Tolerance to Drought, Low pH and Al Combined Stress in Tibetan Wild Barley Is Associated with Improvement of ATPase and Modulation of Antioxidant Defense System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Plant Height, Biomass Accumulation, and Al Concentration
2.2. Photosynthetic Parameters
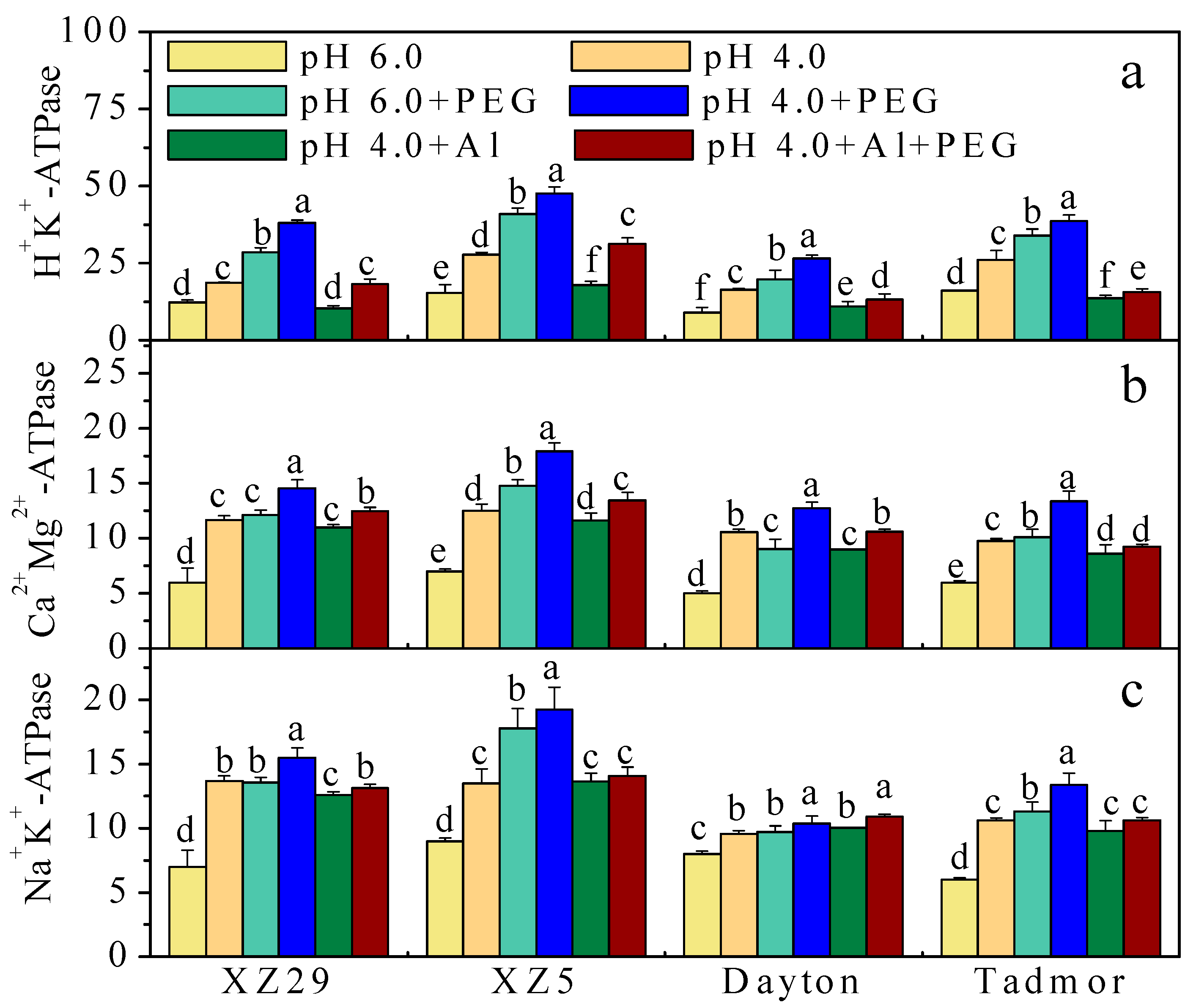
2.3. Activity of H+-ATPase, Ca2+Mg2+-ATPase, and Na+K+-ATPase in Barley Leaves
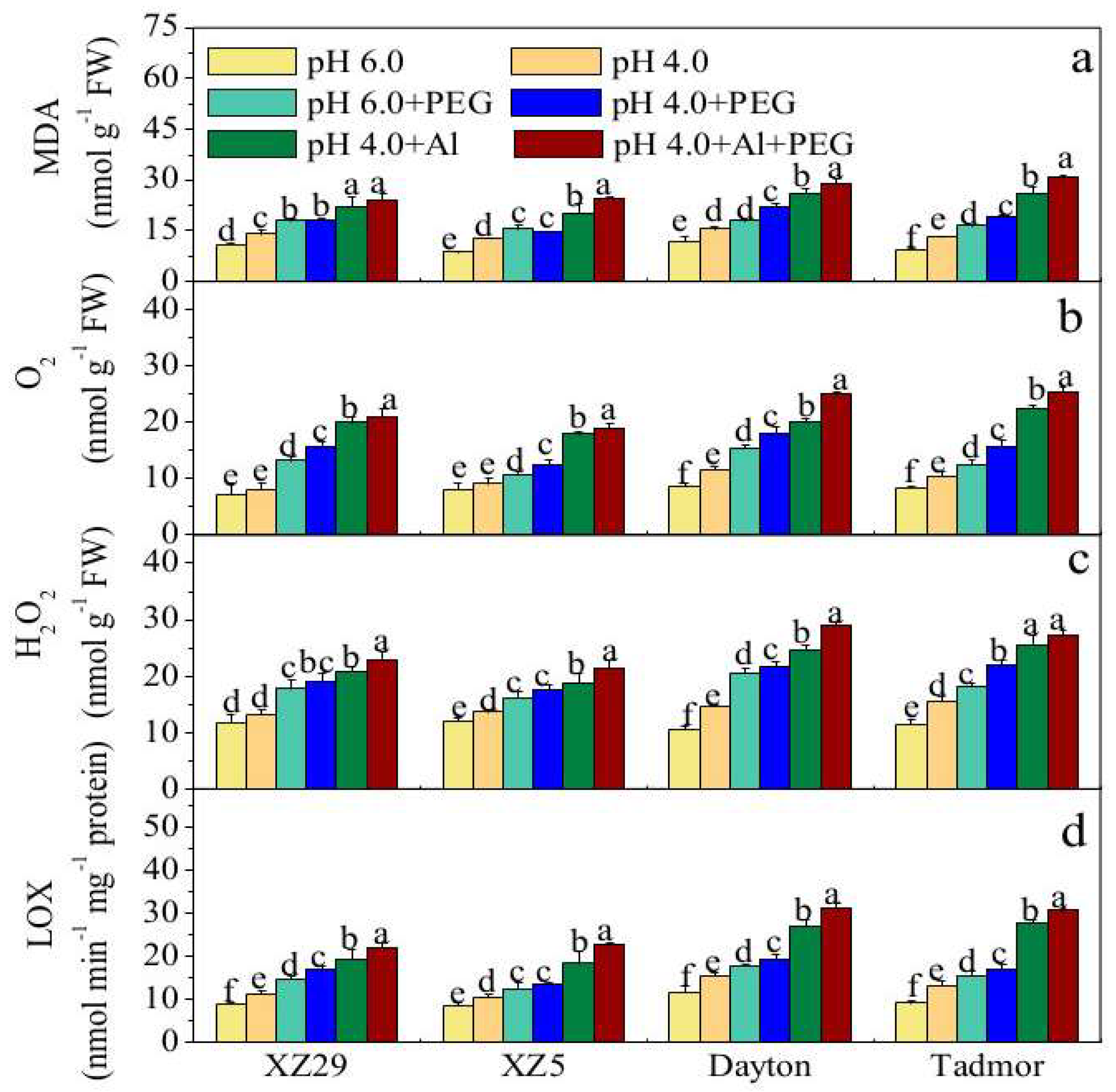
2.4. H2O2, O2−, and Malondialdehyde (MDA) Contents and Lipoxygenase (LOX) Activity
2.5. Antioxidant Enzyme Activities
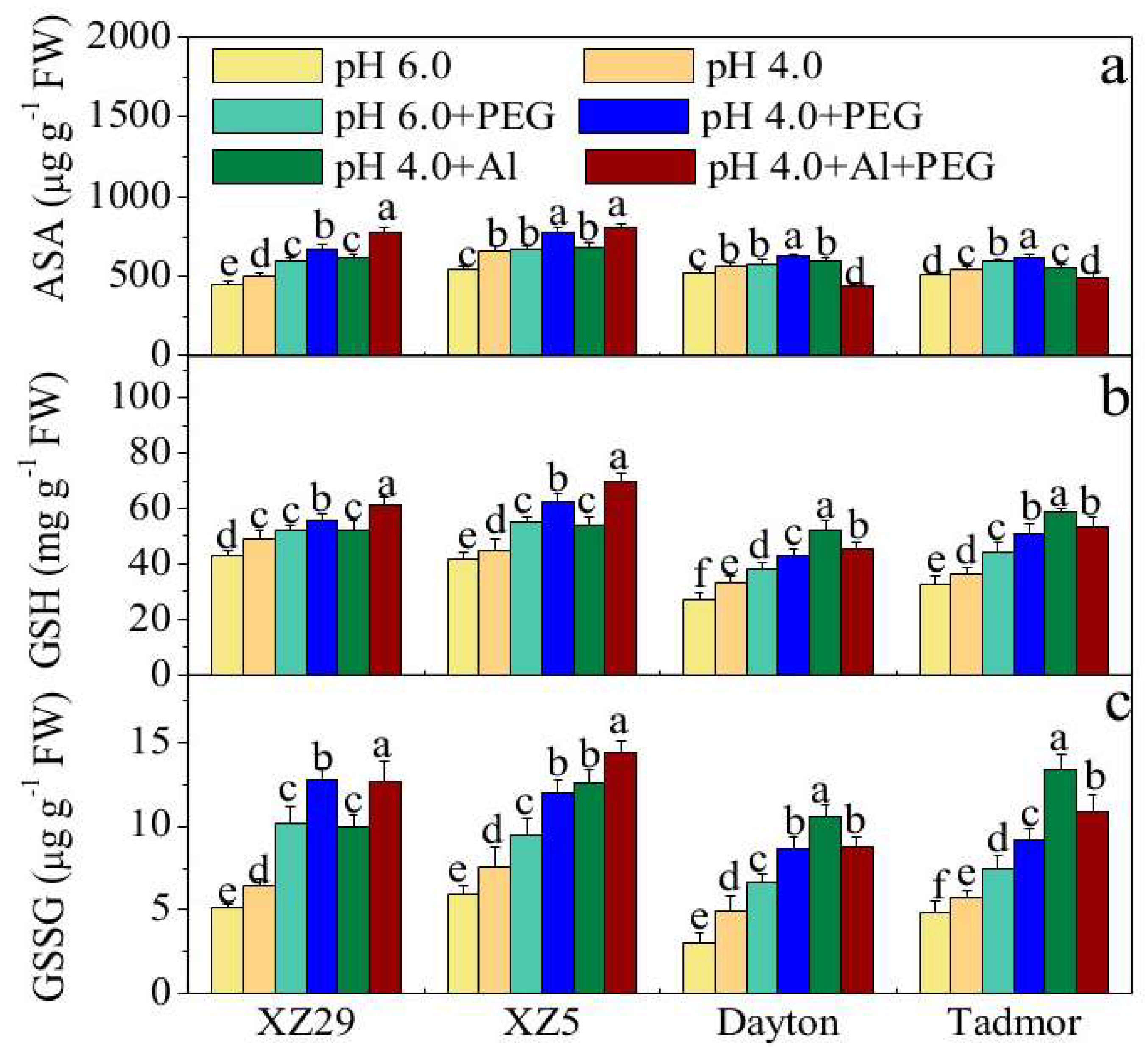
2.6. Contents of Non-Enzymatic Antioxidants
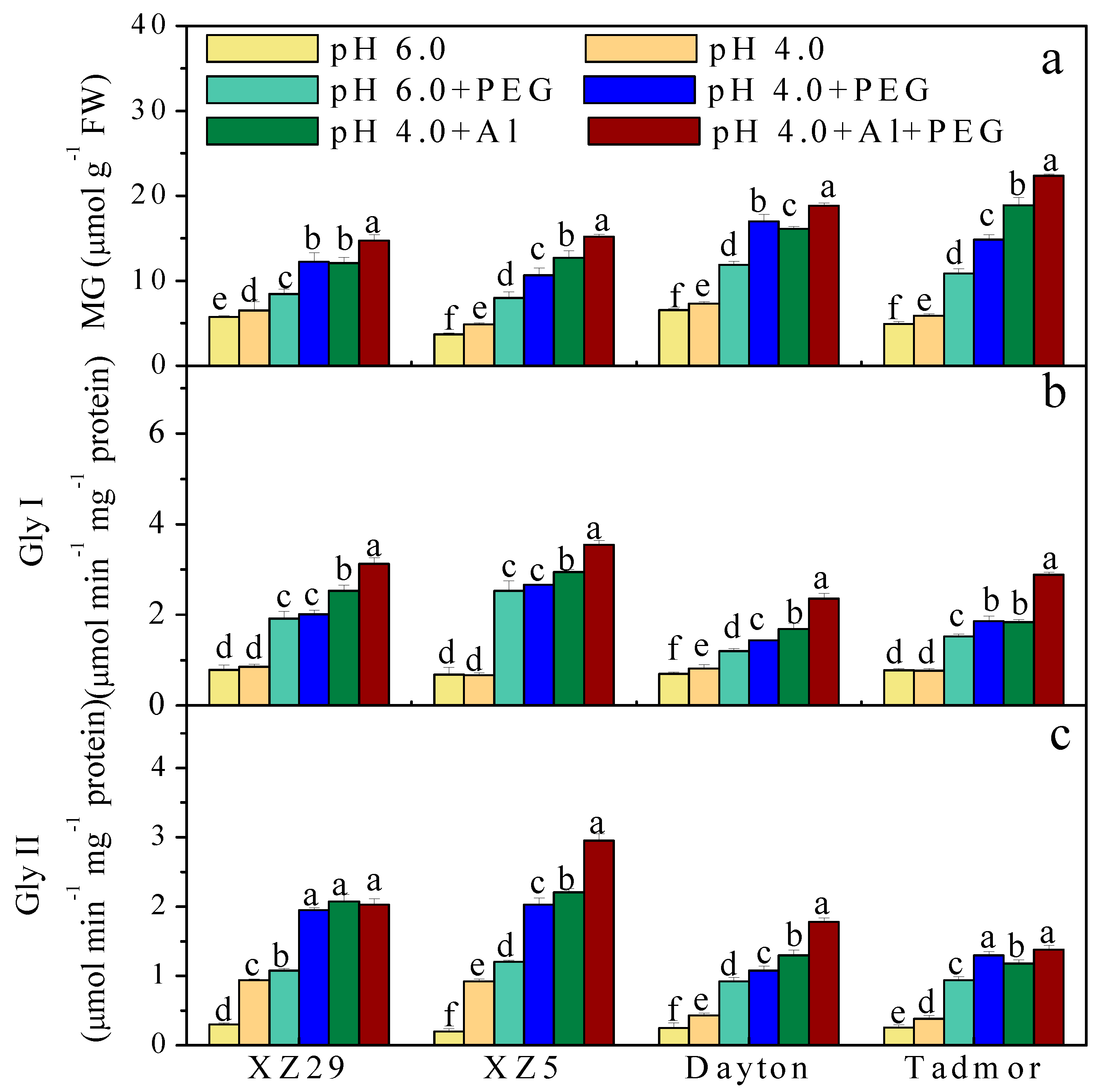
2.7. Methylglyoxal (MG) Content and Activity of Glyoxalase (Gly) I and Gly II Enzymes
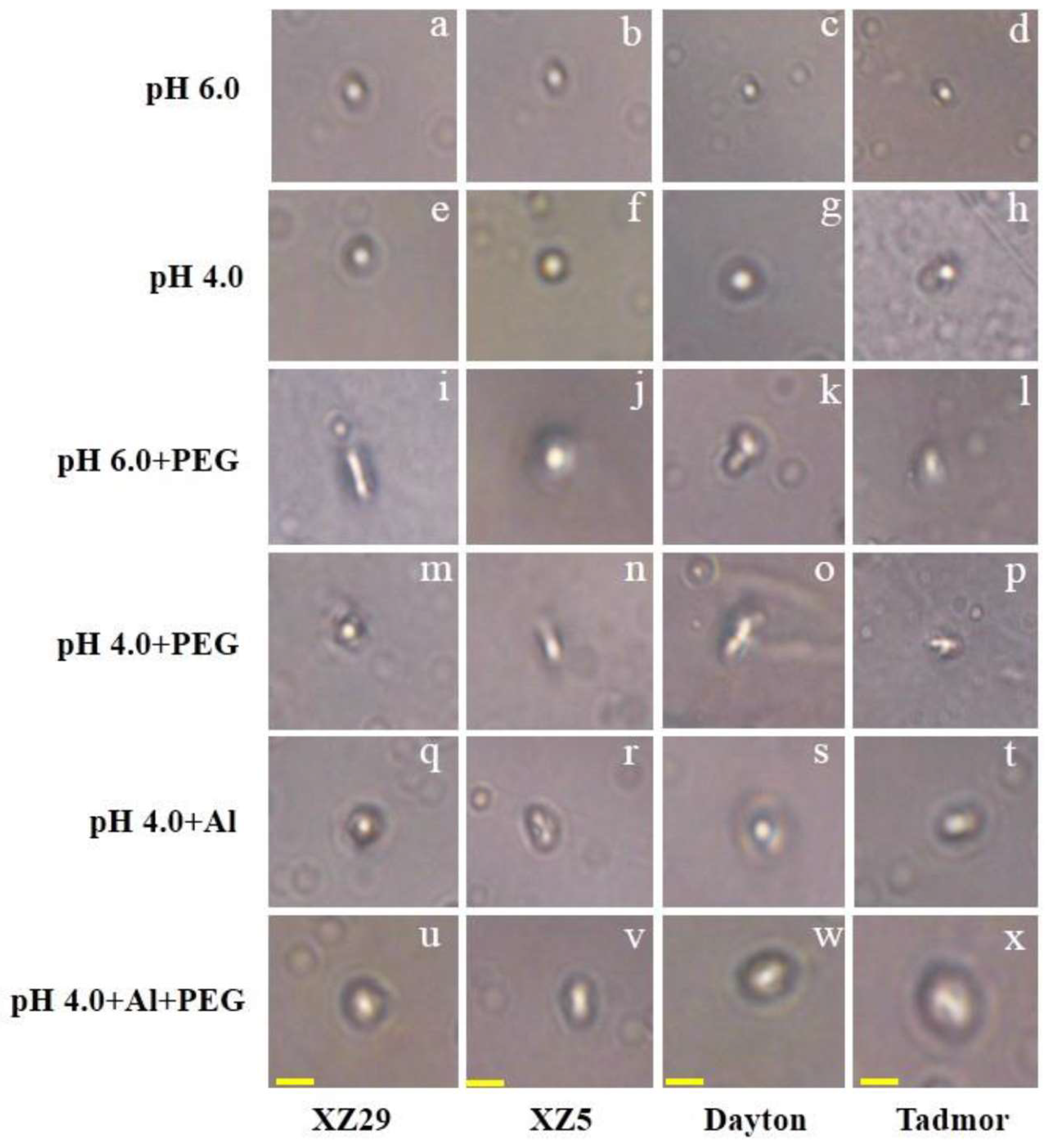
2.8. DNA Damage Induced by Drought and Al Stress
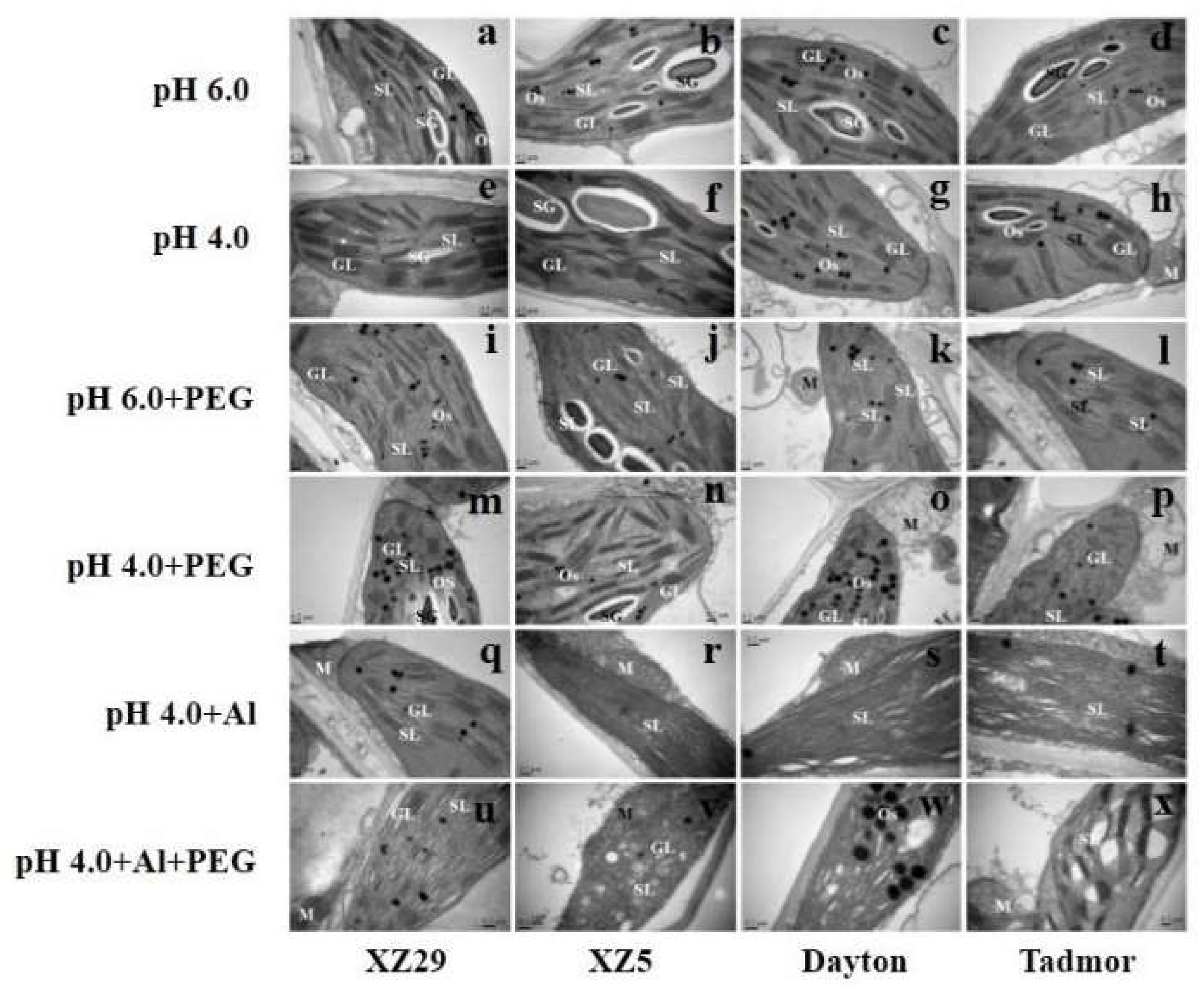
2.9. Ultrastructural Changes in Chloroplast
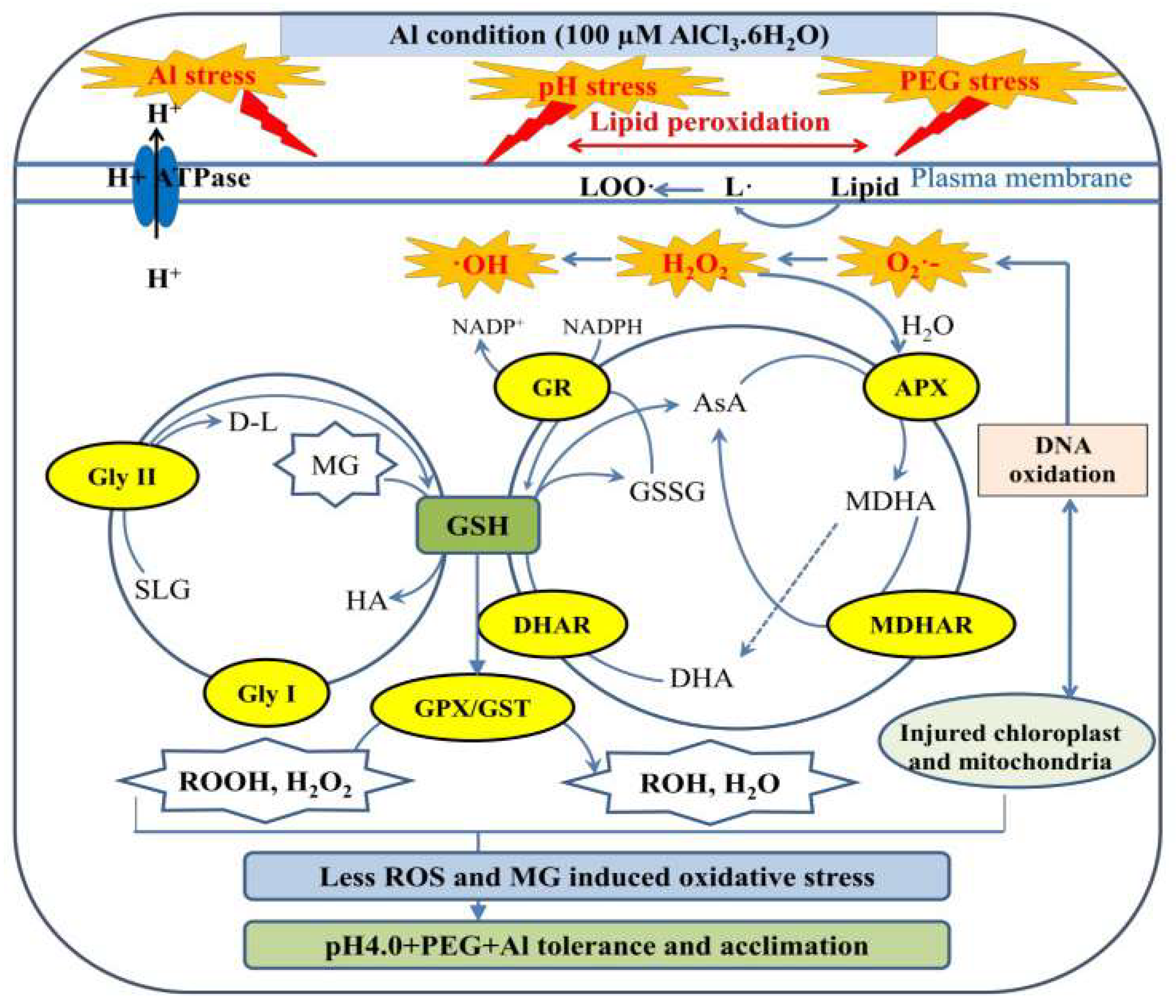
3. Discussion
3.1. The Reduction of Gas Exchange Associated with Combined Stress
3.2. ATPase Activity is Critical for Drought and Al Tolerance
3.3. Tibetan Wild Barley Exhibited Higher ROS-Detoxifying Capacity under Combined Stress Conditions
3.4. XZ29 and XZ5 Maintained a Higher Increase in Activities of Gly I and Gly II with Lower MG Toxicity and DNA Damage under Stress
3.5. Tibetan Wild Barley Showed Fewer Changes in Chloroplasts Organelles under Combined Stress
4. Materials and Methods
4.1. Plant Materials and Hydroponic Culture
4.2. Measurement of Plant Height, Dry Weight, and Al Concentration
