Salvia Species as Sources of Natural Products with Antiprotozoal Activity



Institut für Pharmazeutische Biologie und Phytochemie (IPBP), University of Münster, PharmaCampus, Corrensstraße 48, D-48149 Münster, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(1), 264; https://doi.org/10.3390/ijms19010264
Submission received: 15 December 2017
/
Revised: 5 January 2018
/
Accepted: 11 January 2018
/
Published: 16 January 2018
(This article belongs to the Section Bioactives and Nutraceuticals)
Abstract
:Natural products from plants have been used since ancestral times to treat a wide variety of diseases worldwide. Plants of the genus Salvia (Sage) have been reported to be used for the prevention and treatment of various diseases and ailments. In particular, some Salvia species have been used in traditional medicine to treat diseases caused by protozoan parasites of the genera Trypanosoma, Leishmania and Plasmodium and scientific studies have demonstrated the activity of various isolated constituents from these plants against these pathogens. The current review attempts to give a critical overview of published information about the antiprotozoal activity of species of the genus Salvia and their chemical constituents. It is meant to give a unified overview of these results in order to avoid repetitions caused, e.g., by limited access to some primary reports, and to stimulate further research to possibly facilitate the development of new molecular leads against protozoal neglected tropical diseases (NTDs) based on Salvia constituents.
1. Introduction
Salvia (Sage) is the largest genus of the family Lamiaceae. It is widespread in various regions around the world comprising the Mediterranean area, South Africa, Central and South America as well as Asia [1,2], and some of its representatives are also cultivated and exported to other regions worldwide. The genus comprises around 1000 species [2,3], many of which have long been used as flavoring agents, aromatics, and ornamental plants as well as in perfumery and cosmetics. Salvia species are also known for a wide variety of medicinal uses in folk medicine. In fact, the name Salvia is derived from the Latin word salvare, which refers to its healing qualities. In Europe, the leaves of Salvia officinalis L. and Salvia triloba L. (a synonym of S. fruticosa Mill.) are extensively used as a mouth wash against inflammations in the oral cavity [1]. The roots of Salvia miltiorrhiza Bunge, a species endemic to China and Japan, have long been used in Traditional Chinese Medicine (TCM) to treat circulation-related diseases [4]. Salvia sclarea L. is used as a remedy to alleviate night sweats associated with menopause or tuberculosis [1] and also used in perfumery as a flavoring [5]. South African species such as Salvia stenophylla Burch. ex Benth., Salvia runcinata L. and Salvia repens Burch. ex Benth. have long been used as disinfectants and as purgative [6].
Infectious diseases caused by unicellular eukaryotic parasites (“protozoans”) of the genera Trypanosoma (Chagas disease and sleeping sickness), Leishmania (different forms of leishmaniasis)—which are among the list of 20 neglected tropical diseases (NTDs) recognized by the WHO—and Plasmodium (malaria), threaten the life of millions of people worldwide. The majority of affected populations are from tropical middle and low income countries, but since parasites do not respect national and political borders, protozoal infections threaten other populations and thus constitute a global health problem. Both the lack of safe and available treatments, the threatening increase of drug-resistance against current therapies and the insecticide resistance for vector control make the research on and development of new anti-protozoal drugs an urgent need.
Each of the mentioned parasites has a complex life cycle in which various forms occur subsequently in different host organisms (insect vectors, infected mammalians; for a brief overview see, e.g., [7], or specialized parasitological literature). These life forms differ from each other in physiology and, hence, drug susceptibility. It is hence of great importance when searching for new active entities to use in vitro bioassay models of the clinically most relevant forms of the parasites. These are trypomastigote forms as present in the bloodstream and cerebroventricular fluid in case of T. brucei, amastigote forms as present in host cells in case of T. cruzi and Leishmania species and intraerythrocytic forms in case of Plasmodium spp. (for an overview on assay systems, see, e.g., [7]).
Salvia is distributed worldwide and cultivated so that some species grow where protozoal neglected diseases are endemic. Despite the large spectrum of medicinal uses of Sage species, which mainly describe them against conditions associated with fever and perspiration [6], relatively few references have reported on their traditional use against infectious diseases caused by protozoans. However, a few publications recognize their ethnomedicinal use as antimalarial, antitrypanosomal and antileishmanial remedies [6,8].
Terpenoids (essential oils as well as di- and tri-terpenes) and phenolic compounds, such as flavonoids and caffeic acid derivatives, are known to be present in aerial parts of Salvia species. Hundreds of secondary metabolites have been isolated from different species of Salvia and have been tested for a wide range of biological activities [5].
Different research groups have been investigating Salvia species as well as some of their isolated constituents, reporting them as possible antiprotozoal agents against parasites of the genera Plasmodium, Trypanosoma and Leishmania. Even though some of the reported chemical constituents showed promising activities and encouraging results, to the best of our knowledge, no data on research focused on a direct applicability or further development of Salvia or its constituents (e.g., on mechanism of action studies, structural optimization or clinical trials against protozoal infections) exist.
This review is intended to summarize and give a critical overview of the published information concerning the antiprotozoal activity of Salvia species and their isolated compounds. This collection of data is expected to help avoid research repetitions due, e.g., to limited access to primary data sources, and, possibly, to facilitate the development of new molecular leads against protozoal NTDs based on Salvia constituents.
The following specific abbreviations are used throughout this review: Neglected Tropical Diseases (NTDs), Trypanosoma brucei rhodesiense (Tbr), Trypanosoma brucei gambiense (Tbg), Trypanosoma brucei brucei (Tbb), Trypanosoma cruzi (Tc), Plasmodium falciparum (Pf), Leishmania donovani (Ld), Leishmania major (Lm), Leishmania infantum (Li), Leishmania killicki (Lk), Leishmania guyanensis (Lg), Leishmania tropica (Lt), Chagas disease (CD), sleeping sickness or Human African Trypanosomiasis (HAT), concentration causing half-maximal growth inhibition (IC50), and selectivity index (SI = IC50 (mammalian control cells)/IC50 (parasite)).
2. Methodology
An online literature search for information available about antiprotozoal activity of Salvia species and their constituents was conducted, based on common database sources (Web of Science, SciFinder, PubMed-NCBI and Google Scholar). In this search, combinations of the keyword Salvia with the terms antiprotozoal, antitrypanosomal, antimalarial, antileishmanial, malaria, Chagas disease, sleeping sickness, African trypanosomiasis, Leishmania, Trypanosoma and Plasmodium were applied.
A total number of 34 publications from the years 2002 to 2017 were obtained. The results of these studies have been ordered in this review by biological activity (i.e., the parasite against which the studies were performed) and then by structurally/biosynthetically related secondary metabolites of Salvia species with their antiprotozoal activity. Table 1, Table 2, Table 3, Table 4, Table 5, Table 6, Table 7 and Table 8 and Figure 1, Figure 2, Figure 3 and Figure 4 summarize the content of this review.
Thirty-five species of Salvia and 45 compounds isolated from Salvia species have been reported to be tested against at least one of the protozoans treated in this review.
3. Biological Activity
3.1. Antitrypanosomal Activity
Protozoans of the genus Trypanosoma cause fatal diseases which threaten the lives of millions of people worldwide.
Trypanosoma brucei rhodesiense (Tbr) and T. brucei gambiense (Tbg) are parasites transmitted by tsetse flies (Glossina spp.) and are etiological agents of different forms of Human African Trypanosomiasis (HAT), also known as “sleeping sickness”. HAT is endemic in 36 sub-Saharan countries [9] and its global burden was estimated to be 0.3 million disability-adjusted life years (DALYs) in 2015 [10]. Other species of Trypanosoma such as T. brucei brucei (Tbb) are known to be responsible for diseases in animals. The latter, together with other species such as T. congolense, is a causative agent of the cattle disease Nagana, also responsible for a severe economic and health problem in some parts of Africa [11].
The currently available diagnostic tests and treatment of the HAT require skilled personnel and, in most cases, hospitalization, to which most of the affected people living in rural areas do not have access. Moreover, the current therapies are painful and present severe side effects. All in all, new safe, orally applicable and inexpensive treatments for HAT are urgently needed [11]. The very recent positive outcome of phase III clinical trials with the synthetic compound fexinidazole [12], the first orally applicable drug against Tbg infection, may be seen as an encouraging step in this direction.
The American trypanosomiasis, also known as Chagas disease, is caused by the protozoan Trypanosoma cruzi, which is transmitted mainly by triatomine bugs (Triatoma spp., Rhodnius spp. and Panstrongylus spp.) but also by congenital transmission, blood transfusion or organ transplants. A number of cases of infection after ingestion of the parasites with food or drink have also been reported [13]. It is endemic in 21 countries of South America where the vectors live. Moreover, migration and travelers extended the disease around the world. In non-endemic countries such as Spain or Canada, around 50,000 and 1800 of people, respectively, are estimated to be infected with Tc [14]. Because of its unspecific symptomatology in early phases of infection, thousands of patients remain undiagnosed. It is estimated that the disease affects around 8 million people and causes 20,000 deaths every year [15]. The known resistance and lack of sensitivity of this parasite to existing therapies (as well as in-research-drugs) make the research in this field particularly difficult and challenging.
3.1.1. Activity of Crude Extracts and Isolated Constituents from Salvia Species against Trypanosma Brucei
Extracts and Essential Oils
Some Salvia species and compounds isolated from them have been reported to show biological activity against the etiological agents of HAT and of the cattle disease, Nagana. The reported activity data for 12 extracts obtained from three Salvia species against T. brucei spp. are summarized in Table 1. Methanol extracts and their partitions with n-hexane, chloroform and water of Salvia tomentosa Mill., Salvia sclarea L. and Salvia dichroantha Stapf. were tested against Tbr STIB 900 with IC50 values ranging from 1.24 to 10.96 µg/mL (named as a1–c4 in Table 1), being the n-hexane partition of S. tomentosa Mill. (a2) the most active, with an IC50 = 1.24 µg/mL and significant selectivity with an SI = 21.16, compared with L6 rat skeletal myoblast cells [16]. More and deeper investigations with the most active extracts of these species should be performed to find the compound(s) responsible for their biological activity.
Farimani and coworkers reported on the activity of an n-hexane extract of Salvia hydrangea DC. ex Benth. (IC50 = 18 µg/mL against Tbr STIB 900 strain) (d) (Table 1) [17]. Furthermore, a dichloromethane extract from the roots of S. miltiorrhiza Bunge (named as e in Table 1) showed 97% of growth inhibition against Tbr at 0.81 µg/mL [4]. Moreover, a methanol-dichloromethane extract of S. repens Burch. ex Benth. (f) (Table 1) showed less promising activity with an IC50 value of 10.8 µg/mL [18].
An ethanolic extract of Salvia spathacea Greene. (g) (Table 1) was tested by Jain and coworkers against Tbb showing interesting IC50 values of 1.13 ± 0.78 µg/mL [19].
Ihsan et al. tested the essential oil of Salvia lavandulifolia Vahl. (h) (Table 1) against the trypomastigote form of Tbb which did not show inhibitory effects at concentration up to 20 µg/mL [20].
Recently, as part of the present authors’ own work, an extract obtained by evaporation of a commercial tincture of S. officinalis L. (i) (Table 1) was found to exhibit quite significant activity against Tbr with an IC50 of 1.86 µg/mL and an SI (determined with L6 rat skeletal myoblasts) of about 17 [21]. Isolation and characterization of the main active principles of this extract is in progress.
In most of the cases where active constituents were characterized, diterpenoids and triterpenoids were responsible for the reported antitrypanosomal activity (summarized in Table 2).
Diterpenoids
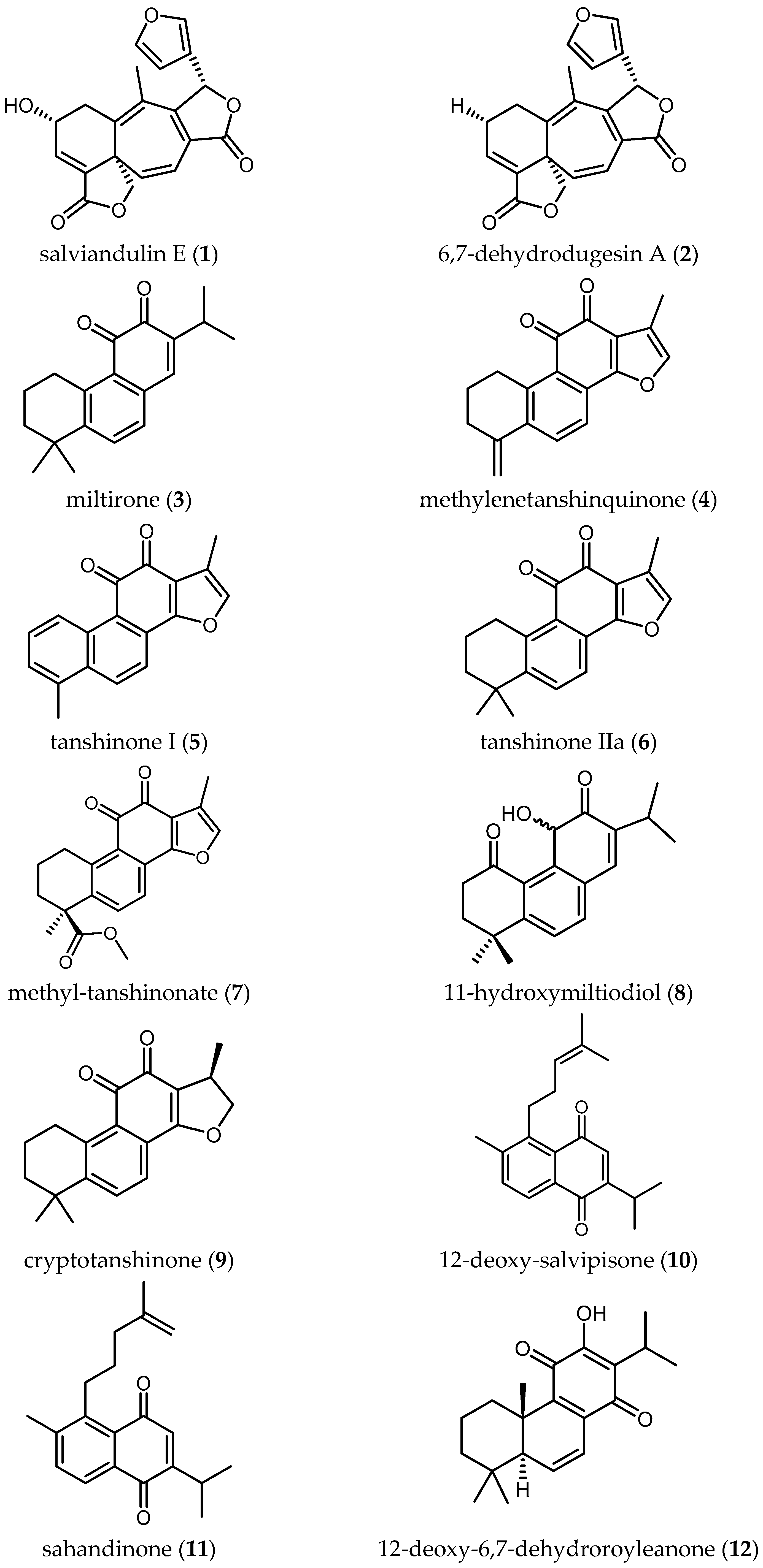
Salviandulin E (1)—a neoclerodane diterpene isolated originally from Salvia leucantha Cav.—and a set of synthetic analogs were tested in vitro against Tbb GUTat 3.1 by Aoyagi and coworkers. Compound 1 itself showed potent biological activity (IC50 = 2.04 µM). A similar diterpene isolated from the same plant, 6,7-dehydrodugesin A (2), did not show antitrypanosomal activity (IC50 > 37.19 µM) [22]. The synthetic derivative butanoyl 3,4-dihydro salviandulin showed the best IC50 = 0.14 µM and SI = 1236 [22]. Although is known that Tbb and Tbr are genetically almost identical [7], some compounds showed different bioactivity against the two parasites [7]. Therefore, studies against Tbr or Tbg should be carried out with these compounds to evaluate the biological activity against human pathogenic parasites. The improvement of the activity and selectivity of the compounds by a semi-synthetical approach are a nice example that relatively simple chemical modification of natural products can be an important step in drug development.
