Early Hippocampal i-LTP and LOX-1 Overexpression Induced by Anoxia: A Potential Role in Neurodegeneration in NPC Mouse Model

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
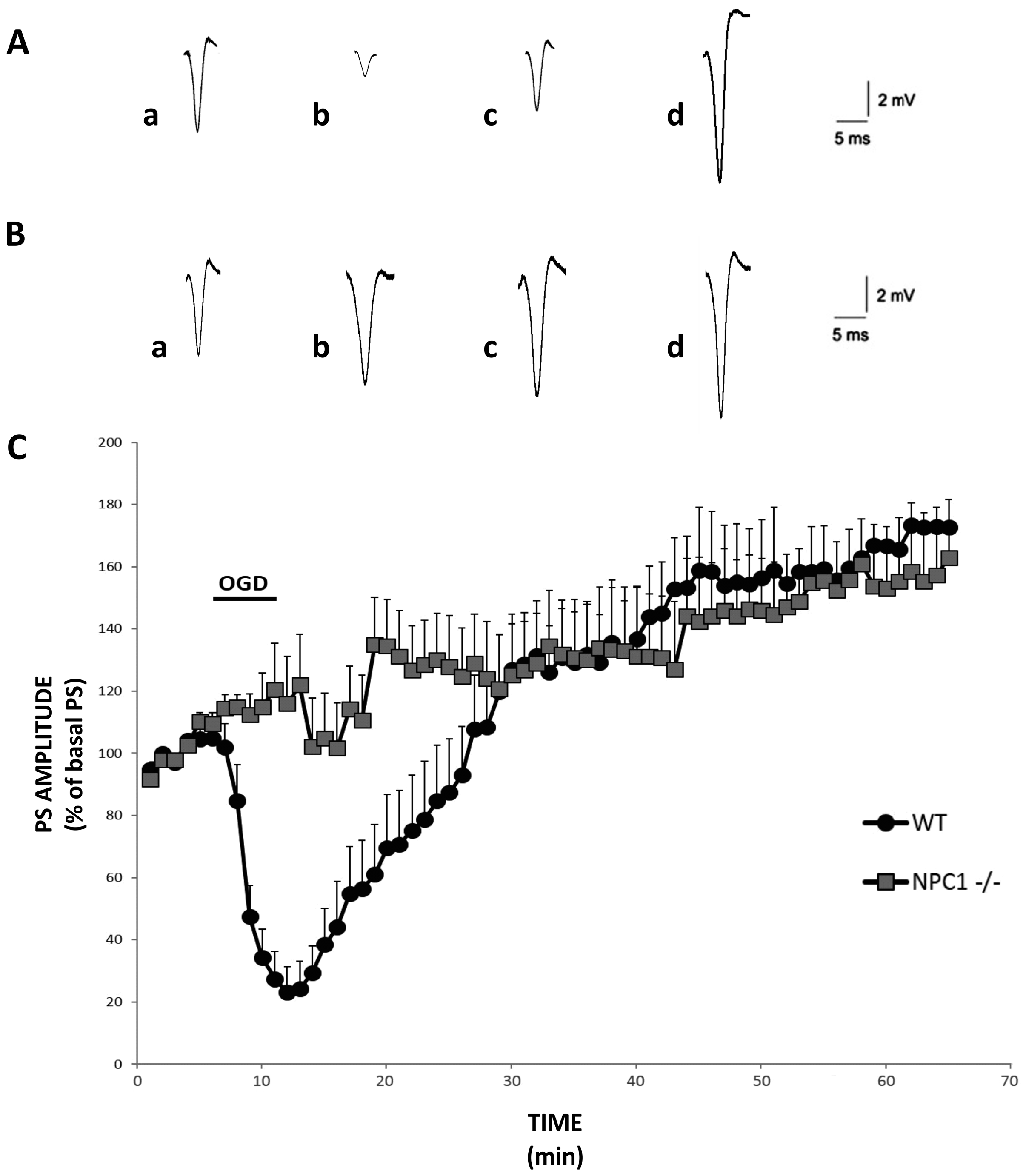
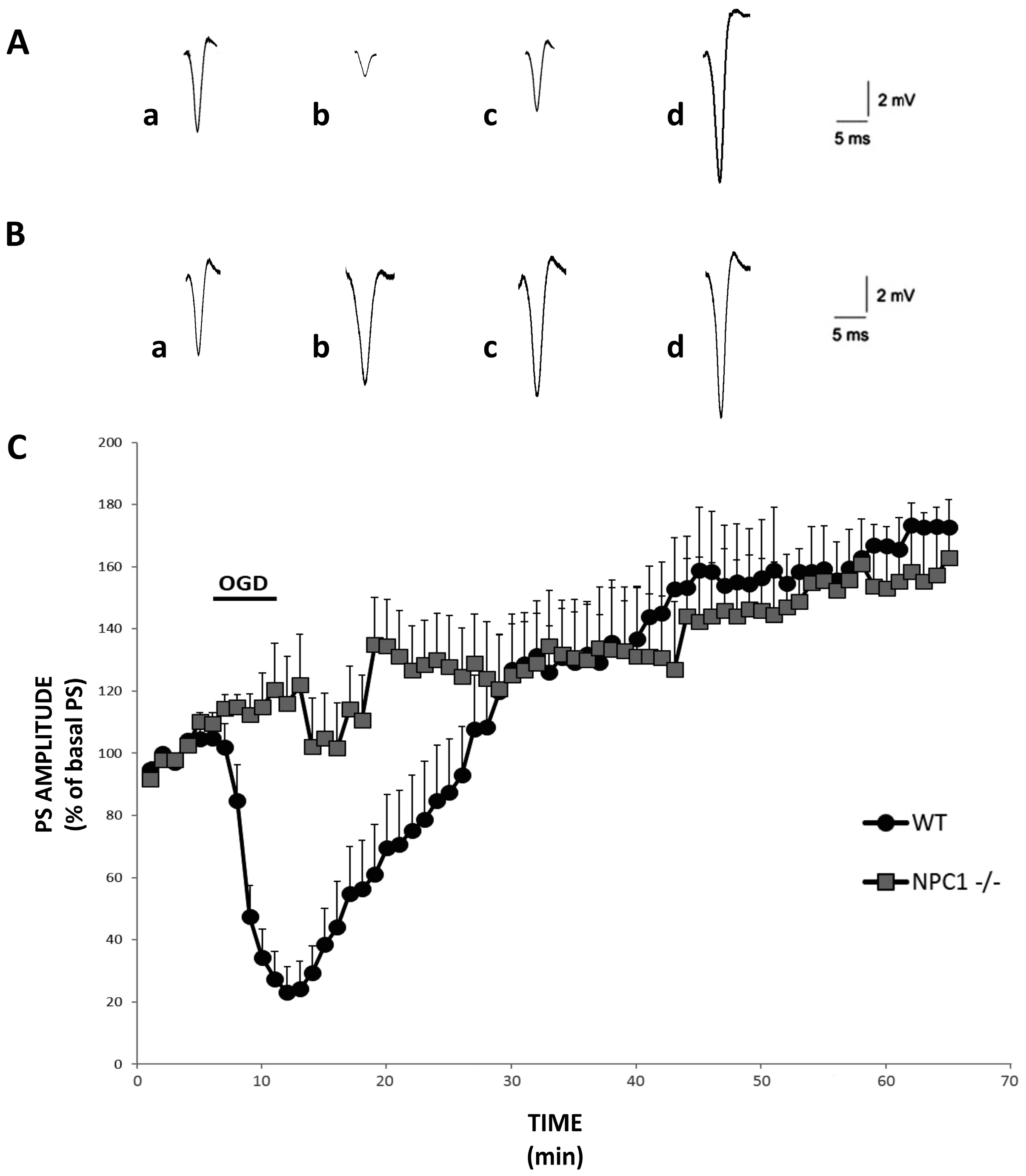
2.1. Precocious i-LTP in NPC1−/− Hippocampal Slices
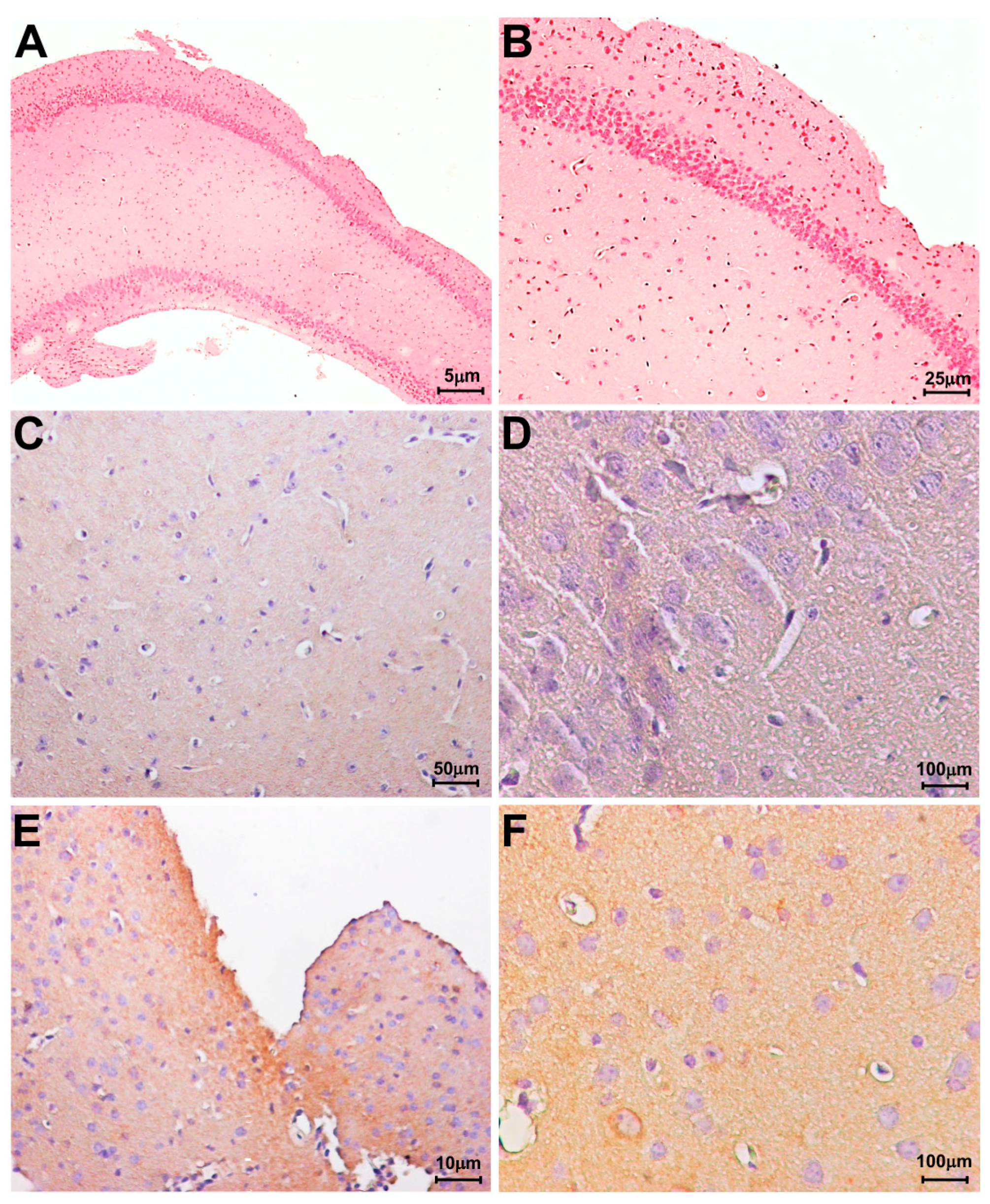
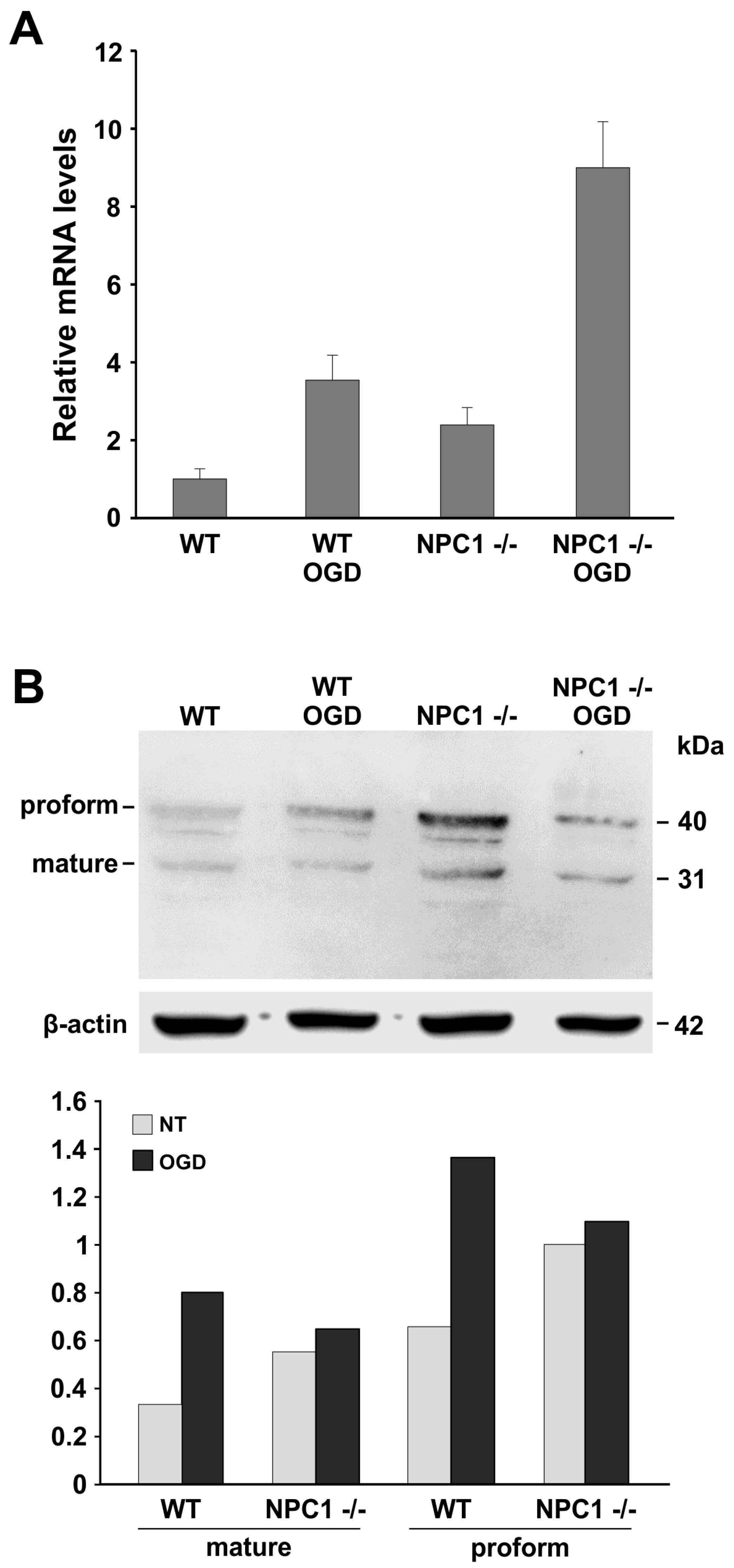
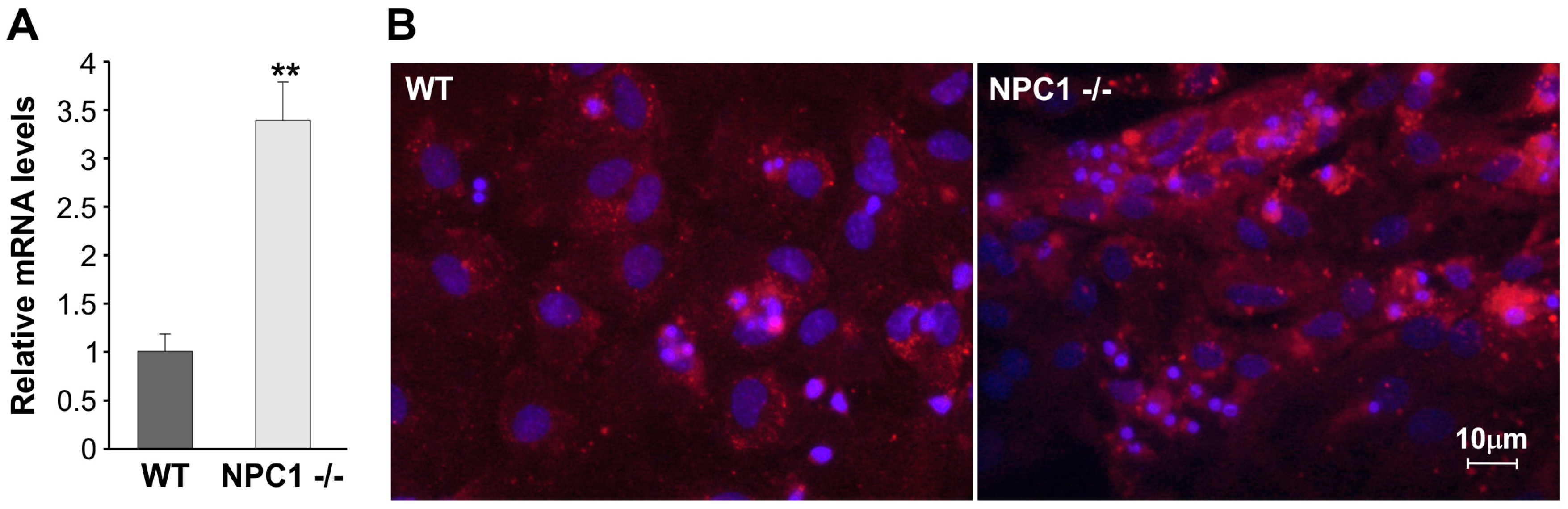
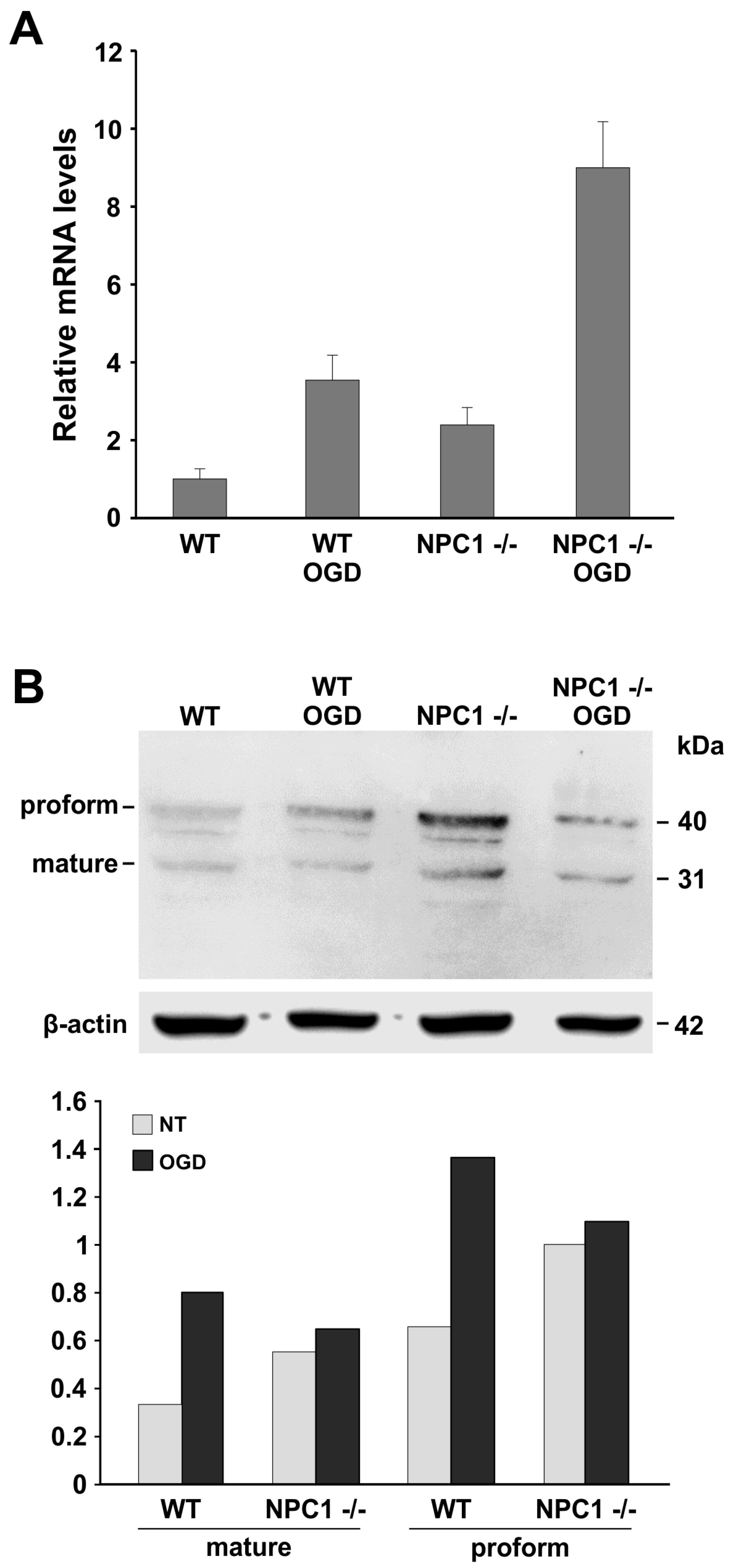
2.2. LOX-1 Analysis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Extracellular Recordings in Mouse Hippocampus
4.3. LOX-1 Expression Analysis
4.4. Immunocytochemistry and Western Blot Analyses of LOX-1 in Hippocampus
4.5. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Totenhagen, J.W.; Lope-Piedrafita, S.; Borbon, I.A.; Yoshimaru, E.S.; Erickson, R.P.; Trouard, T.P. In vivo assessment of neurodegeneration in Niemann-Pick type C mice by quantitative T2 mapping and diffusion tensor imaging. J. Magn. Reson. Imaging 2012, 35, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Zaaraoui, W.; Crespy, L.; Rico, A.; Faivre, A.; Soulier, E.; Confort-Gouny, S.; Cozzone, P.J.; Pelletier, J.; Ranjeva, J.P.; Kaphan, E.; et