Mechanistic Analysis of Physicochemical Cues in Promoting Human Pluripotent Stem Cell Self-Renewal and Metabolism


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
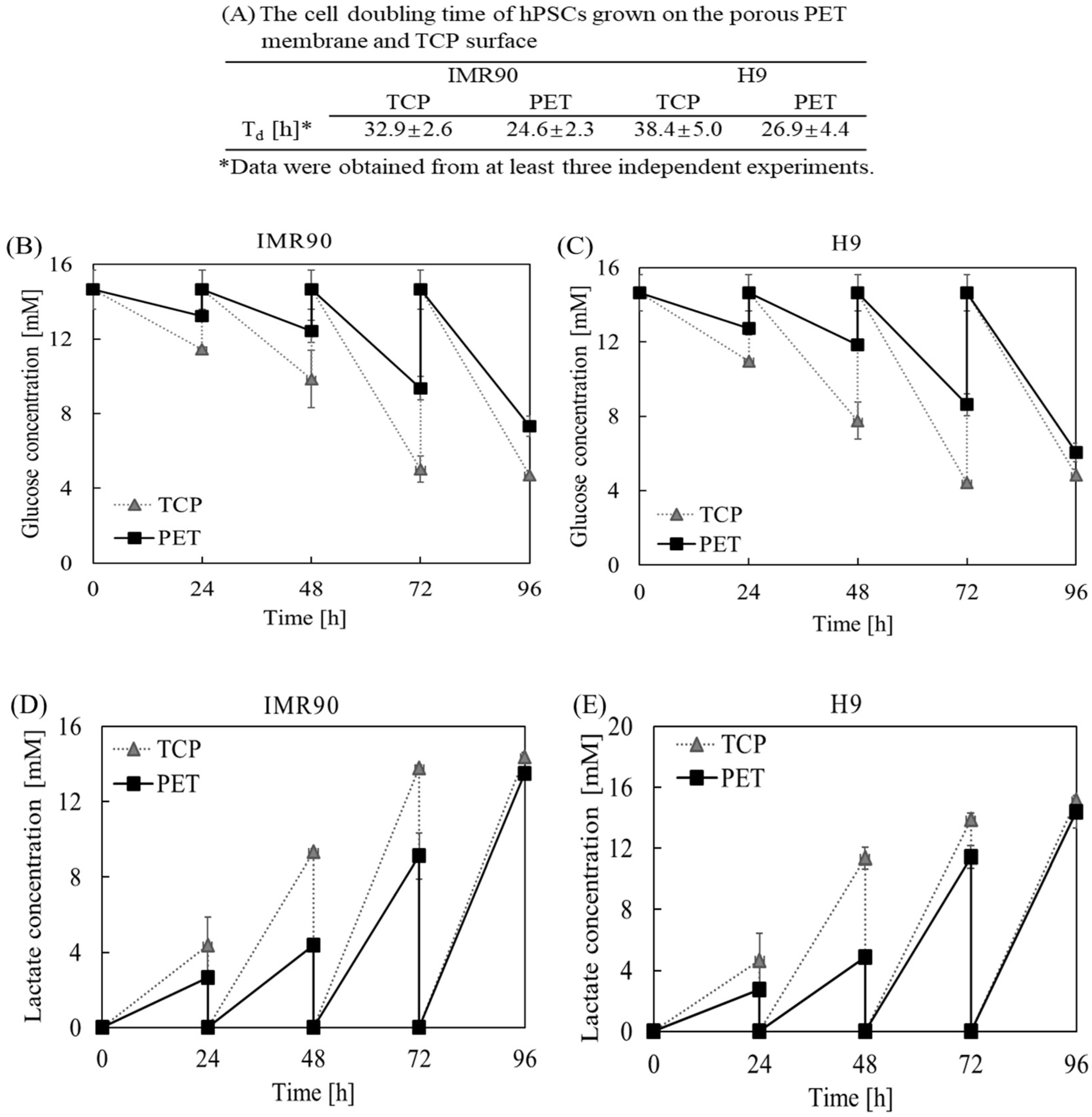
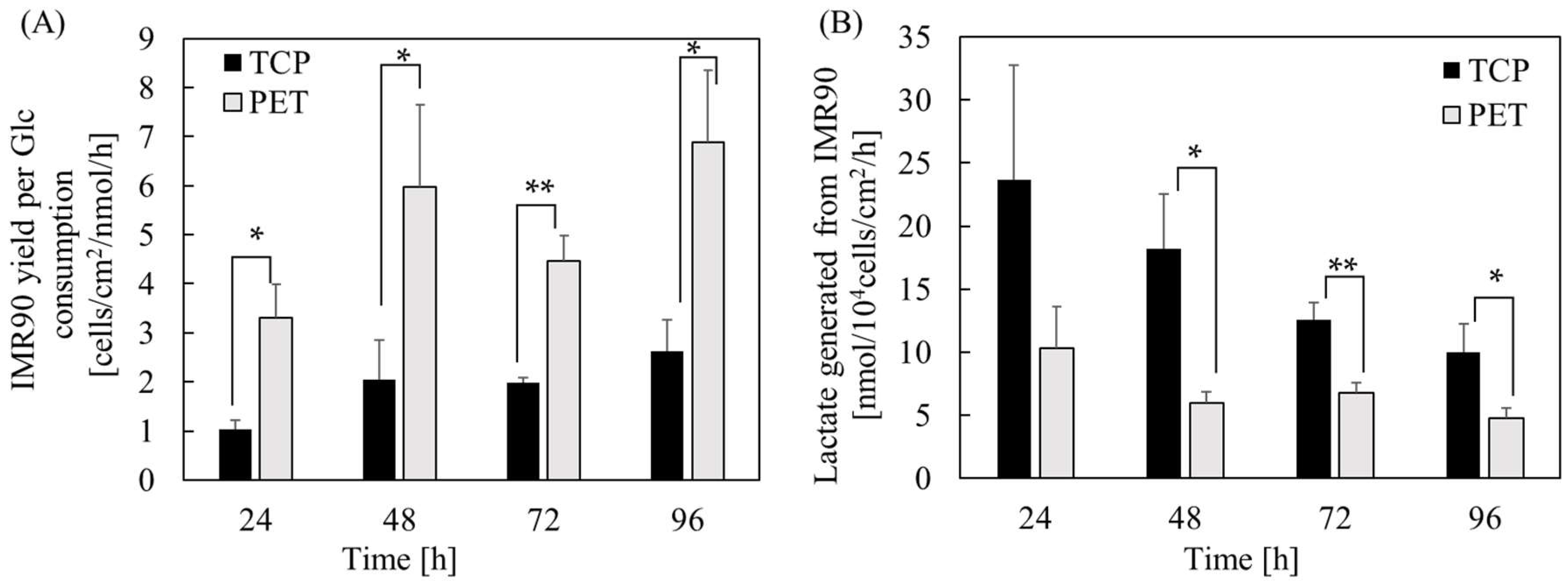
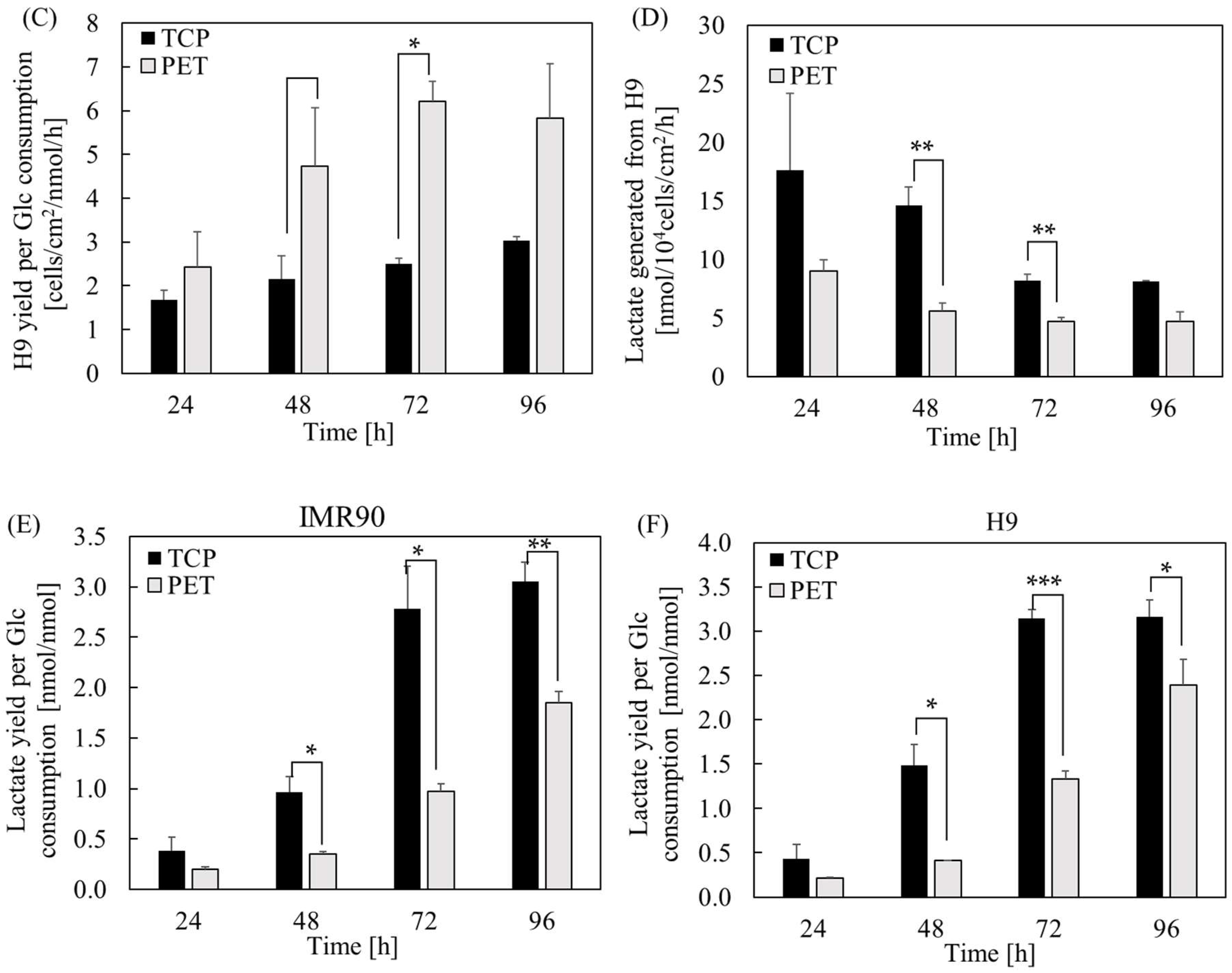
2.1. The Effect of Substrate Cues on hPSC Proliferation and Metabolism
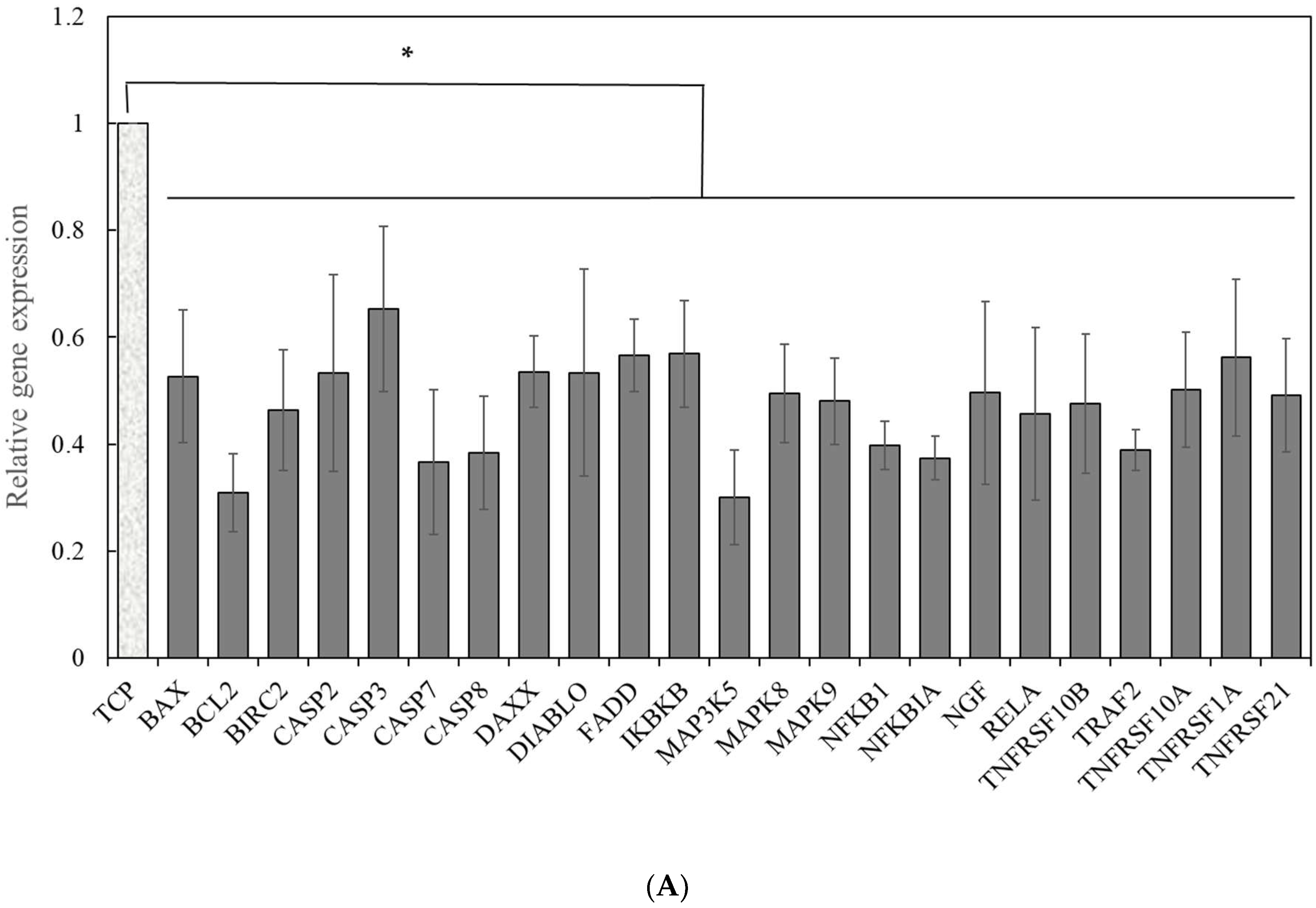
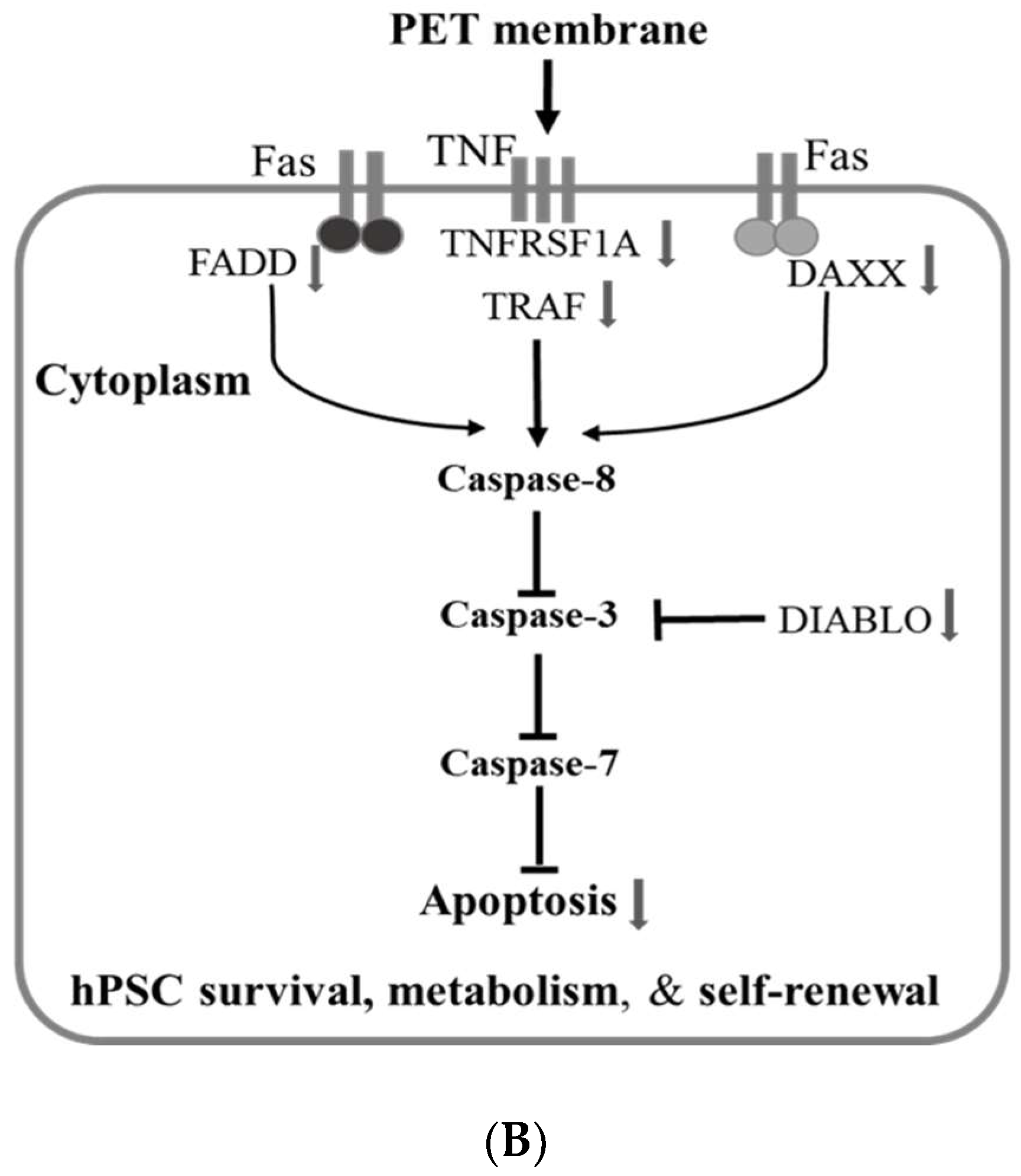
2.2. Signal Pathways Governing the Enhancement of hPSC Proliferation and Metabolism
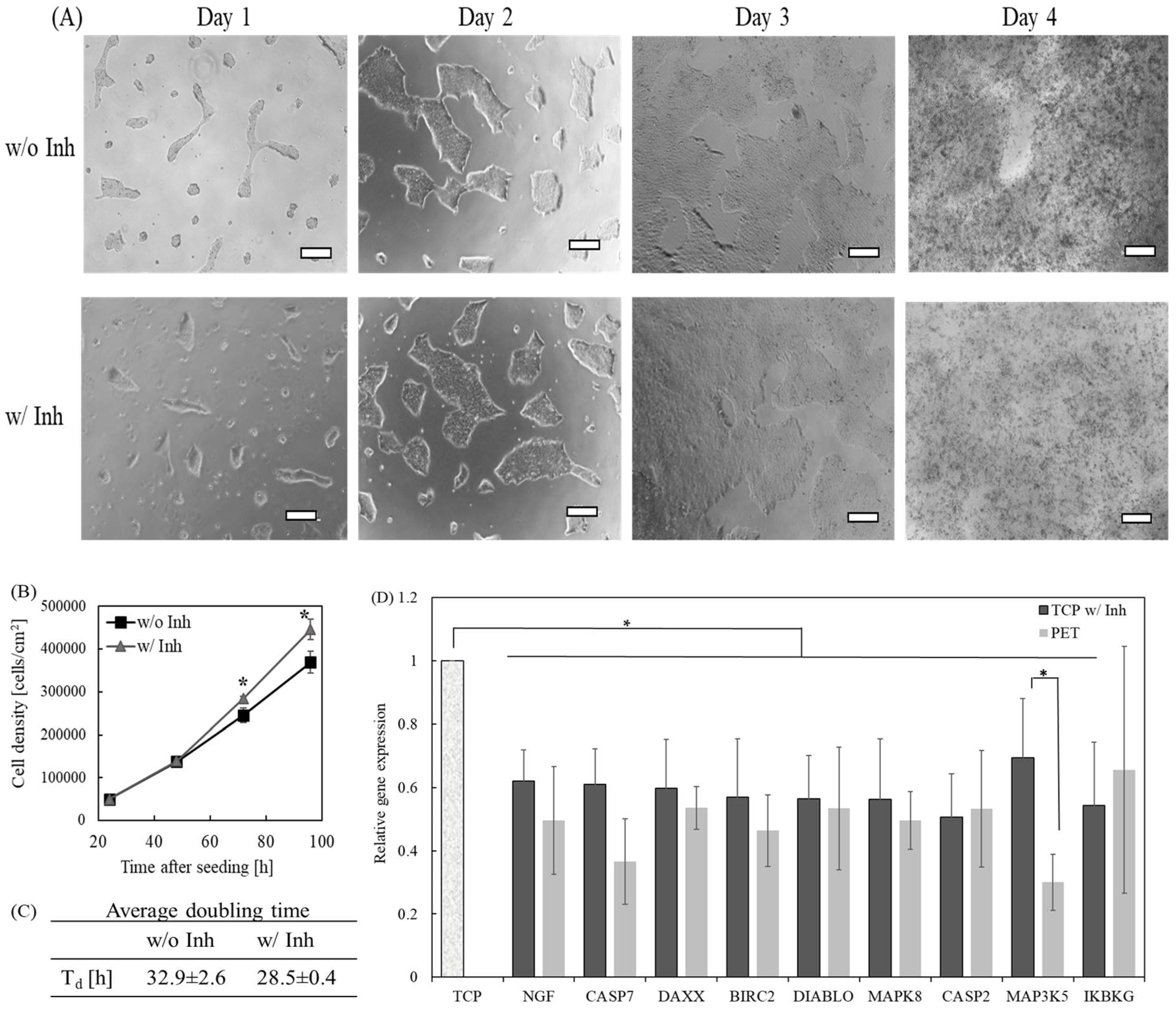
2.3. The Effect of Inhibition of Caspase-8-Mediated Pathway on the Self-Renewal of hPSCs Grown on the TCP Surface
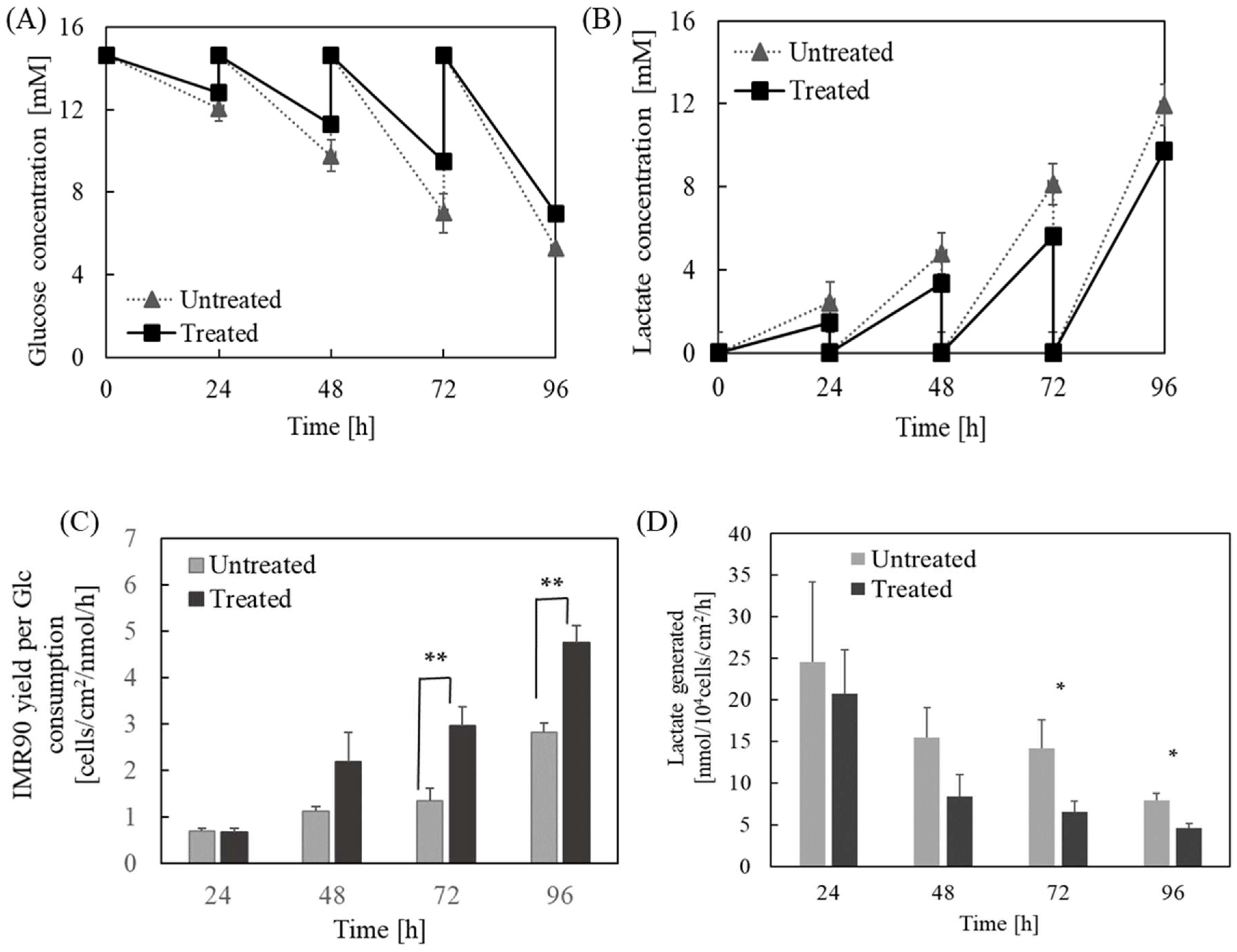
2.4. The Influence of Caspase-8-Mediated Pathway on the Metabolism of hPSCs Grown on the TCP Surface
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culture and Monitor of Cell Proliferation
5.2. Measurement of Glucose and Lactate Concentrations
5.3. Quantitative Real Time-PCR
5.4. Caspase-8 Inhibitor Z-IETD-FMK Treated Cell Culture
5.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Hibino, N.; Duncan, D.R.; Nalbandian, A.; Yi, T.; Qyang, Y.; Shinoka, T.; Breuer, C.K. Evaluation of the use of an induced puripotent stem cell sheet for the construction of tissue-engineered vascular grafts. J. Thorac. Cardiovasc. Surg. 2012, 143, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Villa-Diaz, L.G.; Ross, A.M.; Lahann, J.; Krebsbach, P.H. Concise Review: The Evolution of human pluripotent stem cell culture: From feeder cells to synthetic coatings. Stem Cells 2013, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Burridge, P.W.; Pryzhkova, M.V.; Levine, M.A.; Park, T.S.; Roxbury, C.; Yuan, X.; Peault, B.; Zambidis, E.T. Challenges and strategies for generating therapeutic patient-specific hemangioblasts and hematopoietic stem cells from human pluripotent stem cells. Int J. Dev. Biol 2010, 54, 965–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Yao, H.; Krisanarungson, P.; Haukas, A.; Ye, K. Porous membrane substrates offer better niches to enhance the Wnt signaling and promote human embryonic stem cell growth and differentiation. Tissue Eng. Part A 2012, 18, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yao, H.; Weber, J.L.; Melkoumian, Z.K.; Ye, K. A synthetic, xeno-free Peptide surface for expansion and directed differentiation of human induced pluripotent stem cells. PLoS ONE 2012, 7, e50880. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jin, S.; Ye, K. Development of Islet Organoids from H9 Human Embryonic Stem Cells in Biomimetic 3D Scaffolds. Stem Cells Dev. 2017, 26, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Hazeltine, L.B.; Selekman, J.A.; Palecek, S.P. Engineering the human pluripotent