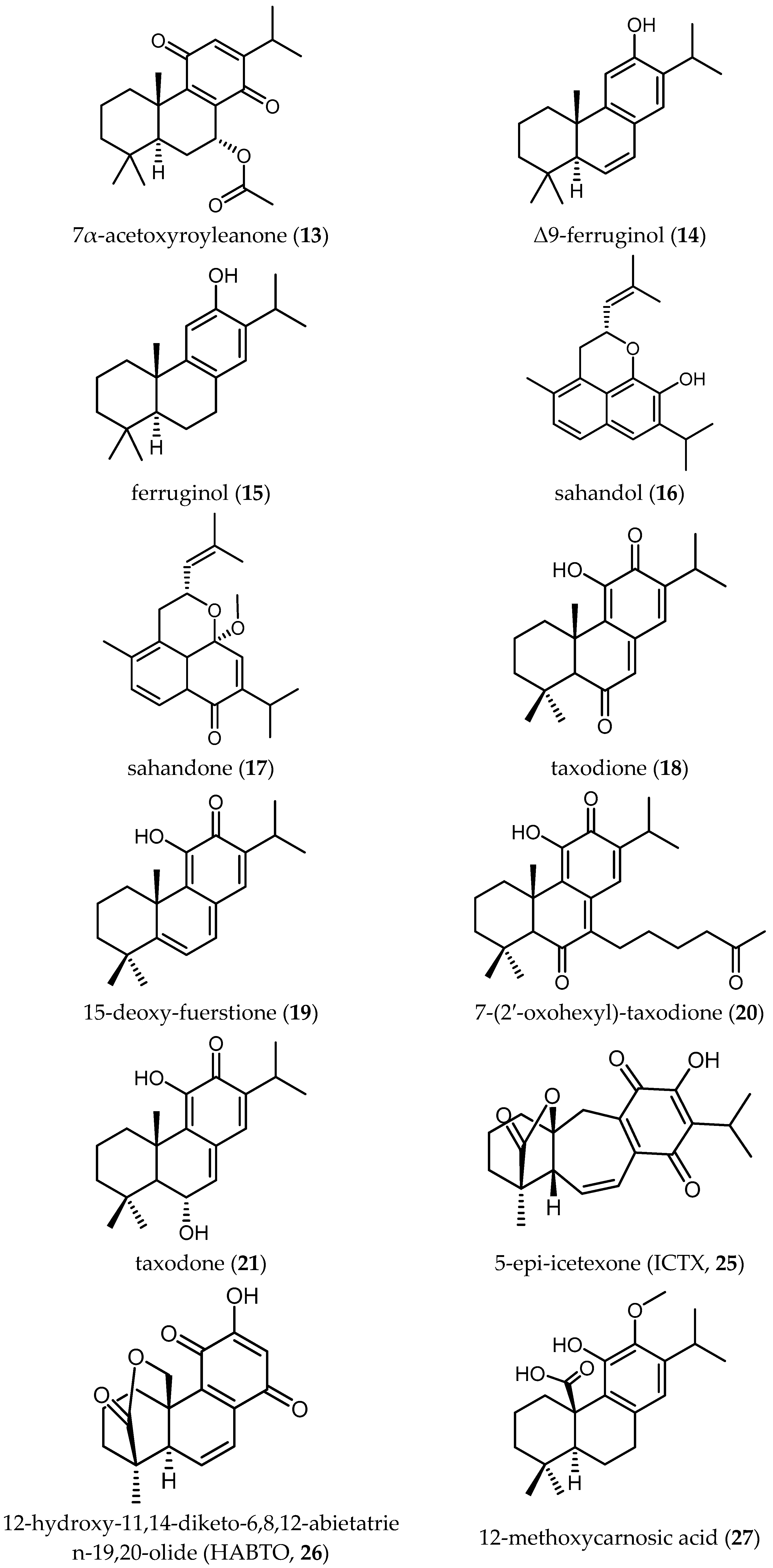
Some tanshinone-type diterpenoids isolated from S. miltiorrhiza Bunge [4] showed activity ranging from 0.5 to 26.2 µM and SI from 0.2 to 24.2. Miltirone (3) and methylenetanshinquinone (4) displayed the highest activity, both with IC50 values of 0.5 µM and SI of 2.6 and 24.2, respectively. Tanshinone I (5) also yielded a promising IC50 of 1.3 µM and an SI of 9.5. Less active diterpenes (6–9) isolated in the same study are also included in Table 2.
Ebrahimi and colleagues reported on four diterpene-quinones (10–13) and four abietane diterpenes (14–17) (Table 2), isolated from Salvia sahendica Boiss & Buhse, to have antitrypanosomal activity [23]. They exhibited interesting IC50 values from 1.8 to 28.1 µM against Tbr STIB 900 strain. Unfortunately, the low SI values (0.1–1.2) indicate the nonselective toxicity of these compounds. Chemical structural modifications of the reported compounds, as demonstrated in the case of Compound 1, might be attempted to decrease their toxicity.
Kuźma and coworkers isolated four quinone methide-type diterpenes from the roots of Salvia austriaca Jacq. (18–21 in Table 2) and tested them against trypomastigotes of Tbr STIB 900. They yielded promising IC50 values from 0.05 to 194.7 µM and SI from 0.69 to 38, taxodione (18) being the most active and promising compound, which showed IC50 of 0.05 µM and a favorable SI of 38 [24].
Triterpenoids
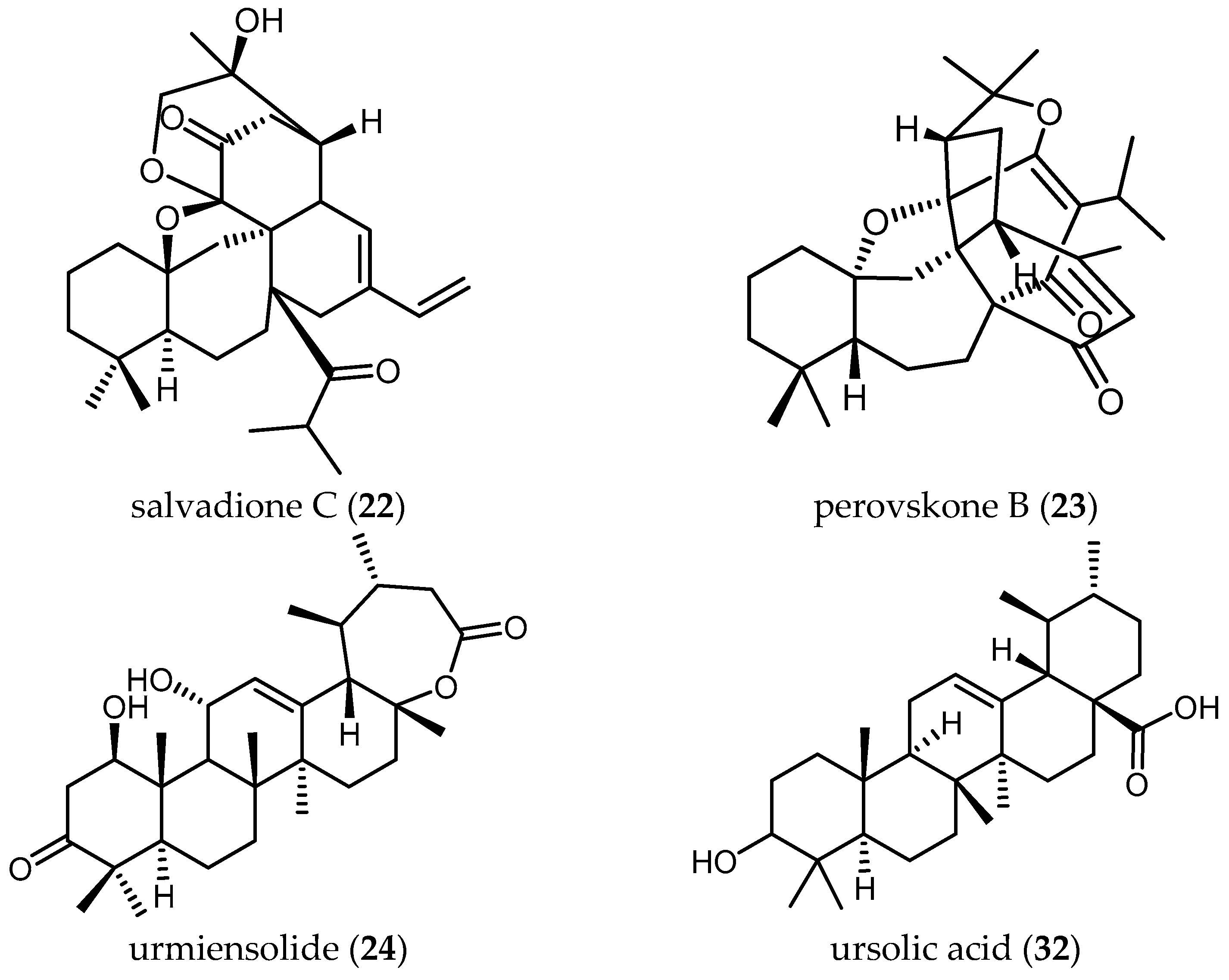
The triterpenoids salvadione C (22) and perovskone B (23) isolated from S. hydrangea DC. ex Benth. displayed low activity against Tbr STIB 900, with IC50 values of 4.33 and 15.92 μM, respectively [25].
Farimani also reported one triterpenoid with an ε-lactone E-Ring showing biological activity against Tbr STIB 900 strain. Urmiensolide (24), isolated from Salvia urmiensis Bunge—an endemic Salvia species in the West Azerbaijan province of northwestern Iran—showed in vitro antitrypanosomal activity with an IC50 value of 5.6 μM and SI of 33 [17].
Further research on the mechanism(s) of action accompanied by studies on in vivo efficacy, bioavailability and possible metabolic routes would appear necessary to evaluate whether such triterpenes have a potential for further drug development.
3.1.2. Activity of Crude Extracts and Isolated Constituents from Salvia Species against Trypanosoma Cruzi
The antitrypanosomal activity of Salvia species against the etiological agent of Chagas disease (CD) has been less studied in comparison with the other protozoan diseases. However, some promising results have been found in this field (Table 3 and Table 4).
Extracts
In the same study cited above, with respect to Tbr [16], the methanol extracts and their partitions with n-hexane, chloroform and water obtained from S. tomentosa Mill., S. sclarea L. and S. dichroantha Stapf. (named as a1–c4 in Table 3) were tested against Tc trypomastigotes. In contrast with the relatively promising biological activity values against Tbr, the IC50 values against Tc were considerably higher, ranging from 18.17 µg/mL to >90 µg/mL and displayed unfavorable SI values ranging from 0.89 to ≥2.15 [16].
Diterpenoids
A novel icetexane diterpene, 5-epi-icetexone (ICTX) (25), isolated from aerial parts of Salvia gilliessi Benth.—a Salvia species native to an endemic Chagas region—was tested against cultured epimastigotes of Tc (Tulahuen strain) to investigate the influence on proliferation, viability and morphology of the parasites and understand the mechanism of action. Compound 25 showed an antiproliferative effect on the Tc epimastigotes beginning at a concentration of 2.8 µM and its IC50 was determined as 6.5 ± 0.75 µM. The compound exhibited low cytotoxicity to mammalian cells. A comparison with other strains (CL-Brener, Dm28c) was also carried out but no significant differences were observed. An interaction with the DNA of the parasite has been suggested as one the mechanisms of action responsible for the cytostatic effect, but it was concluded that multiple mechanisms should be involved [27]. Further investigation with other parasite stages was carried out by Lozano and colleagues testing 25 against amastigotes with observable effects at concentrations of 1.5–3.8 μM. However, no IC50 values have been reported [28]. The promising activity of 25 against Tc justified the further studies in vivo in Swiss albino mice infected with Tc (Tulahuen strain Tc VI). The survival of animals treated daily with 10 mg/kg/day of 25 increased significantly in comparison with the control (saline or 0.1% DMSO), and also a weekly decrease of parasitemia was observed. Moreover, the treatment with 25 prevented the nest formation of amastigotes within muscle fibers [29]. All these data make Compound 25 a favorable lead compound to be considered for further drug development steps.
The effect of the abietane diterpene 12-hydroxy-11,14-diketo-6,8,12-abietatrien-19,20-olide (HABTO) (26) isolated from the aerial parts of Salvia cuspidata subsp. gilliesii (Benth.) J.R.I.Wood on Tc epimastigotes was also studied by Lozano and colleagues [30]. This compound was reported to inhibit parasite growth at moderate concentrations (IC50 ≈ 16.6 µM) and described to be of low toxicity for mammalian cells (SI value not available). In this study, some chemical modifications rendering the compound more lipophilic also succeeded to improve the trypanocidal activity. The 12-trimethylsilyloxy derivative of 26 showed the highest toxicity to the parasite, representing another example that small chemical modifications can already improve the activity of the native natural products.
In the same study cited above with respect to Tbr, Kuźma and coworkers tested four diterpenes with a quinone moiety isolated from the roots of S. austriaca Jaqc. (18–21) against amastigotes of Tc (Tulahuen strain C2C4 containing the β-galactosidase Lac Z) [24]. The IC50 ranged from 7.11 µM to 146.9 µM, taxodione (18) being again the most active compound. Unfortunately, the unfavorable SI values (from 0.27 to 0.91) indicated non-selective toxicity against mammalian cells.
3.2. Antileishmanial Activity
Leishmaniasis is a group of diseases caused by more than 20 protozoan species of the genus Leishmania, transmitted by the bite of infected sandflies (genera Phlebotomus in the Old World, Lutzomyia in the New World). From the different manifested forms of this disease, cutaneous leishmaniasis is the most common and the visceral leishmaniasis—also known as Kala-Azar—is the most serious form, leading to death if untreated. According to estimates by WHO, 1.3 million new cases occur every year, and it is estimated that 20,000–30,000 people die annually of visceral leishmaniasis [31].
Notwithstanding the positive breakthrough in terms of diagnostic programs and the reduction of prices for the treatment, the mortality and morbidity due to leishmaniasis worldwide is still worrying [32]. The lack of vaccines against Leishmania spp. and the increasing resistance of the parasites against current therapies, make research and development of new treatments urgent.
Natural products from plant sources have long been studied as potential antileishmanial leads [33]. However, none of the investigated compounds seem to have reached clinical investigation so far.
In the case of Salvia, infusions from the flowers have reportedly been used in Iranian folk medicine as antileishmanial treatment [8]. Sixty-one species of Salvia occur in Iran, seventeen of which are endemic [17]. No further investigations, however, have been carried out to prove the efficacy of this treatment, but some diterpenes, triterpenes and extracts of other Salvia species have been reported in the published literature to be active against different species of Leishmania (Table 5 and Table 6).
3.2.1. Extracts and Essential Oils
Several research groups investigated the antileishmanial activity of different species of Salvia. The above-mentioned extracts of different polarity from three Salvia species (S. tomentosa Mill., S. sclarea L. and S. dichroantha Stapf.) were also tested against axenic amastigotes of Ld strain MHOM/ET/67/L82, showing IC50 values ranging from 1.81 to >90 µg/mL and favorable SI values ranging from 7.63 to 29.47 (Table 5). The most active extract in this case was the chloroform partition of S. tomentosa Mill. (named as a3 in Table 5) which displayed an IC50 of 1.81 µg/mL and an SI value of 29.47 [16].
In the same study cited above with respect to Tc [26], the dichloromethane-methanol (1:1) extract of S. repens Burch. ex Benth. was tested against axenic amastigotes of Ld. The IC50 was considerably better than in Tb, being 5.36 µg/mL, with an SI value of 7.74.
Ihsan and coworkers tested the essential oil of S. lavandulifolia Vahl. (h) against different stages of Ld. No leishmanicidal effects were observed at concentration up to 20 µg/mL (Table 5) [20].
The essential oil of S. officinalis L. (i1 in Table 5) was tested by Essid and coworkers against the promastigote forms of Lm and Li, yielding IC50 values of 3.40 ± 0.16 and 2.67 ± 0.33 µg/mL, and moderate SI values of 5.92 and 7.54, respectively [34]. Moreover, Nikmehr et al. tested the antiprotozoal activity of the methanolic maceration of S. officinlais L. (i2 in Table 5) against amastigotes and promastigotes of Lm. The macerated extract exhibited an unfavorable IC50 value against the promastigotes of 184 ± 11.17 µg/mL. Furthermore, at this concentration it yielded 58% of lethality of parasite internalized into macrophages (cell line infected with Lm amastigotes) and 8% of toxicity [35].
A.R. Khan and M.J. Khan investigated the antileishmanial activity of a methanolic extract from leaves from Salvia bucharica Popov. and different solvent partitions (named as j-j4 in Table 5) [36]. The extract demonstrated a low IC50 value of 72.31 µg/mL. The following partitions (chloroform, acetone and water partitions) showed IC50 values ranging from 30.51 to >100 µg/mL, the aqueous partition being the most active (Table 5). More research on the polar fraction should be carried to possibly identify single compounds with higher antileishmanial activity.
Three solvent partitions of Salvia verbenaca (L.) Briq. ssp verbenaca Maire (S. clandestina Batt. non L.) were tested by Et-Touys et al. against promastigote forms of Lm, Lt and Li [37]. The methanolic partition did not exhibit antileishmanial activity against any of the tested parasites. However, the dichloromethane partition showed better IC50 values ranging from 24.56 to 33.77 µg/mL. The most active extract was the n-hexane partition, which showed moderate activity against Li (IC50 = 14.11 µg/mL) but lower activity against Lm and Lt, with IC50 of 155.43 and 148.23 µg/mL, respectively (Table 5).
3.2.2. Diterpenes
The reported diterpenes isolated from Salvia species displaying antileishmanial activity belong to the abietane group. Their activity seems to be higher in comparison with the activity against the other parasites reported above, since most of them showed IC50 lower than 1 µM (Table 6).
Mokoka and colleagues reported on 12-methoxycarnosic acid (27), isolated from the moderately active dichloromethane-methanol (1:1) extract of the whole plant of S. repens Burch. ex Benth. (IC50 = 5.36 µg/mL) [26], showing a low IC50 value of 0.75 μM against axenically cultured amastigotes of Ld and a favorable SI of 23.06 [18]. These promising values make this compound interesting as a lead compound. However, no further studies to fully evaluate its potential have been found.
