Optogenetic Investigation of Arousal Circuits


Department of Psychiatry and Behavioral Sciences, Stanford University, 1201 Welch Road, Stanford, CA 94305, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(8), 1773; https://doi.org/10.3390/ijms18081773
Submission received: 16 July 2017
/
Revised: 6 August 2017
/
Accepted: 9 August 2017
/
Published: 15 August 2017
(This article belongs to the Special Issue Optogenetic Approaches in Neuroscience)
{kind=link}
Abstract
:Modulation between sleep and wake states is controlled by a number of heterogeneous neuron populations. Due to the topological proximity and genetic co-localization of the neurons underlying sleep-wake state modulation optogenetic methods offer a significant improvement in the ability to benefit from both the precision of genetic targeting and millisecond temporal control. Beginning with an overview of the neuron populations mediating arousal, this review outlines the progress that has been made in the investigation of arousal circuits since the incorporation of optogenetic techniques and the first in vivo application of optogenetic stimulation in hypocretin neurons in the lateral hypothalamus. This overview is followed by a discussion of the future progress that can be made by incorporating more recent technological developments into the research of neural circuits.
1. Introduction
Before the development of optogenetic tools researchers used methods such as electrophysiological stimulation and recording of neural activity, pharmacological stimulation or inhibition of targeted neuron populations, or the manipulation of gene expression through use of knockout animals to investigate the function and activation patterns of different neural populations. While these methods have undoubtedly produced an immense body of literature and insight into neural functions, structures, and mechanisms across multiple areas of neuroscience research, as new technologies have developed, the weaknesses of these techniques have naturally become highlighted. While electrophysiology allows the temporal precision that is lacking in pharmacological and genetic manipulation, it lacks the ability to target specific neuron populations using their gene expression. Pharmacological and genetic manipulations allow this neuron targeting specificity but lack the temporal flexibility of electrophysiological recordings, either requiring time for the pharmacological agent to peak or washout, or, in genetically manipulated mice, lacking the ability to observe the effect of the gene knockout during specific development periods, or even during specific behaviors.
The development of optogenetic tools has provided researchers with a method to genetically target specific neuron populations with millisecond precision, either for recording (fiber-photometry) or stimulation (optogenetic activation) of neural activity. The advances provided by this technique have allowed researchers to combine the gene-targeting specificity of pharmacological and genetic manipulations with the temporal flexibility of electrophysiological recordings. While the use of electrophysiological recording and stimulation methods is still very useful for applications where the genetic profile of the target neurons are not yet known or are not uniform, research areas in which it is known which neurotransmitters and cell types are involved can benefit greatly from this specificity. One area of research that has benefited greatly from this precision and flexibility is the investigation of neural circuits underlying arousal and transitions between sleeping and waking states. This review will focus on the neural correlates of sleep-wake state modulation, giving an overview of the neural populations involved, and presenting the progress that has been made following the incorporation of optogenetic technologies into the investigation of individual neuron populations, as well as circuit integration.
2. The Importance of Sleep
Considering the ubiquity of sleep across the animal kingdom, the precise function and purpose of sleep remains relatively elusive; however, it is apparent that effectively regulating arousal is crucial for survival. As well as maintaining an appropriate sleep/wake cycle we must also be able to respond appropriately to unexpected stimuli, whether they be an unanticipated reward or the sudden presentation of a stressor. The ability to focus and react can mean the difference between life and death. In states of disordered arousal we may exhibit hyper-arousal—the inability to drop out of a state of heightened vigilance resulting in anxiety disorders, insomnia, etc. or if we are unable to successfully maintain stable sleep-wake states this can lead to either an inability to maintain prolonged sleep or prolonged wakefulness. In order to understand what is going wrong in sleep disorders, researchers first had to understand what normal, healthy sleep-wake states look like.
Defining Different Sleep-Wake States
The first step away from a binary “sleep” versus “wake” definition of sleep-wake states came with the observation of what is now called Rapid Eye Movement (REM) sleep, in which a sleeping subject elicits eye movements and electroencephalogram (EEG) activity that is similar to those elicited from an awake subject. REM sleep was first observed by Aserinsky and Kleitman [1], and this observation lead to the categorization of sleep into two states: REM and non-REM (NREM). In order to study the neurocircuitry underlying different these sleep-wake states it is first important to be able to characterize them. For this purpose sleep-wake states have been broadly categorized into four different states: active/motivated wakefulness, quiet wakefulness, NREM sleep, and REM sleep. In the laboratory, researchers use EEG and electromyogram (EMG) recordings to distinguish between these different sleep-wake states [2]. Broadly, sleep/wake states are categorized as:
- Quiet wakefulness—slower EEG frequencies such as α (8–14 Hz) and β (15–24 Hz) [6] and muscle activity observed in the EMG signal;
Alongside these sleep-wake states, further categorizations are possible to perform more fine-grained analysis of different sleep states, for example in humans NREM sleep can be further categorized into four different stages (I–VI) of increasing depth of sleep [2]. However for the sake of this review a broader categorization of REM, NREM, waking, and occasional specifications of active waking and hyper-arousal states will be used to discuss the neural mechanisms linked to these states.
3. Neural Correlates of Sleep-Wake Modulation
Researchers studying sleep to wake transitions have used many different methods to uncover the numerous neuron populations that are involved in modulating wakefulness and sleep states. Following the influenza epidemic of 1918, Economo [19] performed post-mortem examinations of the brain tissue of patients who had suffered excessive sleepiness after contracting the influenza virus. It was observed that this sleepiness was correlated to brain lesions in the posterior hypothalamus. As well as specific brain areas, human responses to medications and disease states have also implicated specific neuropeptides in the modulation of sleep and wake states. Post-mortem examinations have shown a substantial loss of hypothalamic hypocretin (also known as orexin) neurons in the brains of patients with narcolepsy [20,21], a sleep disorder characterized by excessive sleepiness. Whereas a role for dopamine in modulation of arousal states has been implicated following an observation that patients with Parkinson’s disease, which results in the degeneration of dopaminergic neurons in the substantia nigra (among other neurological changes), experience sleep disturbances such as excessive sleepiness during the day and difficulty sleeping through the night [22].
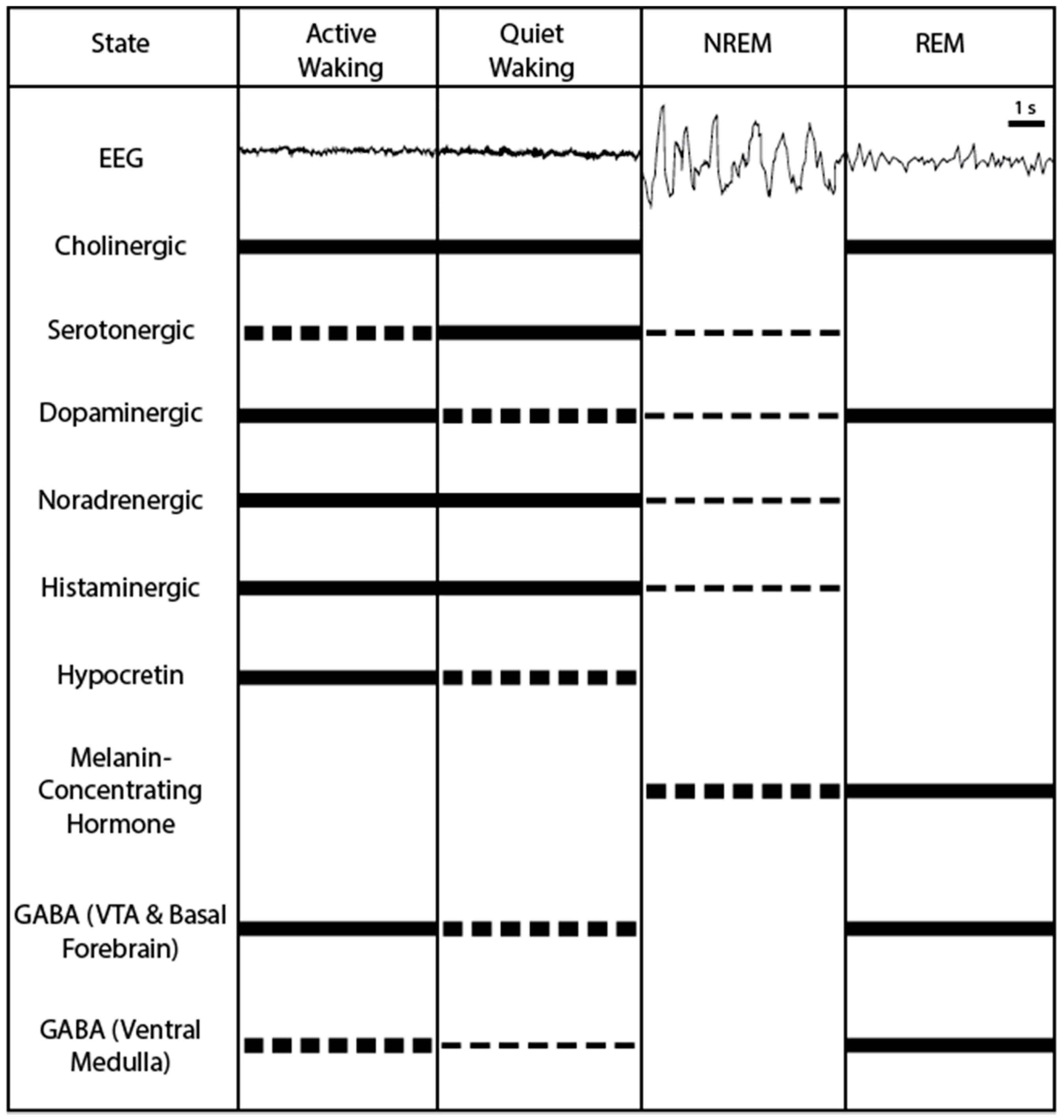
Pharmaceuticals have also highlighted potential candidate systems; particularly the development of first-generation antihistamines (histamine H1 receptor antagonists) and their sedative side effects implicated histamine as a wake-promoting neuropeptide [23,24]. In order to further investigate these neuropeptides and neural structures under more strictly controlled conditions researchers shifted into animals models to observe the mechanisms underlying sleep-wake state modulation on a more fine-grained, cellular level. Although there are some notable differences between humans and other animals when it comes to sleeping patterns, animal models of sleep have proved to be useful for further unraveling the neural circuits involved in modulating sleep and waking states. The use of animal models, particularly rodent models have resulted in the identification of multiple neuropeptides and structures that can be targeted to modulate sleep to wake states. These findings are reviewed below, with an overview of activity patterns of these candidate neurons across sleep-wake states shown in Figure 1.
3.1. Cholinergic Neurons: Active during Waking and REM Sleep
Cholinergic neurons have been shown to modulate arousal, in particular the basal forebrain and the mesopontine tegmentum of the brainstem contain neurons that release acetylcholine and modulate sleep-wake states [63,64]. Cholinergic neurons in the basal forebrain are most active during wakefulness and REM sleep [25,26,30], with a high correlation between their firing rate and cortical activation [25,65,66]. This pattern of wake-active and REM-active cholinergic neurons has also been observed in populations of cholinergic neurons found in the mesopontine tegmentum [31,32,33,34,35] and is supported by evidence of higher levels of acetylcholine in cortical and thalamic areas during wakefulness and REM sleep [27,28,29]. Recordings of cholinergic neurons have also shown that they fire in response to cortical activity occurring following tail-pinch in urethane-anaesthetized animals [67]. While these results suggest a role for cholinergic neurons in processes underlying wakefulness and REM sleep, lesion studies have returned mixed results. Some studies using selective lesions of cholinergic neurons in the basal forebrain only produced minor changes to the wakefulness of the animals [68,69,70], which would suggest that these neurons are not necessary to maintain wakefulness. However, more extensive lesions of cholinergic neurons in the basal forebrain resulted in a reduction of high-frequency EEG power, with a particularly noticeable reduction in γ-activity [71,72], which occurs during active-waking. These more extensive lesions also reduced the normal homeostatic responses to increased sleep-drive following sleep deprivation [73,74,75]. A pharmacological study which infused cholinomimetics into the pontine reticular formation, showed that increasing cholinergic activity in the reticular formation resulted in increased REM sleep [76], which would suggest a REM-sleep promoting role for brainstem cholinergic neurons. The heterogeneity of the basal forebrain and brainstem areas where these cholinergic neurons are found make these populations ideal candidates for genetically targeted optogenetic investigation to further explore these divergent functions of sleep-promoting basal forebrain cholinergic neurons, and REM-promoting brainstem cholinergic neurons.
