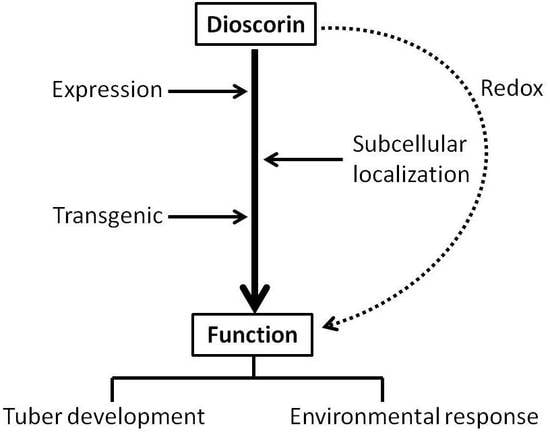
Characterization of the Dioscorin Gene Family in Dioscorea alata Reveals a Role in Tuber Development and Environmental Response

Abstract
:
1. Introduction
2. Results
2.1. Identification of the Da-dio Gene Family
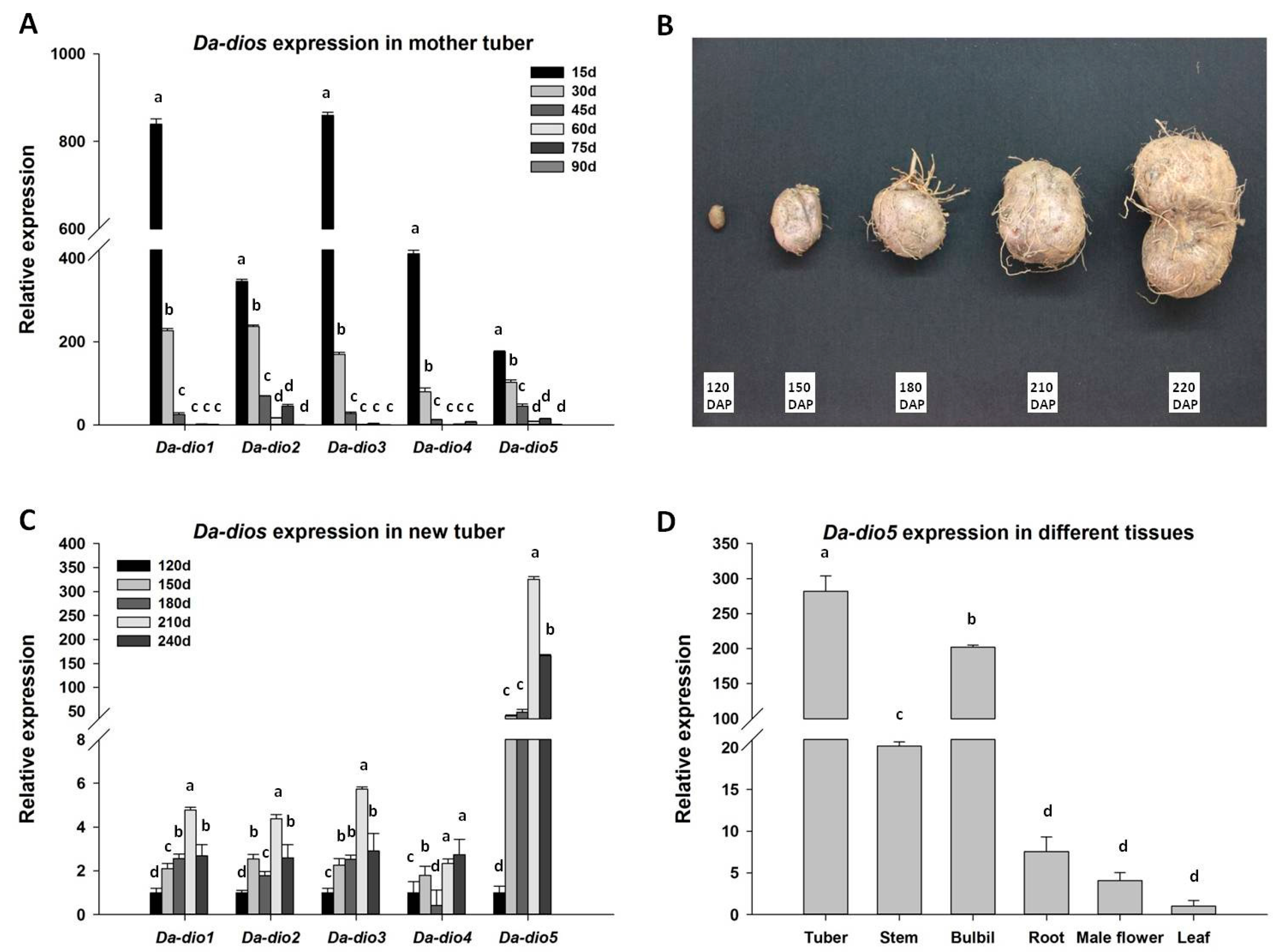
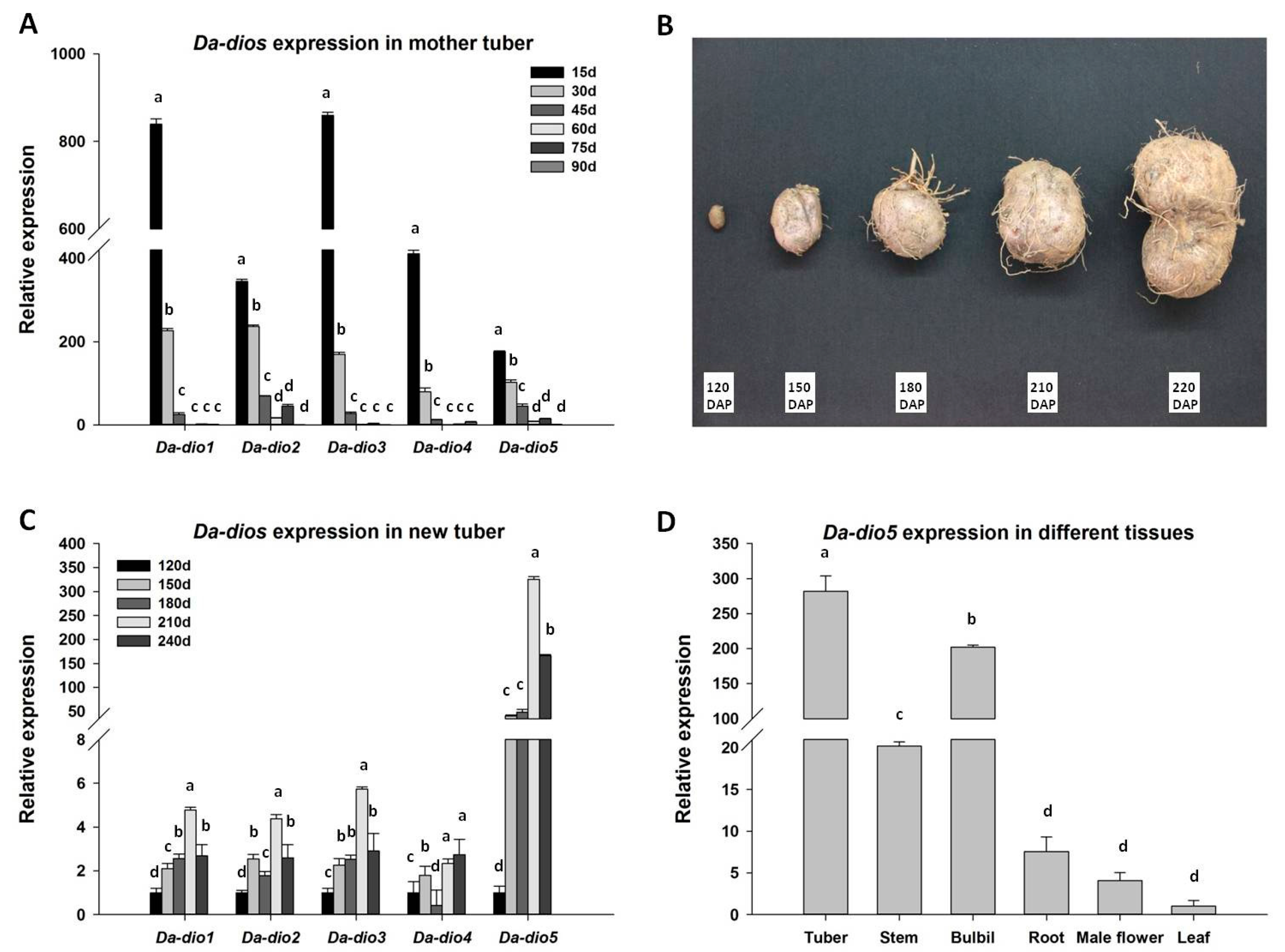
2.2. Expression of the Five Da-dios During Tuber Development
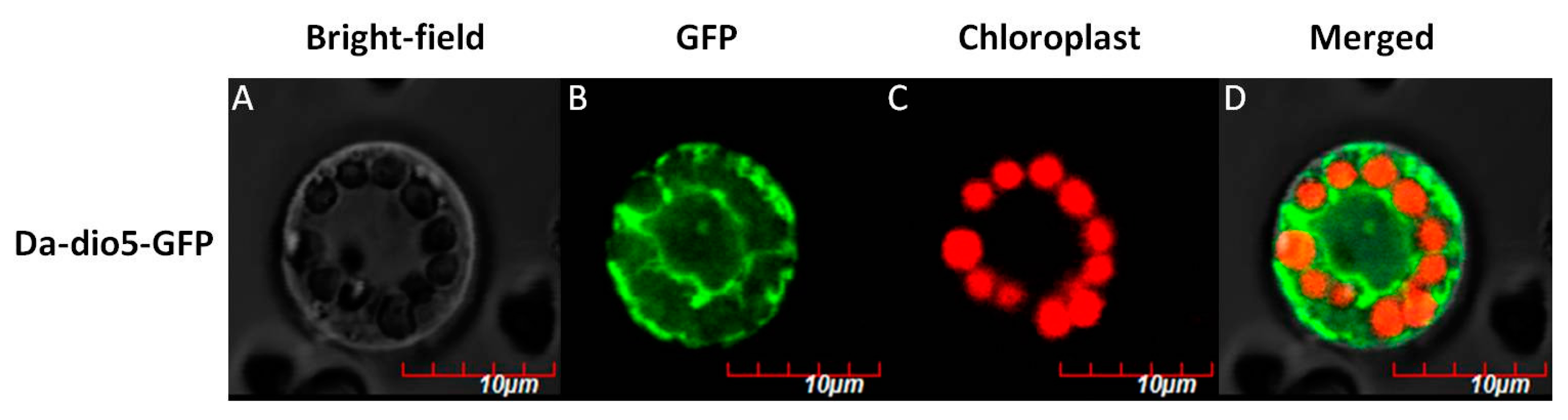
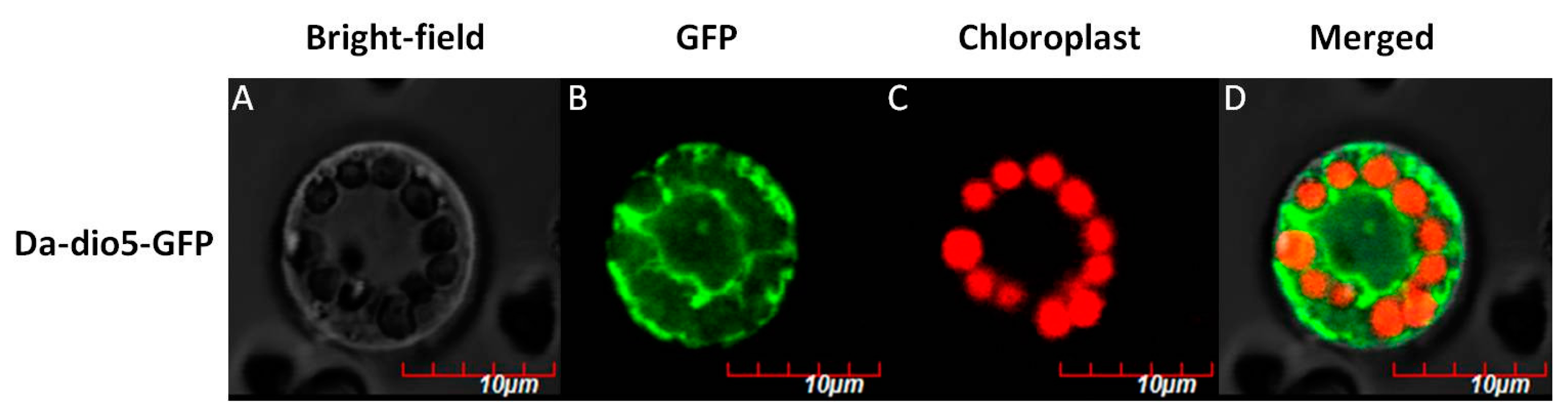
2.3. Expression Pattern and Localization of Da-dio5
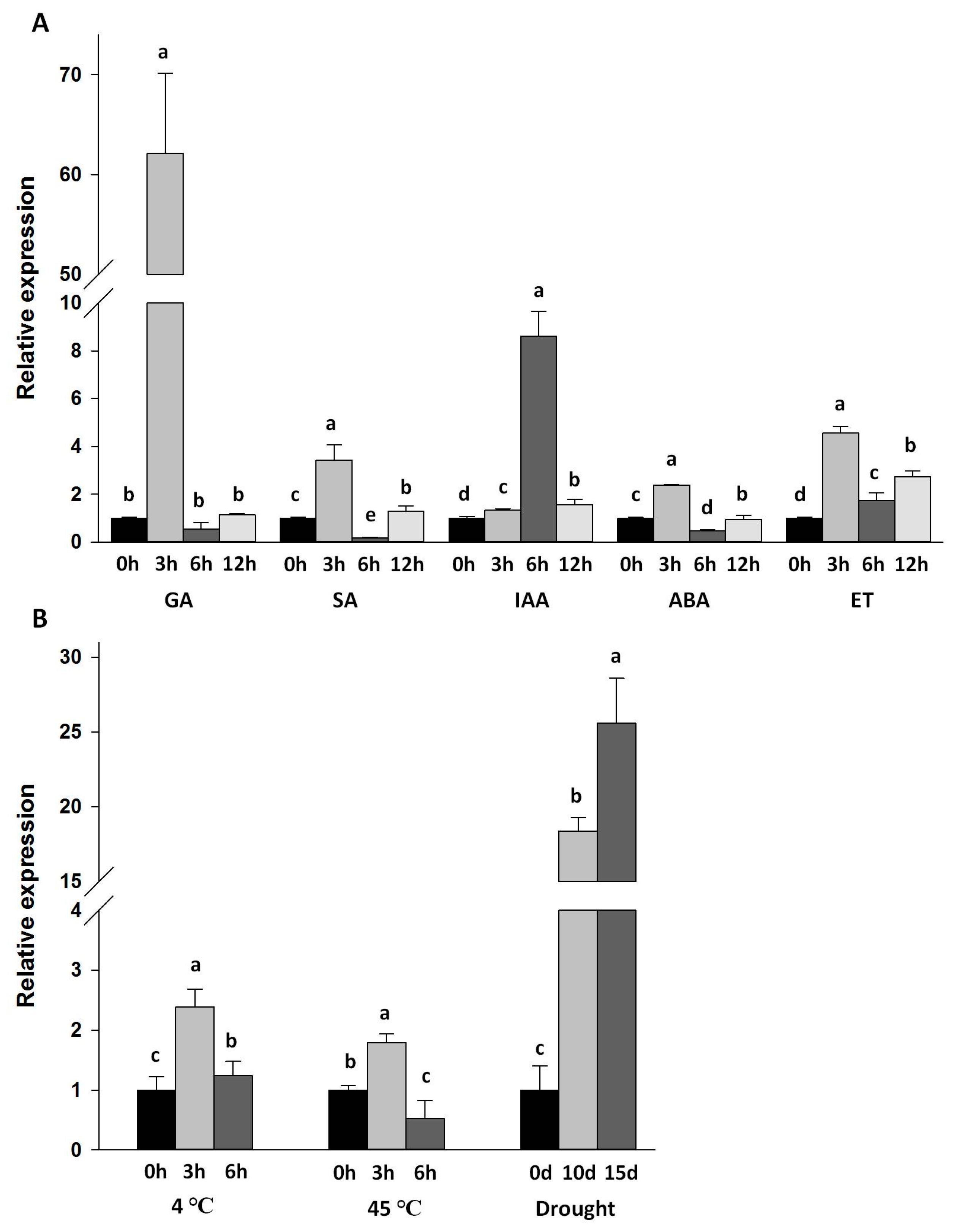
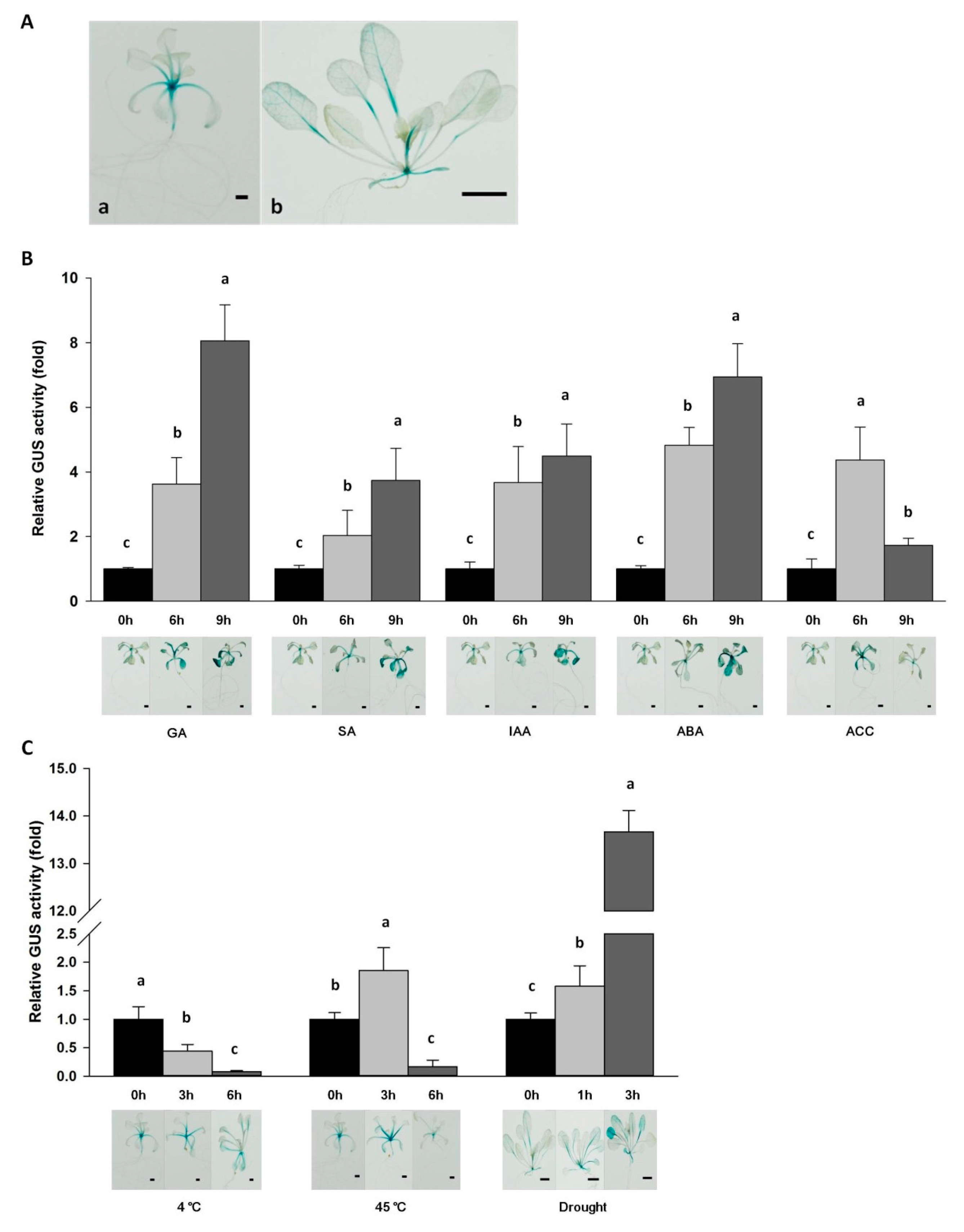
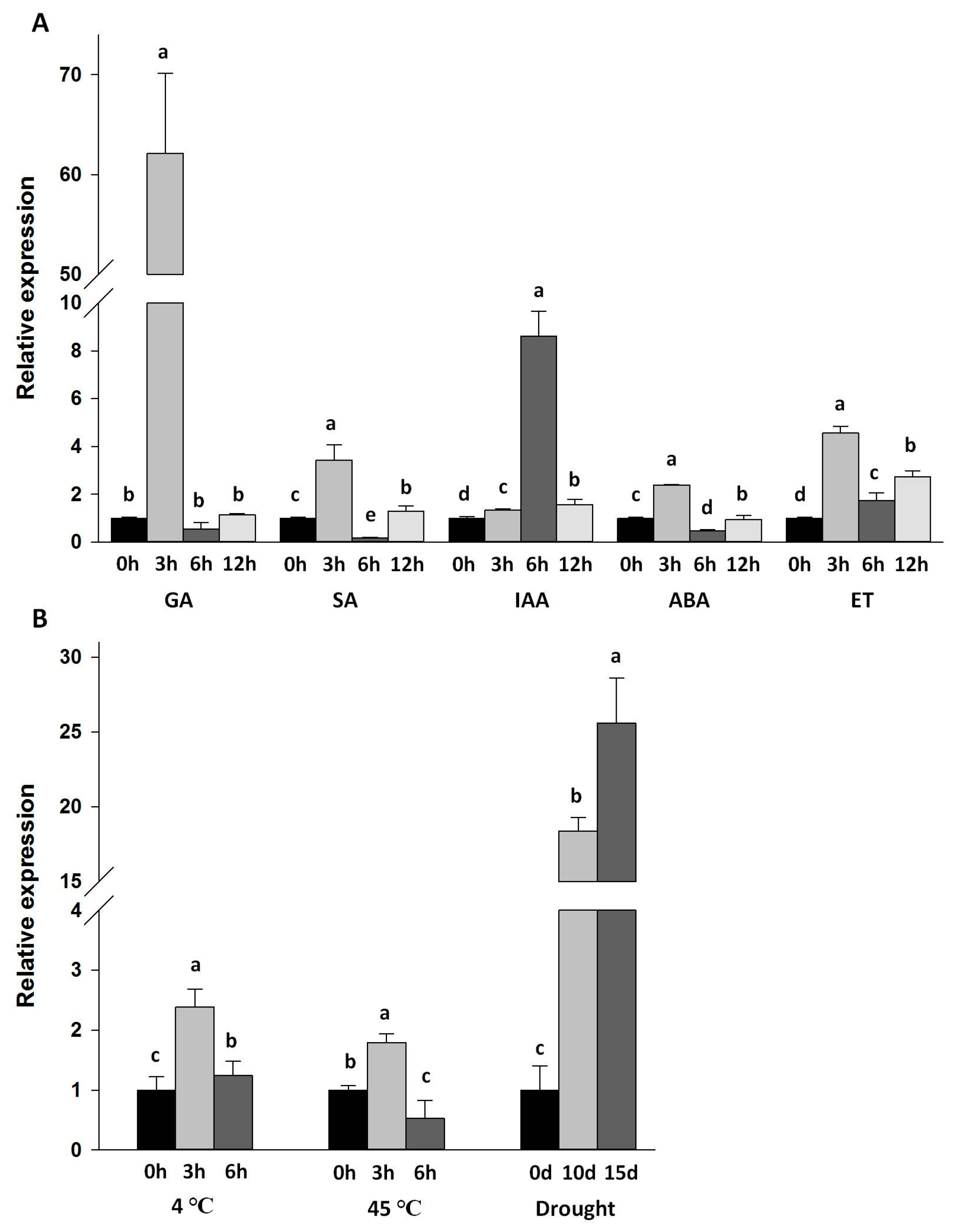
2.4. Expression of Da-dio5 in Response to Hormones and Abiotic Stresses
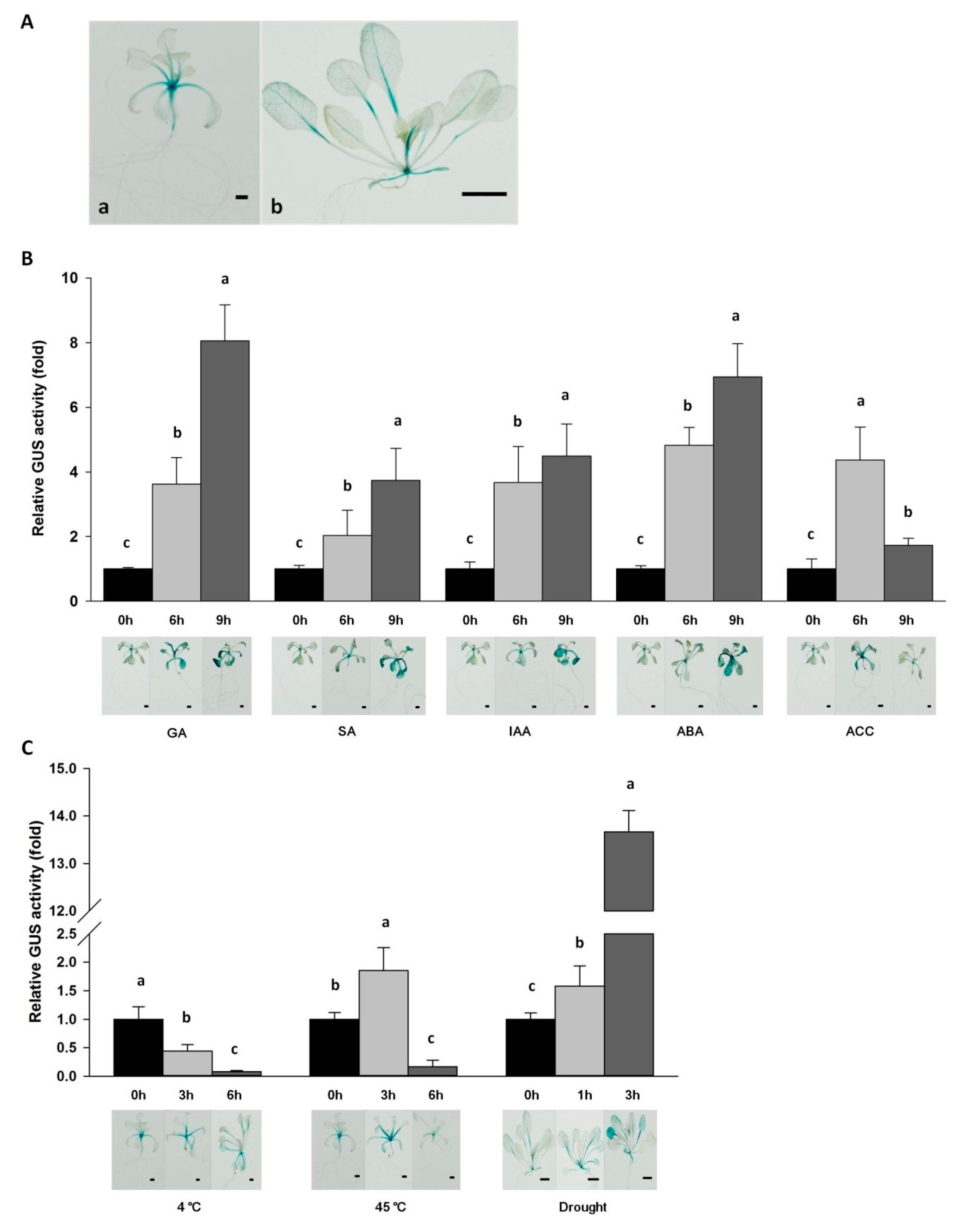
2.5. Functional Characterization of the Da-dio5 Promoter
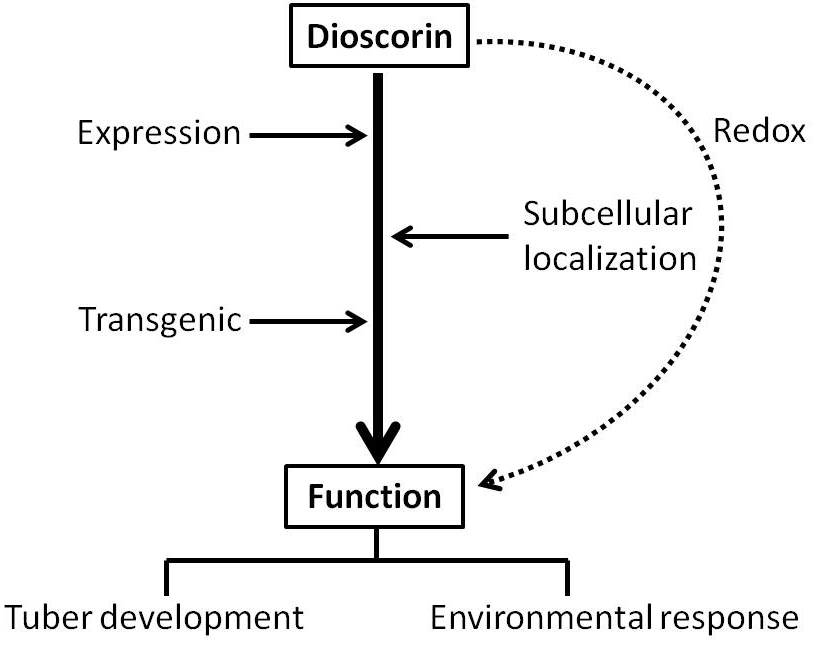
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Hormone and Abiotic Stress Treatments
4.3. Isolation of Dioscorin Genes
4.4. Multiple Sequence Alignments and Bioinformatic Analysis
4.5. RNA Isolation and cDNA Synthesis
4.6. qRT-PCR
4.7. Subcelluar Localization of Da-dio5
4.8. Promoter Cloning and Analysis
4.9. Construction of the Da-dio5 Promoter-GUS Fusion and A. thaliana Transformation
4.10. Histochemical Localization and Quantitative Analysis of GUS Activity
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ORF | Open reading frame |
| aa | Amino acids |
| MW | Molecular weights |
| bp | Base pair |
| BLAST | Basic local alignment search tool |
| TI | Trypsin inhibitor |