4.3. Determination of Photosynthetic Parameters
4.4. Determination of ATPase and Non-Enzymatic Antioxidants
4.5. Determination of Lipid Peroxidation, Hydrogen Peroxide (H2O2), and the Superoxide Radical (O2−)
4.6. Extraction and Assay of Enzymes and Determination of Methylglyoxal (MG) Content
4.7. Single Cell Gel Electrophoresis Assay (Comet Assay)
4.8. Investigation of Leaf Ultrastructure
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Z.C.; Liao, H. Organic acid anions: an effective defensive weapon for plants against aluminum toxicity and phosphorus deficiency in acidic soils. J. Genet. Genom. 2016, 43, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, H.; Xu, J.; Chun, X.; Ni, D. Effects of aluminum on ultrastructure and antioxidant activity in leaves of tea plant. Acta Physiol. Plant. 2011, 33, 973–978. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: from genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.S.; Angle, J.S.; Chaney, R.L.; Thierry, A.; Delorme, T.R.; Reeves, R.D. Soil pH effects on uptake of Cd and Zn by Thlaspi caerulescens. Plant Soil 2006, 281, 325–337. [Google Scholar] [CrossRef]
- Eekhout, T.; Larsen, P.; De Veylder, L. Modification of DNA checkpoints to confer aluminum tolerance. Trends Plant Sci. 2017, 22, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Inostroza-Blancheteau, C.; Reyes-Díaz, M.; Aquea, F.; Nunes-Nesi, A.; Alberdi, M.; Arce-Johnson, P. Biochemical and molecular changes in response to aluminum stress in high bush blueberry (Vaccinium corymbosum L.). Plant Physiol. Biochem. 