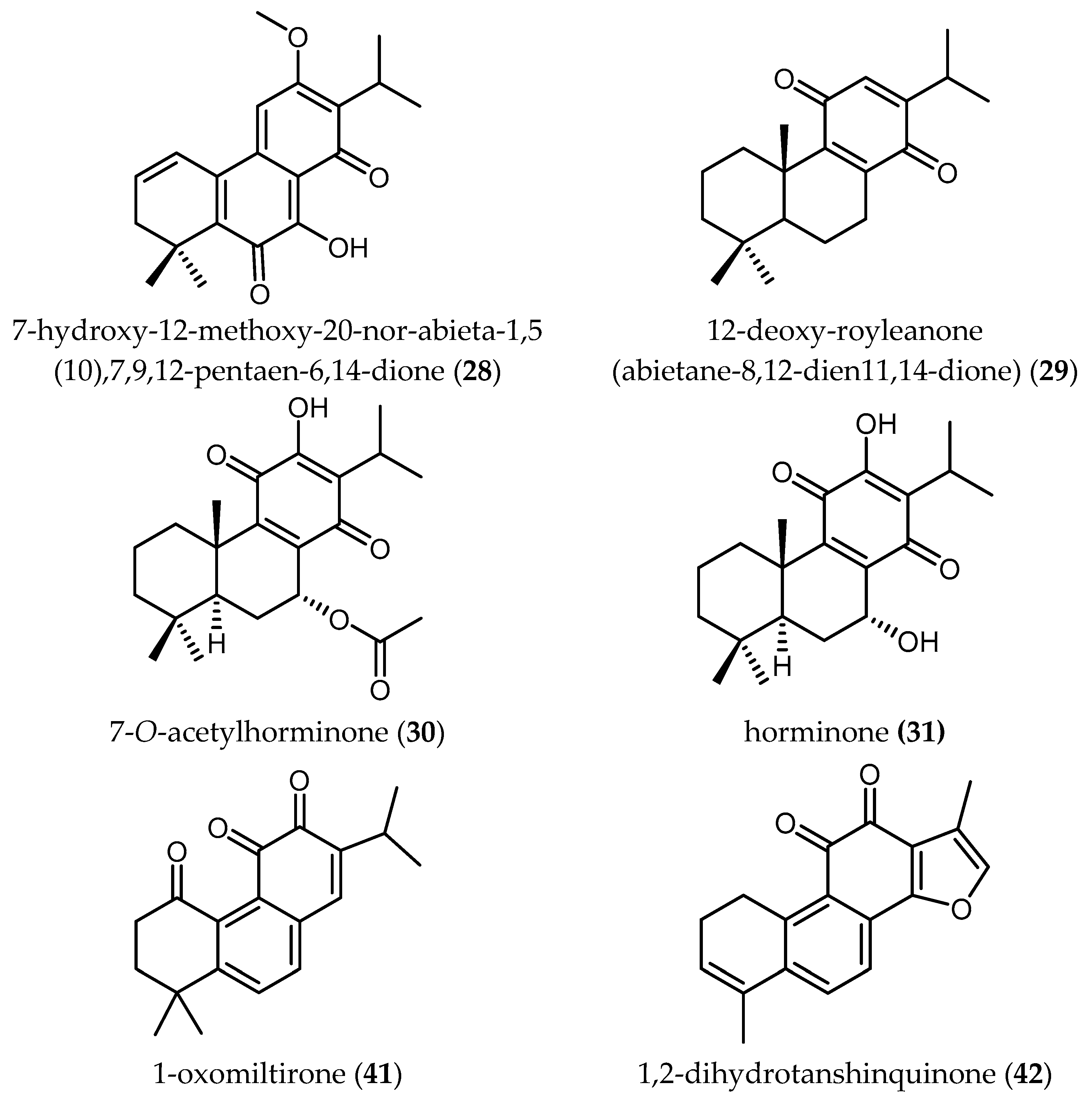
Two new abietane diterpenes derivatives isolated from the acetone extract of the roots of Salvia cilicica Boiss and Kotschy, 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione (28) and abieta-8,12-dien-11,14-dione (12-deoxy-royleanone) (29), were tested against promastigotes and intracellular amastigotes of both Ld and Lm [38]. Both diterpenes were shown to be more active against the clinically more relevant amastigote stages and displayed better antileishmanial activity against Ld, with IC50 values of 0.17 and 0.121 µM, respectively. The activity of 28 and 29 against Lm was lower with IC50 values of 0.287 and 0.18 µM, respectively. Unfortunately, the SI values did not show particularly selective toxicity against the parasites, since they ranged from 0.17 to 2.22.
Búfalo et al. isolated four diterpenes (15, 18, 30 and 31 in Table 6) from the roots of Salvia deserta Schang.—an Asian Salvia—and tested their activity against promastigotes of Ld [39]. The IC50 ranged from 1.46 ± 0.52 μM to 29.43 ± 3.01 μM. Taxodione (15), a diterpenoid quinone methide previously reported to yield promising activity against Tbr (see Section Diterpenoids), was the only compound with significant activity yielding an SI of 10.38, which could also make this compound interesting to be considered a good starting point for further development of novel antileishmanials.
3.2.3. Triterpenes
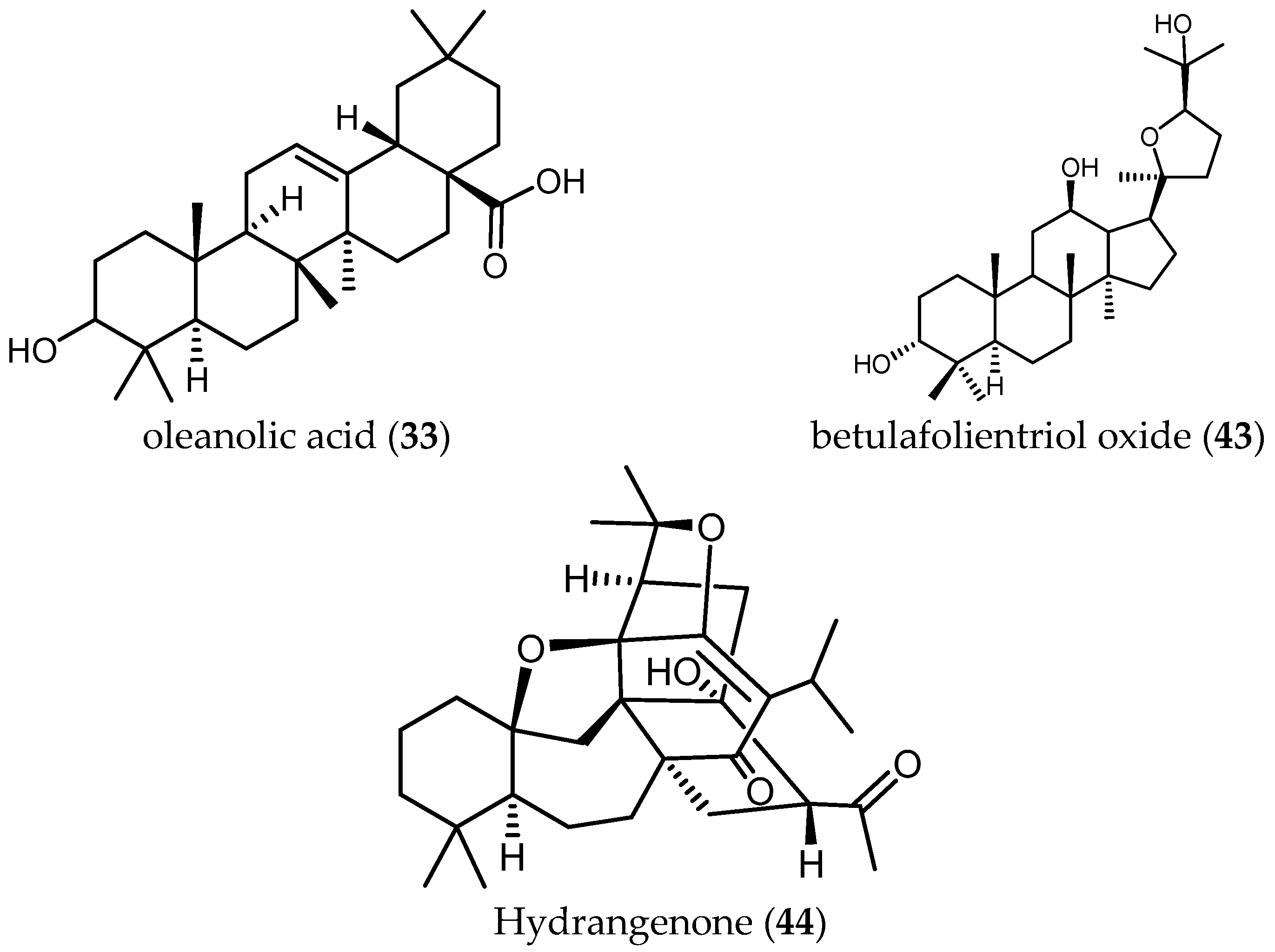
Tan and colleagues tested the antileishmanial activity of two triterpenes, ursolic acid (32) and oleanolic acid (33), isolated from S. cilicica Boiss and Kotschy [38]. They showed to be more potently active against amastigote forms of Ld than Lm, with IC50 values of 0.013 and 0.063 μM against Ld and IC50 values of 0.007 and 0.12 μM in the case of Lm. The ratio of cytotoxic over antileishmanial activity was unfavorable for both compounds with SI values ranging from 0.3 to 2.22. These ubiquitous triterpenes have been reported many times to possess anti-protozoal activity [18,29,40]. Since the level of activity and selectivity is not very high, their usefulness as anti-protozoal lead structures is questionable. Some chemical changes might be an option to improve their selectivity towards parasites.
3.2.4. Phenolic Compounds
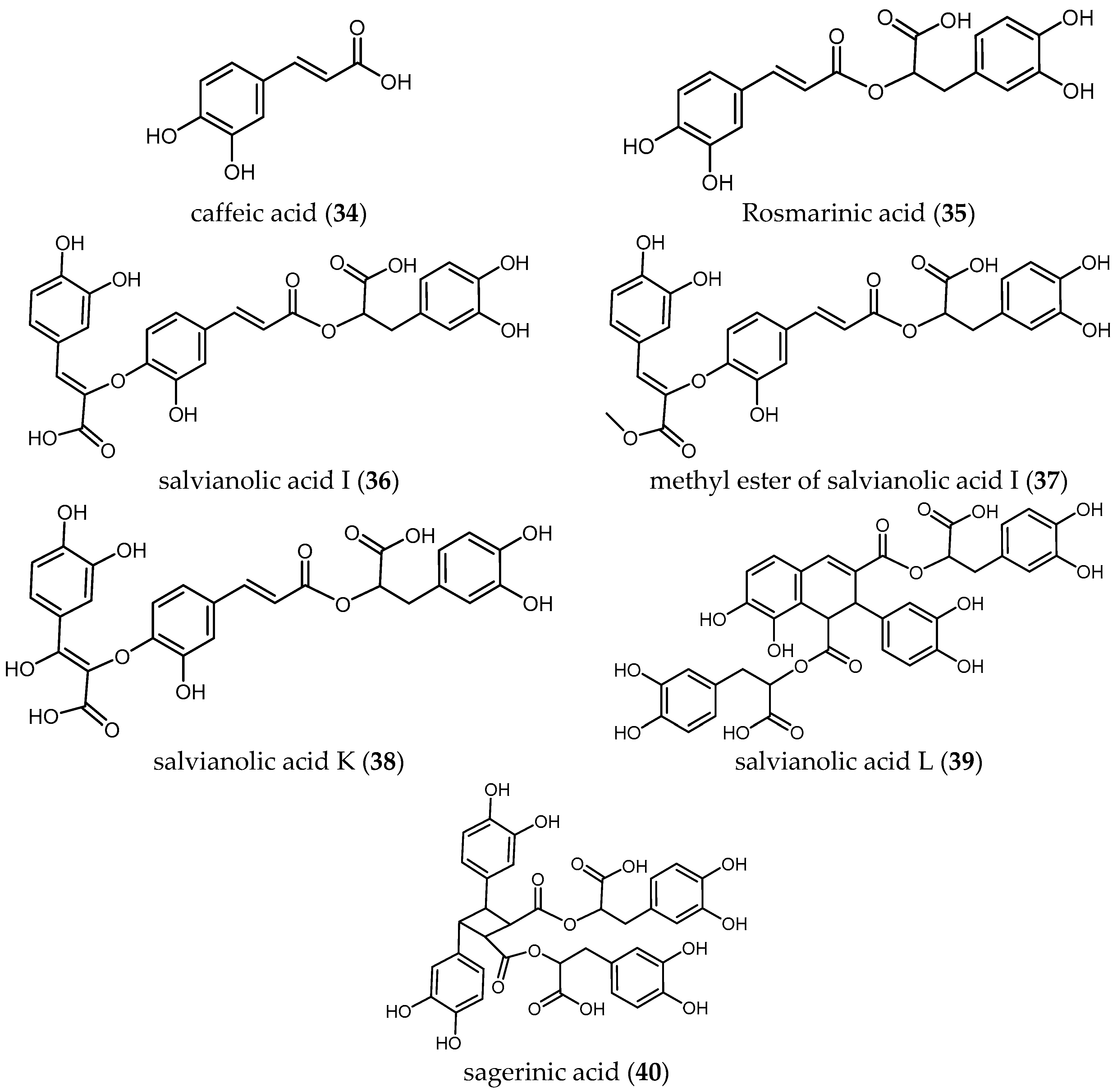
To best of our knowledge, less has been studied about the antiprotozoal activity of phenolic compounds of Salvia species. Radtke et al. isolated seven phenolic compounds (34–40) from S. officinalis L. and tested them against the amastigote and promastigote forms of several Leishmania species (Ld, Lm, Lg and Lk) [41]. None of the phenolic compounds showed selective activity when tested against the promastigote stages of the four Leishmania species (IC50 ranging from 0.70 to 2.4 μM) compared with the high and selective activities observed against the intracellular amastigote stages (IC50 values ranging from 0.004 to 0.176 μM) (Table 6).
Caffeic acid (34), the monomer unit of all tested phenolic compounds, was the most active compound against all tested parasites. This may indicate that the size of the molecule could be an important criterion responsible of the antileishmanial activity.
Concerning the two species of Leishmania with higher morbidity and mortality, the IC50 values of the phenolic compounds against Lm, an etiological agent of cutaneous leishmaniasis, ranged from 4.4 to 160.4 nM. Compound 34 exhibits the higher activity, followed by the mehtyl ester of salvianolic acid I (37), salvianolic acid K (38) and salvianolic acid L (39) with IC50 values of 0.0108, 0.0183 and 0.0203 μM, respectively. Interestingly, salvianolic acid I (36) showed sixteen-fold lower activity (IC50 = 0.1604 μM) than its methyl ester derivative (37), which again confirms that small modifications in molecules can represent a huge difference in terms of bioactivity. Further studies with methylated esters of 38–40 could be an interesting starting point for antileishmanial compounds.
Concerning the antileishmanial activity against Ld, etiological agent of visceral leishmaniasis, compound 34 exhibited the highest activity (IC50 = 0.0061 μM and favorable SI > 360), followed by salvianolic acid L (39) and salvianolic acid K (38) with IC50 values of 0.0154 and 0.0182 μM, respectively. Again, the methyl ester derivative (37), displayed significantly higher antileishmanial activity (IC50 0.0185 μM) than compound (36) (IC50 = 0.1758 μM). Compound 37 also showed higher selectivity against the parasite than compound 36 (SI > 37.63 and 3.9, respectively).
Regarding the activity of the compounds against amastigote forms of Lk and Lg—two causative agents of cutaneous leishmaniasis predominantly in the Tunisian desert [42] and in French Guiana [43], respectively—caffeic acid (34) was the most active compound yielding the higher selective activity against Lk and Lg with IC50 values of 0.0039 and 0.0066 μM, respectively.
These results suggested the potential of 34 and its derivatives as promising compounds to be used as antileishmanials based on their high biological activity and the absence of toxicity against the host cells.
3.3. Antiplasmodial Activity
Malaria is a disease caused by different species of the genus Plasmodium. This disease is endemic in 91 countries and almost half of the world’s population is at risk of it [44], which means a dramatic global health issue. The rate of new cases fell by 37% globally during the period 2000–2015; however, the WHO still estimates that 214 million of cases of malaria and 438,000 deaths were reported in 2015 [44]. Although it does not belong to WHO’s list of NTDs, it represents a neglected disease in many parts of the world, where populations live in Malaria-infested regions under poor socio-economic conditions with no access to health services and adequate medication.
Malaria is a curable and preventable disease and the use of insecticide-treated mosquito nets is an essential and effective means to reduce the transmission and new infections. However, because of the increasing resistance of the parasites against the current therapies and the resistance of the mosquito against the available insecticides, the need of development of new therapies remains urgent.
The majority of results of research on antiplasmodial activity of Salvia species were obtained in studies with Plasmodium falciparum, the most lethal parasite responsible for Malaria tropica. No results on activity against other species of Plasmodium were found in the literature (Table 7 and Table 8).
3.3.1. Extracts and Essential Oils
A series of seventeen extracts obtained with a mixture chloroform and methanol 1:1 (v/v) and eleven essential oils from seventeen different indigenous South African species of Salvia have been tested against the chloroquine-resistant Pf FCR-3 strain (Table 7) [45]. The IC50 values of the essential oils and extracts ranged from 1.2 to 13.50 µg/mL and 3.91 to 25.38 µg/mL, respectively. Most of the essential oils displayed better bioactivity than the solvent-extracts of the respective species. In this respect, the essential oil from S. runcinata L.f. (v2) and the chloroform-methanol extract of S. radula Benth (z1) represent the samples with the best biological activity (Table 7) [45].