stem cell microenvironment to direct cell fate. Biotechnol. Adv. 2013, 31, 1002–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voog, J.; Jones, D.L. Stem cells and the niche: A dynamic duo. Cell Stem Cell 2010, 6, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Desbordes, S.C.; Studer, L. Adapting human pluripotent stem cells to high-throughput and high-content screening. Nat. Protoc. 2013, 8, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Laperle, A.; Hsiao, C.; Lampe, M.; Mortier, J.; Saha, K.; Palecek, S.P.; Masters, K.S. alpha-5 Laminin Synthesized by Human Pluripotent Stem Cells Promotes Self-Renewal. Stem Cell Rep. 2015, 5, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Wu, F.; Zhou, T.; Cai, X.; Zhang, S.; Zhang, X.; Li, Q.; Li, Y.; Zheng, Y.; Wang, M.; et al. Simple and versatile synthetic polydopamine-based surface supports reprogramming of human somatic cells and long-term self-renewal of human pluripotent stem cells under defined conditions. Biomaterials 2016, 87, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Yang, K.; Kim, M.J.; Jang, J.; Lee, M.; Kim, D.W.; Lee, H.; Cho, S.W. Bio-inspired oligovitronectin-grafted surface for enhanced self-renewal and long-term maintenance of human pluripotent stem cells under feeder-free conditions. Biomaterials 2015, 50, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Enam, S.; Jin, S. Substrates