3.2. Serotonergic Neurons: Promoting Wakefulness
Serotonergic interactions with sleep came into the spotlight due to the changes of sleep architecture seen in depressed patients undergoing pharmaceutical treatments affecting the serotonin system, such as a significant decrease in REM sleep duration in patients undergoing treatment with tricyclic antidepressant amitriptyline [77]. In line with this finding, systemic administration of serotonin receptor agonists produces a reduction in NREM and REM sleep and promote waking [78]. Additionally, while normal mice usually exhibit a boost in REM following immobilization stress, this REM rebound is not observed in 5-HT1A knockout mice [79]. Electrophysiological recordings have shown that serotonin is most active during waking, less active during NREM, and quiet during REM [80,81], these findings have also been supported by studies measuring the release of serotonin [37,38]. Serotonin neurons are activated by stressful stimuli [82], however they are most active during feeding and quiet waking, and are less active during active-waking [36], suggesting a possible role for serotonin in promoting a state of quiet-waking.
3.3. Noradrenergic Neurons: Promoting Wakefulness
The LC is the origin of the majority of noradrenergic projections of the forebrain, others arise from small clusters of norepinephrine (NE) neurons throughout the brainstem [83]. Administration of norepinephrine into the forebrain ventricles results in increased wakefulness [84,85]. LC neurons are most active during waking, particularly when the animal is exposed to stressful stimuli [41], they are less active during NREM sleep, and quiet during REM sleep [42,43,44]. Despite this clear link between LC/NE activity and wake states, lesion studies have produced minimal effects on total waking time in LC lesioned animals [86] and increases in REM sleep [87] and reduced expression of wake-related gene transcripts [88,89,90] following norepinephrine depletion. Discrepancies between the pharmacological studies, electrophysiological recordings, and lesion studies could suggest that while norepinephrine activation in the LC is sufficient to induce wakefulness, these neurons are not necessary to maintain wake-states.
3.4. Dopaminergic Neurons: Promoting Wakefulness
The role of dopamine in modulating wakefulness came into the spotlight due to the side effect of increased alertness following the consumption of certain drugs of abuse that are categorized as stimulants. Stimulants such as amphetamine and modafinil interact with other neuromodulatory systems, as well as dopaminergic neurons they also enhance the release of other wake-promoting neuropeptides serotonin and norepinephrine. In order to determine the main mechanism inducing increased alertness Wisor, et al. [91] used a dopamine transporter knockout mouse to show that animals lacking dopamine transporter do not exhibit the stimulant effects of modafinil or amphetamine, suggesting that transportation of dopamine is necessary for these stimulant effects. Interestingly, knockout studies have produced mixed results, with knockouts of the D2 receptor exhibiting bouts of waking, overall decreased waking amounts, and increased sleep duration [92], whereas dopamine transporter knockout mice exhibit reduced NREM sleep and increased wakefulness consolidation [91]. This suggests that dopamine may exhibit more nuanced modulation of sleep-wake states.
Dopaminergic (DA) neurons in both the ventral tegmental area (VTA) and the ventral periaqueductal gray matter (vPAG) have been investigated to observe their activity patterns across different sleep/wake states. Fos studies have shown that DA neurons in the vPAG are active during waking but not sleep states [93], however DA neurons in the VTA and the substantia nigra have a relatively consistent firing rate across all sleep-wake states [94]. An observation of burst-activity revealed that, despite the consistency of firing rates in the VTA, there were a higher number of DA neurons firing bursts in waking and REM sleep as opposed to NREM sleep, which caused increased levels of DA in the prefrontal cortex, nucleus accumbens, and other targets of VTADA neurons [39]. Additionally, Steinfels, et al. [95] showed that increased in DA bursting correlated strongly with salient stimuli that were either aversive or rewarding, suggesting that VTADA neurons may play a role in the directing of attention. The case for DA modulation of arousal is also supported by lesion studies in the vPAG, which showed a ~20% reduction in waking duration [93]. The variations in activity profiles suggest that these wake-active vPAGDA neurons are performing separate functions to the consistently active VTADA neurons that elicit increased burst activity during waking and REM states.
3.5. Neuropeptide S: Promoting Wakefulness
Neuropeptide S (NPS) precursor mRNA (messenger ribonucleic acid) is most strongly expressed around the locus coeruleus (LC), in the the principal sensory 5 nucleus, and in the the lateral parabrachial nucleus, and its receptor mRNA has highest expression levels in the cortex, thalamus, hypothalamus, and amygdala [96]. NPS has been proposed as a mediator of anxiety-related behaviors following evidence that intracerebroventricular (i.c.v.) administration of NPS reduces behavioral markers of anxiety in standard tests including the elevated plus maze and open field [96]. Additionally, considering that i.c.v. NPS administration also produces increased locomotion and reduced the amount of time animals spent in paradoxical REM sleep and the deepest stage of NREM sleep known as slow wave sleep [96]. Alternatively, knocking out the NPS receptor resulted in a decrease explorative behaviors in a novel environment [97], suggesting increased anxiety. Due to these findings it has been suggested that NPS may play a role in modulating states of hyper-vigilance such as fear or anxiety [96,98]. Due to the proximity of NPS to other neurons known to modulate sleep-wake states, such as noradrenergic neurons in the locus coeruleus and neuron populations in the parabrachial nucleus, these pharmacological methods have been used to avoid conflating the effects of unintentional activation of these neighboring populations to NPS.3.6. Histaminergic neurons: promoting wakefulness.
The link between histamine and wake-promotion was first suggested due to the side effect of drowsiness caused by first-generation antihistamines, which antagonized histamine H1 receptors [23,24]. A wake-promoting population of histaminergic neurons is located in the tuberomammillary nucleus, these neurons are most active during waking, less active during NREM sleep, and quiet during REM sleep [45,46,47]. Despite the obvious sedative effect of H1 receptor antagonists, as well as subsequent studies showing that pharmacological inhibition of the histamine system produces drowsiness whereas pharmacological activation of the histamine system promotes wakefulness [23,99], other investigations of histaminergic modulation of wakefulness have produces mixed results. Lesion studies in the cat [100] and the rat [101] produced no effect on sleep/wake duration, suggesting that this system is not critical for the maintenance of sleep/wake architecture. H1 receptor knockout mice exhibited similar sleep/wake durations to controls, only showing fewer micro-arousals (<16 s brief awakening), and fewer transitions between NREM sleep and REM sleep, but no difference in overall sleep/wake duration [102]. Investigation of the histamine system using a knockout model where the histamine-synthesizing enzyme histidine decarboxylase is knocked out (HDC KO) did not change the duration of sleep/wake states under baseline conditions, however HDC KO animals did show decreased wakefulness in a novel environment paradigm [103,104], suggesting a possible role for histamine in hyper-vigilance required in the presence of novel stimuli [23].
3.6. Hypocretin Neurons: Promoting Wakefulness
Hypocretin (Hcrt) neurons are found in the lateral hypothalamus and were first linked to sleep function due to a significant loss of Hcrt neurons in the brains of patients with narcolepsy [20,21] and low levels of Hcrt-1 in the cerebrospinal fluid (CSF) of narcoleptic patients [105]. The role of Hcrt in narcolepsy was further supported by animal models such as canine narcolepsy, which was linked to a mutation in the Hcrt 2 receptor [106]. Similarly, mouse models, which present both behavioral and EEG signs of narcolepsy, have been developed by manipulating the Hcrt system: either by targeting the neuropeptide in a Hcrt knockout mouse line [107] or by genetically ablating the neurons themselves [108].
Studies investigating the role of Hcrt in sleep modulation in non-diseased mouse models have found evidence for a wake-promoting role for Hcrt; Hcrt 1 administration (i.c.v.) increases wakefulness in a dose-dependent manner in rats [109]. Conversely, pharmacological antagonism of the Hcrt receptors results in increased NREM and REM sleep and reduced wakefulness in both animals and humans [110], and inhibition of Hcrt neurons using Designer Receptors Exclusively Activated by Designer Drugs (DREADDs) promotes sleep [111]. Hcrt neurons are most active during active-waking, less active during quiet-waking, and are quiet during sleep, this has been shown in Fos studies [48,49], measurements of Hcrt peptide release [50], and single-unit recordings [51,52,53,54,55]. Additionally, Hcrt levels have been shown to peak toward the end of the day in squirrel monkeys, whose sleep-cycle is similar to that of humans, suggesting that as the wake-cycle progresses wake-promoting Hcrt activity increases, perhaps in opposition to an increased need to rest [56]. Taken together along with the evidence from narcoleptic patients showing reduced ability to maintain wakefulness, these findings suggest a role for Hcrt in maintaining wakefulness, particularly toward the end of the waking-period of the animal.
While considering the role of different neuropeptides in modulating sleep-wake states across the day/night cycle, researchers must also consider how outside influences on sleep behaviors interact with these neurotransmitters. The two strongest determinants of sleeping behaviors are time of day and food availability, humans generally adhere to an active light-period and a quiet dark-period. EEG rhythms that are used to distinguish between sleep-wake states can oscillate according to these 24 h light–dark cycles, for example hippocampal θ rhythm, most often associated with waking states, is modulated on a circadian rhythm and can be entrained by food availability [112]. Considering evidence that genetic ablation of Hcrt neurons results in reduced mRNA expression of numerous clock genes in the forebrain and restricted feeding can shift the peak of Hcrt activity [113], it is possible that Hcrt may play a role in mediating food-related shifts in circadian rhythms, and thus effect sleeping behaviors. This idea is supported by evidence suggesting that nutritional status can affect Hcrt activity, in particular, wake-active Hcrt neurons are inhibited following food-intake and Hcrt neuron activity increases during periods of fasting in non-human primates [114]. Additionally, normal behavioral responses to fasting, such as increased waking and foraging behaviors, are not observed in mice with a genetic ablation of Hcrt neurons [113,115,116], suggesting that Hcrt is not only involved in, but also necessary for these behavioral responses to food availability. Mechanistically there are many candidates to investigate how Hcrt activity is modulated by food intake, Hcrt neurons interact with multiple biomarkers related to nutritional state, and in particular appear to be inhibited by biomarkers whose release is triggered by food intake, such as leptin, glucose, and neuropeptide Y [115,116,117,118,119,120]. Taken together these findings suggest a role for Hcrt in modulating not only sleep-wake transitions, but also more general sleeping behaviors such shifting circadian rhythms in response to changes in food availability or nutritional status.
3.7. Melanin-Concentrating Hormone Neurons: Promoting REM Sleep
Melanin-concentrating hormone (MCH) is another sleep-relevant neuron population located in the hypothalamus. MCH appears to have an opposing function to the other hypothalamic population of Hcrt neurons, while Hcrt promotes wakefulness it appears that MCH plays a role in promoting sleep, particularly REM sleep. Fos studies have shown that MCH neurons are most responsive after REM sleep [57,58]. This finding has been supported by a study using cellular recordings, which observed that MCH neurons only fire during REM sleep [59]. Observation of MCH levels across the sleep-wake cycle in the amygdala of humans also showed that the onset of sleep produces an increase in MCH levels [60]. In order to further explore the role of MCH in sleep-wake states researchers have also performed a variety of manipulation studies such as administration of MCH (i.c.v.) at the beginning of the light period (sleep phase for rodents), which showed a dramatic ~300% increase in the amount of REM sleep, and a slightly lower~150% increase in NREM sleep, as opposed to controls [58]. Concordantly, administration of an MCH antagonist decreases the amount of REM and NREM sleep [121], together these findings support the idea of MCH being a sleep-promoting neuropeptide. These findings would suggest that any sleep-relevant function of MCH neurons must be involved in REM sleep, however studies using genetic manipulation to investigate MCH function have returned mixed results. MCH receptor 1 knockout animals showed no change in sleep levels and no change in sleep rebound following a sleep deprivation paradigm [122], whereas MCH knockout mice showed minor decreases in sleep duration at baseline conditions, but showed a significant decrease in REM sleep and an increase in behavioral hyperactivity in response to fasting [123]. This link to fasting has also been supported by in vitro evidence that MCH neurons can be excited by increased circulating glucose levels [124]. This evidence for a modulatory effect of MCH function could suggest a possible mechanism for the modulatory effect of nutritional status on sleep function and appears to be in opposition to the effects seen in Hcrt neurons.
3.8. Glutamatergic and GABAergic Neurons
Alongside the other neuropeptides mentioned researchers must also consider the contribution of glutamatergic neurons and γ-amino butyric acidergic (GABAergic) neurons to sleep-wake states. These neuron types are found in all of the brain areas previously discussed [125,126,127,128,129,130,131], and in some cases are co-expressed in the same neurons with these other neuropeptides. For example, Vincent, Hokfelt, Skirboll and Wu [130] observed neurons co-expressing GABA and histamine in the hypothalamus, this has also been observed with GABA and MCH [129], and it has been suggested that glutamate may be a co-transmitter of Hcrt neurons in the hypothalamus [132], NPS neurons near the LC [96,133], and serotonin neurons in the dorsal raphe nuclei [134]. Additionally, animals with Vglut2 (vesicular glutamate transporter 2) deletion in the lateral parabrachial nucleus exhibited impaired ability to arouse from sleep and animals with the same treatment in the medial parabrachial nucleus exhibited shorter waking periods (~20%) and longer NREM duration (~43%) [135], suggesting a role for glutamatergic parabrachial nucleus neurons in modulating sleep–wake states.