Futhermore, S. repens Burch. ex Benth. and its extracts obtained with solvents of different polarity (named as f–f5 in Table 7) have been extensively studied by different research groups. Kamatou and coworkers reported on the IC50 of a methanol extract of this plant with IC50 = 78.9 µg/mL [6]. Moreover, IC50 values for the chloroform-methanol (1:1) extract and also the essential oil of this species against chloroquine-sensitive Pf strain D10 were reported as 8.25 and 1.65 µg/mL, respectively [45]. Clarkson and coworkers tested the dichloromethane-methanol (1:1) extract of the whole plant against the chloroquine-sensitive strain (D10) of Pf resulting in an IC50 of 10.8 µg/mL [46]. The same extract was tested by Mokoka and colleagues against the chloroquine- and pyrimethamine-resistant K1 strain of Pf resulting IC50 values of 7.65 µg/mL and SI = 5.4 [6]. The results of the extracts thus indicated some significant differences.
Extracts of different polarity from three Salvia species were tested by Kirmizibekmez and coworkers against the resistant K1 strain and compared with the standard drug chloroquine [16]. The chloroform extracts of the three selected plants displayed the best IC50 values ranging from 2.54 to 3.72 µg/mL, the chloroform extract of S. sclarea L. (b3) being the most active one (Table 7).
Quite recently, Hammoud and coworkers tested different solvent extract of Salvia chudaei Batt. & Trab. and its essential oil against chloroquine resistance and sensitive Pf strains [47]. The essential oil (Aa3) showed the highest activity against both chloroquine resistance and sensitive parasites with IC50 of 2.39 ± 0.24, 2.40 ± 0.77 µg/mL, respectively, followed by the n-hexane extract (Aa) (IC50 = 4.91 ± 2.91 µg/mL against Pf K1 strain) (Table 7). No data about toxicity in mammalian cells were reported.
More studies to identify the compounds most probably responsible for the biological activity of these extracts are still required.
3.3.2. Diterpenoids
Several Salvia diterpenoids have been reported to possess antiplasmodial activity. Diterpenes 10–17, which also were tested against Tbr (see Section Diterpenoids) showed antiplasmodial activity with IC50 values ranging from 0.9 to 17.8 µM against Pf K1 with SI ranging from 0.1 to 18.2 [23] (Table 8). The mostly unfavorable values of selectivity call into question the security of most of these compounds and their use as lead compounds for new drugs. Only two compounds, Δ9-ferruginol (14) and ferruginol (15), were shown to be active and selective against Pf K1 strain. Both displayed IC50 values of 0.9 µM and SI values of 18.2 and 15.6, respectively, and may thus be considered promising hits [23]. Compound 15 was also isolated from the roots of S. deserta Schang. and tested against D6 chloroquine sensitive and W2 chloroquine resistance strains of Pf yielding IC50 values of 5.64 and 6.35 μM, respectively [39]. In this study, the ratio of cytotoxicity of Compound 15 against VERO cells over antiplasmodial activity was unfavorable with SI values of 0.8 and 0.7, respectively. This results contrasts with the results published by Ebrahimi et al. [23] (Table 8).
Ślusarczyk and colleagues isolated nine compounds from the dichloromethane extract of S. miltiorrhiza Bunge roots and identified them as tanshinone and abietane type diterpenoids (Compounds 3–9 and 41–42 in Table 8) [4]. They were tested against the chloroquine resistant K1 strain of Pf. The total extract showed 34% of growth inhibition at 0.81 µg/mL, while the isolated compounds displayed IC50 values ranging from 0.5 to ≥30 µM. Moreover, the SI values of these compounds were unfavorable, ranging from only 0.3 to 2.6. The relatively high activity of some of these compounds has been correlated with the presence of a furan D ring in their structure [4]. More investigations to relate structural elements with the activity and toxicity are necessary.
Another diterpene, betulafolientriol oxide (43), isolated from an active extract of Salvia radula Benth. (IC50 = 3.91 µg/mL, see z1 in Table 7), showd antiplasmodial activity comparable with that of the crude extract, displaying an IC50 value of 10.39 μM against the FCR3 strain of Pf and a SI > 20, tested on kidney human cells [48].
In the same study cited above with respect to Tbr and Tc [24], Kuźma and coworkers tested in vitro the antiplasmodial activity of the four quinone methide-type diterpenes (18–21 in Table 8) isolated from the roots of S. austriaca Jacq. against erythrocytic stages of Pf NF54. The IC50 values of the tested compound ranged from 1.9 µM to up to 167 µM, with taxodione (18) once more representing the most active compound. However, the SI of 18 was not favorable (SI = 1). In contrast, the antiplasmodial activity of Compound 18—isolated from the roots of S. deserta Schang—was also determined against chloroquine sensitive (D6) and chloroquine resistant (W2) strains of Pf showing higher IC50 values (10.49 μM and 9.66 μM 3, respectively) [39].
Kuźma et al. also reported interesting biological activities of the diterpenes 7-(2′-oxohexyl)-taxodione (20) and taxadone (21), which demonstrated antiplasmodial activity at IC50 values of 3.37 and 3.66 µM, respectively. However, less promising was the antiplasmodial activity of 7-O-acetylhorminone (30) and horminone (31), for which the IC50 values were shown to be higher than 12.72 μM [39].
In view of the presented data it can be concluded that diterpenoids showed some favorable activity results but also present some toxicity to mammalian cells, which renders their direct usefulness in unmodified form as antimalarials unlikely. However, it might be useful to conduct further investigations, such as quantitative structure-activity relationship (QSAR) analysis with series of chemically modified compounds of this type, which could help to determine which characteristics and substituents of the compounds are most important to increase the antiplasmodial activity and their selectivity. A progress in this direction might make it possible to improve the properties of such natural products with respect to a more favorable activity/selectivity ratio.
3.3.3. Triterpenes and Related Terpenoids
The two triterpenoids isolated from S. hydrangea DC. ex Benth., salvadione C (22) and perovskone B (23), were not only reported to show antitrypanosomal effects (see Section Triterpenoids) but also shown to be antiplasmodial agents (Table 8). The n-hexane extract of the plant (d) displayed an IC50 value of 3.2 µg/mL (Table 7) against Pf and the two isolated triterpenoids, 22 and 23, were reported to have antiplasmodial activity with IC50 values of 1.43 and 0.18 µM, and SI values of 86.2 and 69.6, respectively [25]. Both the high bioactivity and the favorable SI values indicate that they can be considered as promising lead compounds to be further investigated. More research in terms of in vivo-activity and bioavailability as well as their mechanism of action should be performed.
The common triterpene oleanolic acid (33) (Table 8)—also isolated from the extract of S. hydrangea DC. ex Benth. (IC50 < 12.5 µg/mL)—displayed much lower activity with an IC50 value of 19.3 µM when tested against the 3D7 strain of Pf [8].
Farimani and colleagues also isolated from S. hydrangea DC. ex Benth. the unusual isoprenoid hydrangenone (44), which showed antiplasmodial activity with IC50 = 1.4 µM and a favorable SI = 6 [49].
3.3.4. Flavonoids
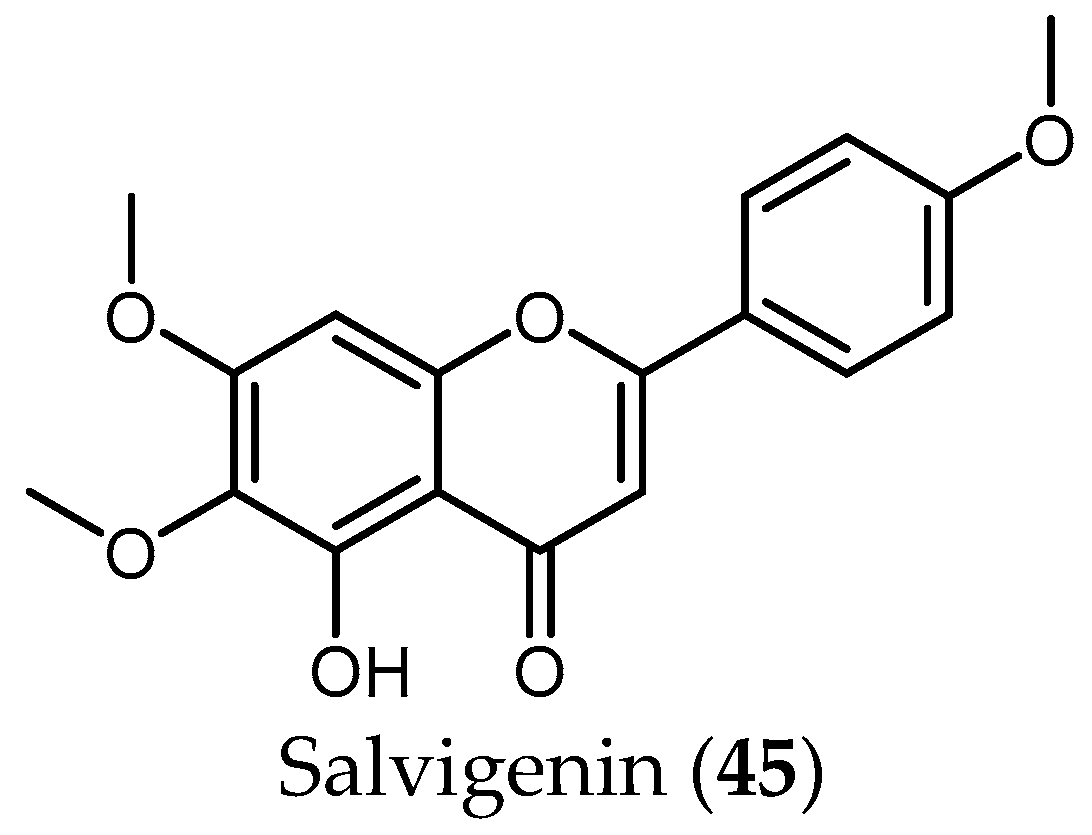
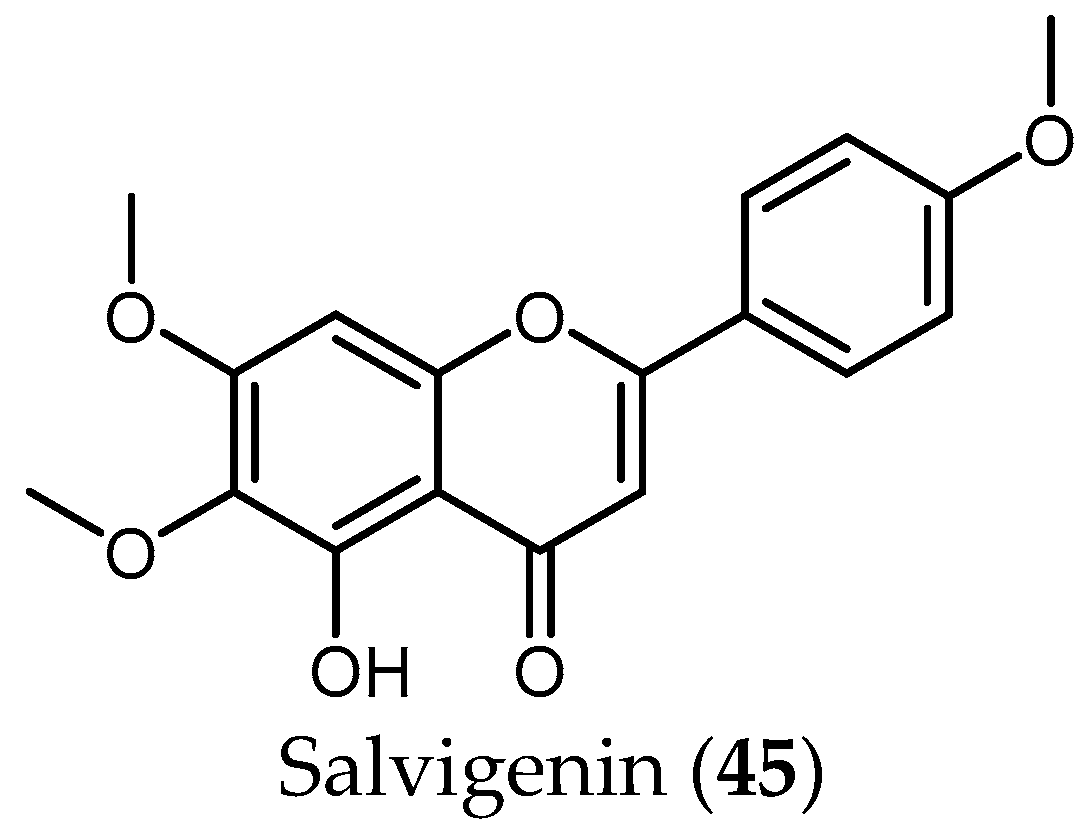
Although the antiprotozoal activity of some flavonoids has been reported by different authors [40,50], little has been published in the case of Salvia. To the best of the knowledge, only the flavonoid salvigenin (45), extracted from the active extract from S. radula Benth. (IC50 = 3.91 µg/mL), was tested against Pf, yielding a low antiplasmodial activity (IC50 value 74.98 μM and SI > 4) [48].
4. Conclusions
The present review has summarized the current literature knowledge on the antiprotozoal activity of crude extracts from various Salvia species and of a variety of isolated compounds.
Relatively little has been reported on the traditional use of Salvia against the parasites under study. Thirty-five species of Salvia—or compounds islated from them—have been tested against at least one of these pathogens. Antiplasmodial activity was most frequently reported and antitrypanosomal activity against T. cruzi was the least well studied. However, the latter was the only parasite for which in vivo studies have been performed with a compound from Salvia.
From the 35 species, 45 compounds were isolated and have been tested against the protozoans treated in this review. Diterpenes of the abietane group represent the most active compounds against T. brucei and Pf, whereas phenolic compounds seem to show higher activity against Leishmania spp. The only flavonoid isolated so far from Salvia species—salvigenin (45)—and tested against protozoa did not show any promising results.
When inspecting the data summarized here, the bioactivity of the various extracts and compounds against the different parasites under study vary over a rather wide range and it is not trivial to make a judgement which level of activity should be considered strong or which entity could be promising for further studies. Naturally, the view on this issue of authors publishing such activity of their compounds will vary almost as widely as the data reported. Generally, from the authors’ own experience, T. brucei appears to be the most sensitive of the parasites treated here and T. cruzi, in particular if the clinically most relevant intracellular amastigote stage is assayed, is the hardest to kill in vitro. For a rough orientation, the IC50 values of standard positive controls routinely used in many studies, such as those conducted by our group in collaboration with the Swiss Tropical and Public Health Institute (Basel, Switzerland), are typically in the following ranges of magnitude: melarsoprol (Tbr, STIB 9000 strain, bloodstream trypomastigotes): 0.01 µM; benznidazole (Tc, Tulahuen C4 strain, intracellular amastigotes in L6 rat skeletal myoblasts): 4.5 µM; miltefosine (Ld MHOM-ET-67/L82, axenic amastigotes): 0.2 µM; and chloroquine (Pf, NF54 strain, intraerythrocytic forms): 0.02 µM (see e.g., [21]; data reported there are in µg/mL). It would appear overly critical, however, to concentrate only on such entities that outmatch these positive controls’ activity because all of them are clinically used drugs and the native natural products may only represent starting points for further optimization. Thus, in our own work, we would consider any isolated natural product that displays an IC50 value <0.1, <10, <2 and <1 µM against the mentioned forms of Tbr, Tc, Ld and Pf, respectively, a valuable hit for further studies, in case it also displays a favorable selectivity index (determined with L6 cells), e.g., >5–10.
Some of the reported compounds showed interesting in vitro biological activity, which could be considered a starting point for further investigations, such as structure-activity, mechanistic and in vivo studies. There appears to be a lack of further research-steps beyond the stage of in vitro phenotypic assays to exploit the full potential of these compounds in terms of the development of lead compounds into drugs. This may be due to the lack of funding within the current R&D system which is mainly guided by economical interest and often reluctant to support research in this field, which makes it difficult for many academic research groups to advance promising hits to further development steps. Most information on native natural compounds with promising activity that might be worth optimization is published at the stage of in vitro tests and then tends to be dissipated in the literature. The compounds often “vanish” in refrigerators in many laboratories around the globe but this valuable knowledge and material is not further followed up in a systematic way [7,40]. Further research steps such as a thorough investigation of structure-activity and structure-selectivity relationships, studies on the (possibly new) mechanisms of action and studies on the in vivo fate and efficacy of promising compounds often never happen. The information on Salvia metabolites and their antiprotozoal activity collected in this review will be useful for further studies on related species and their chemical constituents in the quest for new effective and affordable drugs against NTDs. A review such as this can help to avoid repetition of research, e.g., if it is used as a guide to dereplicate the pattern of constituents in further, hitherto unstudied Salvia species. Most importantly, however, it would be useful to stimulate further research on structure-activity relationship (SAR) studies for compounds of the classes treated here.
Acknowledgments
Núria Llurba-Montesino receives financial support from Heinrich Böll Fundation in the form of a promotion fellowship which is most gratefully acknowledged. We acknowledge support by Open Access Publication Fund of University of Muenster.