| CA | Carbonic anhydrase |
| DHA | Dehydroascorbate |
| MDA | Monodehydroascorbate |
| pI | Isoelectric point |
| qRT-PCR | Quantitative real-time RT-PCR |
| DAP | Days after planting |
| SAM | Shoot apical meristem |
| DAG | Days after germination |
| WT | Wild-type |
| ET | Ethylene |
| MeJA | Methyl jasmonate |
| ABA | Abscisic acid |
| IAA | Indole-3-acetic acid |
| SA | Salicylic acid |
| GA | Gibberellic acid |
| ACC | 1-Aminocyclopropanecarboxylic acid |
| GUS | β-Glucuronidase |
| GFP | Green fluorescent protein |
References
- Shewry, P.R. Tuber storage proteins. Ann. Bot. 2003, 91, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Wanasundera, J.P.; Ravindran, G. Nutritional assessment of yam (Dioscorea alata) tubers. Plant Foods Hum. Nutr. 1994, 46, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.R.; Kasai, T.; Kawabata, J. Nutritional evaluation of wild yam (Dioscorea spp.) tubers of Nepal. Food Chem. 2003, 82, 619–623. [Google Scholar] [CrossRef]
- Bhandari, M.R.; Kawabata, J. Organic acid, phenolic content and antioxidant activity of wild yam (Dioscorea spp.) tubers of Nepal. Food Chem. 2004, 88, 163–168. [Google Scholar] [CrossRef]
- Nagai, T.; Nagashima, T. Functional properties of dioscorin, a soluble viscous protein from Japanese yam (Dioscorea opposita thunb.) tuber mucilage Tororo. Z. Naturforsch. C 2006, 61, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.L.; Hsu, Y.H.; Lee, P.Y.; Hou, W.C.; Hung, L.C.; Lin, C.H.; Chen, C.M.; Huang, Y.J. Dioscorin isolated from Dioscorea alata activates TLR4-signaling pathways and induces cytokine expression in macrophages. Biochem. Biophys. Res. Commun. 