Within the hypothalamus it has been shown that a population of wake-active neurons will increase their firing rate in response to antagonism of GABAA receptors [136], whereas GABAergic neurons in the BF and VTA show increased firing during waking and REM sleep [26,137]. It is thought that these VTAGABA neurons in particular may be involved in directing arousal linked to the reward system following evidence that their firing increases before intracranial self-stimulation in the medial forebrain bundle [61]. GABAergic neurons in the preoptic area have also been put forward as a candidate for mediating sleep-wake states. Electrophysiological studies have shown that these preoptic area neurons are predominantly sleep-active, and lesions of the ventrolateral preoptic area results in insomnia [138]. Additionally, GABAergic neurons have also been shown to project to the lateral hypothalamic area from the ventrolateral preoptic area and median preoptic nucleus [139]. These opposing actions of the hypothalamus and the preoptic area, coupled with the synaptic connections between the two structures have lead researchers to propose a possible state-switch mechanism, whereby the sleep-promoting preoptic area and the wake-promoting hypothalamus communicate to effectively switch between sleep-wake states [43]. The heterogeneity of these neuron populations makes functional investigation using traditional methods such as lesions problematic, as it is difficult to know if the effects are due to lesioning/recording/stimulating the other sleep–wake-relevant neuron populations that are known to be neighboring, or even co-expressed within, the neurons being targeted.
4. Optogenetic Investigation of Individual Neuron Populations
The development of optogenetics has revolutionized neuroscience research. It has been particularly useful for studying heterogeneous neuron populations in areas like the hypothalamus where many neuropeptides may be co-expressed within the same cell. Within the hypothalamus investigation of Hcrt and MCH neurons were made particularly difficult using traditional research methods due to their proximity and their apparent duality of function [59]. The gene-targeting specificity permitted by optogenetics has allowed researchers to overcome these problems studying sleep circuits.
4.1. Hypocretin Neurons
The first illustration of in vivo optogenetic manipulation was an investigation of Hcrt activation in the lateral hypothalamus (LH), which showed that optogenetic activation (5–30 Hz) of Hcrt neurons increases the probability of a sleep-wake transition from either NREM or REM sleep [140]. This effect was apparent throughout both light and dark phases unless the animal was placed in a 2 or 4 h sleep-deprivation paradigm resulting in increased sleep pressure [141]. Taken together these results suggest that Hcrt activity may modulate sleep-wake transitions according to homeostatic sleep-need. While these optogenetic stimulation studies have investigated the effect of activating Hcrt neurons [142,143] have also investigated the effect of optogenetically silencing Hcrt neurons using two transgenic mouse lines. Optogenetic inhibition of Hcrt neurons during the sleep-phase in mice expressing halorhodopsin in Hcrt neurons resulted in EEG and EMG patterns characteristically seen during NREM sleep, whereas no effect was seen following inhibition during the wake-phase [142]. Using a second transgenic mouse line with archaerhodopsin expressed in Hcrt neurons, [143] found that 1 h of Hcrt inhibition during the wake-phase can increase the total amount of time spent sleeping and reduce the total waking-time. These experiments have shown that stimulating Hcrt neurons can increase the likelihood of transitions into wakefulness, whereas inhibition of these neurons increases the likelihood of transitions into sleeping states.
4.2. Melanin-Concentrating Hormone Neurons
Optogenetic techniques have also been used to investigate MCH neurons in the hypothalamus showing that optogenetic activation of LHMCH neurons at the start of the dark phase (waking period) produces a dramatic increase in sleep duration, NREM sleep by 60% and REM sleep by 95% across the first 6 h [144]. Whereas optogenetic activation of MCH neurons during NREM sleep did not change the duration of NREM sleep but instead increased the probability of a transition from NREM sleep to REM sleep, and MCH neuron activation at the onset of REM sleep resulted in longer periods of REM sleep [145,146]. In order to investigate the downstream structures involved in this modulation of REM sleep, Jego, Glasgow, Herrera, Ekstrand, Reed, Boyce, Friedman, Burdakov and Adamantidis [145] also used optogenetics to target MCH fibers innervating the medial septum and tuberomammillary nucleus and found that activating these innervating fibers produced the same REM extension exhibited following optogenetic stimulation of the MCH neurons themselves. These results support the role of MCH in promoting REM sleep.
4.3. Dopaminergic Neurons
Optogenetic investigation of DA neurons in the VTA has shown that semi-chronic stimulation of VTADA neurons over 6 h is sufficient to maintain wakefulness and inhibit sleep-relevant behaviors such as nest building [40]. Conversely, when these neurons were inhibited animals exhibited nest building behaviors, suggesting that quieting these neurons triggers behaviors linked to sleep preparation [40]. Additionally, using fiber photometry to observe fluorescent calcium signals emitted by activation of Th+ VTA neurons, Eban-Rothschild, Rothschild, Giardino, Jones and de Lecea [40] observed that these neurons are more active during waking and REM sleep than NREM sleep, showed a decrease in activity immediately prior to wake-to-NREM transitions, and showed an increase in activity prior to either NREM-to-REM transitions or NREM-to-wake transitions, as had been shown previously [39]. In line with the results of inhibiting these neurons it was also observed that these Th+ VTA neurons were quiet during nest building behaviors [40]. Similarly, investigation of DA neurons in the dorsal raphe nucleus with fiber photometry showed increased activity to both rewarding and aversive salient stimuli, and increased activation during waking and REM sleep compared to NREM [147]. Similarly to the VTA, optogenetic activation of dorsal raphe nucleus (DRN) DA neurons promotes wakefulness, whereas their inhibition promotes sleeping [147]. Interestingly, fiber photometry results showed that VTADA neurons appeared to have increased activity during REM sleep compared to waking states [40], whereas DRNDA showed higher activation during waking states compared to REM sleep, especially during the early stages of waking, showing a gradual decrease in activity across the waking phase [147]. Considering previous evidence from measurements of Hcrt1 in CSF suggesting that wake-promoting Hcrt neurons activity peaks in the latter third of the day [56], it is possible that different wake-promoting neurons populations modulate wake-states throughout the day, with DRNDA neurons promoting wakefulness in the earlier stages of the waking period, and Hcrt neurons taking over modulation of arousal as the wake-phase progresses.
4.4. Cholinergic Neurons
Wake- and REM-active cholinergic neurons are found in the basal forebrain [25,26,30] and the mesopontine tegmentum [31,32,33,34,35]. Due to the heterogeneity of these areas results from lesion studies and electrophysiological stimulation studies have produced mixed results (as discussed in Section 3), however the use of optogenetics has provided more precise targeting of these neurons. Optogenetic activation of cholinergic basal forebrain neurons increases NREM-wake transitions, resulting in increased waking-duration and decreased NREM-duration [30], suggesting that these are a population of wake-promoting neurons. Interestingly, optogenetic activation of cholinergic neurons in the pedunculopontine tegmentum or the laterodorsal tegmentum increase the likelihood of REM sleep, and this increase in REM sleep is due to a greater number of REM sleep episodes, rather than an increase in the duration of the REM sleep episodes [148]. This suggests that cholinergic neurons in the pedunculopontine tegmentum and laterodorsal tegmentum are playing a role in the initiation of REM sleep episodes, rather than the maintenance of REM sleep episodes. These results suggest a wake-promoting role for cholinergic neurons in the basal forebrain, and a REM-initiation-promoting role for cholinergic neurons in the mesopontine tegmentum.
4.5. GABAergic Neurons
As previously stated, GABAergic neurons are located throughout the central nervous system and are co-localized with many different sleep-relevant neuropeptides. Optogenetic activation of GABAergic neurons found in the ventral medulla has been shown to rapidly and reliably induce REM sleep, or extend REM sleep episodes when stimulated during an already initiated REM sleep episode, whereas pharmacogenetic inhibition of these neurons results in dose-dependently reduced REM sleep [62]. Genetically targeted optrode recordings also showed that these ventral medulla GABAergic neurons are most active during REM sleep, and during wakefulness they responded preferentially to feeding and grooming behaviors [62]. Interestingly, optogenetic activation of GABAergic neurons in the bed nucleus of the stria terminalis (BdNST) during NREM sleep actually triggers transitions into wakefulness, however stimulating these neurons during REM sleep produces no transition from REM sleep [149]. Additionally, investigation of a group of GABAergic neurons in the LH that are known to project to the ventrolateral preoptic area, and do not appear to co-express either Hcrt or MCH showed that chemogenetic activation of these neurons produces increased waking-duration, whereas their inhibition causes increased sleep-duration [150]. Conversely, optogenetic activation of a population of GABAergic neurons in the preoptic area that project to the tuberomammillary nucleus resulted in increased NREM and REM sleep, and decreased waking, whereas their inhibition increased waking and decreased both NREM and REM sleep duration [151]. Taken together this suggests that LH and BdNST GABAergic neurons promote wakefulness, whereas preoptic area GABAergic neurons promote both NREM and REM sleep, and ventral medulla GABAergic neurons promote REM sleep.
A recent optogenetic investigation of the basal forebrain studied the role of two GABAergic neuron populations in sleep/wake states. It was observed that parvalbumin-positive GABAergic neurons are primarily wake- and REM-active, and optogenetic activation of these neurons results in increased NREM-wake transitions, and an overall increase in the duration of waking states and decrease in the duration of NREM sleep [30]. Conversely, somatostatin-positive GABAergic neurons showed more activity during NREM sleep states, and activation of these neurons promotes NREM sleep [30]. Further investigation revealed that the parvalbumin-positive GABAergic neurons received inhibitory connections from the somatostatin-positive GABAergic neurons [30]. These results suggest a possible circuit for controlling sleep-wake transitions in the basal forebrain via different neuron populations.
4.6. Astrocytes
While DA, Hcrt, MCH, and GABAergic neurons are well accepted as being part of the sleep-wake state machine, recent studies have also taken advantage of optogenetic tools to investigate possible roles in sleep regulation in other non-neuron cell types in the brain. Pelluru, et al. [152] used the promoter for the astrocyte-specific cytoskeletal protein (GFAP) to optogenetically target astrocytes to investigate the possibility of astrocyte regulation of sleep-wake states. Although a range of stimulation frequencies were used (0, 5, 10, 30 Hz), only 10 Hz stimulation of astrocytes produced a significant decrease in waking and a significant increase in both in REM and NREM sleep duration [152]. This finding suggests a possible role for astrocytes in the maintenance of sleep-states; however further investigation is required to understand the precise role of these cells, and the importance of a 10 Hz stimulation frequency.
5. Optogenetic Tools for Circuit Investigation
While optogenetic tools have provided insight into each of these neuron populations individually, the most beneficial quality of these tools is the possibilities they present for circuit investigation. By combining optogenetic methods with other pharmacological and genetic approaches such as DREADDs, chemogenetic inhibition/excitation, or genetic mouse models researchers can observe how manipulating one part of the sleep–wake state circuitry affects the relevant upstream/downstream targets. The majority of studies using optogenetic methods to investigate these neuropeptides have focused on the wake promoting neurons, with a particular focus on interactions with Hcrt neurons.
5.1. Hypocretin Interactions with Histamine
Hcrt and histamine are both wake-active neuron populations and the possibility of some kind of sleep-wake state mediating circuit was suggested by in vitro evidence that Hcrt neurons excite histamine neurons [153], and reports of low histamine levels in the CSF of narcolepsy patients, particularly those who are not taking medications to regulate their narcolepsy [154,155]. Low levels of histamine were also observed in the brains of narcoleptic dogs [156]. Additionally, administration (i.c.v.) of Hcrt 1 promotes wakefulness, and this effect is attenuated by interruption of histamine H1 receptor function either by application of an H1 receptor antagonist [157,158] or altered in via gene manipulation in knockout mouse models [159], and patch clamp electrophysiological recordings have also shown that infusion of Hcrt increases the activity of histaminergic neurons [158]. Conversely, while the wake promoting effect of Hcrt 1 was reduced in histamine H1 receptor knockout mice [159], optogenetic stimulation of Hcrt neurons in histidine decarboxylase (HDC) knockout mice was sufficient to increase the probability of a sleep-wake transition [141]. These findings suggest that there is some interaction between the hypocretin system and the histamine system and their modulation of arousal.