2011, 49, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Konarska, A. Effects of aluminum on growth and structure of red pepper (Capsicum annuum L.) leaves. Acta Physiol. Plant. 2010, 32, 145–151. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Vardar, F.; Yasar, U.; Akinci, S. Long-term effects of aluminum and cadmium on growth leaf anatomy and photosynthetic pigments of cotton. Commun. Soil Sci. Plant Anal. 2013, 44, 3076–3091. [Google Scholar] [CrossRef]
- Moustakas, M.; Ouzounidou, G.; Eleftheriou, E.P.; Lannoye, R. Indirect effects of aluminum stress on the function of the photosynthetic apparatus. Plant Physiol. Biochem. 1996, 34, 553–560. [Google Scholar]
- Mukhopadyay, M.; Bantawa, P.; Das, A.; Sarkar, B.; Bera, B.; Ghosh, P.; Mondal, T.K. Changes of growth photosynthesis and alteration of leaf antioxidative defense system of tea [Camellia sinensis (L.) O. Kuntze] seedlings under aluminum stress. BioMetals 2012, 25, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.R.; Tyerman, S.D.; Sasaki, T.; Furuichi, T.; Yamamoto, Y.; Zhang, W.H.; Delhaize, E. The identification of aluminum-resistance genes provides opportunities for enhancing crop production on acid soils. J. Exp. Bot. 2011, 62, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, X.; Yao, S.; LwinOo, T.; He, H.; Wang, A.; Li, C.; He, L. Reactive oxygen species burst induced by aluminum stress triggers mitochondria-dependent programmed cell death in peanut root tip cells. Plant Physiol. Biochem. 2014, 82, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Brito, DS.; Inostroza-Blancheteau, C.; Fernie, A.R.; Araújo, W.L. The complex role of mitochondrial metabolism in plant aluminum resistance. Trends in Plant Sci. 2014, 19, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Kichigina, N.E.; Puhalsky, J.V.