Author Contributions
Núria Llurba-Montesino performed the literature research and prepared the manuscript together with Thomas J. Schmidt who supervised the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Topçu, G. Bioactive triterpenoids from Salvia species. J. Nat. Prod. 2006, 69, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.B.; Sytsma, K.J. Staminal evolution in the genus Salvia (Lamiaceae): Molecular phylogenetic evidence for multiple origins of the staminal lever. Ann. Bot. 2007, 100, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Goñalons, L. Salvia L. In Flora Ibérica; Morales, R., Quintanar, A., Cabezas, F., Pujadas, A.J., Cirujano, S., Eds.; Real Jardín Botánico, CSIC: Madrid, Sapin, 1974; Volume 12, pp. 1194–1196. [Google Scholar]
- Ślusarczyk, S.; Zimmermann, S.; Kaiser, M.; Matkowski, A.; Hamburger, M.; Adams, M. Antiplasmodial and antitrypanosomal activity of tanshinone-type diterpenoids from Salvia miltiorrhiza. Planta Med. 2011, 77, 1594–1596. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.B.; Ni, Z.Y.; Shi, Q.W.; Dong, M.; Kiyota, H.; Gu, Y.C.; Cong, B. Constituents from Salvia species and their biological activities. Chem. Rev. 2012, 112, 5967–6026. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.P.; Viljoen, A.M.; Gono-Bwalya, A.B.; van Zyl, R.L.; van Vuuren, S.F.; Lourens, A.C.U.; Başer, K.H.C.; Demirci, B.; Lindsey, K.L.; van Staden, J.; et al. The in vitro pharmacological activities and a chemical investigation of three South African Salvia species. J. Ethnopharmacol. 2005, 102, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.; Biavatti, M.W.; Brun, R.; da Costa, B.D.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; Lago, J.H.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads Against Protozoan Neglected Diseases—Part I. Curr. Med. Chem. 2012, 19, 2128–2175. [Google Scholar] [CrossRef] [PubMed]
- Sairafianpour, M.; Bahreininejad, B.; Witt, M.; Ziegler, H.L.; Jaroszewski, J.W.; Stærk, D. Terpenoids of Salvia hydrangea: Two New, Rearranged 20-Norabietanes and the Effect of Oleanolic Acid on Erythrocyte Membranes. Planta Med. 2003, 69, 846–850. [Google Scholar] [CrossRef] [PubMed]
- WHO Press. Research priorities for Chagas Disease, Human African Trypanosomiasis and Leishmaniasis. World Health Organ. Tech. Rep. Ser. 2012, 975, 1–100. [Google Scholar]
- WHO. Health Statistics and Information Systems, Estimates for 2000–2015. Available online: http://www.who.int/healthinfo/global_burden_disease/estimates/en/index2.html (accessed on 1 July 2017).
- WHO. Human African Trypanosomiasis. Available online: http://www.who.int/trypanosomiasis_african/en/ (accessed on 1 December 2017).
- Mesu, V.K.B.K.; Kalonji, W.M.; Bardonneau, C.; Mordt, O.V.; Blesson, S.; Simon, F.; Delhomme, S.; Bernhard, S.; Kuziena, W.; Lubaki, J.-P.F.; et al. Oral fexinidazole for late-stage African Trypanosoma brucei gambiense trypanosomiasis: A pivotal multicentre, randomised, non-inferiority trial. Lancet 2017, 6736, 1–11. [Google Scholar] [CrossRef]
- Pinazo, M.J.; Gascon, J. The importance of the multidisciplinary approach to deal with the new epidemiological scenario of Chagas disease (global health). Acta Trop. 2015, 151, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gascon, J.; Bern, C.; Pinazo, M.J. Chagas disease in Spain, the United States and other non-endemic countries. Acta Trop. 2010, 115, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Info Chagas. Available online: http://www.infochagas.org/en/ (accessed on 13 April 2017).
- Kirmizibekmez, H.; Atay, I.; Kaiser, M.; Yesiladaa, E.; Tasdemir, D. In vitro antiprotozoal activity of extracts of five Turkish Lamiaceae species. Nat. Prod. Commun. 2011, 6, 1697–1700. [Google Scholar]
- Farimani, M.M.; Ebrahimi, S.N.; Salehi, P.; Bahadori, M.B.; Sonboli, A.; Khavasi, H.R.; Zimmermann, S.; Kaiser, M.; Hamburger, M. Antitrypanosomal triterpenoid with an ε-lactone E-ring from Salvia urmiensis. J. Nat. Prod. 2013, 76, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Mokoka, T.A.; Peter, X.K.; Fouche, G.; Moodley, N.; Adams, M.; Hamburger, M.; Kaiser, M.; Brun, R.; Maharaj, V.; Koorbanally, N. Antileishmanial activity of 12-methoxycarnosic acid from Salvia repens Burch. ex Benth. (Lamiaceae). S. Afr. J. Bot. 2014, 90, 93–95. [Google Scholar] [CrossRef]
- Jain, S.; Jacob, M.; Walker, L.; Tekwani, B. Screening North American plant extracts in vitro against Trypanosoma brucei for discovery of new antitrypanosomal drug leads. BMC Complement. Altern. Med. 2016, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Ihsan, S.A.; Mei, W.; Ali, Z.; Zaki, A.A.; Khan, S.I.; Khan, I.A. Chemical Analysis and Biological Activities of Salvia lavandulifolia Vahl. Essential Oil. Chem. Anal. 2017, 7, 71–78. [Google Scholar]
- Llurba-Montesino, N.; Kaiser, M.; Brun, R.; Schmidt, T.J. Search for Antiprotozoal Activity in Herbal Medicinal Preparations; New Natural Leads against Neglected Tropical Diseases. Molecules 2015, 20, 14118–14138. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, Y.; Fujiwara, K.; Yamazaki, A.; Sugawara, N.; Yano, R.; Fukaya, H.; Hitotsuyanagi, Y.; Takeya, K.; Ishiyama, A.; Iwatsuki, M.; et al. Semisynthesis of salviandulin e analogues and their antitrypanosomal activity. Bioorg. Med. Chem. Lett. 2014, 24, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.N.; Zimmermann, S.; Zaugg, J.; Smiesko, M.; Brun, R.; Hamburger, M. Abietane diterpenoids from Salvia sahendica—Antiprotozoal activity and determination of their absolute configurations. Planta Med. 2013, 79, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Kuźma, Ł.; Kaiser, M.; Wysokińska, H. The production and antiprotozoal activity of abietane diterpenes in Salvia austriaca hairy roots grown in shake flasks and bioreactor. Prep. Biochem. Biotechnol. 2017, 47, 58–66. [Google Scholar] [CrossRef]
- Farimani, M.M.; Bahadori, M.B.; Taheri, S.; Ebrahimi, S.N.; Zimmermann, S.; Brun, R.; Amin, G.; Hamburger, M. Triterpenoids with rare carbon skeletons from Salvia hydrangea: Antiprotozoal activity and absolute configurations. J. Nat. Prod. 2011, 74, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Mokoka, T.A.; Zimmermann, S.; Julianti, T.; Hata, Y.; Moodley, N.; Cal, M.; Adams, M.; Kaiser, M.; Brun, R.; Koorbanally, N.; et al. In vitro screening of traditional South African malaria remedies against Trypanosoma brucei rhodesiense, Trypanosoma cruzi, Leishmania donovani, and Plasmodium falciparum. Planta Med. 2011, 77, 1663–1667. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Jimenez-Ortiz, V.; Sartor, T.; Tonn, C.E.; García, E.E.; Nieto, M.; Burgos, M.H.; Sosa, M.A. A novel icetexane diterpene, 5-epi-icetexone from Salvia gilliessi is active against Trypanosoma cruzi. Acta Trop. 2006, 98, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Lozano, E.; Barrera, P.; Salinas, R.; Vega, I.; Nieto, M.; Tonn, C.; Kemmerling, U.; Mortara, R.A.; Sosa, M.A. Sesquiterpene lactones and the diterpene 5-epi-icetexone affect the intracellular and extracellular stages of Trypanosoma cruzi. Parasitol. Int. 2012, 61, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Lozano, E.; Strauss, M.; Spina, R.; Cifuente, D.; Tonn, C.; Rivarola, H.W.; Sosa, M.A. The in vivo trypanocidal effect of the diterpene 5-epi-icetexone obtained from Salvia gilliesii. Parasitol. Int. 2016, 65, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Lozano, E.S.; Spina, R.M.; Tonn, C.E.; Sosa, M.A.; Cifuente, D.A. An abietane diterpene from Salvia cuspidata and some new derivatives are active against Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2015, 25, 5481–5484. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Investing to Overcome the Global Impact of Neglected Tropical Diseases. Third WHO Report on Neglected Tropical Diseases; Holmes, P., Ed.; WHO Press: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. WHO technical report series: Control of the leishmaniases. In WHO Technical Report Series; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789241209. [Google Scholar]
- Singh, N.; Mishra, B.B.; Bajpai, S.; Singh, R.K.; Tiwari, V.K. Natural product based leads to fight against leishmaniasis. Bioorg. Med. Chem. 2014, 22, 18–45. [Google Scholar] [CrossRef] [PubMed]
- Essid, R.; Zahra, F.; Msaada, K.; Sghair, I.; Hammami, M.; Bouratbine, A.; Aoun, K.; Limam, F. Antileishmanial and cytotoxic potential of essential oils from medicinal plants in Northern Tunisia. Ind. Crops Prod. 2015, 77, 795–802. [Google Scholar] [CrossRef]
- Nikmehr, B.; Ghaznavi, H.; Rahbar, A.; Sadr, S.; Mehrzadi, S. In vitro anti-leishmanial activity of methanolic extracts of Calendula officinalis flowers, Datura stramonium seeds, and Salvia officinalis leaves. Chin. J. Nat. Med. 2014, 12, 423–427. [Google Scholar] [CrossRef]
- Khan, A.R.; Khan, M.J. In vitro Antileishmanial, Cytotoxic and Antioxidant activities of Salvia bucharica leaves extract and its fractions. Int. J. Basic Appl. Sci. 2003, 13, 74–78. [Google Scholar]
- Et-Touys, A.; Fellah, H.; Sebti, F.; Mniouil, M.; Aneb, M.; Elboury, H.; Talbaoui, A.; Dakka, N.; Sadak, A.; Bakri, Y. In vitro Antileishmanial Activity of Extracts from Endemic Moroccan Medicinal Plant Salvia verbenaca (L.) Briq. Ssp. verbenaca Maire (S. clandestina Batt. non L). Eur. J. Med. Plants 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Tan, N.; Kaloga, M.; Radtke, O.A.; Kiderlen, A.F.; Öksüz, S.; Ulubelen, A.; Kolodziej, H. Abietane diterpenoids and triterpenoic acids from Salvia cilicica and their antileishmanial activities. Phytochemistry 2002, 61, 881–884. [Google Scholar] [CrossRef]
- Búfalo, J.; Cantrell, C.L.; Jacob, M.R.; Schrader, K.K.; Tekwani, B.L.; Kustova, T.S.; Ali, A.; Boaro, C.S.F. Antimicrobial and Antileishmanial Activities of Diterpenoids Isolated from the Roots of Salvia deserta. Planta Med. 2015, 82, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.; Biavatti, M.W.; Brun, R.; da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads Against Protozoan Neglected Diseases—Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Radtke, O.A.; Yeap Foo, L.; Lu, Y.; Kiderlen, A.F.; Kolodziej, H. Evaluation of sage phenolics for their antileishmanial activity and modulatory effects on interleukin-6, interferon and tumour necrosis factor-α-release in RAW 264.7 Cells. Z. Naturforsch. C 2003, 58, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Maubon, D.; Thurot-Guillou, C.; Ravel, C.; Leccia, M.-T.; Pelloux, H. Leishmania killicki imported from Tunisian desert. Emerg. Infect. Dis. 2009, 15, 1864–1865. [Google Scholar] [CrossRef] [PubMed]
- Rotureau, B.; Ravel, C.; Nacher, M.; Couppié, P.; Curtet, I.; Dedet, J.P.; Carme, B. Molecular epidemiology of Leishmania (Viannia) guyanensis in French Guiana. J. Clin. Microbiol. 2006, 44, 468–473. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Fact Sheet Malaria. Available online: http://www.who.int/mediacentre/factsheets/fs094/en/ (accessed on 1 July 2017).
- Kamatou, G.P.P. Indigenous Salvia Species—An Investigation of Their Pharmacological Activities and Phytochemistry. Ph.D. Thesis, University of the Witwatersrand, Johannesburg, South Africa, 2006. [Google Scholar]
- Clarkson, C.; Maharaj, V.J.; Crouch, N.R.; Grace, O.M.; Pillay, P.; Matsabisa, M.G.; Bhagwandin, N.; Smith, P.J.; Folb, P.I. In vitro antiplasmodial activity of medicinal plants native to or naturalised in South Africa. J. Ethnopharmacol. 2004, 92, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, R.; Dehak, K.; Sanon, S.; Mahammed, M.H.; Ouldelhadj, M.D. In Vitro Antimalarial, Antimicrobial and Antioxidants Activities of Salvia chudaei Batt. & Trab. (Lamiaceae) Extracts Roukia. Der Pharma Chem. 2017, 9, 82–89. [Google Scholar]
- Kamatou, G.P.P.; van Zyl, R.L.; Davids, H.; van Heerden, F.R.; Lourens, A.C.U.; Viljoen, A.M. Antimalarial and anticancer activities of selected South African Salvia species and isolated compounds from S. radula. S. Afr. J. Bot. 2008, 74, 238–243. [Google Scholar] [CrossRef]
- Farimani, M.M.; Taheri, S.; Ebrahimi, S.N.; Bahadori, M.B.; Khavasi, H.R.; Zimmermann, S.; Brun, R.; Hamburger, M. Hydrangenone, a new isoprenoid with an unprecedented skeleton from Salvia hydrangea. Org. Lett. 2012, 14, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Kaiser, M.; Brun, R.; Yardley, V.; Schmidt, T.J.; Tosun, F.; Rüedi, P. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: In vitro, in vivo, structure-activity relationship, and quantitative structure-activity relationship studies. Antimicrob. Agents Chemother. 2006, 50, 1352–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Diterpenoids of Salvia with antiprotozoal activity.