To observe Hcrt activation of histamine neurons more closely Schone, et al. [160] combined optogenetic stimulation with in vitro slice electrophysiology to observe the response profiles of histamine neurons to optogenetic stimulation of Hcrt neurons. Hcrt neuron stimulation resulted in fast postsynaptic currents in histamine neurons and a robust connection of Hcrt neurons to histamine neurons was observed (~60% of histamine neurons received connections from ~40% of Hcrt neurons) [160]. Subsequent investigation of the role of the Hcrt receptor 2 (HcrtR2) in this connectivity revealed that Hcrt activation of histamine neurons could produce two distinct phases of histamine neuron firing: an initial fast transient firing peak which is unaffected by HcrtR2 antagonism and is followed by a slow firing phase that increases linearly during constant Hcrt activation and is abolished when HcrtR2 is blocked [161]. The same combination of methods was used to observe targets of tuberomammillary nucleus (TMN) histamine neurons following optogenetic stimulation of histamine neurons. Results showed that stimulating histamine neurons results in disinhibition of the wake-active ventrolateral TMN by decreasing inhibitory GABAergic inputs to the vlTMN, whereas histamine activation resulted in the inhibition of activity in the sleep-active ventrolateral preoptic nucleus (VLPO) [162]. These observations of two phases of histamine activation differentially affected by the blocking of HcrtR2 could explain inconsistencies in the role of histamine in Hcrt modulation of arousal seen in previous studies.
5.2. Hypocretin Interactions with the Locus Coeruleus
The densest afferent projections from Hcrt neurons project to the wake-promoting locus coeruleus [163] and in vivo studies have shown that Hcrt injections into the LC promote wakefulness and reduce REM sleep [164,165]. Additionally, while mice lacking the Hcrt receptor show narcolepsy-like symptoms, targeted restoration of Hcrt receptors in the LC can reduce symptoms of chronic sleepiness and fragmented wakefulness [166]. Similarly to Hcrt neurons, optogenetic low frequency stimulation of LC neurons during sleep induces immediate sleep-wake transitions, and in awake mice it triggers increases in activity and total waking-duration [167]. Additionally, c-Fos stainings have shown that optogenetic activation of Hcrt neurons in the LH results in increased neural activity in the LC [141]. To investigate how these neurons interact with the Hcrt system Carter, et al. [168] carried out a dual optogenetic approach in which they could simultaneously stimulate Hcrt neurons in the LH and either inhibit or stimulate LC neurons. Using this method they found that bilateral inhibition of LC neurons blocked the Hcrt-mediated sleep-wake transitions normally induced by optogenetic stimulation. Conversely, concomitant optogenetic stimulation of Hcrt neurons and LC neurons enhances Hcrt-mediated sleep-wake transitions. While the Hcrt neurons and LC neurons have similar wake-promoting effects, the millisecond precision allowed using optogenetics has allowed researchers to investigate the temporal distinctions between these neuron populations. While optogenetic stimulation of LC neurons triggers rapid, reliable sleep-wake transitions within ~5 s, Hcrt neurons act over a longer 10–30 s to induce arousal [169]. It is possible that, while Hcrt modulates sleep-wake transitions according to homeostatic need, the LC which is known to be active during exposure to stressful stimuli [41] might play a role in enhancing Hcrt modulation of arousal when the animal is in a state of hyper-vigilance.
5.3. Hypocretin Interactions with Leptin
While the majority of arousal and sleep–wake state research focuses on transitions between waking, NREM, and REM sleep due to their clear delineations, there is also the possibility of subcategorizing wake-states. Although there are not currently widely accepted criteria to define categories of wakefulness, states of quiet-waking and active-waking are discussed frequently as well as the state following presentation of salient stimuli to induce either exploratory behaviors (rewards) or stress responses (aversive/fearful) stimuli. Hypocretin has been linked to states of hyper-arousal such as stress in a study by Bonnavion, et al. [170], which showed that extended phasic high-frequency optogenetic stimulation of Hcrt neurons produced multiple physiological and behavioral markers of stress including elevated plasma corticosterone concentrations, increased heart rate, increased blood pressure, and reduced exploratory behavior in the open field task. Interestingly, in food deprived animals optogenetic stimulation of Hcrt neurons resulted in a 3-fold increase in corticosterone concentrations above that of mice fed ad libitum [170], suggesting that food deprivation heightened the stress-response induced by Hcrt activation.
Hcrt is inhibited following food intake [114] and by biomarkers that are released following food intake such as leptin, glucose, and neuropeptide Y [115,117,118,119,120]. Of these compounds it has been shown that leptin, which is released in response to dietary fat intake, is behaviorally anxiolytic [171,172]. LepRb neurons are also known to be present in the lateral hypothalamic area, and directly innervate Hcrt neurons in the LH [173]. By combining optogenetic activation of Hcrt with leptin infusion into the lateral hypothalamic area, Bonnavion, Jackson, Carter and de Lecea [170] showed that Hcrt-mediated increase in corticosterone that is enhanced by food restriction, is attenuated by leptin administration. Then, to observe the direct effect of leptin receptor activation they optogenetically activated LepRb neurons during a behavioral restraint stress paradigm, which is known to trigger increased corticosterone release. Results showed that activation of LepRb neurons resulted in a suppression of Hcrt activity and resulted in decreased corticosterone concentrations [170]. Taken together, these results show that nutritional status modulates physiological and behavioral markers of stress via the Hcrt system, and Hcrt-induced stress can be attenuated by leptin, which is endogenously released following dietary fat intake.
5.4. Hypocretin Interactions with Melanin-Concentrating Hormone (Feat. GABA)
Optogenetic methods have also been used to investigate the apparent opposing functions of wake-promoting Hcrt neurons [140] and REM-promoting MCH neurons [145,146] in the lateral hypothalamus. Gene-deletion studies have also provided evidence for contrasting functions of these two neuron populations: deletion of Hcrt neurons results in increased sleepiness (as seen in narcolepsy) and weight gain [107,108], whereas deletion of MCH neurons results in increased hyperactivity and reduced bodyweight [174,175,176]. Recent research investigating the interactions between these neurons using combinations of pharmacological manipulation, optogenetic manipulation, and network-level calcium imaging in in vitro slice recordings have shown that bath-application of Hcrt in the LH results in the activation of approximately 30% of MCH neurons [177]. Further investigation showed that optogenetic activation of Hcrt neurons resulted in a rapid reduction of firing in approximately 80% of the MCH neurons recorded [177], however this effect was attenuated following application of a GABAA receptor blocker (10 μm gabazine), suggesting a possible role for GABA in mediating interactions between MCH and Hcrt neurons [177]. Indeed, optogenetic activation of Hcrt neurons produces increased GABAergic tone in the LH (also seen following bath application of Hcrt [161]) and this effect is reduced following application of Hcrt receptor antagonists [177]. Taken together, these results show that Hcrt neural activity can mediate MCH neuron activity either by excitation or inhibition, and that these interactions may be mediated via GABAA receptors.
6. Further Developments and Future Directions
Optogenetic stimulation dramatically improved the ability of researchers to stimulate specific populations of neurons without unintentionally stimulating neighboring neuron populations. This specificity can also be achieved for the recording of neural activity of targeted neurons populations without inadvertently recording activity from other adjacent populations by using fiber-photometry [178,179]. The implementation of this method will be crucial for understanding endogenous function of neuron populations—without first observing the naturally occurring activity patterns of these neuron populations, the optogenetic stimulation of them will only allow researchers to understand what artificial activation of the neurons produces. Greater insight into the naturally occurring neural activity patterns of different types of neurons will be important for driving further studies manipulating these activity patterns using optogenetic stimulation.
The main improvement that optogenetic tools provide over traditional electrophysiological recording and stimulating methods is the ability to target specific neurons based on their gene-expression. However, this does not help if the population of neurons being studied is not yet genetically defined. Therefore, in order for researchers to receive the maximum benefit from these tools it is important to carry out a systematic characterization of the target population. Recent developments in next-generation sequencing methods now allow researchers to investigate gene-expression profiles of single neurons [180]. These methods will be extremely useful for defining subpopulations of neurons within highly heterogeneous structures, such as the hypothalamus, to allow greater specificity when genetically targeting neurons. In particular, the use of drop-seq to sequence the RNA of a single cell separated into nanoliter-sized droplets has now been used to successfully define 50 genetically distinct neuron types in the heterogeneous hypothalamic arcuate–median eminence complex [181]. These different methods are impressive when used alone, but the real benefits for circuit investigation are apparent when they are used in combination. A recent study by [151] investigated GABAergic neurons in the preoptic area, beginning with viral tracing methods, combined EEG and EMG sleep state monitoring, optogenetic manipulation, and optrode recordings they found a population of sleep-active, sleep-promoting GABAergic neurons in the preoptic area that project to the tuberomammillary nucleus. Further investigations identified multiple candidate markers found within the neurons by using cingle cell RNA sequencing and translating ribosome affinity purification. With these candidate genes [151] were then able to use further optogenetic and pharmacogenetic stimulation experiments to show a role for each of these gene candidates in promoting sleep. By following this example of using a combination of these techniques, researchers will be able to define their target neuron populations, observe the endogenous neuronal activity with fiber-photometry, and then optogenetically manipulate it.
7. Conclusions
While the optogenetic investigations discussed here have all taken great steps toward understanding the mechanisms underlying how the brain modulates between REM, NREM, quiet waking, active waking, and hyper-arousal states, there is still much to be learned. Each of the neural populations discussed here hold their own interest for the function of sleep and wake states, however, no behavior as complex as the modulation of sleep-wake transitions occurs in a single-structure vacuum, and it will only be by investigating circuits that researchers can finally comprehend the mechanisms via which animals can modulate sleep behaviors. Optogenetic technologies have vastly improved the ability of researchers to investigate the complex circuitry underlying sleep behaviors and these technologies will be fundamental in increasing our current knowledge of the sleep functions carried out in the brain. While much attention has been paid to optogenetic stimulation in particular, without first observing the endogenously occurring neuronal activation it would be impossible to successfully mimic neural activity using optogenetic stimulation. Therefore preliminary studies using optogenetic techniques such as fiber photometry should be taken advantage of to first observe the naturally occurring neural patterns researchers wish to recreate in the neurons expressing their neuropeptide of interest.
Acknowledgments
Susan M. Tyree is currently funded by the Philip Wrightson Postdoctoral Fellowship, awarded by the New Zealand Neurological Foundation. Luis de Lecea is supported by grants from the NIMH (5R01MH087592-05; R01MH102638-01A1), the NIH (1R01AG047671-01), the US-Israel Binational Science Foundation (BSF #2011335), Merck, and Johnson & Johnson. We thank Robert G. K. Munn and Shane A. Liddelow for helpful comments on this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BdNST | Bed nucleus of the stria terminalis |
| CSF | Cerebrospinal fluid |
| DA | Dopaminergic |
| DREADD | Designer receptors exclusively activated by designer drugs |
| DRN | Dorsal raphe nucleus |
| EEG | Electroencephalogram |
| EMG | Electromyogram |
| GABA | γ Amino butyric acid |
| GFAP | Glial fibrillary acidic protein |
| Hcrt | Hypocretin |
| HcrtR2 | Hypocretin receptor 2 |
| Hz | Hertz |
| i.c.v. | Intracerebroventricular |
| LC | Locus coeruleus |
| LH | Lateral hypothalamus |
| MCH | Melanin-concentrating hormone |
| mRNA | Messenger ribonucleic acid |
| NE | Norepinephrine |
| NPS | Neuropeptide S |
| NREM | Non-rapid eye movement |
| REM | Rapid eye movement |
| TMN | Tuberomammillary nucleus |
| Vglut2 | Vesicular glutamate transporter 2 |
| VLPO | Ventrolateral pre-optic nucleus |
| vlTMN | Ventrolateral tuberomammillary nucleus |
| vPAG | Ventral periaqueductal gray |
| VTA | Ventral tegmental area |
References
- Aserinsky, E.; Kleitman, N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science 1953, 118, 273–274. [Google Scholar] [CrossRef] [PubMed]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques, and Scoring Systems for Sleep Stages of Human Subjects; U.S. National Institute of Neurological Diseases and Blindness, Neurological Information Network: Maryland, MD, USA, 1969; Volum 26, p. 644.