for clinical applicability of stem cells. World J. Stem Cells 2015, 7, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yoshioka, M.; Nakajima, M.; Ogasawara, A.; Liu, J.; Hasegawa, K.; Li, S.; Zou, J.; Nakatsuji, N.; Kamei, K.; et al. Nanofibrous gelatin substrates for long-term expansion of human pluripotent stem cells. Biomaterials 2014, 35, 6259–6267. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, K.; Narayanan, K.; Ni, M.; Du, C.; Wan, A.C.; Zink, D. Polysulfone membranes coated with polymerized 3,4-dihydroxy-l-phenylalanine are a versatile and cost-effective synthetic substrate for defined long-term cultures of human pluripotent stem cells. Biomacromolecules 2014, 15, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Musah, S.; Morin, S.A.; Wrighton, P.J.; Zwick, D.B.; Jin, S.; Kiessling, L.L. Glycosaminoglycan-binding hydrogels enable mechanical control of human pluripotent stem cell self-renewal. ACS Nano 2012, 6, 10168–10177. [Google Scholar] [CrossRef] [PubMed]
- Klim, J.R.; Li, L.; Wrighton, P.J.; Piekarczyk, M.S.; Kiessling, L.L. A defined glycosaminoglycan-binding substratum for human pluripotent stem cells. Nat. Methods 2010, 7, 989–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, Y.; Saha, K.; Bogatyrev, S.R.; Yang, J.; Hook, A.L.; Kalcioglu, Z.I.; Cho, S.W.; Mitalipova, M.; Pyzocha, N.; Rojas, F.; et al. Combinatorial development of biomaterials for clonal growth of human pluripotent stem cells. Nat. Mater. 