J.; Castro, G.; Walsh Vockley, C.; Coppock, D.L.; Pettit, K.M.; Heidenreich, R.A.; et al. The National Niemann-Pick C1 disease database: Report of clinical features and health problems. Am. J. Med. Genet. A 2007, 143, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Beltroy, E.P.; Richardson, J.A.; Horton, J.D.; Turley, S.D.; Dietschy, J.M. Cholesterol accumulation and liver cell death in mice with Niemann-Pick type C disease. Hepatology 2005, 42, 886–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Yamada, Y.; Ishitsuka, Y.; Matsuo, M.; Shiraishi, K.; Wada, K.; Uchio, Y.; Kondo, Y.; Takeo, T.; Nakagata, N.; et al. Efficacy of 2-Hydroxypropyl-beta-cyclodextrin in Niemann-Pick Disease Type C Model Mice and Its Pharmacokinetic Analysis in a Patient with the Disease. Biol. Pharm. Bull. 2015, 38, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Pentchev, P.G.; Gal, A.E.; Booth, A.D.; Omodeo-Sale, F.; Fouks, J.; Neumeyer, B.A.; Quirk, J.M.; Dawson, G.; Brady, R.O. A lysosomal storage disorder in mice characterized by a dual deficiency of sphingomyelinase and glucocerebrosidase. Biochim. Biophys. Acta 1980, 619, 669–679. [Google Scholar] [CrossRef]
- Imrie, J.; Dasgupta, S.; Besley, G.T.; Harris, C.; Heptinstall, L.; Knight, S.; Vanier, M.T.; Fensom, A.H.; Ward, C.; Jacklin, E.; et al. The natural history of Niemann-Pick disease type C in the UK. J. Inherit. Metab. Dis. 2007, 30, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Sevin, M.; Lesca, G.; Baumann, N.; Millat, G.; Lyon-Caen, O.; Vanier, M.T.; Sedel, F. The adult form of Niemann-Pick disease type C. Brain 2007, 130, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Patterson, M.C.; Hendriksz, C.J.; Walterfang, M.; Sedel, F.; Vanier, M.T.; Wijburg, F.; Group, N.-C.G.W. Recommendations for the diagnosis and management of Niemann-Pick disease type C: An update. Mol. Genet. Metab. 2012, 106, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Vanier, M.T. Complex lipid trafficking in Niemann-Pick disease type C. J. Inherit. Metab. Dis. 2015, 38, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Loftus, S.K.; Morris, J.A.; Carstea, E.D.; Gu, J.Z.; Cummings, C.; Brown, A.; Ellison, J.; Ohno, K.; Rosenfeld, M.A.; Tagle, D.A.; et al. Murine model of Niemann-Pick C disease: Mutation in a cholesterol homeostasis gene. Science 1997, 277, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Claudepierre, T.; Paques, M.; Simonutti, M.; Buard, I.; Sahel, J.; Maue, R.A.; Picaud, S.; Pfrieger, F.W. Lack of Niemann-Pick type C1 induces age-related degeneration in the mouse retina. Mol. Cell Neurosci. 2009. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Ma, L.; Hovakimyan, M.; Lukas, J.; Wree, A.; Frank, M.; Guthoff, R.; Rolfs, A.; Witt, M.; Luo, J. Defects in the retina of Niemann-pick type C 1 mutant mice. BMC Neurosci. 2014, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Aisen, M.; Rapoport, S.; Solomon, G. Brain stem auditory evoked potentials in two siblings with Niemann-Pick disease. Brain Dev. 1985, 7, 431–433. [Google Scholar] [CrossRef]
- King, K.A.; Gordon-Salant, S.; Yanjanin, N.; Zalewski, C.; Houser, A.; Porter, F.D.; Brewer, C.C. Auditory phenotype of Niemann-Pick disease, type C1. Ear Hear. 2014, 35, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Aki, T.; Funakoshi, T.; Unuma, K.; Uemura, K. Impairment of autophagy: From hereditary disorder to drug intoxication. Toxicology 2013, 311, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Schwob, J.E. Neural regeneration and the peripheral olfactory system. Anat. Rec. 2002, 269, 33–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwob, J.E.; Jang, W.; Holbrook, E.H.; Lin, B.; Herrick, D.B.; Peterson, J.N.; Hewitt Coleman, J. Stem and progenitor cells of the mammalian olfactory epithelium: Taking poietic license. J. Comp. Neurol. 2017, 525, 1034–1054. [Google Scholar] [CrossRef] [PubMed]