- Huber, R.; Deboer, T.; Tobler, I. Prion protein: A role in sleep regulation? J. Sleep Res. 1999, 8, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.-P.; Rétey, J.V.; Tönz, K.; Gottselig, J.M.; Khatami, R.; Buckelmüller, I.; Achermann, P. Caffeine attenuates waking and sleep electroencephalographic markers of sleep homeostasis in humans. Neuropsychopharmacology 2004, 29, 1933. [Google Scholar] [CrossRef] [PubMed]
- Aeschbach, D.; Matthews, J.R.; Postolache, T.T.; Jackson, M.A.; Giesen, H.A.; Wehr, T.A. Two circadian rhythms in the human electroencephalogram during wakefulness. Am. J. Physiol. 1999, 277, R1771–R1779. [Google Scholar] [PubMed]
- Niedermeyer, E. The normal eeg of the waking adult. In Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Schomer, D.L., Lopes da Silva, F., Eds.; Lippincott Williams & Wilkins: Pennsyivania, PA, USA, 2005; Volum 9, pp. 155–164. [Google Scholar]
- Cirelli, C.; Tononi, G. Is sleep essential? PLoS Biol. 2008, 6, e216. [Google Scholar] [CrossRef] [PubMed]
- Franken, P.; Chollet, D.; Tafti, M. The homeostatic regulation of sleep need is under genetic control. J. Neurosci. 2001, 21, 2610–2621. [Google Scholar] [PubMed]
- Vyazovskiy, V.V.; Ruijgrok, G.; Deboer, T.; Tobler, I. Running wheel accessibility affects the regional electroencephalogram during sleep in mice. Cereb. Cortex 2006, 16, 328. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Tobler, I. Regional differences in NREM sleep slow-wave activity in mice with congenital callosal dysgenesis. J. Sleep Res. 2005, 14, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Harris, K.D. Sleep and the single neuron: The role of global slow oscillations in individual cell rest. Nat. Rev. Neurosci. 2013, 14, 443. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Achermann, P.; Bo rbely, A.A.; Tobler, I. The dynamics of spindles and eeg slow-wave activity in NREM sleep in mice. Arch. Ital. Biol. 2004, 142, 511–523. [Google Scholar] [PubMed]
- Lüthi, A. Sleep spindles: Where they come from, what they do. Neuroscientist 2014, 20, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Nir, Y.; Staba, R.J.; Andrillon, T.; Vyazovskiy, V.V.; Cirelli, C.; Fried, I.; Tononi, G. Regional slow waves and spindles in human sleep. Neuron 2011, 70, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Callaway, C.W.; Lydic, R.; Baghdoyan, H.A.; Hobson, J.A. Pontogeniculooccipital waves: Spontaneous visual system activity during rapid eye movement sleep. Cell. Mol. Neurobiol. 1987, 7, 105–149. [Google Scholar] [CrossRef] [PubMed]
- Datta, S. Cellular and chemical neuroscience of mammalian sleep. Sleep Med. 2010, 11, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Karashima, A.; Katayama, N.; Nakao, M. Enhancement of synchronization between hippocampal and amygdala θ waves associated with pontine wave density. J. Neurophysiol. 2010, 103, 2318–2325. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.M. Clues to the functions of mammalian sleep. Nature 2005, 437, 1264. [Google Scholar] [CrossRef] [PubMed]
- Economo, C.V. Sleep as a problem of localization. J. Nerv. Ment. Dis. 1930, 71, 249–259. [Google Scholar] [CrossRef]
- Peyron, C.; Faraco, J.; Rogers, W.; Ripley, B.; Overeem, S.; Charnay, Y.; Nevsimalova, S.; Aldrich, M.; Reynolds, D.; Albin, R.; et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat. Med. 2000, 6, 991–997. [Google Scholar] [PubMed]
- Thannickal, T.C.; Moore, R.Y.; Nienhuis, R.; Ramanathan, L.; Gulyani, S.; Aldrich, M.; Cornford, M.; Siegel, J.M. Reduced number of hypocretin neurons in human narcolepsy. Neuron 2000, 27, 469–474. [Google Scholar] [CrossRef]
- Rye, D.B. The two faces of eve: Dopamine’s modulation of wakefulness and sleep. Neurology 2004, 63, S2–S7. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.E.; Stevens, D.R.; Haas, H.L. The physiology of brain histamine. Prog. Neurobiol. 2001, 63, 637–672. [Google Scholar] [CrossRef]
- White, J.M.; Rumbold, G.R. Behavioural effects of histamine and its antagonists: A review. Psychopharmacology (Berl) 1988, 95, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.G.; Hassani, O.K.; Alonso, A.; Jones, B.E. Cholinergic basal forebrain neurons burst with θ during waking and paradoxical sleep. J. Neurosci. 2005, 25, 4365–4369. [Google Scholar] [CrossRef] [PubMed]
- Hassani, O.K.; Lee, M.G.; Henny, P.; Jones, B.E. Discharge profiles of identified GABAergic in comparison to cholinergic and putative glutamatergic basal forebrain neurons across the sleep–wake cycle. J. Neurosci. 2009, 29, 11828–11840. [Google Scholar] [CrossRef] [PubMed]
- Celesia, G.G.; Jasper, H.H. Acetylcholine released from cerebral cortex in relation to state of activation. Neurology 1966, 16, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Jasper, H.H.; Tessier, J. Acetylcholine liberation from cerebral cortex during paradoxical (rem) sleep. Science 1971, 172, 601–602. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A.; Comisarow, J.; Day, J.; Fibiger, H.C.; Reiner, P.B. State–dependent release of acetylcholine in rat thalamus measured by in vivo microdialysis. J. Neurosci. 1994, 14, 5236–5242. [Google Scholar] [PubMed]
- Xu, M.; Chung, S.; Zhang, S.; Zhong, P.; Ma, C.; Chang, W.C.; Weissbourd, B.; Sakai, N.; Luo, L.; Nishino, S. Basal forebrain circuit for sleep–wake control. Nat. Neurosci. 2015, 18, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- El Mansari, M.; Sakai, K.; Jouvet, M. Unitary characteristics of presumptive cholinergic tegmental neurons during the sleep–waking cycle in freely moving cats. Exp. Brain Res. 1989, 76, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Koyama, Y. Are there cholinergic and non-cholinergic paradoxical sleep-on neurones in the pons? Neuroreport 1996, 7, 2449–2454. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.J.; Highfield, D.A. Extracellular characteristics of putative cholinergic neurons in the rat laterodorsal tegmental nucleus. Brain Res. 1991, 559, 64–74. [Google Scholar] [CrossRef]
- Steriade, M.; Datta, S.; Pare, D.; Oakson, G.; Dossi, R.C.C. Neuronal activities in brain-stem cholinergic nuclei related to tonic activation processes in thalamocortical systems. J. Neurosci. 1990, 10, 2541–2559. [Google Scholar] [PubMed]
- Boucetta, S.; Cissé, Y.; Mainville, L.; Morales, M.; Jones, B.E. Discharge profiles across the sleep–waking cycle of identified cholinergic, GABAergic, and glutamatergic neurons in the pontomesencephalic tegmentum of the rat. J. Neurosci. 2014, 34, 4708–4727. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.L.; Fornal, C.A. Activity of brain serotonergic neurons in the behaving animal. Pharmacol. Rev. 1991, 43, 563–578. [Google Scholar] [PubMed]
- Wilkinson, L.O.; Auerbach, S.B.; Jacobs, B.L. Extracellular serotonin levels change with behavioral state but not with pyrogen-induced hyperthermia. J. Neurosci. 1991, 11, 2732–2741. [Google Scholar] [PubMed]
- Portas, C.M.; Bjorvatn, B.; Ursin, R. Serotonin and the sleep/wake cycle: Special emphasis on microdialysis studies. Prog. Neurobiol. 2000, 60, 13–35. [Google Scholar] [CrossRef]
- Dahan, L.; Astier, B.; Vautrelle, N.; Urbain, N.; Kocsis, B.; Chouvet, G. Prominent burst firing of dopaminergic neurons in the ventral tegmental area during paradoxical sleep. Neuropsychopharmacology 2007, 32, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Eban–Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. Vta dopaminergic neurons regulate ethologically relevant sleep–wake behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.; Morilak, D.A.; Jacobs, B.L. Single unit activity of locus coeruleus neurons in the freely moving cat. I. During naturalistic behaviors and in response to simple and complex stimuli. Brain Res. 1986, 371, 324–334. [Google Scholar] [CrossRef]
- Hobson, J.A.; McCarley, R.W.; Wyzinski, P.W. Sleep cycle oscillation: Reciprocal discharge by two brainstem neuronal groups. Science 1975, 189, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep state switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E. Arousal systems. Front. Biosci. 2003, 8, s438–s451. [Google Scholar] [CrossRef] [PubMed]
- Vanni-Mercier, G.; Gigout, S.; Debilly, G.; Lin, J.S. Waking selective neurons in the posterior hypothalamus and their response to histamine H3-receptor ligands: An electrophysiological study in freely moving cats. Behav. Brain Res. 2003, 144, 227–241. [Google Scholar] [CrossRef]
- John, J.; Wu, M.F.; Boehmer, L.N.; Siegel, J.M. Cataplexy-active neurons in the hypothalamus: Implications for the role of histamine in sleep and waking behavior. Neuron 2004, 42, 619–634. [Google Scholar] [CrossRef]
- Takahashi, K.; Lin, J.S.; Sakai, K. Neuronal activity of histaminergic tuberomammillary neurons during wake–sleep states in the mouse. J. Neurosci. 2006, 26, 10292–10298. [Google Scholar] [CrossRef] [PubMed]
- Estabrooke, I.V.; McCarthy, M.T.; Ko, E.; Chou, T.C.; Chemelli, R.M.; Yanagisawa, M.; Saper, C.B.; Scammell, T.E. Fos expression in orexin neurons varies with behavioral state. J. Neurosci. 2001, 21, 1656–1662. [Google Scholar] [PubMed]
- Modirrousta, M.; Mainville, L.; Jones, B.E. Orexin and MCH neurons express c-Fos differently after sleep deprivation vs. recovery and bear different adrenergic receptors. Eur. J. Neurosci. 2005, 21, 2807–2816. [Google Scholar] [CrossRef] [PubMed]
- Kiyashchenko, L.I.; Mileykovskiy, B.Y.; Maidment, N.; Lam, H.A.; Wu, M.F.; John, J.; Peever, J.; Siegel, J.M. Release of hypocretin (orexin) during waking and sleep states. J. Neurosci. 2002, 22, 5282–5286. [Google Scholar] [PubMed]
- Alam, M.N.; Gong, H.; Alam, T.; Jaganath, R.; McGinty, D.; Szymusiak, R. Sleep–waking discharge patterns of neurons recorded in the rat perifornical lateral hypothalamic area. J. Physiol. 2002, 538, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Takahashi, K.; Kodama, T.; Kayama, Y. State-dependent activity of neurons in the perifornical hypothalamic area during sleep and waking. Neuroscience 2003, 119, 1209–1219. [Google Scholar] [CrossRef]
- Lee, M.G.; Hassani, O.K.; Jones, B.E. Discharge of identified orexin/hypocretin neurons across the sleep–waking cycle. J. Neurosci. 2005, 25, 6716–6720. [Google Scholar] [CrossRef] [PubMed]
- Mileykovskiy, B.Y.; Kiyashchenko, L.I.; Siegel, J.M. Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron 2005, 46, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Lin, J.S.; Sakai, K. Neuronal activity of orexin and non-orexin waking-active neurons during wake–sleep states in the mouse. Neuroscience 2008, 153, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Buckmaster, C.L.; Parker, K.J.; Hauck, C.M.; Lyons, D.M.; Mignot, E. Circadian and homeostatic regulation of hypocretin in a primate model: Implications for the consolidation of wakefulness. J. Neurosci. 2003, 23, 3555–3560. [Google Scholar] [PubMed]