al. In vivo quantification of brain injury in adult Niemann-Pick Disease Type C. Mol. Genet. Metab. 2011, 103, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Auer, I.A.; Schmidt, M.L.; Lee, V.M.Y.; Curry, B.; Suzuki, K.; Shin, R.W.; Pentchev, P.G.; Carstea, E.D.; Trojanowski, J.Q. Paired helical filament tau (PHFtau) in Niemann-Pick type C disease is similar to PHFtau in Alzheimer’s disease. Acta Neuropathol. 1995, 90, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Millard, E.E.; Gale, S.E.; Dudley, N.; Zhang, J.; Schaffer, J.E.; Ory, D.S. The sterol-sensing domain of the Niemann-Pick C1 (NPC1) protein regulates trafficking of low density lipoprotein cholesterol. J. Biol. Chem. 2005, 80, 28581–28590. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.N.; Hanni, K.B.; Markesbery, W.R. Oxidized low-density lipoprotein induces neuronal death: Implications for calcium, reactive oxygen species, and caspases. J. Neurochem. 1999, 72, 2601–2609. [Google Scholar] [CrossRef] [PubMed]
- Sugawa, M.; Ikeda, S.; Kushima, Y.; Takashima, Y.; Cynshi, O. Oxidized low density lipoprotein caused CNS neuron cell death. Brain Res. 1997, 761, 165–172. [Google Scholar] [CrossRef]
- Sawamura, T.; Kume, N.; Aoyama, T.; Moriwaki, H.; Hoshikawa, H.; Aiba, Y.; Tanaka, T.; Miwa, S.; Katsura, Y.; Kita, T.; et al. An endothelial receptor for oxidized low density lipoprotein. Nature 1997, 386, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Xie, L.; Greenberg, D.A. LOX-1 expression and oxidized LDL uptake and toxicity in the HN33 neuronal cell line. Neurosci. Lett. 2014, 580, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Doyle, K.P.; Simon, R.P.; Stenzel-Poore, M.P. Mechanisms of ischemic brain damage. Neuropharmacology 2008, 55, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H.; Moskowitz, M.A.; Jacobs, T.P. Exciting, radical, suicidal: How brain cells die after stroke. Stroke 2005, 36, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Calabresi, P.; Saulle, E.; Centonze, D.; Pisani, A.; Marfia, G.A.; Bernardi, G. Post-ischaemic long-term synaptic potentiation in the striatum: A putative mechanism for cell type-specific vulnerability. Brain 2002, 125, 844–860. [Google Scholar] [CrossRef] [PubMed]