- Haehner, A.; Hummel, T.; Hummel, C.; Sommer, U.; Junghanns, S.; Reichmann, H. Olfactory loss may be a first sign of idiopathic Parkinson’s disease. Mov. Disord. 2007, 22, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfactory dysfunction in Parkinson disease. Nat. Rev. Neurol. 2012, 8, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Deems, D.A.; Stellar, S. Olfactory dysfunction in parkinsonism: A general deficit unrelated to neurologic signs, disease stage, or disease duration. Neurology 1988, 38, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Hovakimyan, M.; Meyer, A.; Lukas, J.; Luo, J.; Gudziol, V.; Hummel, T.; Rolfs, A.; Wree, A.; Witt, M. Olfactory deficits in Niemann-Pick type C1 (NPC1) disease. PLoS ONE 2013, 8, e82216. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Gläser, A.; Bräuer, A.U.; Wree, A.; Strotmann, J.; Rolfs, A.; Witt, M. Olfactory Performance as an Indicator for Protective Treatment Effects in an Animal Model of Neurodegeneration. Front. Integr. Neurosci. 2018, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Wree, A.; Günther, R.; Holzmann, C.; Schmitt, O.; Rolfs, A.; Witt, M. Increased Regenerative Capacity of the Olfactory Epithelium in Niemann-Pick Disease Type C1. Int. J. Mol. Sci. 2017, 18, 777. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, R.H.; Platt, F.M. Substrate reduction therapy for glycosphingolipid storage disorders. Expert Opin. Investig. Drugs 2001, 10, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Platt, F.M.; Jeyakumar, M. Substrate reduction therapy. Acta Paediatr. Suppl. 2008, 97, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.D.; Gong, W.; Verot, L.; Mellon, S.H. Niemann-Pick type C disease involves disrupted neurosteroidogenesis and responds to allopregnanolone. Nat. Med. 2004, 10, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Davidson, C.D.; Ali, N.F.; Micsenyi, M.C.; Stephney, G.; Renault, S.; Dobrenis, K.; Ory, D.S.; Vanier, M.T.; Walkley, S.U. Chronic cyclodextrin treatment of murine Niemann-Pick C disease ameliorates neuronal cholesterol and glycosphingolipid storage and disease progression. PLoS ONE 2009, 4, e6951. [Google Scholar] [CrossRef] [PubMed]
- Hovakimyan, M.; Maass, F.; Petersen, J.; Holzmann, C.; Witt, M.; Lukas, J.; Frech, M.J.; Hubner, R.; Rolfs, A.; Wree, A. Combined therapy with cyclodextrin/allopregnanolone and miglustat improves motor but not cognitive functions in Niemann-Pick Type C1 mice. Neuroscience 2013, 252, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E.; Karten, B. Niemann-Pick C disease and mobilization of lysosomal cholesterol by cyclodextrin. J. Lipid Res. 2014, 55, 1609–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, E.; McCulloch, M.A.; Farbman, A.I. Proliferation in the vomeronasal organ of the rat during postnatal development. Eur. J. Neurosci. 1999, 11, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Marcos, A.; Jia, C.; Quan, W.; Halpern, M. Neurogenesis, migration, and apoptosis in the vomeronasal epithelium of adult mice. J. Neurobiol. 2005, 63, 173–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Rosa-Prieto, C.; Saiz-Sanchez, D.; Ubeda-Banon, I.; Argandona-Palacios, L.; Garcia-Munozguren, S.; Martinez-Marcos, A. Fate of marginal neuroblasts in the vomeronasal epithelium of adult mice. J. Comp. Neurol. 2009, 517, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Baldisseri, D.M.; Margolis, J.W.; Weber, D.J.; Koo, J.H.; Margolis, F.L. Olfactory marker protein (OMP) exhibits a beta-clam fold in solution: Implications for target peptide interaction and olfactory signal transduction. J. Mol. Biol. 2002, 319, 823–837. [Google Scholar] [CrossRef]
- Margolis, F.L. Olfactory marker protein (OMP). Scand. J. Immunol. Suppl. 1982, 9, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Dibattista, M.; Reisert, J. The Odorant Receptor-Dependent Role of Olfactory Marker Protein in Olfactory Receptor Neurons. J. Neurosci. 2016, 36, 2995–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witt, M.; Hummel, T. Vomeronasal versus olfactory epithelium: Is there a cellular basis for human vomeronasal perception? Int. Rev. Cytol. 2006, 248, 209–259. [Google Scholar] [PubMed]