- Hanriot, L.; Camargo, N.; Courau, A.C.; Leger, L.; Luppi, P.H.; Peyron, C. Characterization of the melanin-concentrating hormone neurons activated during paradoxical sleep hypersomnia in rats. J. Comp. Neurol. 2007, 505, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Verret, L.; Goutagny, R.; Fort, P.; Cagnon, L.; Salvert, D.; Leger, L.; Boissard, R.; Salin, P.; Peyron, C.; Luppi, P.H. A role of melanin-concentrating hormone producing neurons in the central regulation of paradoxical sleep. BMC Neurosci. 2003, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Hassani, O.K.; Lee, M.G.; Jones, B.E. Melanin-concentrating hormone neurons discharge in a reciprocal manner to orexin neurons across the sleep–wake cycle. Proc. Natl. Acad. Sci. USA 2009, 106, 2418–2422. [Google Scholar] [CrossRef] [PubMed]
- Blouin, A.M.; Fried, I.; Wilson, C.L.; Staba, R.J.; Behnke, E.J.; Lam, H.A.; Maidment, N.T.; Karlsson, K.A.E.; Lapierre, J.L.; Siegel, J.M. Human hypocretin and melanin-concentrating hormone levels are linked to emotion and social interaction. Nat. Commun. 2013, 4, 1547. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, S.C.; Lee, R.S.; Stobbs, S.H.; Henriksen, S.J. Responses of ventral tegmental area GABA neurons to brain stimulation reward. Brain Res. 2001, 906, 190–197. [Google Scholar] [CrossRef]
- Weber, F.; Chung, S.; Beier, K.T.; Xu, M.; Luo, L.; Dan, Y. Control of REM sleep by ventral medulla GABAergic neurons. Nature 2015, 526, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.M.; Saper, C.B.; Levey, A.I.; Wainer, B.H.; Terry, R.D. Distribution of cholinergic neurons in rat brain: Demonstrated by the immunocytochemical localization of choline acetyltransferase. J. Comp. Neurol. 1983, 216, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Mesulam, M.M.; Mufson, E.J.; Wainer, B.H.; Levey, A.I. Central cholinergic pathways in the rat: An overview based on an alternative nomenclature (ch1–ch6). Neuroscience 1983, 10, 1185–1201. [Google Scholar] [CrossRef]
- Duque, A.; Balatoni, B.; Detari, L.; Zaborszky, L. EEG correlation of the discharge properties of identified neurons in the basal forebrain. J. Neurophysiol. 2000, 84, 1627–1635. [Google Scholar] [PubMed]
- Manns, I.D.; Alonso, A.; Jones, B.E. Rhythmically discharging basal forebrain units comprise cholinergic, GABAergic, and putative glutamatergic cells. J. Neurophysiol. 2003, 89, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Boucetta, S.; Jones, B.E. Activity profiles of cholinergic and intermingled GABAergic and putative glutamatergic neurons in the pontomesencephalic tegmentum of urethane-anesthetized rats. J. Neurosci. 2009, 29, 4664–4674. [Google Scholar] [CrossRef] [PubMed]
- Fisahn, A.; Pike, F.G.; Buhl, E.H.; Paulsen, O. Cholinergic induction of network oscillations at 40 Hz in the hippocampus in vitro. Nature 1998, 394, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Huerta, P.T.; Lisman, J.E. Heightened synaptic plasticity of hippocampal ca1 neurons during a cholinergically induced rhythmic state. Nature 1993, 364, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Shimono, K.; Brucher, F.; Granger, R.; Lynch, G.; Taketani, M. Origins and distribution of cholinergically induced β rhythms in hippocampal slices. J. Neurosci. 2000, 20, 8462–8473. [Google Scholar] [PubMed]
- Berntson, G.G.; Shafi, R.; Sarter, M. Specific contributions of the basal forebrain corticopetal cholinergic system to electroencephalographic activity and sleep/waking behaviour. Eur. J. Neurosci. 2002, 16, 2453–2461. [Google Scholar] [CrossRef] [PubMed]
- Kapas, L.; Obal, F., Jr.; Book, A.A.; Schweitzer, J.B.; Wiley, R.G.; Krueger, J.M. The effects of immunolesions of nerve growth factor-receptive neurons by 192 IgG-saporin on sleep. Brain Res. 1996, 712, 53–59. [Google Scholar] [CrossRef]
- Blanco-Centurion, C.; Xu, M.; Murillo–Rodriguez, E.; Gerashchenko, D.; Shiromani, A.M.; Salin-Pascual, R.J.; Hof, P.R.; Shiromani, P.J. Adenosine and sleep homeostasis in the basal forebrain. J. Neurosci. 2006, 26, 8092–8100. [Google Scholar] [CrossRef] [PubMed]
- Kalinchuk, A.V.; McCarley, R.W.; Stenberg, D.; Porkka-Heiskanen, T.; Basheer, R. The role of cholinergic basal forebrain neurons in adenosine–mediated homeostatic control of sleep: Lessons from 1 to 92 IgG-saporin lesions. Neuroscience 2008, 157, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Junek, A.; Black, M.A.; Semba, K. Effects of ibotenate and 192 IgG-saporin lesions of the nucleus basalis magnocellularis/substantia innominata on spontaneous sleep and wake states and on recovery sleep after sleep deprivation in rats. J. Neurosci. 2008, 28, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Shiromani, P.J.; Fishbein, W. Continuous pontine cholinergic microinfusion via mini-pump induces sustained alterations in rapid eye movement (REM) sleep. Pharmacol. Biochem. Behav. 1986, 25, 1253–1261. [Google Scholar] [CrossRef]
- Gillin, J.C.; Wyatt, R.J.; Fram, D.; Snyder, F. The relationship between changes in REM sleep and clinical improvement in depressed patients treated with amitriptyline. Psychopharmacology 1978, 59, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.M.; Jantos, H. The roles of dopamine and serotonin, and of their receptors, in regulating sleep and waking. Prog. Brain Res. 2008, 172, 625–646. [Google Scholar] [PubMed]
- Boutrel, B.; Monaca, C.; Hen, R.; Hamon, M.; Adrien, J. Involvement of 5-HT1A receptors in homeostatic and stress-induced adaptive regulations of paradoxical sleep: Studies in 5-HT1A knock-out mice. J. Neurosci. 2002, 22, 4686–4692. [Google Scholar] [PubMed]
- McGinty, D.J.; Harper, R.M. Dorsal raphe neurons: Depression of firing during sleep in cats. Brain Res. 1976, 101, 569–575. [Google Scholar] [CrossRef]
- Trulson, M.E.; Jacobs, B.L. Raphe unit activity in freely moving cats: Correlation with level of behavioral arousal. Brain Res. 1979, 163, 135–150. [Google Scholar] [CrossRef]
- Hammack, S.E.; Schmid, M.J.; LoPresti, M.L.; Der-Avakian, A.; Pellymounter, M.A.; Foster, A.C.; Watkins, L.R.; Maier, S.F. Corticotropin releasing hormone type 2 receptors in the dorsal raphe nucleus mediate the behavioral consequences of uncontrollable stress. J. Neurosci. 2003, 23, 1019–1025. [Google Scholar] [PubMed]
- Carlsson, A.; Falck, B.; Hillarp, N.A. Cellular localization of brain monoamines. Acta Physiol. Scand. Suppl. 1962, 56, 1–28. [Google Scholar] [PubMed]
- Flicker, C.; Geyer, M.A. The hippocampus as a possible site of action for increased locomotion during intracerebral infusions of norepinephrine. Behav. Neural. Biol. 1982, 34, 421–426. [Google Scholar] [CrossRef]
- Segal, D.S.; Mandell, A.J. Behavioral activation of rats during intraventricular infusion of norepinephrine. Proc. Natl. Acad. Sci. USA 1970, 66, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Harper, S.T.; Halaris, A.E. Effects of locus coeruleus lesions upon cerebral monoamine content, sleep–wakefulness states and the response to amphetamine in the cat. Brain Res. 1977, 124, 473–496. [Google Scholar] [CrossRef]
- Monti, J.M.; D’Angelo, L.; Jantos, H.; Barbeito, L.; Abo, V. Effect of DSP-4, a noradrenergic neurotoxin, on sleep and wakefulness and sensitivity to drugs acting on adrenergic receptors in the rat. Sleep 1988, 11, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Pompeiano, M.; Tononi, G. Neuronal gene expression in the waking state: A role for the locus coeruleus. Science 1996, 274, 1211–1215. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Tononi, G. Differential expression of plasticity-related genes in waking and sleep and their regulation by the noradrenergic system. J. Neurosci. 2000, 20, 9187–9194. [Google Scholar] [PubMed]
- Cirelli, C.; Tononi, G. Locus ceruleus control of state-dependent gene expression. J. Neurosci. 2004, 24, 5410–5419. [Google Scholar] [CrossRef] [PubMed]
- Wisor, J.P.; Nishino, S.; Sora, I.; Uhl, G.H.; Mignot, E.; Edgar, D.M. Dopaminergic role in stimulant-induced wakefulness. J. Neurosci. 2001, 21, 1787–1794. [Google Scholar] [PubMed]
- Qu, W.-M.; Xu, X.-H.; Yan, M.-M.; Wang, Y.-Q.; Urade, Y.; Huang, Z.-L. Essential role of dopamine D2 receptor in the maintenance of wakefulness, but not in homeostatic regulation of sleep, in mice. J. Neurosci. 2010, 30, 4382–4389. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jhou, T.C.; Saper, C.B. Identification of wake-active dopaminergic neurons in the ventral periaqueductal gray matter. J. Neurosci. 2006, 26, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Farber, J.; Gatz, P.; Roffwarg, H.; German, D.C. Activity of mesencephalic dopamine and non-dopamine neurons across stages of sleep and waking in the rat. Brain Res. 1983, 273, 133–141. [Google Scholar] [CrossRef]
- Steinfels, G.F.; Heym, J.; Strecker, R.E.; Jacobs, B.L. Behavioral correlates of dopaminergic unit activity in freely moving cats. Brain Res. 1983, 258, 217–228. [Google Scholar] [CrossRef]
- Xu, Y.L.; Reinscheid, R.K.; Huitron-Resendiz, S.; Clark, S.D.; Wang, Z.; Lin, S.H.; Brucher, F.A.; Zeng, J.; Ly, N.K.; Henriksen, S.J.; et al. Neuropeptide s: A neuropeptide promoting arousal and anxiolytic-like effects. Neuron 2004, 43, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Duangdao, D.M.; Clark, S.D.; Okamura, N.; Reinscheid, R.K. Behavioral phenotyping of neuropeptide s receptor knockout mice. Behav. Brain Res. 2009, 205, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jungling, K.; Seidenbecher, T.; Sosulina, L.; Lesting, J.; Sangha, S.; Clark, S.D.; Okamura, N.; Duangdao, D.M.; Xu, Y.L.; Reinscheid, R.K.; et al. Neuropeptide S-mediated control of fear expression and extinction: Role of intercalated GABAergic neurons in the amygdala. Neuron 2008, 59, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.S. Physiological review article: Brain structures and mechanisms involved in the control of cortical activation and wakefulness, with emphasis on the posterior hypothalamus and histaminergic neurons. Sleep Med. Rev. 2000, 4, 471–503. [Google Scholar] [CrossRef] [PubMed]
- Denoyer, M.; Sallanon, M.; Buda, C.; Kitahama, K.; Jouvet, M. Neurotoxic lesion of the mesencephalic reticular formation and/or the posterior hypothalamus does not alter waking in the cat. Brain Res. 1991, 539, 287–303. [Google Scholar] [CrossRef]
- Gerashchenko, D.; Chou, T.C.; Blanco-Centurion, C.A.; Saper, C.B.; Shiromani, P.J. Effects of lesions of the histaminergic tuberomammillary nucleus on spontaneous sleep in rats. Sleep 2004, 27, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.L.; Mochizuki, T.; Qu, W.M.; Hong, Z.Y.; Watanabe, T.; Urade, Y.; Hayaishi, O. Altered sleep–wake characteristics and lack of arousal response to h3 receptor antagonist in histamine h1 receptor knockout mice. Proc. Natl. Acad. Sci. USA 2006, 103, 4687–4692. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, R.; Ohtsu, H.; Djebbara-Hannas, Z.; Valatx, J.L.; Watanabe, T.; Lin, J.S. Anatomical, physiological, and pharmacological characteristics of histidine decarboxylase knock-out mice: Evidence for the role of brain histamine in behavioral and sleep–wake control. J. Neurosci. 2002, 22, 7695–7711. [Google Scholar] [PubMed]