- Calabresi, P.; Centonze, D.; Pisani, A.; Cupini, L.M.; Bernardi, G. Synaptic plasticity in the ischaemic brain. Lancet Neurol. 2003, 2, 622–629. [Google Scholar] [CrossRef]
- Calabresi, P.; Cupini, L.M.; Centonze, D.; Pisani, F.; Bernardi, G. Antiepileptic drugs as a possible neuroprotective strategy in brain ischemia. Ann. Neurol. 2003, 53, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Karten, B.; Peake, K.B.; Vance, J.E. Mechanisms and consequences of impaired lipid trafficking in Niemann-Pick type C1-deficient mammalian cells. Biochim. Biophys. Acta 2009, 1791, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Rudolphi, K.A.; Schubert, P.; Parkinson, F.E.; Fredholm, B.B. Neuroprotective role of adenosine in cerebral ischaemia. Trends Pharmacol. Sci. 1992, 13, 439–445. [Google Scholar] [CrossRef]
- Sweeney, M.I. Neuroprotective effects of adenosine in cerebral ischemia: Window of opportunity. Neurosci. Biobehav. Rev. 1997, 21, 207–217. [Google Scholar] [CrossRef]
- Pearson, T.; Frenguelli, B.G. Volume-regulated anion channels do not contribute extracellular adenosine during the hypoxic depression of excitatory synaptic transmission in area CA1 of rat hippocampus. Eur. J. Neurosci. 2000, 12, 3064–3066. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.Y.; Xu, S.J.; Yan, Y.G.; Yu, H.M.; Ling, S.C.; Luo, J.H. Decreased purinergic inhibition of synaptic activity in a mouse model of Niemann-Pick disease type C. Hippocampus 2011, 21, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Nicoll, R.A. Long-term potentiation—A decade of progress? Science 1999, 285, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- D’Arcangelo, G.; Grossi, D.; de Chiara, G.; de Stefano, M.C.; Cortese, G.; Citro, G.; Rufini, S.; Tancredi, V.; Merlo, D.; Frank, C. Glutamatergic neurotransmission in a mouse model of Niemann–Pick Type C Disease. Brain Res. 2011, 1396, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Frank, C.; Rufini, S.; Tancredi, V.; Forcina, R.; Grossi, D.; D’arcangelo, G. Cholesterol depletion inhibits synaptic transmission and synaptic plasticity in rat hippocampus. Exp. Neurol. 