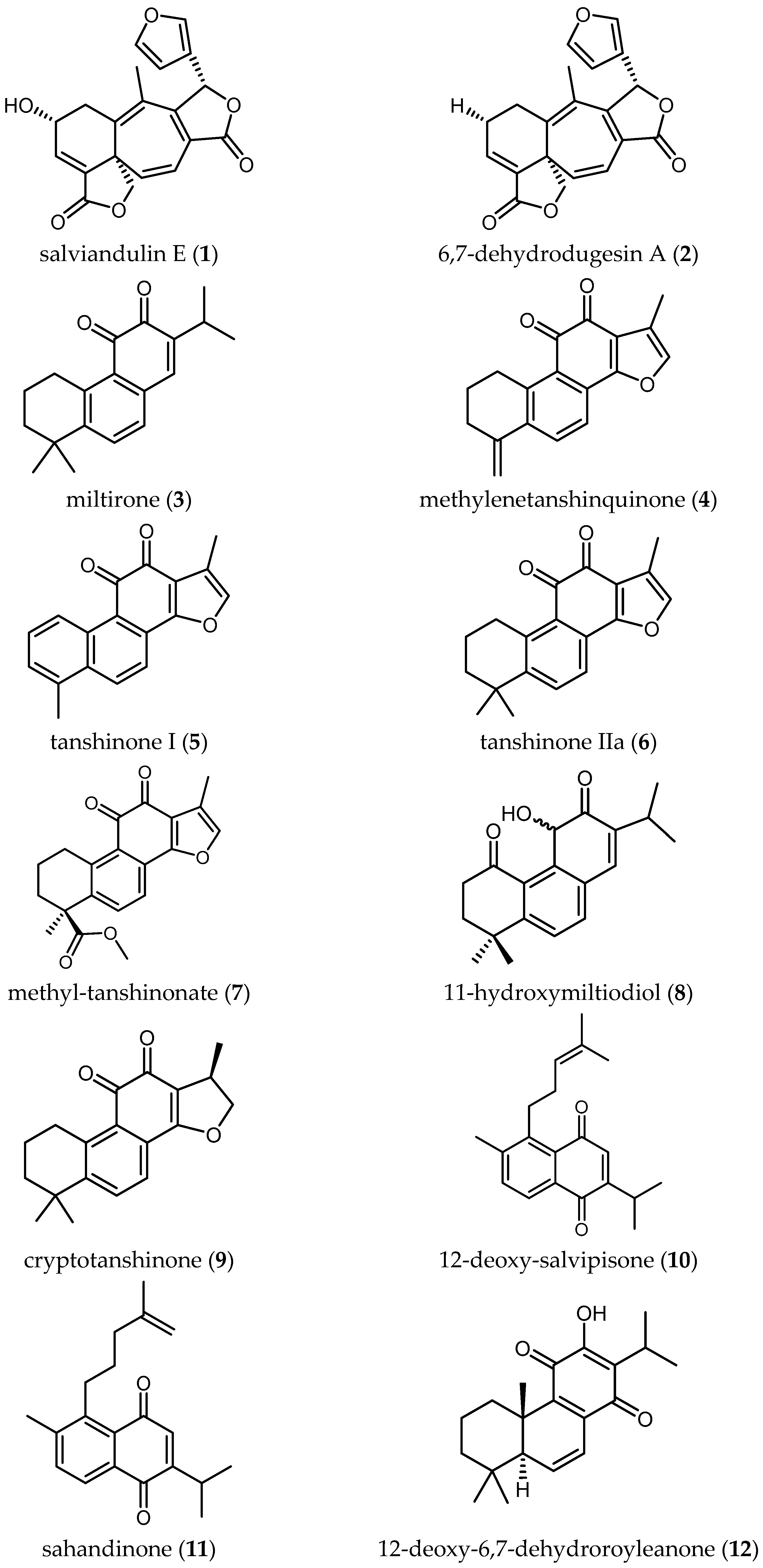

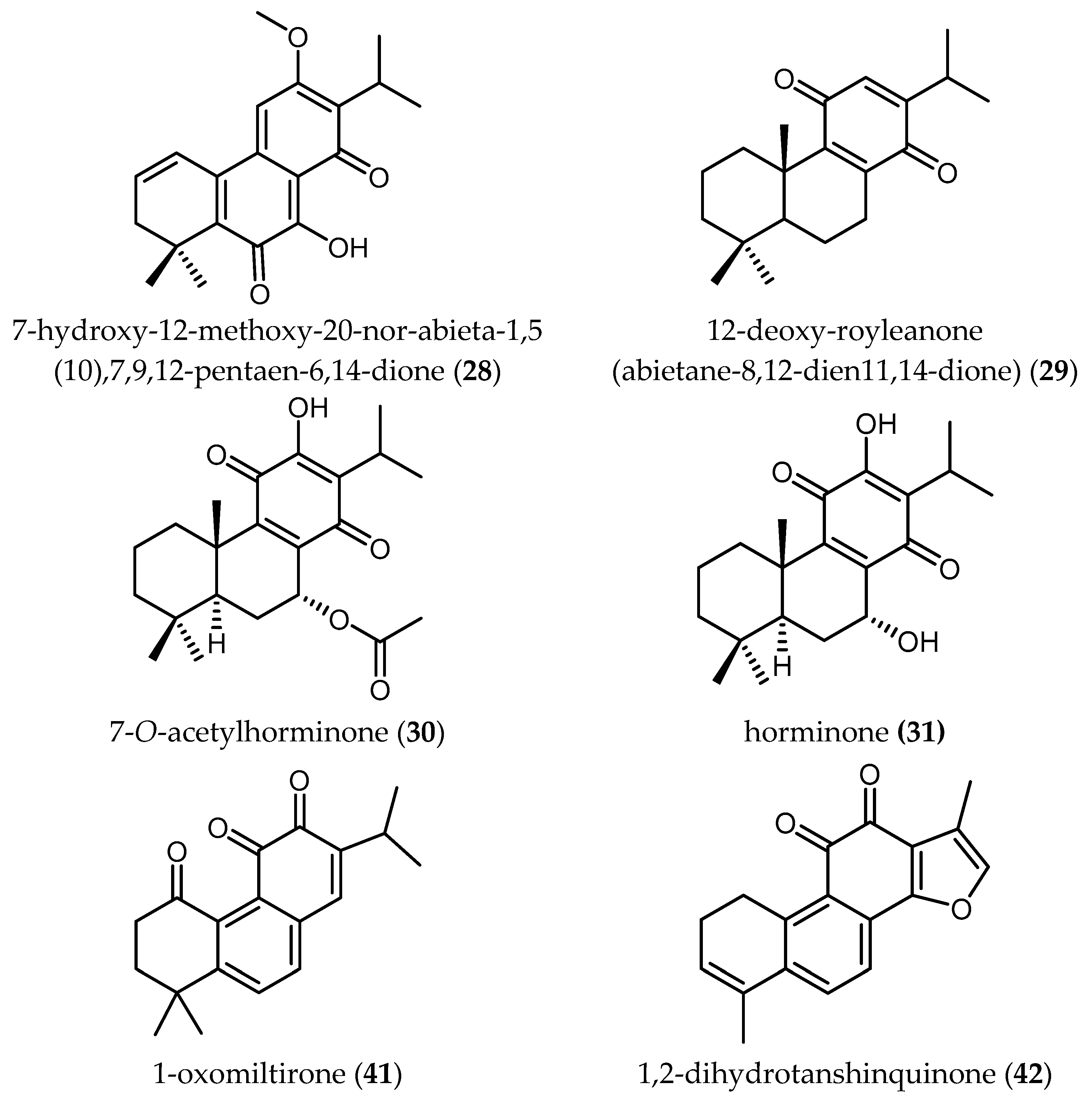
Figure 2.
Triterpenes and related terpenoids of Salvia with antiprotozoal activity.
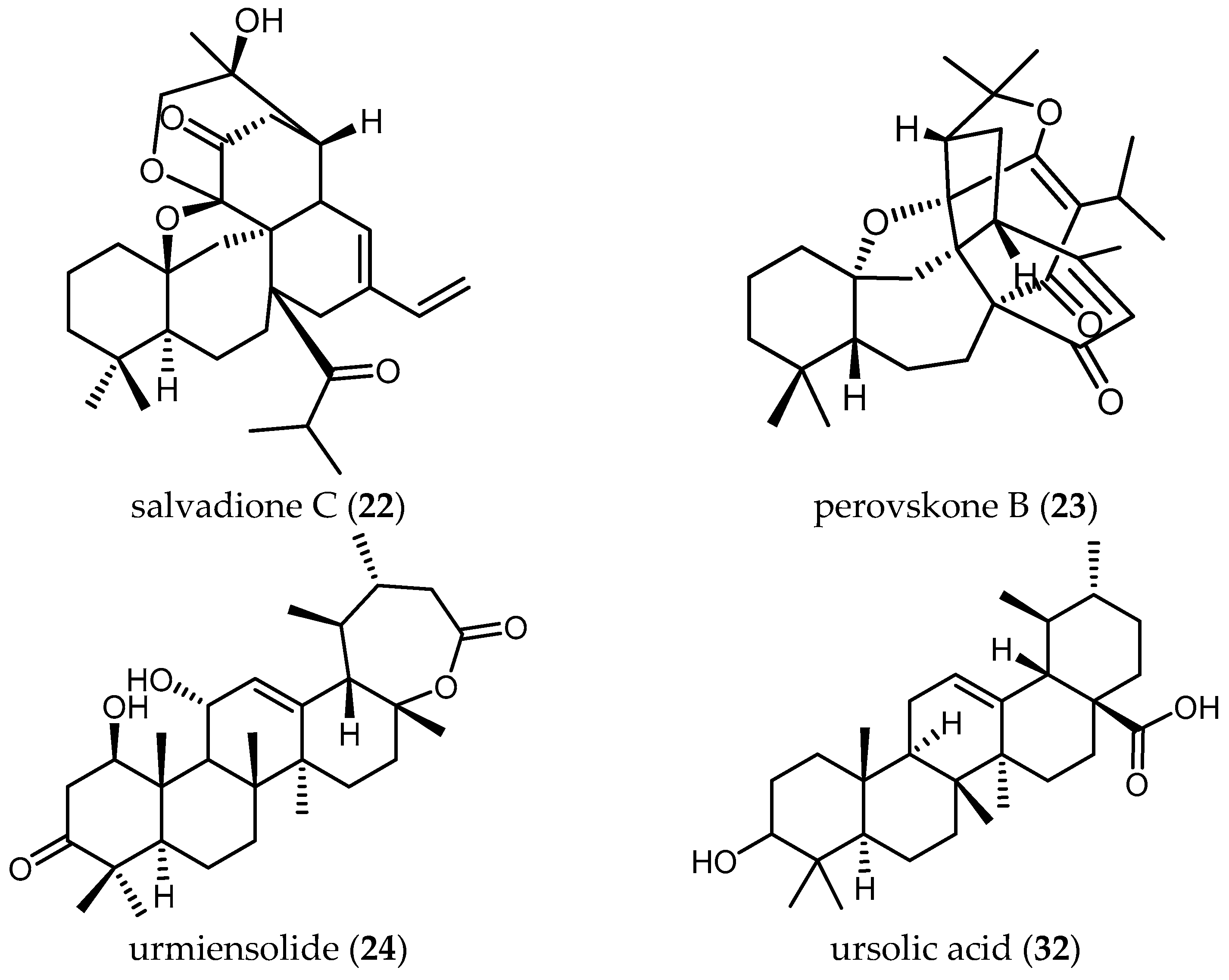
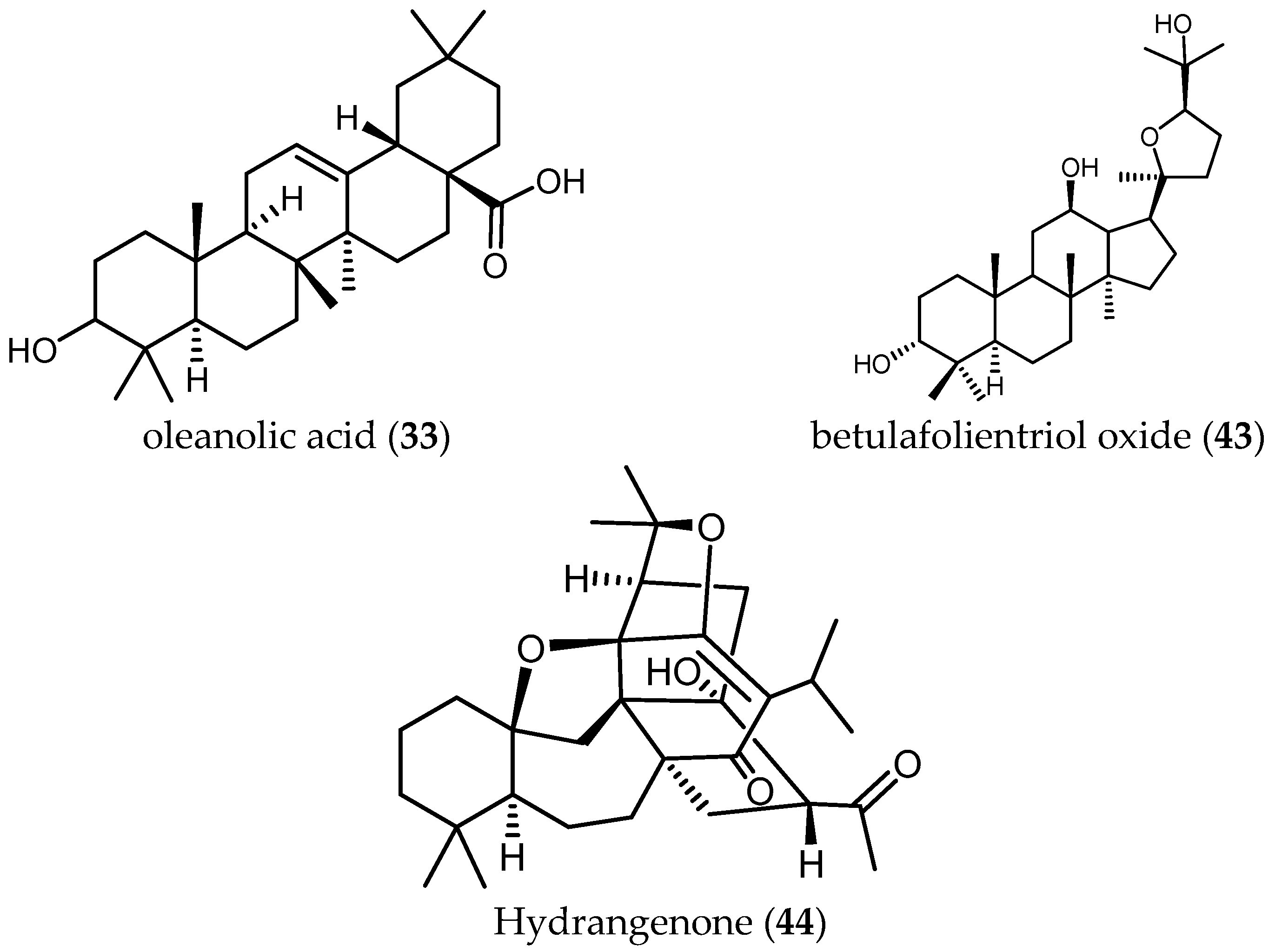
Figure 3.
Phenolic compounds of Salvia with antiprotozoal activity.

Figure 4.
Flavonoids of Salvia with antiprotozoal activity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biological activity of extracts from different species of Salvia against Trypanosoma brucei (n.d./n.f. = not determined/not found).
Table 1.
Biological activity of extracts from different species of Salvia against Trypanosoma brucei (n.d./n.f. = not determined/not found).
| Parasite | Strain | Plant Species | IC50 Extract | Type of Solvent Extract | SI | Ref. |
|---|---|---|---|---|---|---|
| Tbr | STIB 900 | S. tomentosa Mill. (a1) | 3.64 µg/mL | MeOH-extract | 24.73 | [16] |
| Tbr | STIB 900 | S. tomentosa Mill. (a2) | 1.24 µg/mL | n-Hexane-extract | 21.16 | [16] |
| Tbr | STIB 900 | S. tomentosa Mill. (a3) | 2.33 µg/mL | Chloroform-extract | 13.65 | [16] |
| Tbr | STIB 900 | S. tomentosa Mill. (a4) | 10.96 µg/mL | H2O-extract | >8 | [16] |
| Tbr | STIB 900 | S. sclarea L. (b1) | 6.44 µg/mL | MeOH-extract | 13.6 | [16] |
| Tbr | STIB 900 | S. sclarea L. (b2) | 2.4 µg/mL | n-Hexane-extract | 7.63 | [16] |
| Tbr | STIB 900 | S. sclarea L. (b3) | 4.4 µg/mL | Chloroform-extract | 19.07 | [16] |
| Tbr | STIB 900 | S. sclarea L. (b4) | 10.31 µg/mL | H2O-extract | >8.7 | [16] |
| Tbr | STIB 900 | S. dichroantha Stapf. (c1) | 3.58 µg/mL | MeOH-extract | >25 | [16] |
| Tbr | STIB 900 | S. dichroantha Stapf. (c2) | 3.5 µg/mL | n-Hexane-extract | >25.7 | [16] |
| Tbr | STIB 900 | S. dichroantha Stapf. (c3) | 4.4 µg/mL | Chloroform-extract | 19.27 | [16] |
| Tbr | STIB 900 | S. dichroantha Stapf. (c4) | 7.77 µg/mL | H2O-extract | >11.5 | [16] |
| Tbr | STIB 900 | S. hydrangea DC. ex Benth. (d) | 18 µg/mL | n-Hexane-extract | n.d. | [25] |
| Tbr | STIB 900 | S. miltiorrhiza Bunge (e) | 97% inhibition at 0.81 µg/mL | DCM-extract | n.d. | [4] |
| Tbr | STIB 900 | S. repens Burch.ex Benth. (f) | 10.8 µg/mL | DCM-MeOH (1:1) | 3.82 | [26] |
| Tbb | Strain 427 | S. spathacea Greene (g) | 1.13 ± 0.78 µg/mL | Ethanolic extract | n.d. | [19] |
| Tbb | n.d. | S. lavandulifolia Vahl. (h) | No activity at >20 mg/mL | Essential oil | n.d. | [20] |
| Tbr | STIB 900 | S. officinalis L. (i) | 1.86 µg/mL | Ethanolic tincture | 17.3 | [21] |
Table 2.
Biological activity of extracts of Salvia spp. and their isolated compounds against Trypanosoma brucei, (n.d./n.f.d. = not determined/not found).
Table 2.