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Loskutov, S.I.; Safronova, V.I.; Tikhonovich, I.A.; Vishnyakova, M.A.; Semenova, E.V.; et al. Aluminum exclusion from root zone and maintenance of nutrient uptake are principal mechanisms of Al tolerance in Pisum sativum L. Physiol. Mol. Biol. Plants 2017, 23, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Xia, S.; Ye, J.; Huang, Y.; Liu, C.; Zhang, Z. PEG-simulated drought stress decreases cadmium accumulation in castor bean by altering root morphology. Environ. Exp. Bot. 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Goldman, I.L.; Carter, T.E.; Patterson, R.P. A detrimental interaction of subsoil aluminum and drought stress on the leaf water status of soybean. Agron. J. 1989, 81, 461–463. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Forster, B.P.; Ellis, R.P.; Thomas, W.; Newton, A.C.; Tuberosa, R.; This, D.; El-Enein, R.; Bahri, M.; Salem, M.B. The development and application of molecular markers for abiotic stress tolerance in barley. J. Exp. Bot. 2000, 51, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.Q.; Ma, J.F.; Sato, K.; Takeda, K. Differential Al resistance and citrate secretion in barley (Hordeum vulgare L.). Planta 2003, 217, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Pickering, R.; Johnston, P.A. Recent progress in barley improvement using wild species of Hordeum. Cytogenet. Genome Res. 2005, 109, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Nevo, E.; Wu, D.Z.; Comadran, J.; Zhou, M.; Qiu, L.; Chen, Z.H.; Beiles, A.; Chen, G.; Zhang, G.P. Tibet