2006, 339, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.S.; Ko, Y.H.; Lin, K.W.; Hsu, J.Y.; Chu, J.J.; Chi, C.S. Dioscorin protects tight junction protein expression in A549 human airway epithelium cells from dust mite damage. J. Microbiol. Immunol. Infect. 2009, 42, 457–463. [Google Scholar] [PubMed]
- Araghiniknam, M.; Chung, S.; Nelson-White, T.; Eskelson, C.; Watson, R.R. Antioxidant activity of dioscorea and dehydroepiandrosterone (DHEA) in older humans. Life Sci. 1996, 59, PL147–PL157. [Google Scholar] [CrossRef]
- Akanbi, C.T.; Gureje, P.O.; Adeyemi, I.A. Effect of heat-moisture pre-treatment on physical characteristics of dehydrated yam. J. Food Eng. 1996, 28, 45–54. [Google Scholar] [CrossRef]
- Omonigho, S.E.; Ikenebomeh, M.J. Effect of temperature treatment on the chemical composition of pounded white yam during storage. Food Chem. 2000, 71, 215–220. [Google Scholar] [CrossRef]
- Harvey, P.J.; Boulter, B. Isolation and characterization of the storage protein of yam tubers (Dioscorea rotundata). Phytochemistry 1983, 22, 1687–1693. [Google Scholar] [CrossRef]
- Tsai, W.Y.; Jheng, Y.J.; Chen, K.H.; Lin, K.W.; Ho, Y.P.; Yang, C.C.; Lin, K.C. Molecular cloning, structural analysis and mass spectrometric identification of native dioscorins of various yam species. J. Sci. Food Agric. 2013, 93, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.C.; Chen, H.J.; Lin, Y.H. Dioscorins from different Dioscorea species all exhibit both carbonic anhydrase and trypsin inhibitor activities. Bot. Bull. Acad. Sin. 2000, 41, 191–196. [Google Scholar]