Biological activity of extracts of Salvia spp. and their isolated compounds against Trypanosoma brucei, (n.d./n.f.d. = not determined/not found).
| Parasite | Form/Strain | Plant Species | IC50 Extract | Isolated Compound | Molecular Formula | Molar Mass (g/mol) | IC50 of the Compound (µM) | SI | Ref |
|---|---|---|---|---|---|---|---|---|---|
| Tbb | GUT at 3.1 | S. leucantha Cav. | n.d./n.f. | salviandulin E (1) | C20H16O6 | 352.09 | 2.04 µM (0.72 µg/mL) | 1.17 | [22] |
| Tbb | GUT at 3.1 | S. leucantha Cav. | n.d./n.f. | 6,7-dehydrodugesin A (2) | C20H16O5 | 336.09 | >37.19 µM (>12.5 µg/mL) | n.d. | [22] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | miltirone (3) | C19H22O2 | 282.16 | 0.5 µM | 2.6 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | methylenetanshinquinone (4) | C18H14O3 | 278.09 | 0.5 µM | 24.2 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | tanshinone I (5) | C18H12O3 | 276.08 | 1.3 µM | 9.5 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | tanshinone IIa (6) | C19H18O3 | 294.13 | 1.5 µM | 5 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | methyl-tanshinonate (7) | C20H18O5 | 338.12 | 17 µM | 0.4 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | 11-hydroxymiltiodiol (8) | C19H22O3 | 298.16 | 3.6 µM | n.d. | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. miltiorrhiza Bunge. | 97% inhib. at 0.81 µg/mL | cryptotanshinone (9) | C19H20O3 | 296.14 | 26.2 µM | 0.2 | [4] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | 12-deoxy-salvipisone (10) | C20H24O2 | 296.18 | 2.5 µM | 0.2 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | sahandinone (11) | C20H24O2 | 296.18 | 1.8 µM | 0.2 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | 12-deoxy-6,7-dehydroroyleanone (12) | C20H26O2 | 298.19 | 32.3 µM | 0.9 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | 7α-acetoxyroyleanone (13) | C22H30O5 | 374.21 | 2.9 µM | 0.1 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | Δ 9-ferruginol (14) | C20H28O | 284.21 | 12.8 µM | 1.2 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | ferruginol (15) | C20H30O | 286.23 | 28.1 µM | 0.5 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | sahandol (16) | C20H24O2 | 296.18 | 18.4 µM | 0.8 | [23] |
| Tbr | Trypomastigotes/ STIB 900 | S. sahendica Boiss. & Buhse | n.d./n.f. | sahandone (17) | C21H26O3 | 326.19 | 19.5 µM | 0.6 | [23] |
| Tbr | STIB 900 | S.austriaca Jacq. | n.d./n.f. | taxodione (18) | C20H26O3 | 314.19 | 0.05 µM | 38 | [24] |
| Tbr | STIB 900 | S.austriaca Jacq. | n.d./n.f. | 15-deoxy-fuerstione (19) | C20H26O2 | 298.19 | 194.7 µM | 0.69 | [24] |
| Tbr | STIB 900 | S.austriaca Jacq. | n.d./n.f. | 7-(2′-oxohexyl)-taxodione (20) | C26H36O4 | 412.26 | 0.62 µM | 5.0 | [24] |
| Tbr | STIB 900 | S.austriaca Jacq. | n.d./n.f. | taxodone (21) | C20H28O3 | 316.20 | 1.67 µM | 2.4 | [24] |
| Tbr | STIB 900 | S. hydrangea DC. Ex Benth. | 18 µg/mL | salvadione C (22) | C30H40O5 | 480.29 | 4.33 µM | 43.2 | [25] |
| Tbr | STIB 900 | S. hydrangea DC. Ex Benth. | 18 µg/mL | perovskone B (23) | C30H40O4 | 464.29 | 15.92 µM | 0.78 | [25] |
| Tbr | STIB 900 | S. urmiensis Bunge. | n.d./n.f. | urmiensolide (24) | C30H46O5 | 486.33 | 5.6 μM | 33 | [17] |
Table 3.
Biological activity of extracts from different species of Salvia against Trypanosoma cruzi.
Table 3.
Biological activity of extracts from different species of Salvia against Trypanosoma cruzi.
| Parasite | Form/Strain | Plant Species | IC50 Extract | Type of Solvent Extract | SI | Ref. |
|---|---|---|---|---|---|---|
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. tomentosa Mill. (a1) | >90 µg/mL | MeOH-extract | <1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. tomentosa Mill. (a2) | 28.46 µg/mL | n-Hexane-extract | 0.92 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. tomentosa Mill. (a3) | 35.72 µg/mL | Chloroform-extract | 0.89 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. tomentosa Mill. (a4) | >90 µg/mL | H2O-extract | ≈1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. sclarea L. (b1) | 56.82 µg/mL | MeOH-extract | 1.54 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. sclarea L. (b2) | 18.17 µg/mL | n-Hexane-extract | 1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. sclarea L. (b3) | 52.51 µg/mL | Chloroform-extract | 1.59 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. sclarea L. (b4) | >90 µg/mL | H2O-extract | ≈1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. dichroantha Stapf. (c1) | >90 µg/mL | MeOH-extract | ≈1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. dichroantha Stapf. (c2) | 41.85 µg/mL | n-Hexane-extract | >2.15 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. dichroantha Stapf. (c3) | 48.99 µg/mL | Chloroform-extract | 1.73 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. dichroantha Stapf. (c4) | >90 µg/mL | H2O-extract | ≈1 | [16] |
| Tc | Trypomastigote/Tulahuen strain C2C4 | S. repens Burch.ex Benth. (f) | 36.2 µg/mL | DCM-MeOH (1:1) | 1.15 | [26] |
Table 4.
Biological activity of extracts of Salvia spp. and their isolated compounds against Trypanosoma cruzi. (n.d.= not determined).
Table 4.
Biological activity of extracts of Salvia spp. and their isolated compounds against Trypanosoma cruzi. (n.d.= not determined).
| Parasite | Form/Strain | Plant Species | IC50 Extract | Isolated Compound | Molecular Formula | Molar Mass (g/mol) | IC50 of the Compound (µM) | SI | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Tc | Epimastigotes/Tulhauen | S. gilliessi Benth. | n.d. | 5-epi-icetexone (ICTX) (25) | C20H22O5 | 342.15 | 6.5 ± 0.75 µM | n.d. | [27] |
| Tc | Amastigotes/Y Strain | S. gilliessi Benth. | n.d. | 5-epi-icetexone (ICTX) (25) | C20H22O5 | 342.15 | n.d. | n.d. | [28] |
| Tc | Epimastigotes/Dm28c strain | S. cuspidata (Ruiz & Pav. Subsp. gilliesii (Benth.) J.R.I. Wood | n.d. | 12-hydroxy-11,14-diketo-6,8,12-abietatrien-19,20-olide (HABTO) (26) | C17H16O5 | 300.10 | ≈16.6 µM (≈5 µg/mL) | n.d. | [30] |
| Tc | Amastigotes/Tulahuen strain C2C4 containing Lac Z | S. austriaca Jacq. | n.d. | taxodione (18) | C20H26O3 | 314.19 | 7.11 µM | 0.27 | [24] |
| Tc | Amastigotes/Tulahuen strain C2C4 containing Lac Z | S. austriaca Jacq. | n.d. | 15-Deoxy-fuerstione (19) | C20H26O2 | 298.19 | 146.9 µM | 0.91 | [24] |
| Tc | Amastigotes/Tulahuen strain C2C4 containing Lac Z | S. austriaca Jacq. | n.d. | 7-(2′-oxohexyl)-taxodione (20) | C26H36O4 | 412.26 | 7.76 µM | 0.4 | [24] |
| Tc | Amastigotes/Tulahuen strain C2C4 containing Lac Z | S. austriaca Jacq. | n.d. | taxodone (21) | C20H28O3 | 316.20 | 7.63 µM | 0.5 | [24] |
Table 5.
Biological activity of extracts from different species of Salvia against Leishmania spp. (n.d./n.f. = not determined/not found). *.
Table 5.
Biological activity of extracts from different species of Salvia against Leishmania spp. (n.d./n.f. = not determined/not found). *.
| Parasite | Form/Strain | Plant Species | IC50 Extract | Type of Solvent Extract | SI | Ref. |
|---|---|---|---|---|---|---|
| n.d. | (Popular use antileishmanial remedy) | S. hydrangea DC ex. Benth. (d1) | n.d. | n.d. | n.d. | [8] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. tomentosa Mill. (a1) | 14.92 µg/mL | MeOH-extract | 24.73 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. tomentosa Mill. (a2) | 2.49 µg/mL | n-Hexane-extract | 21.16 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. tomentosa Mill. (a3) | 1.81 µg/mL | Chloroform-extract | 29.47 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. tomentosa Mill. (a4) | >90 µg/mL | H2O-extract | >10 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. sclarea L. (b1) | 12.95 µg/mL | MeOH-extract | 13.6 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. sclarea L. (b2) | 5.25 µg/mL | n-Hexane-extract | 7.63 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. sclarea L. (b3) | 8.31 µg/mL | Chloroform-extract | 19.07 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. sclarea L. (b4) | 47.88 µg/mL | H2O-extract | >9 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. dichroantha Stapf. (c1) | 4.93 µg/mL | MeOH-extract | >25 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. dichroantha Stapf. (c2) | 3.48 µg/mL | n-Hexane-extract | 25.7 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. dichroantha Stapf. (c3) | 2.31 µg/mL | Chloroform-extract | 19.27 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. dichroantha Stapf. (c4) | >90 µg/mL | H2O-extract | >11 | [16] |
| Ld | Axenic amastigotes/ strain MHOM/ET/67/L82 | S. repens Burch. ex Benth. (f) | 5.36 µg/mL | DCM-MeOH (1:1) | 7.74 | [26] |
| Ld | Promastigotes | S. lavandulifolia Vahl. (h) | No activity up to 20 µg/mL | Essential oil | n.d. | [20] |
| Ld | Axenic amastigotes | S. lavandulifolia Vahl. (h) | No activity up to 20 µg/mL | Essential oil | n.d. | [20] |
| Ld | Intracellular amastigotes | S. lavandulifolia Vahl. (h) | No activity up to 20 µg/mL | Essential oil | n.d. | [20] |
| Lm | Promastigote/ LCO3 | S.officinalis L. (i) | 3.40 ± 0.16 µg/mL | Essential oil | 5.92 | [34] |
| Li | Promastigote/ LV20 | S.officinalis L. (i1) | 2.67 ± 0.33 µg/mL | Essential oil | 7.54 | [34] |
| Lm | Promastigotes/ MROH/IR/75/IR | S.officinalis L. (i2) | 184 ± 11.17 µg/mL | MeOH-maceration | n.d. | [35] |
| Lm | Amastogotes | S.officinalis L. (i2) | 58% letally at 184 ± 11.17 µg/mL | MeOH-maceration | n.d. | [35] |
| Lm | Promastigotes | S. bucharica Popov (j) | 72.31 µg/mL | MeOH-extract | n.d. | [36] |
| Lm | Promastigotes | S. bucharica Popov (j1) | 50.51 µg/mL | Chloroform-extract | n.d. | [36] |
| Lm | Promastigotes | S. bucharica Popov (j2) | >100 µg/mL | Acetone-extract | n.d. | [36] |
| Lm | Promastigotes | S. bucharica Popov (j3) | 30.51 µg/mL | H2O extract | n.d. | [36] |
| Lm | Promastigotes/ MHOM/MA/2009/LCER19-09 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k) | 155.43 µg/mL | n-Hexane-extract | n.d. | [37] |
| Lm | Promastigotes/ MHOM/MA/2009/LCER19-09 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k1) | 24.56 µg/mL | DCM-extract | n.d. | [37] |
| Lm | Promastigotes/ MHOM/MA/2009/LCER19-09 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k2) | >1000 µg/mL | MeOH-extract | n.d. | [37] |
| Li | Promastigotes/ MHOM/MA/1998/LVTA | S. verbenaca (L.) Briq. ssp verbenaca Maire (k) | 14.11 µg/mL | n-Hexane-extract | n.d. | [37] |
| Li | Promastigotes/ MHOM/MA/1998/LVTA | S. verbenaca (L.) Briq. ssp verbenaca Maire (k1) | 31.57 µg/mL | DCM-extract | n.d. | [37] |
| Li | Promastigotes/ MHOM/MA/1998/LVTA | S. verbenaca (L.) Briq. ssp verbenaca Maire (k2) | >1000 µg/mL | MeOH-extract | n.d. | [37] |
| Lt | Promastigotes/ MHOM/MA/2010/LCTIOK-4 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k) | 148.23 µg/mL | n-Hexane-extracts | n.d. | [37] |
| Lt | Promastigotes/ MHOM/MA/2010/LCTIOK-4 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k1) | 33.77 µg/mL | DCM-extract | n.d. | [37] |
| Lt | Promastigotes/ MHOM/MA/2010/LCTIOK-4 | S. verbenaca (L.) Briq. ssp verbenaca Maire (k2) | >1000 µg/mL | MeOH-extract | n.d. | [37] |
* Leishmania donovani (Ld), Leishmania major (Lm), Leishmania infantum (Li), Leishmania guyanensis (Lg), Leishnmania killicki (Lk), and Leishmania tropica (Lt).
Table 6.
Biological activity of extracts of Salvia spp. and their isolated compounds against Leishmania spp. (n.d./n.f. = not determined/not found). *
Table 6.
Biological activity of extracts of Salvia spp. and their isolated compounds against Leishmania spp. (n.d./n.f. = not determined/not found). *
| Parasite | Form/Strain | Plant Species | IC50 Extract | Isolated Compound | Molecular Formula | Molar Mass (g/mol) | IC50 of the Compound | SI | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Ld | Axenic amastigotes/ MHOM/ET/67/L82 | S.repens Burch.ex Benth. | 5.36 µg/mL | 12-methoxycarnosic acid (27) | C21H30O4 | 346.21 | 0.75 μM | 23.06 | [18] |
| Ld | Intracellular Amastgitotes /n.d. | S. cilicica Boiss and Kotschy | n.d. | 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione (28) | C20H22O4 | 326.15 | 0.17 μM (170 nM) | >1.76 | [38] |
| Ld | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione (28) | C20 H22 O4 | 326.15 | >0.3 μM (>300 nM) | ≈1 | [38] |
| Lm | Amastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione (28) | C20H22O4 | 326.15 | 0.287 μM (287.4 nM) | 1.04 | [38] |
| Lm | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione (28) | C20H22O4 | 326.15 | ≥0.3 μM (≥300 nM) | ≈1 | [38] |
| Ld | Amastgitotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | abieta-8,12-dien-11,14-dione (12-deoxy-royleanone) (29) | C20H28O2 | 300.21 | 0.121 μM (121 nM) | 1.58 | [38] |
| Ld | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | abieta-8,12-dien-11,14-dione (12-deoxy-royleanone) (29) | C20H28O2 | 300.21 | >0.3 μM (>300 nM) | ≤0.64 | [38] |
| Lm | Amastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | abieta-8,12-dien-11,14-dione (12-deoxy-royleanone) (29) | C20H28O2 | 300.21 | 0.182 μM (182.3 nM) | 1.04 | [38] |
| Lm | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | abieta-8,12-dien-11,14-dione (12-deoxy-royleanone) (29) | C20H28O2 | 300.21 | ≥0.30 μM (≥300 nM) | ≤0.63 | [38] |
| Ld | Promastigotes/n.d. | S. deserta Schang. | n.d. | taxodione (18) | C20H26O3 | 314.42 | 1.46 ± 0.52 μM | 10.34 | [39] |
| Ld | Promastigotes/n.d. | S. deserta Schang. | n.d. | ferruginol (15) | C20H30O | 286.45 | 11.39 ± 1.05 μM | n.d. | [39] |
| Ld | Promastigotes/n.d. | S. deserta Schang. | n.d. | 7-O-acetylhorminone (30) | C22H30O5 | 374.47 | 19.69 ± 0.80 μM | n.d. | [39] |
| Ld | Promastigotes/n.d. | S. deserta Schang. | n.d. | horminone (31) | C20H28O4 | 332.20 | 29.43 ± 3.01 μM | n.d. | [39] |
| Ld | Amastgitotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | ursolic acid (32) | C30H48O3 | 456.36 | 0.0127 μM (12.7 nM) | 2.22 | [38] |
| Ld | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | ursolic acid (32) | C30H48O3 | 456.36 | 0.091 μM (91 nM) | 0.17 | [38] |
| Lm | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | ursolic acid (32) | C30H48O3 | 456.36 | 0.051 μM (51.3 nM) | 0.3 | [38] |
| Lm | Amastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | ursolic acid (32) | C30H48O3 | 456.36 | 0.007 μM (7 nM) | 2.22 | [38] |
| Ld | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | oleanolic acid (33) | C30H48O3 | 456.36 | 0.091 μM (91 nM) | 1.45 | [38] |
| Ld | Amastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | oleanolic acid (33) | C30H48O3 | 456.36 | 0.063 μM (62.9 nM) | 2.1 | [38] |
| Lm | Promastigotes/n.d. | S. cilicica Boiss and Kotschy | n.d. | oleanolic acid (33) | C30H48O3 | 456.36 | 0.137 μM (137 nM) | 0.97 | [38] |
| Lm | Amastigotes | S. cilicica Boiss and Kotschy | n.d. | oleanolic acid (33) | C30H48O3 | 456.36 | 0.120 μM (119.9 nM) | 1.1 | [38] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | caffeic acid (34) | C9H8O4 | 180.04 | 0.0044 μM (4.4 nM) | >2200 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | caffeic acid (34) | C9H8O4 | 180.04 | 0.0061 μM (6.1 nM) | >500 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | caffeic acid (34) | C9H8O4 | 180.04 | 0.0066 μM (6.6 nM) | >360 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | caffeic acid (34) | C9H8O4 | 180.04 | 0.0039 μM (3.9 nM) | >333 | [41] |
| Lm/Ld/Lk/Lg | Promastigotes | S. officinalis L. | n.d. | caffeic acid (34) | C9H8O4 | 180.04 | >2.8 μM (>2800 nM) | ≈0.78 | [41] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | rosmarinic acid (35) | C18H16O8 | 360.08 | 0.0592 μM (59.2 nM) | >18 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | rosmarinic acid (35) | C18H16O8 | 360.08 | 0.0744 μM (74.4 nM) | >15 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | rosmarinic acid (35) | C18H16O8 | 360.08 | 0.0842 μM (84.2 nM) | >13 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | rosmarinic acid (35) | C18H16O8 | 360.08 | 0.0694 μM (69.4 nM) | >15 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | rosmarinic acid (35) | C18H16O8 | 360.08 | >1.4 μM (>1400 nM) | ≈0.78 | [41] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | salvianolic acid I (36) | C27H22O12 | 538.11 | 0.1604 μM (160.4 nM) | > 4.35 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | salvianolic acid I (36) | C27H22O12 | 538.11 | 0.1758 μM (175.8 nM) | >3.9 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | salvianolic acid I (36) | C27H22O12 | 538.11 | 0.1515 μM (151.5 nM) | >4.62 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | salvianolic acid I (36) | C27H22O12 | 538.11 | 0.1678 μM (167.8 nM) | >4.17 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | salvianolic acid I (36) | C27H22O12 | 538.11 | >0.9 μM (>900 nM) | ≈0.77 | [41] |
| Lm | Amastigotes/strain | S. officinalis L. | n.d. | methyl ester of salvianolic acid I (37) | C28H24O12 | 552.13 | 0.0108 μM (10.8 nM) | >64.81 | [41] |
| Ld | Amastigotes | S. officinalis L. | n.d. | methyl ester of salvianolic acid I (37) | C28H24O12 | 552.13 | 0.0186 μM (18.6 nM) | >37.63 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | methyl ester of salvianolic acid I (37) | C28H24O12 | 552.13 | 0.0152 μM (15.2 nM) | >46 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | methyl ester of salvianolic acid I (37) | C28H24O12 | 552.13 | 0.0136 μM (13.6 nM) | >51.47 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | methyl ester of salvianolic acid I (37) | C28H24O12 | 552.13 | >0.9 μM (>900 nM) | ≈0.77 | [41] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | salvianolic acid K (38) | C27H24O13 | 556.12 | 0.0183 μM (18.3 nM) | >38.25 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | salvianolic acid K (38) | C27H24O13 | 556.12 | 0.0182 μM (18.2 nM) | >38.46 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | salvianolic acid K (38) | C27H24O13 | 556.12 | 0.0133 μM (13.3 nM) | >52.63 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | salvianolic acid K (38) | C27H24O13 | 556.12 | 0.0145 μM (14.5 nM) | >48.27 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | salvianolic acid K (38) | C27H24O13 | 556.12 | >0.9 μM (>900 nM) | ≈0.77 | [41] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | salvianolic acid L (39) | C36H30O16 | 718.15 | 0.0203 μM (20.3 nM) | >34,48 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | salvianolic acid L (39) | C36H30O16 | 718.15 | 0.0154 μM (15.4 nM) | >45.45 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | salvianolic acid L (39) | C36H30O16 | 718.15 | 0.0226 μM (22.6 nM) | >30.97 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | salvianolic acid L (39) | C36H30O16 | 718.15 | 0.013 μM (13.0 nM) | >53.84 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | salvianolic acid L (39) | C36H30O16 | 718.15 | >0.9 μM (>900 nM) | ≈0.77 | [41] |
| Lm | Amastigotes/ LV39 strain | S. officinalis L. | n.d. | sagerinic acid (40) | C36H32O16 | 720.17 | 0.128 μM (128.7 nM) | >4.66 | [41] |
| Ld | Amastigotes/ LV9 strain | S. officinalis L. | n.d. | sagerinic acid (40) | C36H32O16 | 720.17 | 0.122 μM (122.1 nM) | >4.91 | [41] |
| Lg | Amastigotes | S. officinalis L. | n.d. | sagerinic acid (40) | C36H32O16 | 720.17 | 0.142 μM (141.5 nM) | >4.24 | [41] |
| Lk | Amastigotes | S. officinalis L. | n.d. | sagerinic acid (40) | C36H32O16 | 720.17 | 0.155 μM (154.8 nM) | >3.88 | [41] |
| Lm/Ld/ Lk/Lg | Promastigote | S. officinalis L. | n.d. | sagerinic acid (40) | C36H32O16 | 720.17 | >0.7 μM (>700 nM) | ≈0.85 | [41] |
* Leishmania donovani (Ld), Leishmania major (Lm), Leishmania infantum (Li), Leishmania guyanensis (Lg), Leishnmania killicki (Lk), and Leishmania tropica (Lt).