- Johnson, E.W.; Eller, P.M.; Jafek, B.W. An immuno-electron microscopic comparison of olfactory marker protein localization in the supranuclear regions of the rat olfactory epithelium and vomeronasal organ neuroepithelium. Acta Otolaryngol. 1993, 113, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Rodewald, A.; Gisder, D.; Gebhart, V.M.; Oehring, H.; Jirikowski, G.F. Distribution of olfactory marker protein in the rat vomeronasal organ. J. Chem. Neuroanat. 2016, 77, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Kaluza, J.F.; Gussing, F.; Bohm, S.; Breer, H.; Strotmann, J. Olfactory receptors in the mouse septal organ. J. Neurosci. Res. 2004, 76, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Weiler, E.; Farbman, A.I. The Septal Organ of the Rat During Postnatal Development. Chem. Sens. 2003, 28, 581–593. [Google Scholar] [CrossRef] [Green Version]
- Brechbühl, J.; Klaey, M.; Broillet, M.C. Grueneberg ganglion cells mediate alarm pheromone detection in mice. Science 2008, 321, 1092–1095. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, J.; Breer, H. The Grueneberg ganglion: A novel sensory system in the nose. Histol. Histopathol. 2010, 25, 909–915. [Google Scholar] [PubMed]
- Dulac, C.; Axel, R. A novel family of genes encoding putative pheromone receptors in mammals. Cell 1995, 83, 195–206. [Google Scholar] [CrossRef]
- Meredith, M.; Fernandez-Fewell, G. Vomeronasal system, LHRH, and sex behaviour. Psychoneuroendocrinology 1994, 19, 657–672. [Google Scholar] [CrossRef]
- Meredith, M. Sensory processing in the main and accessory olfactory systems: Comparisons and contrasts. J. Steroid Biochem. Mol. Biol. 1991, 39, 601–614. [Google Scholar] [CrossRef]
- Kimoto, H.; Haga, S.; Sato, K.; Touhara, K. Sex-specific peptides from exocrine glands stimulate mouse vomeronasal sensory neurons. Nature 2005, 437, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Halpern, M. Subclasses of vomeronasal receptor neurons: Differential expression of G proteins (Gi alpha 2 and G(o alpha)) and segregated projections to the accessory olfactory bulb. Brain Res. 1996, 719, 117–128. [Google Scholar] [CrossRef]
- Seo, Y.; Kim, H.-S.; Shin, Y.; Kang, I.; Choi, S.W.; Yu, K.-R.; Seo, K.-W.; Kang, K.-S. Excessive microglial activation aggravates olfactory dysfunction by impeding the survival of newborn neurons in the olfactory bulb of Niemann–Pick disease type C1 mice. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 2193–2203. [Google Scholar] [CrossRef] [PubMed]
- Amritraj, A.; Wang, Y.; Revett, T.J.; Vergote, D.; Westaway, D.; Kar, S. Role of Cathepsin D in U18666A-induced Neuronal Cell Death: Potential implication in Niemann-Pick Type C disease pathogenesis. J. Biol. Chem. 2012, 288, 3136–3152. [Google Scholar] [CrossRef] [PubMed]
- Cluzeau, C.V.; Watkins-Chow, D.E.; Fu, R.; Borate, B.; Yanjanin, N.; Dail, M.K.; Davidson, C.D.; Walkley, S.U.; Ory, D.S.; Wassif, C.A.; et al. Microarray expression analysis and identification of serum biomarkers for Niemann-Pick disease, type C1. Hum. Mol. Genet. 2012, 21, 3632–3646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cataldo, A.M.; Paskevich, P.A.; Kominami, E.; Nixon, R.A. Lysosomal hydrolases of different classes are abnormally distributed in brains of patients with Alzheimer disease. Proc. Natl. Acad. Sci. USA 1991, 88, 10998–11002. [Google Scholar] [CrossRef] [PubMed]
- Vitner, E.B.; Dekel, H.; Zigdon, H.; Shachar, T.; Farfel-Becker, T.; Eilam, R.; Karlsson, S.; Futerman, A.H. Altered expression and distribution of cathepsins in neuronopathic forms of Gaucher disease and in other sphingolipidoses. Hum. Mol. Genet. 