2008, 212, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Hering, H.; Lin, C.C.; Sheng, M. Lipid rafts in the maintenance of synapses, dendritic spines, and surface AMPA receptor stability. J. Neurosci. 2003, 23, 3262–3271. [Google Scholar] [PubMed]
- Rufini, S.; Grossi, D.; Luly, P.; Tancredi, V.; Frank, C.; D’Arcangelo, G. Cholesterol depletion inhibits electrophysiological changes induced by anoxia in CA1 region of rat hippocampal slices. Brain Res. 2009, 1298, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. LOX-1 and neurodegeneration. Neurosci. Lett. 2014, 580, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Masaki, T.; Sawamura, T. LOX-1, the receptor for oxidized low-density lipoprotein identified from endotelial cells: Implications in endotelial dysfunction and atherosclerosis. Pharmacol. Ther. 2002, 95, 89–100. [Google Scholar] [CrossRef]
- Loftus, S.K.; Morris, J.A.; Carstea, E.D.; Gu, J.Z.; Cummings, C.; Brown, A.; Ellison, J.; Ohno, K.; Rosenfeld, M.A.; Tagle, D.A.; et al. Murine model of Niemann–Pick C disease: Mutation in a cholesterol homeostasis gene. Science 1997, 277, 232–235. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Castro, A.; Murdocca, M.; Pucci, S.; Zaratti, A.; Greggi, C.; Sangiuolo, F.; Tancredi, V.; Frank, C.; D’Arcangelo, G. Early Hippocampal i-LTP and LOX-1 Overexpression Induced by Anoxia: A Potential Role in Neurodegeneration in NPC Mouse Model. Int. J. Mol. Sci. 2017, 18, 1442. https://doi.org/10.3390/ijms18071442
Lo Castro A, Murdocca M, Pucci S, Zaratti A, Greggi C, Sangiuolo F, Tancredi V, Frank C, D’Arcangelo G. Early Hippocampal i-LTP and LOX-1 Overexpression Induced by Anoxia: A Potential Role in Neurodegeneration in NPC Mouse Model. International Journal of Molecular Sciences. 2017; 18(7):1442. https://doi.org/10.3390/ijms18071442
Chicago/Turabian StyleLo Castro, Adriana, Michela Murdocca, Sabina Pucci, Anna Zaratti, Chiara Greggi, Federica Sangiuolo, Virginia Tancredi, Claudio Frank, and Giovanna D’Arcangelo. 2017. "Early Hippocampal i-LTP and LOX-1 Overexpression Induced by Anoxia: A Potential Role in Neurodegeneration in NPC Mouse Model" International Journal of Molecular Sciences 18, no. 7: 1442. https://doi.org/10.3390/ijms18071442