Table 7.
Biological activity of extracts and essential oils of Salvia spp. against P. falciparum (n.d./n.f. = not determined/not found).
Table 7.
Biological activity of extracts and essential oils of Salvia spp. against P. falciparum (n.d./n.f. = not determined/not found).
| Parasite | Strain | Plant Species | Type of Solvent Extract | IC50 Solvent Extract | SI | Ref. |
|---|---|---|---|---|---|---|
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. tomentosa Mill. (a1) | MeOH-extract | 9.94 µg/mL | 9.05 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. tomentosa Mill. (a2) | n-Hexane-extract | 3.47 µg/mL | 7.56 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. tomentosa Mill. (a3) | Chloroform-extract | 3.14 µg/mL | 10.12 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. tomentosa Mill. (a4) | H2O-extract | >20 µg/mL | ≈4.5 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. sclarea L. (b1) | MeOH-extract | 6.6 µg/mL | 13.27 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. sclarea L. (b2) | n-Hexane-extract | 3.78 µg/mL | 4.84 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. sclarea L. (b3) | Chloroform-extract | 2.54 µg/mL | 33.03 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. sclarea L. (b4) | H2O-extract | >20 µg/mL | ≈4.5 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. dichroantha Stapf. (c1) | MeOH-extract | 8.85 µg/mL | >10.17 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. dichroantha Stapf. (c2) | n-Hexane-extract | 4.17 µg/mL | >21.58 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. dichroantha Stapf. (c3) | Chloroform-extract | 3.72 µg/mL | 22.79 | [16] |
| Pf | Chloroquine- and Pyrimethamine-resistant K1 strain | S. dichroantha Stapf. (c4) | H2O-extract | >20 µg/mL | ≈4.5 | [16] |
| Pf | K1 strain | S. hydrangea DC ex. Benth. (d) | n-Hexane-extract | 3.2 µg/mL | n.d. | [25] |
| Pf | 3D7 strain | S. hydrangea DC ex. Benth. (d2) | n.d. | <12.5 µg/mL | n.d. | [8] |
| Pf | K1 strain | S. repens Burch. ex Benth. (f) | DCM/MeOH 1:1 | 7.65 µg/mL | 5.4 | [26] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. repens Burch. ex Benth. (f1) | Chloroform/MeOH (1:1) | 8.25 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. repens Burch. ex Benth. (f2) | Essential oil | 1.65 µg/mL | n.d. | [45] |
| Pf | Chloroquine-sensitive strain D10 | S. repens Burch. ex Benth. (f3) | DCM/MeOH 1:1 | 10.8 µg/mL | n.d. | [46] |
| Pf | n.d. | S. repens Burch. ex Benth. (f4) | Methanol extract | 78.9 µg/mL | 1.26 | [6] |
| Pf | n.d. | S. repens Burch. ex Benth. (f5) | Essential oil | 1.68 µg/mL | 5.28 | [6] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. africana–caerulea L. (l) | Essential oil | 4.76 µg/m | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. africana–lutea L. (m) | Chloroform/MeOH (1:1) | 15.86 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. africana–lutea L (m1) | Essential oil | 5.45 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. albicaulis Benth. (n) | Chloroform/MeOH (1:1) | 15.83 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. albicaulis Benth. (n1) | Essential oil | 6.41 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. muirii L. Bol. (o) | Chloroform/MeOH (1:1) | 11.87 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. muirii L. Bol. (o1) | Essential oil | 5.93 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. lanceolata Lam. (p) | Chloroform/MeOH (1:1) | 26.01 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. lanceolata Lam. (p1) | Essential oil | 7.83 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. garipensis E. Mey. (q) | Chloroform/MeOH (1:1) | 13.95 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. dolomitica Codd (r) | Chloroform/MeOH (1:1) | 7.62 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. dolomitica Codd (r1) | Essential oil | 4.81 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. disermas L. (s) | Chloroform/MeOH (1:1) | 24.17 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. chamelaeagnea Berg. (t) | Chloroform/MeOH (1:1) | 8.71 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. chamelaeagnea Berg. (t1) | Essential oil | 8.63 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. aurita L.f. (u) | Chloroform/MeOH (1:1) | 8.92 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. namaensis Schinz (v) | Chloroform/MeOH (1:1) | 25.38 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. runcinata L.f. (v1) | Chloroform/MeOH (1:1) | 16.61 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. runcinata L.f. (v2) | Essential oil | 1.20 µg/mL | n.d. | [45] |
| Pf | n.d. | S. runcinata L.f. (v3) | MeOH extract | 29 µg/mL | >3.44 | [6] |
| Pf | n.d. | S. runcinata L.f. (v4) | Essential oil | 1.23 µg/mL | 2.06 | [6] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. schlechteri Briq. (w) | Chloroform/MeOH (1:1) | 17.51 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. stenophylla Burch. Ex. Benth. (x1) | Chloroform/MeOH (1:1) | 6.5 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. stenophylla Burch. Ex. Benth. (x2) | Essential oil | 4.13 µg/mL | n.d. | [45] |
| Pf | n.d. | S. stenophylla Burch. Ex. Benth. (x3) | MeOH-extract | 17.03 µg/mL | 1.27 | [6] |
| Pf | n.d. | S. stenophylla Burch. Ex. Benth. (x4) | Essential oil | 4.38 µg/mL | 1.53 | [6] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. verbenaca L. (y) | Chloroform/MeOH (1:1) | 23.97 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. radula Benth. (z1) | Chloroform/MeOH (1:1) | 3.91 µg/mL | n.d. | [45] |
| Pf | Chloroquine-resistant Pf FCR-3 strain | S. radula Benth. (z2) | Essential oil | 13.5 µg/mL | n.d. | [45] |
| Pf | Chloroquine resistance Pf K1 strain | S. chudaei Batt. & Trab. (Aa) | n-Hexane-extract | 4.91 ± 2.91 µg/mL | n.d. | [47] |
| Pf | 3D7-chloroquine sensitive strain | S. chudaei Batt. & Trab. (Aa) | n-Hexane extract | 5.69 ± 0.3 µg/mL | n.d. | [47] |
| Pf | Chloroquine resistance Pf K1 strain | S. chudaei Batt. & Trab. (Aa1) | Chloroform-extract | 6.41 ± 0.31 µg/mL | n.d. | [47] |
| Pf | 3D7-chloroquine sensitive strain | S. chudaei Batt. & Trab. (Aa1) | Chloroform-extract | 5.69 ± 2.71 µg/mL | n.d. | [47] |
| Pf | Chloroquine resistance Pf K1 strain | S. chudaei Batt. & Trab. (Aa2) | Ethanolic-extract | 7.99 ± 1.06 µg/mL | n.d. | [47] |
| Pf | 3D7-chloroquine sensitive strain | S. chudaei Batt. & Trab. (Aa2) | Ethanolic-extract | 8.43 ± 0.38 µg/mL | n.d. | [47] |
| Pf | Chloroquine resistance Pf K1 strain | S. chudaei Batt. & Trab. (Aa3) | Essential oil | 2.39 ± 0.24 µg/mL | n.d. | [47] |
| Pf | 3D7-chloroquine sensitive strain | S. chudaei Batt. & Trab. (Aa3) | Essential oil | 2.40 ± 0,77 µg/mL | n.d. | [47] |
| Pf | Chloroquine sensitive D6 strain | S. lavandulifolia Vahl (h) | Essential oil | 47% inhibition at 15.86 µg/mL | n.d. | [20] |
Table 8.
Biological activity of extracts of Salvia spp. and their isolated compounds against Plasmodium (n.d./n.f. = not determined/not found).
Table 8.
Biological activity of extracts of Salvia spp. and their isolated compounds against Plasmodium (n.d./n.f. = not determined/not found).
| Parasite | Strain | Plant Species | IC50 Extract | Isolated Compound | Molecular Formula | Molar Mass (g/mol) | IC50 of the Compound | SI | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | miltirone (3) | C19H22O2 | 282.38 | 0.5 µM | 2.6 | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | methylenetanshinquinone (4) | C18H14O3 | 278.09 | 6.3 µM | 1.9 | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | tanshinone I (5) | C18H12O3 | 276.08 | 7.2 µM | 1.7 | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | tanshinone IIa (6) | C19H18O3 | 294.13 | 4 µM | 1.9 | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | methyl-tanshinonate (7) | C20H18O5 | 338.12 | 4.1 µM | 1.7 | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | 11-hydroxymiltiodiol (8) | C19H22O3 | 298.16 | ≥30 µM | n.d. | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | cryptotanshinone (9) | C19H20O3 | 296.14 | 16.1 µM | 0.3 | [4] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | 12-deoxy-salvipisone (10) | C20H24O2 | 296.18 | 8.8 µM | 0.1 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | sahandinone (11) | C20H24O2 | 296.18 | 5.1 µM | 0.1 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | 12-deoxy-6,7-dehydroroyleanone (12) | C20H26O2 | 298.19 | 17.8 µM | 0.6 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | 7α-acetoxyroyleanone (13) | C22H30O5 | 374.21 | 1.3 µM | 0.1 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | Δ9-ferruginol (14) | C20H28O | 284.21 | 0.9 µM | 18.2 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | ferruginol (15) | C20H30O | 286.23 | 0.9 µM | 15.6 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | sahandol (16) | C20H24O2 | 296.18 | 4.7 µM | 3.3 | [23] |
| Pf | K1 strain | S. sahendica Boiss. & Buhse | n.d. | sahandone (17) | C21H26O3 | 326.19 | 17.2 µM | 0.7 | [23] |
| Pf | K1 strain | S. hydrangea DC. ex Benth. | 3.2 µg/mL | salvadione C (22) | C30H40O5 | 480.29 | 1.43 µM | 86.2 | [25] |
| Pf | K1 strain | S. hydrangea DC. ex Benth. | 3.2 µg/mL | perovskone B (23) | C30H40O4 | 464.29 | 18 µM | 69.6 | [25] |
| Pf | 3D7 strain | S. hydrangea DC. ex Benth. | <12.5 µg/mL | oleanolic acid (33) | C30H48O3 | 456.36 | 19.3 µM | n.d. | [8] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | 1-oxomiltirone (41) | C19H20O3 | 296.14 | ≥30 µM | n.d. | [4] |
| Pf | K1 strain | S. miltiorrhiza Bunge. | 34% inhib. at 0.81 µg/mL | 1,2-dihydrotanshinquinone (42) | C18H14O3 | 278.09 | 5.3 µM | 0.8 | [4] |
| Pf | NF54 strain | S. austriaca Jacq. | n.d. | taxodione (18) | C20H26O3 | 314.19 | 1.9 µM | 1.0 | [24] |
| Pf | NF54 strain | S. austriaca Jacq. | n.d. | 15-deoxy-fuerstione (19) | C20H26O2 | 298.19 | >167 µM | n.d. | [24] |
| Pf | NF54 strain | S. austriaca Jacq. | n.d. | 7-(2′-oxohexyl)-taxodion (20) | C26H36O4 | 412.26 | 3.37 µM | 0.9 | [24] |
| Pf | NF54 strain | S. austriaca Jacq. | n.d. | taxodone (21) | C20H28O3 | 316.20 | 3.66 µM | 1.1 | [24] |
| Pf | D6 strain | S. deserta Schang. | n.d. | taxodione (18) | C20H26O3 | 314.19 | 10.49 μM (3.297 mg/L) | >1.4 | [39] |
| Pf | W2 strain | S. deserta Schang. | n.d. | taxodione (18) | C20H26O3 | 314.19 | 9.66 μM (3.036 mg/L) | >1.6 | [39] |
| Pf | D6 strain | S. deserta Schang. | n.d. | ferruginol (15) | C20H30O | 286.23 | 5.64 μM (1.616 mg/L) | 0.8 | [39] |
| Pf | W2 strain | S. deserta Schang. | n.d. | ferruginol (15) | C20H30O | 286.23 | 6.35 μM (1.817 mg/L) | 0.7 | [39] |
| Pf | D6 strain | S. deserta Schang. | n.d. | 7-o-acetylhorminone (30) | C22H30O5 | 374.21 | >12.72 μM (>4.760 mg/L) | 1 | [39] |
| Pf | W2 strain | S. deserta Schang. | n.d. | 7-o-acetylhorminone (30) | C22H30O5 | 374.21 | >12.72 μM (>4.760 mg/L) | 1 | [39] |
| Pf | D6 strain | S. deserta Schang. | n.d. | horminone (31) | C20H28O4 | 332.20 | >14.33 μM (>4.760 mg/L) | 1 | [39] |
| Pf | W2 strain | S. deserta Schang. | n.d. | horminone (31) | C20H28O4 | 332.20 | >14.33 μM (>4.760 mg/L) | 1 | [39] |
| Pf | FCR3 strain | S. radula Benth. | 3.91 µg/mL | betulafolientriol oxide (43) | C30H52O4 | 476.39 | 10.39 μM (4.95 µg/mL) | >20 | [48] |
| Pf | K1 strain | S. hydrangea DC. ex. Benth. | n.d. | hydrangenone (44) | C30H42O5 | 482.30 | 1.4 µM | 6 | [49] |
| Pf | FCR3 strain | S. radula Benth. | 3.91 µg/mL | salvigenin (45) | C18H16O6 | 328.09 | 74.98 μM (24.60 µg/mL) | >4 | [48] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Llurba-Montesino, N.; Schmidt, T.J. Salvia Species as Sources of Natural Products with Antiprotozoal Activity. Int. J. Mol. Sci. 2018, 19, 264. https://doi.org/10.3390/ijms19010264
AMA Style
Llurba-Montesino N, Schmidt TJ. Salvia Species as Sources of Natural Products with Antiprotozoal Activity. International Journal of Molecular Sciences. 2018; 19(1):264. https://doi.org/10.3390/ijms19010264
Chicago/Turabian StyleLlurba-Montesino, Núria, and Thomas J. Schmidt. 2018. "Salvia Species as Sources of Natural Products with Antiprotozoal Activity" International Journal of Molecular Sciences 19, no. 1: 264. https://doi.org/10.3390/ijms19010264
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.