- Anaclet, C.; Parmentier, R.; Ouk, K.; Guidon, G.; Buda, C.; Sastre, J.P.; Akaoka, H.; Sergeeva, O.A.; Yanagisawa, M.; Ohtsu, H.; et al. Orexin/hypocretin and histamine: Distinct roles in the control of wakefulness demonstrated using knock-out mouse models. J. Neurosci. 2009, 29, 14423–14438. [Google Scholar] [CrossRef] [PubMed]
- Nishino, S.; Ripley, B.; Overeem, S.; Lammers, G.J.; Mignot, E. Hypocretin (orexin) deficiency in human narcolepsy. Lancet 2000, 355, 39–40. [Google Scholar] [CrossRef]
- Lin, L.; Faraco, J.; Li, R.; Kadotani, H.; Rogers, W.; Lin, X.Y.; Qiu, X.H.; de Jong, P.J.; Nishino, S.; Mignot, E. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell 1999, 98, 365–376. [Google Scholar] [CrossRef]
- Chemelli, R.M.; Willie, J.T.; Sinton, C.M.; Elmquist, J.K.; Scammell, T.; Lee, C.; Richardson, J.A.; Williams, S.C.; Xiong, Y.; Kisanuki, Y.; et al. Narcolepsy in orexin knockout mice: Molecular genetics of sleep regulation. Cell 1999, 98, 437–451. [Google Scholar] [CrossRef]
- Hara, J.; Beuckmann, C.T.; Nambu, T.; Willie, J.T.; Chemelli, R.M.; Sinton, C.M.; Sugiyama, F.; Yagami, K.; Goto, K.; Yanagisawa, M.; et al. Genetic ablation of orexin neurons in mice results in narcolepsy, hypophagia, and obesity. Neuron 2001, 30, 345–354. [Google Scholar] [CrossRef]
- Piper, D.C.; Upton, N.; Smith, M.I.; Hunter, A.J. The novel brain neuropeptide, Orexin-A, modulates the sleep-wake cycle of rats. Eur. J. Neurosci. 2000, 12, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Brisbare-Roch, C.; Dingemanse, J.; Koberstein, R.; Hoever, P.; Aissaoui, H.; Flores, S.; Mueller, C.; Nayler, O.; van Gerven, J.; de Haas, S.L.; et al. Promotion of sleep by targeting the orexin system in rats, dogs and humans. Nat. Med. 2007, 13, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Suzuki, M.; Mieda, M.; Tsujino, N.; Roth, B.; Sakurai, T. Pharmacogenetic modulation of orexin neurons alters sleep/wakefulness states in mice. PLoS ONE 2011, 6, e20360. [Google Scholar] [CrossRef] [PubMed]
- Munn, R.G.K.; Tyree, S.M.; McNaughton, N.; Bilkey, D.K. The frequency of hippocampal θ rhythm is modulated on a circadian period and is entrained by food availability. Front. Behav. Neurosci. 2015, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Yuasa, T.; Hayasaka, N.; Horikawa, K.; Sakurai, T.; Shibata, S. Reduced food anticipatory activity in genetically orexin (hypocretin) neuron-ablated mice. Eur. J. Neurosci. 2004, 20, 3054–3062. [Google Scholar] [CrossRef] [PubMed]
- Diano, S.; Horvath, B.; Urbanski, H.F.; Sotonyi, P.; Horvath, T.L. Fasting activates the nonhuman primate hypocretin (orexin) system and its postsynaptic targets. Endocrinology 2003, 144, 3774–3778. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Beuckmann, C.T.; Willie, J.T.; Hara, J.; Tsujino, N.; Mieda, M.; Tominaga, M.; Yagami, K.; Sugiyama, F.; Goto, K.; et al. Hypothalamic orexin neurons regulate arousal according to energy balance in mice. Neuron 2003, 38, 701–713. [Google Scholar] [CrossRef]
- Mieda, M.; Williams, S.C.; Sinton, C.M.; Richardson, J.A.; Sakurai, T.; Yanagisawa, M. Orexin neurons function in an efferent pathway of a food-entrainable circadian oscillator in eliciting food-anticipatory activity and wakefulness. J. Neurosci. 2004, 24, 10493–10501. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Lu, M.; Ge, F.; Marsh, D.J.; Qian, S.; Wang, A.H.; Picciotto, M.R.; Gao, X.B. Regulation of synaptic efficacy in hypocretin/orexin-containing neurons by melanin concentrating hormone in the lateral hypothalamus. J. Neurosci. 2008, 28, 9101–9110. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.Y.; Acuna-Goycolea, C.; van den Pol, A.N. Neuropeptide y inhibits hypocretin/orexin neurons by multiple presynaptic and postsynaptic mechanisms: Tonic depression of the hypothalamic arousal system. J. Neurosci. 2004, 24, 8741–8751. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Goycolea, C.; van den Pol, A. Glucagon-like peptide 1 excites hypocretin/orexin neurons by direct and indirect mechanisms: Implications for viscera-mediated arousal. J. Neurosci. 2004, 24, 8141–8152. [Google Scholar] [CrossRef] [PubMed]
- Burdakov, D.; Jensen, L.T.; Alexopoulos, H.; Williams, R.H.; Fearon, I.M.; O’Kelly, I.; Gerasimenko, O.; Fugger, L.; Verkhratsky, A. Tandem-pore K+ channels mediate inhibition of orexin neurons by glucose. Neuron 2006, 50, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Ahnaou, A.; Drinkenburg, W.H.I.M.; Bouwknecht, J.A.; Alcazar, J.; Steckler, T.; Dautzenberg, F.M. Blocking melanin-concentrating hormone mch 1 receptor affects rat sleep–wake architecture. Eur. J. Pharmacol. 2008, 579, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Adamantidis, A.R.; Salvert, D.; Goutagny, R.; Lakaye, B.; Gervasoni, D.; Grisar, T.; Luppi, P.H.; Fort, P. Sleep architecture of the melanin–concentrating hormone receptor 1-knockout mice. Eur. J. Neurosci. 2008, 27, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Willie, J.T.; Sinton, C.M.; Maratos-Flier, E.; Yanagisawa, M. Abnormal response of melanin-concentrating hormone deficient mice to fasting: Hyperactivity and rapid eye movement sleep suppression. Neuroscience 2008, 156, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Burdakov, D.; Gerasimenko, O.; Verkhratsky, A. Physiological changes in glucose differentially modulate the excitability of hypothalamic melanin-concentrating hormone and orexin neurons in situ. J. Neurosci. 2005, 25, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Hur, E.E.; Zaborszky, L. Vglut2 afferents to the medial prefrontal and primary somatosensory cortices: A combined retrograde tracing in situ hybridization study [corrected]. J. Comp. Neurol. 2005, 483, 351–373. [Google Scholar] [CrossRef] [PubMed]
- Henny, P.; Jones, B.E. Projections from basal forebrain to prefrontal cortex comprise cholinergic, GABAergic and glutamatergic inputs to pyramidal cells or interneurons. Eur. J. Neurosci. 2008, 27, 654–670. [Google Scholar] [CrossRef] [PubMed]
- Gritti, I.; Mainville, L.; Mancia, M.; Jones, B.E. GABAergic and other noncholinergic basal forebrain neurons, together with cholinergic neurons, project to the mesocortex and isocortex in the rat. J. Comp. Neurol. 1997, 383, 163–177. [Google Scholar] [CrossRef]
- Freund, T.F.; Meskenaite, V. Γ-aminobutyric acid-containing basal forebrain neurons innervate inhibitory interneurons in the neocortex. Proc. Natl. Acad. Sci. USA 1992, 89, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Bäckberg, M.; Ultenius, C.; Fritschy, J.M.; Meister, B. Cellular localization of GABA receptor α subunit immunoreactivity in the rat hypothalamus: Relationship with neurones containing orexigenic or anorexigenic peptides. J. Neuroendocrinol. 2004, 16, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.R.; Hokfelt, T.; Skirboll, L.R.; Wu, J.Y. Hypothalamic γ-aminobutyric acid neurons project to the neocortex. Science 1983, 220, 1309–1311. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, S.C.; Svingos, A.L.; Pickel, V.M.; Henriksen, S.J. Electrophysiological characterization of GABAergic neurons in the ventral tegmental area. J. Neurosci. 1998, 18, 8003–8015. [Google Scholar] [PubMed]
- Rosin, D.L.; Weston, M.C.; Sevigny, C.P.; Stornetta, R.L.; Guyenet, P.G. Hypothalamic orexin (hypocretin) neurons express vesicular glutamate transporters vglut1 or vglut2. J. Comp. Neurol. 2003, 465, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; Gall, C.M.; Jackson, V.R.; Civelli, O.; Reinscheid, R.K. Distribution of neuropeptide S receptor mRNA and neurochemical characteristics of neuropeptide S-expressing neurons in the rat brain. J. Comp. Neurol. 2007, 500, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Gras, C.; Herzog, E.; Bellenchi, G.C.; Bernard, V.; Ravassard, P.; Pohl, M.; Gasnier, B.; Giros, B.; El Mestikawy, S. A third vesicular glutamate transporter expressed by cholinergic and serotoninergic neurons. J. Neurosci. 2002, 22, 5442–5451. [Google Scholar] [PubMed]
- Kaur, S.; Pedersen, N.P.; Yokota, S.; Hur, E.E.; Fuller, P.M.; Lazarus, M.; Chamberlin, N.L.; Saper, C.B. Glutamatergic signaling from the parabrachial nucleus plays a critical role in hypercapnic arousal. J. Neurosci. 2013, 33, 7627–7640. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Kumar, S.; Suntsova, N.; Bashir, T.; Szymusiak, R.; McGinty, D. GABAergic regulation of the perifornical-lateral hypothalamic neurons during non-rapid eye movement sleep in rats. Neuroscience 2010, 167, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.S.; Steffensen, S.C.; Henriksen, S.J. Discharge profiles of ventral tegmental area GABA neurons during movement, anesthesia, and the sleep–wake cycle. J. Neurosci. 2001, 21, 1757–1766. [Google Scholar] [PubMed]
- Lu, J.; Greco, M.A.; Shiromani, P.; Saper, C.B. Effect of lesions of the ventrolateral preoptic nucleus on NREM and rem sleep. J. Neurosci. 2000, 20, 3830–3842. [Google Scholar] [PubMed]
- Szymusiak, R.; McGinty, D. Hypothalamic regulation of sleep and arousal. Ann. N. Y. Acad. Sci. 2008, 1129, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Adamantidis, A.R.; Zhang, F.; Aravanis, A.M.; Deisseroth, K.; de Lecea, L. Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature 2007, 450, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Adamantidis, A.; Ohtsu, H.; Deisseroth, K.; de Lecea, L. Sleep homeostasis modulates hypocretin-mediated sleep-wake transitions. J. Neurosci. 2009, 29, 10939–10949. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, T.; Kilduff, T.S.; Boyden, E.S.; Takahashi, S.; Tominaga, M.; Yamanaka, A. Acute optogenetic silencing of orexin/hypocretin neurons induces slow–wave sleep in mice. J. Neurosci. 2011, 31, 10529–10539. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, T.; Tabuchi, S.; Tanaka, K.F.; Boyden, E.S.; Tominaga, M.; Yamanaka, A. Long-lasting silencing of orexin/hypocretin neurons using archaerhodopsin induces slow-wave sleep in mice. Behav. Brain Res. 2013, 255, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Konadhode, R.R.; Pelluru, D.; Blanco-Centurion, C.; Zayachkivsky, A.; Liu, M.; Uhde, T.; Glen, W.B., Jr.; van den Pol, A.N.; Mulholland, P.J.; Shiromani, P.J. Optogenetic stimulation of mch neurons increases sleep. J. Neurosci. 2013, 33, 10257–10263. [Google Scholar] [CrossRef] [PubMed]
- Jego, S.; Glasgow, S.D.; Herrera, C.G.; Ekstrand, M.; Reed, S.J.; Boyce, R.; Friedman, J.; Burdakov, D.; Adamantidis, A.R. Optogenetic identification of a rapid eye movement sleep modulatory circuit in the hypothalamus. Nat. Neurosci. 2013, 16, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Tsunematsu, T.; Ueno, T.; Tabuchi, S.; Inutsuka, A.; Tanaka, K.F.; Hasuwa, H.; Kilduff, T.S.; Terao, A.; Yamanaka, A. Optogenetic manipulation of activity and temporally controlled cell-specific ablation reveal a role for mch neurons in sleep/wake regulation. J. Neurosci. 2014, 34, 6896–6909. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.R.; Treweek, J.B.; Robinson, J.E.; Xiao, C.; Bremner, L.R.; Greenbaum, A.; Gradinaru, V. Dorsal raphe dopamine neurons modulate arousal and promote wakefulness by salient stimuli. Neuron 2017, 94, 1205–1219. [Google Scholar] [CrossRef] [PubMed]