2010, 19, 3583–3590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, F.; Petersen, J.; Hovakimyan, M.; Schmitt, O.; Witt, M.; Hawlitschka, A.; Lukas, J.; Rolfs, A.; Wree, A. Reduced cerebellar neurodegeneration after combined therapy with cyclodextrin/allopregnanolone and miglustat in NPC1: A mouse model of Niemann-Pick type C1 disease. J. Neurosci. Res. 2015, 93, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Turley, S.D.; Burns, D.K.; Miller, A.M.; Repa, J.J.; Dietschy, J.M. Reversal of defective lysosomal transport in NPC disease ameliorates liver dysfunction and neurodegeneration in the npc1-/- mouse. Proc. Natl. Acad. Sci. USA 2009, 106, 2377–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqul, A.; Liu, B.; Ramirez, C.M.; Pieper, A.A.; Estill, S.J.; Burns, D.K.; Liu, B.; Repa, J.J.; Turley, S.D.; Dietschy, J.M. Unesterified cholesterol accumulation in late endosomes/lysosomes causes neurodegeneration and is prevented by driving cholesterol export from this compartment. J. Neurosci. 2011, 31, 9404–9413. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.M.; Liu, B.; Aqul, A.; Taylor, A.M.; Repa, J.J.; Turley, S.D.; Dietschy, J.M. Quantitative role of LAL, NPC2, and NPC1 in lysosomal cholesterol processing defined by genetic and pharmacological manipulations. J. Lipid Res. 2011, 52, 688–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megias-Vericat, J.E.; Garcia-Robles, A.; Company-Albir, M.J.; Fernandez-Megia, M.J.; Perez-Miralles, F.C.; Lopez-Briz, E.; Casanova, B.; Poveda, J.L. Early experience with compassionate use of 2 hydroxypropyl-beta-cyclodextrin for Niemann-Pick type C disease: Review of initial published cases. Neurol. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Crumling, M.A.; Liu, L.; Thomas, P.V.; Benson, J.; Kanicki, A.; Kabara, L.; Halsey, K.; Dolan, D.; Duncan, R.K. Hearing loss and hair cell death in mice given the cholesterol-chelating agent hydroxypropyl-beta-cyclodextrin. PLoS ONE 2012, 7, e53280. [Google Scholar] [CrossRef] [PubMed]
- Davidson, C.D.; Fishman, Y.I.; Puskas, I.; Szeman, J.; Sohajda, T.; McCauliff, L.A.; Sikora, J.; Storch, J.; Vanier, M.T.; Szente, L.; et al. Efficacy and ototoxicity of different cyclodextrins in Niemann-Pick C disease. Ann. Clin. Transl. Neurol. 2016, 3, 366–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, S.; O’Donnell, P.; Fernandez, S.; Vite, C.H. 2-hydroxypropyl-beta-cyclodextrin raises hearing threshold in normal cats and in cats with Niemann-Pick type C disease. Pediatr. Res. 2010, 68, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Dover, R.; Patel, K. Improved methodology for detecting bromodeoxyuridine in cultured cells and tissue sections by immunocytochemistry. Histochemistry 1994, 102, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Onda, K.; Davis, R.L.; Shibuya, M.; Wilson, C.B.; Hoshino, T. Correlation between the bromodeoxyuridine labeling index and the MIB-1 and Ki-67 proliferating cell indices in cerebral gliomas. Cancer 1994, 74, 1921–1926. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, H.J. Stereology of arbitrary particles. A review of unbiased number and size estimators and the presentation of some new ones, in memory of William R. Thompson. J. Microsc. 1986, 143, 3–45. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Group | VNE Cells/mm³ ± SEM | OE * Cells/mm³ ± SEM | ||
|---|---|---|---|---|
| sham-treated Npc1+/+ | 37,827 ± 3,907 | 17,417 ± 1,317 | ||
| sham-treated Npc1−/− | 81,287 ± 9,900 | +215% | 25,180 ± 1,605 | +144% |
| HPβCD-treated Npc1−/− | 98,626 ± 10,388 | +260% | 44,693 ± 4,191 | +257% |
| Combination-treated Npc1−/− | 102,080 ± 10.097 | +269% | 70,558 ± 8159 | +405% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witt, M.; Thiemer, R.; Meyer, A.; Schmitt, O.; Wree, A. Main Olfactory and Vomeronasal Epithelium Are Differently Affected in Niemann-Pick Disease Type C1. Int. J. Mol. Sci. 2018, 19, 3563. https://doi.org/10.3390/ijms19113563
Witt M, Thiemer R, Meyer A, Schmitt O, Wree A. Main Olfactory and Vomeronasal Epithelium Are Differently Affected in Niemann-Pick Disease Type C1. International Journal of Molecular Sciences. 2018; 19(11):3563. https://doi.org/10.3390/ijms19113563
Chicago/Turabian StyleWitt, Martin, René Thiemer, Anja Meyer, Oliver Schmitt, and Andreas Wree. 2018. "Main Olfactory and Vomeronasal Epithelium Are Differently Affected in Niemann-Pick Disease Type C1" International Journal of Molecular Sciences 19, no. 11: 3563. https://doi.org/10.3390/ijms19113563