- Gaidamashvili, M.; Ohizumi, Y.; Iijima, S.; Takayama, T.; Ogawa, T.; Muramoto, K. Characterization of the yam tuber storage proteins from Dioscorea batatas exhibiting unique lectin activities. J. Biol. Chem. 2004, 279, 26028–26035. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.C.; Chen, H.J.; Lin, Y.H. Dioscorins, the major tuber storage proteins of yam (Dioscorea batatas Decne), with dehydroascorbate reductase and monodehydroascorbate reductase activities. Plant Sci. 1999, 149, 151–156. [Google Scholar] [CrossRef]
- Hou, W.C.; Liu, J.S.; Chen, H.J.; Chen, T.E.; Chang, C.F.; Lin, Y.H. Dioscorin, the major tuber storage protein of yam (Dioscorea batatas decne) with carbonic anhydrase and trypsin inhibitor activities. J. Agric. Food Chem. 1999, 47, 2168–2172. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.C.; Lee, M.H.; Chen, H.J.; Liang, W.L.; Han, C.H.; Liu, Y.W.; Lin, Y.H. Antioxidant activities of dioscorin, the storage protein of yam (Dioscorea batatas Decne) tuber. J. Agric. Food Chem. 2001, 49, 4956–4960. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.L.; Lin, Y.H.; Lee, M.H.; Lin, C.L.; Hou, W.C. Both dioscorin, the tuber storage protein of yam (Dioscorea alata cv. Tainong No. 1), and its peptic hydrolysates exhibited angiotensin converting enzyme inhibitory activities. J. Agric. Food Chem. 2002, 50, 6109–6113. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.W.; Chen, J.C.; Lin, M.I.; Chen, Y.M.; Lin, C.Y. Functional activity of sporamin from sweet potato (Ipomoea batatas Lam.): A tuber storage protein with trypsin inhibitory activity. Plant Mol. Biol. 1997, 33, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Racusen, D. Lipid acyl hydrolase of patatin. Can. J. Bot. 1984, 62, 1640–1644. [Google Scholar] [CrossRef]
- Hendriks, T.; Vreugdenhil, D.; Stiekema, W.J. Patatin and four serine proteinase inhibitor genes are differentially expressed during potato tuber development. Plant Mol. Biol. 1991, 17, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Hannapel, D.; Mignery, G.; Pikaard, C. Molecular approaches to the study of the major tuber proteins. Potato Physiol. 1985. [Google Scholar] [CrossRef]
- Maeshima, M.; Sasaki, T.; Asahi, T. Characterization of major proteins in sweet potato tuberous roots. Phytochemistry 1985, 24, 1899–1902. [Google Scholar] [CrossRef]
- Hattori, T.; Nakagawa, T.; Maeshima, M.; Nakamura, K.; Asahi, T. Molecular cloning and nucleotide sequence of cDNA for sporamin, the major soluble protein of sweet potato tuberous roots. Plant Mol. Biol. 1985, 5, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Hannapel, D.J. Characterization of the early events of potato tuber development. Physiol. Plant 1991, 83, 568–573. [Google Scholar] [CrossRef]
- Paiva, E.; Lister, R.M.; Park, W.D. Induction and accumulation of major tuber proteins of potato in stems and petioles. Plant Physiol. 1983, 71, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Gupta, R.; Deswal, R. Dioscorea alata tuber proteome analysis shows over thirty dioscorin isoforms and novel tuber proteins. Plant Physiol. Biochem. 