- Van Dort, C.J.; Zachs, D.P.; Kenny, J.D.; Zheng, S.; Goldblum, R.R.; Gelwan, N.A.; Ramos, D.M.; Nolan, M.A.; Wang, K.; Weng, F.J. Optogenetic activation of cholinergic neurons in the PPT or LDT induces REM sleep. Proc. Natl. Acad. Sci. USA 2015, 112, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.; Soya, S.; Sakurai, T. Excitation of GABAergic neurons in the bed nucleus of the stria terminalis triggers immediate transition from non-rapid eye movement sleep to wakefulness in mice. J. Neurosci. 2017, 37, 7164–7176. [Google Scholar] [CrossRef] [PubMed]
- Venner, A.; Anaclet, C.; Broadhurst, R.Y.; Saper, C.B.; Fuller, P.M. A novel population of wake-promoting GABAergic neurons in the ventral lateral hypothalamus. Curr. Biol. 2016, 26, 2137–2143. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Weber, F.; Zhong, P.; Tan, C.L.; Nguyen, T.N.; Beier, K.T.; Hörmann, N.; Chang, W.C.; Zhang, Z.; Do, J.P. Identification of preoptic sleep neurons using retrograde labelling and gene profiling. Nature 2017, 545, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Pelluru, D.; Konadhode, R.R.; Bhat, N.R.; Shiromani, P.J. Optogenetic stimulation of astrocytes in the posterior hypothalamus increases sleep at night in C57BL/6J mice. Eur. J. Neurosci. 2016, 43, 1298–1306. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, K.S.; Sergeeva, O.; Brown, R.E.; Haas, H.L. Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus. J Neurosci 2001, 21, 9273–9279. [Google Scholar] [PubMed]
- Nishino, S.; Sakurai, E.; Nevsimalova, S.; Yoshida, Y.; Watanabe, T.; Yanai, K.; Mignot, E. Decreased csf histamine in narcolepsy with and without low csf hypocretin-1 in comparison to healthy controls. Sleep 2009, 32, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Kanbayashi, T.; Kodama, T.; Kondo, H.; Satoh, S.; Inoue, Y.; Chiba, S.; Shimizu, T.; Nishino, S. Csf histamine contents in narcolepsy, idiopathic hypersomnia and obstructive sleep apnea syndrome. Sleep 2009, 32, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Nishino, S.; Fujiki, N.; Ripley, B.; Sakurai, E.; Kato, M.; Watanabe, T.; Mignot, E.; Yanai, K. Decreased brain histamine content in hypocretin/orexin receptor-2 mutated narcoleptic dogs. Neurosci. Lett. 2001, 313, 125–128. [Google Scholar] [CrossRef]
- Shigemoto, Y.; Fujii, Y.; Shinomiya, K.; Kamei, C. Participation of histaminergic H1 and noradrenergic α 1 receptors in orexin A-induced wakefulness in rats. Brain Res. 2004, 1023, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Tsujino, N.; Funahashi, H.; Honda, K.; Guan, J.L.; Wang, Q.P.; Tominaga, M.; Goto, K.; Shioda, S.; Sakurai, T. Orexins activate histaminergic neurons via the orexin 2 receptor. Biochem. Biophys. Res. Commun. 2002, 290, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.L.; Qu, W.M.; Li, W.D.; Mochizuki, T.; Eguchi, N.; Watanabe, T.; Urade, Y.; Hayaishi, O. Arousal effect of orexin a depends on activation of the histaminergic system. Proc. Natl. Acad. Sci. USA 2001, 98, 9965–9970. [Google Scholar] [CrossRef] [PubMed]
- Schone, C.; Cao, Z.F.; Apergis-Schoute, J.; Adamantidis, A.; Sakurai, T.; Burdakov, D. Optogenetic probing of fast glutamatergic transmission from hypocretin/orexin to histamine neurons in situ. J. Neurosci. 2012, 32, 12437–12443. [Google Scholar] [CrossRef] [PubMed]
- Schone, C.; Apergis-Schoute, J.; Sakurai, T.; Adamantidis, A.; Burdakov, D. Coreleased orexin and glutamate evoke nonredundant spike outputs and computations in histamine neurons. Cell Rep. 2014, 7, 697–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.H.; Chee, M.J.; Kroeger, D.; Ferrari, L.L.; Maratos-Flier, E.; Scammell, T.E.; Arrigoni, E. Optogenetic-mediated release of histamine reveals distal and autoregulatory mechanisms for controlling arousal. J. Neurosci. 2014, 34, 6023–6029. [Google Scholar] [CrossRef] [PubMed]
- Peyron, C.; Tighe, D.K.; van den Pol, A.N.; de Lecea, L.; Heller, H.C.; Sutcliffe, J.G.; Kilduff, T.S. Neurons containing hypocretin (orexin) project to multiple neuronal systems. J. Neurosci. 1998, 18, 9996–10015. [Google Scholar] [PubMed]
- Hagan, J.J.; Leslie, R.A.; Patel, S.; Evans, M.L.; Wattam, T.A.; Holmes, S.; Benham, C.D.; Taylor, S.G.; Routledge, C.; Hemmati, P.; et al. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc. Natl. Acad. Sci. USA 1999, 96, 10911–10916. [Google Scholar] [CrossRef] [PubMed]
- Bourgin, P.; Huitron–Resendiz, S.; Spier, A.D.; Fabre, V.; Morte, B.; Criado, J.R.; Sutcliffe, J.G.; Henriksen, S.J.; de Lecea, L. Hypocretin-1 modulates rapid eye movement sleep through activation of locus coeruleus neurons. J. Neurosci. 2000, 20, 7760–7765. [Google Scholar] [PubMed]
- Hasegawa, E.; Yanagisawa, M.; Sakurai, T.; Mieda, M. Orexin neurons suppress narcolepsy via 2 distinct efferent pathways. J. Clin. Investig. 2014, 124, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; de Lecea, L. Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat. Neurosci. 2010, 13, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Brill, J.; Bonnavion, P.; Huguenard, J.R.; Huerta, R.; de Lecea, L. Mechanism for hypocretin–mediated sleep-wake transitions. Proc. Natl. Acad. Sci. USA 2012, 109, E2635–E2644. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; de Lecea, L.; Adamantidis, A. Functional wiring of hypocretin and LC-NE neurons: Implications for arousal. Front. Behav. Neurosci. 2013, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnavion, P.; Jackson, A.C.; Carter, M.E.; de Lecea, L. Antagonistic interplay between hypocretin and leptin in the lateral hypothalamus regulates stress responses. Nat. Commun. 2015, 6, 6266. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Garza, J.C.; Bronner, J.; Kim, C.S.; Zhang, W.; Lu, X.-Y. Acute administration of leptin produces anxiolytic-like effects: A comparison with fluoxetine. Psychopharmacology 2010, 207, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Tyree, S.M.; Munn, R.G.K.; McNaughton, N. Anxiolytic-like effects of leptin on fixed interval responding. Pharmacol. Biochem. Behav. 2016, 148, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Louis, G.W.; Leinninger, G.M.; Rhodes, C.J.; Myers, M.G. Direct innervation and modulation of orexin neurons by lateral hypothalamic LepRb neurons. J. Neurosci. 2010, 30, 11278–11287. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Tritos, N.A.; Lowell, B.B.; Flier, J.S.; Maratos–Flier, E. Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature 1998, 396, 670–674. [Google Scholar] [PubMed]
- Marsh, D.J.; Weingarth, D.T.; Novi, D.E.; Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Guan, X.M.; Jiang, M.M.; Feng, Y.; Camacho, R.E. Melanin-concentrating hormone 1 receptor-deficient mice are lean, hyperactive, and hyperphagic and have altered metabolism. Proc. Natl. Acad. Sci. USA 2002, 99, 3240–3245. [Google Scholar] [CrossRef] [PubMed]
- Takase, K.; Kikuchi, K.; Tsuneoka, Y.; Oda, S.; Kuroda, M.; Funato, H. Meta-analysis of melanin–concentrating hormone signaling-deficient mice on behavioral and metabolic phenotypes. PLoS ONE 2014, 9, e99961. [Google Scholar] [CrossRef] [PubMed]
- Apergis-Schoute, J.; Iordanidou, P.; Faure, C.; Jego, S.; Schone, C.; Aitta-Aho, T.; Adamantidis, A.; Burdakov, D. Optogenetic evidence for inhibitory signaling from orexin to mch neurons via local microcircuits. J. Neurosci. 2015, 35, 5435–5441. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhou, J.; Feng, Q.; Lin, R.; Gong, H.; Luo, Q.; Zeng, S.; Luo, M.; Fu, L. Multi–channel fiber photometry for population neuronal activity recording. Biomed. Opt. Express 2015, 6, 3919–3931. [Google Scholar] [CrossRef] [PubMed]
- Adelsberger, H.; Grienberger, C.; Stroh, A.; Konnerth, A. In vivo calcium recordings and channelrhodopsin-2 activation through an optical fiber. Cold Spring Harb. Protoc. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.N.; Macosko, E.Z.; Fenselau, H.; Pers, T.H.; Lyubetskaya, A.; Tenen, D.; Goldman, M.; Verstegen, A.M.J.; Resch, J.M.; McCarroll, S.A. A molecular census of arcuate hypothalamus and median eminence cell types. Nat. Neurosci. 2017, 20, 484–496. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A visual representation of neural activity profiles of sleep/wake-active neurons populations. Showing high (solid line), moderate (thick broken line), low (thin broken line), and quiet (no line) activity profiles during active waking, quiet waking, non-rapid eye movement sleep (NREM), and rapid eye movement sleep (REM) states in basal forebrain [25,26,27,28,29,30] and brainstem [31,32,33,34,35] cholinergic, dorsal raphe nuclei serotonergic [36,37,38], ventral tegmental area (VTA) dopaminergic [39,40], locus coeruleus noradrenergic [41,42,43,44], tuberomammillary nucleus histaminergic [45,46,47], lateral hypothalamic hypocretin [48,49,50,51,52,53,54,55,56], lateral hypothalamic area melanin-concentrating hormone [57,58,59,60], VTA and basal forebrain γ amino butyric acidergic (GABAergic) [26,61], and ventral medulla GABAergic neurons [62]. 1 s = 1 second.
Figure 1.
A visual representation of neural activity profiles of sleep/wake-active neurons populations. Showing high (solid line), moderate (thick broken line), low (thin broken line), and quiet (no line) activity profiles during active waking, quiet waking, non-rapid eye movement sleep (NREM), and rapid eye movement sleep (REM) states in basal forebrain [25,26,27,28,29,30] and brainstem [31,32,33,34,35] cholinergic, dorsal raphe nuclei serotonergic [36,37,38], ventral tegmental area (VTA) dopaminergic [39,40], locus coeruleus noradrenergic [41,42,43,44], tuberomammillary nucleus histaminergic [45,46,47], lateral hypothalamic hypocretin [48,49,50,51,52,53,54,55,56], lateral hypothalamic area melanin-concentrating hormone [57,58,59,60], VTA and basal forebrain γ amino butyric acidergic (GABAergic) [26,61], and ventral medulla GABAergic neurons [62]. 1 s = 1 second.
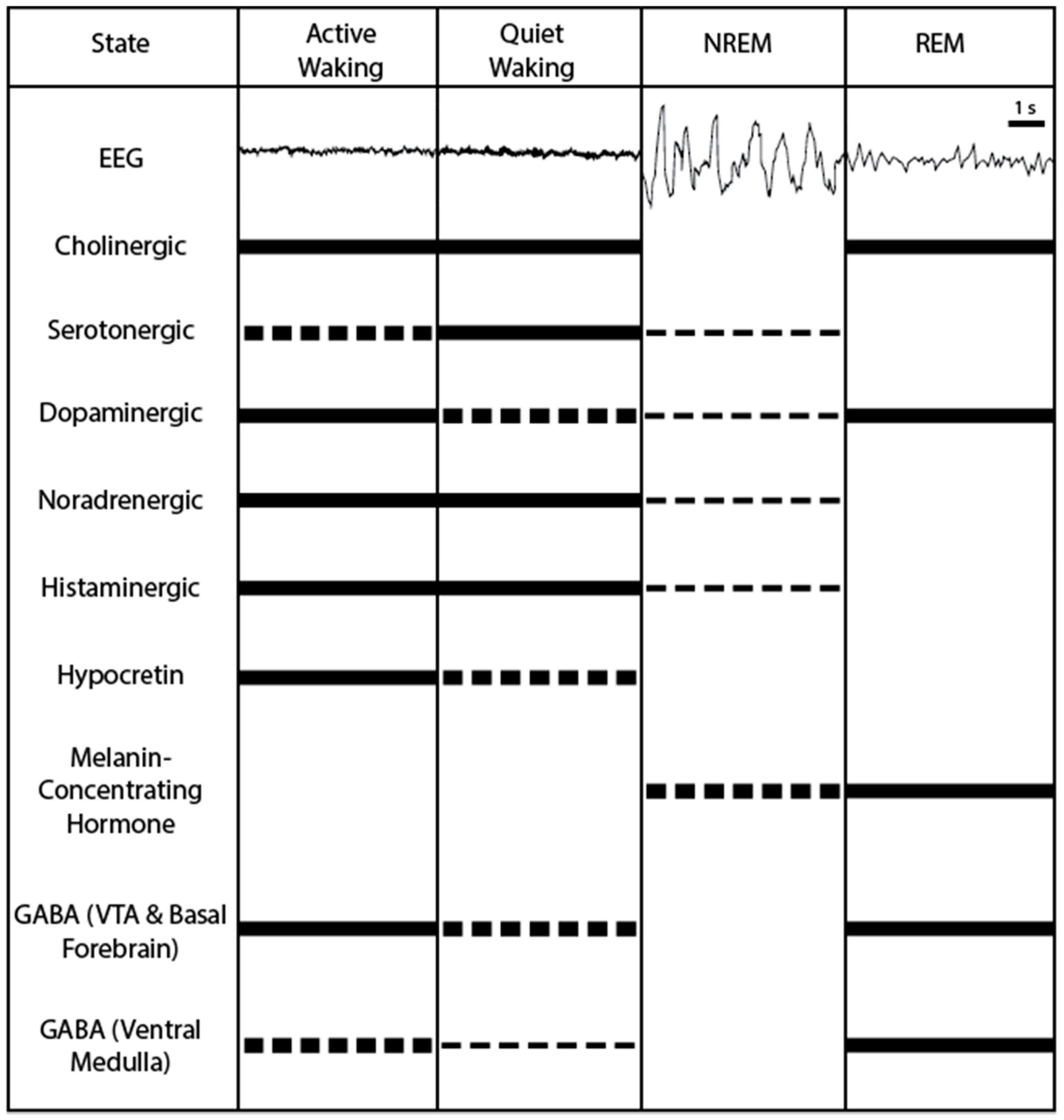
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tyree, S.M.; De Lecea, L. Optogenetic Investigation of Arousal Circuits. Int. J. Mol. Sci. 2017, 18, 1773. https://doi.org/10.3390/ijms18081773
AMA Style
Tyree SM, De Lecea L. Optogenetic Investigation of Arousal Circuits. International Journal of Molecular Sciences. 2017; 18(8):1773. https://doi.org/10.3390/ijms18081773
Chicago/Turabian StyleTyree, Susan M., and Luis De Lecea. 2017. "Optogenetic Investigation of Arousal Circuits" International Journal of Molecular Sciences 18, no. 8: 1773. https://doi.org/10.3390/ijms18081773
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.