2010, 9, 768–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, S.; Riggs, M.J.; Nelson, K.; Lee, V.; Rao, R.R. Characterization of human fibroblast-derived extracellular matrix components for human pluripotent stem cell propagation. Acta Biomater. 2010, 6, 4622–4633. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.R.; Kanninen, L.; Kuisma, T.; Niklander, J.; Noon, L.A.; Burks, D.; Urtti, A.; Yliperttula, M. The use of nanofibrillar cellulose hydrogel as a flexible three-dimensional model to culture human pluripotent stem cells. Stem Cells Dev. 2014, 23, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Kamei, K.I.; Koyama, Y.; Tokunaga, Y.; Mashimo, Y.; Yoshioka, M.; Fockenberg, C.; Mosbergen, R.; Korn, O.; Wells, C.; Chen, Y. Characterization of Phenotypic and Transcriptional Differences in Human Pluripotent Stem Cells under 2D and 3D Culture Conditions. Adv. Healthc Mater. 2016, 5, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Johansson, J.; Damdimopoulou, P.; Shahsavani, M.; Falk, A.; Hovatta, O.; Rising, A. Spider silk for xeno-free long-term self-renewal and differentiation of human pluripotent stem cells. Biomaterials 2014, 35, 8496–8502. [Google Scholar] [CrossRef] [PubMed]