2017, 114, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sehrawat, A.; Deswal, R. Asada-Halliwell pathway maintains redox status in Dioscorea alata tuber which helps in germination. Plant Sci. 2016, 250, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S.; Lin, I.W.; Chen, M.J.; Chen, C.Y.; Yeh, K.W. Differential activation of sporamin expression in response to abiotic mechanical wounding and biotic herbivore attack in the sweet potato. BMC Plant Biol. 2014, 14, 112. [Google Scholar] [CrossRef] [PubMed]
- Twell, D.; Ooms, G. The 5′ flanking DNA of a patatin gene directs tuber specific expression of a chimaeric gene in potato. Plant Mol. Biol. 1987, 9, 345–375. [Google Scholar] [CrossRef] [PubMed]
- Rosahl, S.; Schell, J.; Willmitzer, L. Expression of a tuber-specific storage protein in transgenic tobacco plants: Demonstration of an esterase activity. EMBO J. 1987, 6, 1155–1159. [Google Scholar] [PubMed]
- Zhou, M.S.; Wang, P.L.; Xiang, X.; Wei, H.B.; Li, Z.L.; Li, R.Y.; Fang, X.F.; Cao, S.J. Cloning and molecular characteristics of ANS gene and its correlations with anthocyan in accumulation in yam. Acta Hortic. Sin. 2009, 9, 1317–1326. [Google Scholar]
- Wang, S.J.; Lan, Y.C.; Chen, S.F.; Chen, Y.M.; Yeh, K.W. Wound-response regulation of the sweet potato sporamin gene promoter region. Plant Mol. Biol. 2002, 48, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Eun, C.H.; Kim, S.U.; Kim, I.J. The promoter from the Citrus unshiucarotenoid isomerase gene directs differential GUS expression in transgenic Arabidopsis. Mol. Breed. 2015, 35, 116. [Google Scholar] [CrossRef]