is one of the centers of domestication of cultivated barley. Proc. Natl. Acad. Sci. USA 2012, 109, 16969–16973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, I.M.; Nadira, U.A.; Cao, F.B.; He, X.Y.; Zhang, G.P.; Wu, F.B. Physiological and molecular analysis on root growth associated with the tolerance to aluminum and drought individual and combined in Tibetan wild and cultivated barley. Planta 2016, 243, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, S.; Zhao, H.; Korpelainen, H.; Li, C. Sex-related adaptive responses to interaction of drought and salinity in Populus yunnanensis. Plant Cell Environ. 2010, 33, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Zeng, F.R.; Qiu, B.Y.; Zhang, G.P. The effect of chromium and aluminum on growth, root morphology, photosynthetic parameters and transpiration of the two barley cultivars. Biologia. Planta 2011, 55, 291–296. [Google Scholar] [CrossRef]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J.Plant Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G.; Harper, J.F. Pumping with plant P-type ATPases. J. Exp. Bot. 1999, 50, 883–893. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Cao, F.B.; Zhang, M.; Chen, X.; Zhang, G.P.; Wu, F.B. Difference in yield and physiological features in response to drought and salinity combined stress during anthesis in Tibetan wild and cultivated barleys. PLoS ONE 2013, 8, e77869. [Google Scholar] [CrossRef] [PubMed]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Szalai, G.; Kellõs, T.; Galiba, G.; Kocsy, G. Glutathione as an Antioxidant and Regulatory Molecule in Plants Under Abiotic Stress Conditions. J. Plant Growth Regul. 2009, 28, 66–80. [Google Scholar] [CrossRef]