- Abbasalizadeh, S.; Larijani, M.R.; Samadian, A.; Baharvand, H. Bioprocess development for mass production of size-controlled human pluripotent stem cell aggregates in stirred suspension bioreactor. Tissue Eng. Part C Methods 2012, 18, 831–851. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Gupta, S.; Yang, Y.; Forsyth, N.R. αVβ5 and CD44 are oxygen-regulated human embryonic stem cell attachment factors. Biomed. Res. Int. 2013, 2013, 729281. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Eshghi, S.; Li, Y.J.; Schmidt, R.; Schaffer, D.V.; Healy, K.E. Characterization of integrin engagement during defined human embryonic stem cell culture. FASEB J. 2010, 24, 1056–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Kim, J.; Park, T.J.; Shin, Y.; Lee, S.Y.; Han, Y.M.; Kang, S.; Park, H.S. The effects of the physical properties of culture substrates on the growth and differentiation of human embryonic stem cells. Biomaterials 2011, 32, 8816–8829. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Khosravi-Far, R.; Chang, H.Y.; Baltimore, D. Daxx, a novel Fas-binding protein that activates JNK and apoptosis. Cell 1997, 89, 1067–1076. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Desouza, M.; Gunning, P.W.; Stehn, J.R. The actin cytoskeleton as a sensor and mediator of apoptosis. Bioarchitecture 2012, 2, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinarsky, V.; Krivanek, J.; Rankel, L.; Nahacka, Z.; Barta, T.; Jaros, J.; Andera, L.; Hampl, A. Human embryonic and induced pluripotent stem cells express TRAIL receptors and can be sensitized to TRAIL-induced apoptosis. Stem Cells Dev. 2013, 22, 2964–2974. [Google Scholar] [CrossRef] [PubMed]