- Lü, S.; Gu, H.; Yuan, X.; Wang, X.; Wu, A.M.; Qu, L.; Liu, J.Y. The GUS reporter-aided analysis of the promoter activities of a rice metallothionein gene reveals different regulatory regions responsible for tissue-specific and inducible expression in transgenic Arabidopsis. Transgenic Res. 2007, 16, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-Glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J. Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant Physiol. 2001, 127, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, G.J.; Singh, M.; Chhabra, R. DNA-based diagnostics for genetically modified cotton: Decaplex PCR assay to differentiate MON531 and MON15985 Bt cotton events. Methods Mol. Biol. 2013, 958, 139–151. [Google Scholar] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | ORF Length (bp) | Predicted Protein | ||
|---|---|---|---|---|---|
| Length (aa) | MW (kDa) | pI | |||
| Da-dio1 | KX237676 | 822 | 273 | 30.89 | 7.02 |
| Da-dio2 | KX237677 | 822 | 273 | 31.29 | 6.17 |
| Da-dio3 | KX237678 | 822 | 273 | 30.97 | 7.02 |
| Da-dio4 | KX237679 | 831 | 276 | 31.31 | 6.25 |
| Da-dio5 | KX237680 | 819 | 272 | 31.01 | 6.45 |
| Name | Sequence | Function |
|---|---|---|
| GCN4_motif | CAAGCCA | involved in endosperm expression element |
| Skn-1_motif | GTCAT | required for endosperm expression regulatory element |
| ABRE | GCCACGTACA | abscisic acid-responsiveness element |