- Tan, S.; Zhu, M.; Zhang, Q. Physiological responses of bermudagrass (Cynodon dactylon) to submergence. Acta Physiol. Plant. 2010, 32, 133–140. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hossain, M.Z.; Fujita, M. Stress induced changes of methylglyoxal level and glyoxalase I activity in pumpkin seedlings and cDNA cloning of glyoxalase I gene. Aust. J. Crop Sci. 2009, 3, 53–64. [Google Scholar]
- Kaur, C.; Ghosh, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Glyoxalases and stress tolerance in plants. Biochem. Soc. Trans. 2014, 42, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, C.P.; Venkatesh, J.; Gururani, M.A.; Asnin, L.; Sharma, K.; Ajappala, H.; Park, S.W. Transgenic potato overproducing L-ascorbic acid resisted an increase in methylglyoxal under salinity stress via maintaining higher reduced glutathione level and glyoxalase enzyme activity. Biotechnol. Lett. 2011, 33, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Álvarez Viveros, M.F.; Inostroza-Blancheteau, C.; Timmermann, T.; González, M.; Arce-Johnson, P. Overexpression of Gly I and Gly II genes in transgenic tomato (Solanum lycopersicum Mill.) plants confers salt tolerance by decreasing oxidative stress. Mol. Biol. Rep. 2013, 40, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Östling, O.; Johanson, K.J. Microelectrophoretic study of radiation-induced DNA damages in individual mammalian cells. Biochem. Biophys. Res. Commun. 1984, 123, 291–298. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Bibi, N.; Cao, F.B.; He, X.Y.; Zhang, G.P.; Wu, F.B. Secondary metabolism and antioxidants are involved in the tolerance to drought and salinity separately and combined in Tibetan wild barley. Environ. Exp. Bot. 2015, 111, 1–12. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, Y.; Ding, Z.; Fan, K.; Ma, D.; Zhang, Y.; Yin, Q. Aluminum induced physiological and proteomic responses in tea (Camellia sinensis) roots and leaves. Plant Physiol. Biochem. 2017, 115, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Pääkkönen, E.; Vahala, J.; Pohjola, M.; Holopainen, T.; Kärenlampi, L. Physiological stomatal and ultrastructural ozone responses in birch (Betula pendula Roth) are modified by water stress. Plant Cell Environ. 1998, 21, 671–684. [Google Scholar] [CrossRef]
- Rauser, W.E.; Samarakoon, A.B. Vein loading in seedlings of Phaseolus vulgaris exposed to excess cobalt nickel and zinc. Plant Physiol. 1980, 65, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hua, X.; Guo, J.; Qi, D.; Wang, L.; Liu, Z.; Jin, Z.; Chen, S.; Liu, G. Enhanced tolerance to drought stress in transgenic tobacco plants overexpressing VTE1 for increased tocopherol production from Arabidopsis thaliana. Biotechnol. Lett. 2008, 30, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Dutilleul, C.; Driscoll, S.; Cornic, G.; De Paepe, R.; Foyer, C.H.; Noctor, G. Functional mitochondrial complex I is required by tobacco leaves for optimal photosynthetic performance in photo respiratory conditions and during transients. Plant Physiol. 2003, 131, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Dai, H.; Zheng, W.; Cao, F.B.; Zhang, G.P.; Sun, D.; Wu, F.B. Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiol. Biochem. 2013, 63, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Fujita, M.; Tran, L.S.P. Nitric oxide mediates hydrogen peroxide- and salicylic acid-induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, F.B.; Wei, K.; Zhang, G.P.; Wu, F.B. Genotypic dependent effect of exogenous glutathione on Cd-induced changes in proteins ultrastructure and antioxidant defense enzymes in rice seedlings. J. Hazard. Mater. 2011, 192, 1056–1066. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, I.M.; Nadira, U.A.; Qiu, C.-W.; Cao, F.; Zhang, G.; Holford, P.; Wu, F. Tolerance to Drought, Low pH and Al Combined Stress in Tibetan Wild Barley Is Associated with Improvement of ATPase and Modulation of Antioxidant Defense System. Int. J. Mol. Sci. 2018, 19, 3553. https://doi.org/10.3390/ijms19113553
Ahmed IM, Nadira UA, Qiu C-W, Cao F, Zhang G, Holford P, Wu F. Tolerance to Drought, Low pH and Al Combined Stress in Tibetan Wild Barley Is Associated with Improvement of ATPase and Modulation of Antioxidant Defense System. International Journal of Molecular Sciences. 2018; 19(11):3553. https://doi.org/10.3390/ijms19113553
Chicago/Turabian StyleAhmed, Imrul Mosaddek, Umme Aktari Nadira, Cheng-Wei Qiu, Fangbin Cao, Guoping Zhang, Paul Holford, and Feibo Wu. 2018. "Tolerance to Drought, Low pH and Al Combined Stress in Tibetan Wild Barley Is Associated with Improvement of ATPase and Modulation of Antioxidant Defense System" International Journal of Molecular Sciences 19, no. 11: 3553. https://doi.org/10.3390/ijms19113553