- Heng, B.C.; Clement, M.V.; Cao, T. Caspase inhibitor Z-VAD-FMK enhances the freeze-thaw survival rate of human embryonic stem cells. Biosci. Rep. 2007, 27, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Chai, J.; Suber, T.L.; Wu, J.W.; Du, C.; Wang, X.; Shi, Y. Structural basis of IAP recognition by Smac/DIABLO. Nature 2000, 408, 1008–1012. [Google Scholar] [CrossRef] [PubMed]
- Eletto, D.; Chevet, E.; Argon, Y.; Appenzeller-Herzog, C. Redox controls UPR to control redox. J. Cell Sci. 2014, 127, 3649–3658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Street, C.A.; Bryan, B.A. Rho kinase proteins--pleiotropic modulators of cell survival and apoptosis. Anticancer Res. 2011, 31, 3645–3657. [Google Scholar] [PubMed]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton (Hoboken) 2010, 67, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaibuchi, K.; Kuroda, S.; Amano, M. Regulation of the cytoskeleton and cell adhesion by the Rho family GTPases in mammalian cells. Annu. Rev. Biochem. 1999, 68, 459–486. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yang, H.; Yu, L.; Xu, C.L.; Zeng, Y.; Qiu, Y.; Li, D.S. Feeder-free and xeno-free culture of human pluripotent stem cells using UCBS matrix. Cell Biol. Int. 2015, 39, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Hourigan, K.; Jadhav, S.; Bellare, J.; Verma, P. Effect of lactate and pH on mouse pluripotent stem cells: Importance of media analysis. Biochem. Eng. J. 2017, 118, 25–33. [Google Scholar] [CrossRef]
- Wen, Y.; Jin, S. Production of neural stem cells from human pluripotent stem cells. J. Biotechnol. 2014, 188, 122–129. [Google Scholar] [CrossRef] [PubMed]



 signifies inhibition.
signifies inhibition.
signifies inhibition.
signifies inhibition.



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hai, N.; Shin, D.W.; Bi, H.; Ye, K.; Jin, S. Mechanistic Analysis of Physicochemical Cues in Promoting Human Pluripotent Stem Cell Self-Renewal and Metabolism. Int. J. Mol. Sci. 2018, 19, 3459. https://doi.org/10.3390/ijms19113459
Hai N, Shin DW, Bi H, Ye K, Jin S. Mechanistic Analysis of Physicochemical Cues in Promoting Human Pluripotent Stem Cell Self-Renewal and Metabolism. International Journal of Molecular Sciences. 2018; 19(11):3459. https://doi.org/10.3390/ijms19113459
Chicago/Turabian StyleHai, Nan, Dong Woo Shin, Huanjing Bi, Kaiming Ye, and Sha Jin. 2018. "Mechanistic Analysis of Physicochemical Cues in Promoting Human Pluripotent Stem Cell Self-Renewal and Metabolism" International Journal of Molecular Sciences 19, no. 11: 3459. https://doi.org/10.3390/ijms19113459