| AuxRR-core | GGTCCAT | auxin -responsiveness element |
| ERE | ATTTCAAA | ethylene-responsive element |
| GARE- motif | TCTGTTG | gibberellin-responsive element |
| P- box | CCTTTTG | gibberellin-responsive element |
| TCA-element | CCATCTTTTT | salicylic acid-responsiveness element |
| HSE | AAAAAATTTC | heat stress-responsiveness element |
| LTR | CCGAAA | low temperature-responsiveness |
| MBS | CAACTG(CGGTCA) | MYB binding site involved in drought-inducibility |
| TC-rich repeats | ATTTTCTTCA | defense and stress responsiveness element |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Huang, Y.; Huang, X.; Yang, J.; Wu, W.; Xu, Y.; Cong, Z.; Xie, J.; Xia, W.; Huang, D. Characterization of the Dioscorin Gene Family in Dioscorea alata Reveals a Role in Tuber Development and Environmental Response. Int. J. Mol. Sci. 2017, 18, 1579. https://doi.org/10.3390/ijms18071579
Liu L, Huang Y, Huang X, Yang J, Wu W, Xu Y, Cong Z, Xie J, Xia W, Huang D. Characterization of the Dioscorin Gene Family in Dioscorea alata Reveals a Role in Tuber Development and Environmental Response. International Journal of Molecular Sciences. 2017; 18(7):1579. https://doi.org/10.3390/ijms18071579
Chicago/Turabian StyleLiu, Linya, Yacheng Huang, Xiaolong Huang, Jianghua Yang, Wenqiang Wu, Yun Xu, Ziwen Cong, Jun Xie, Wei Xia, and Dongyi Huang. 2017. "Characterization of the Dioscorin Gene Family in Dioscorea alata Reveals a Role in Tuber Development and Environmental Response" International Journal of Molecular Sciences 18, no. 7: 1579. https://doi.org/10.3390/ijms18071579