Fortifying Horticultural Crops with Essential Amino Acids: A Review

1
College of Horticulture, South China Agricultural University, Guangzhou 510642, China
2
Department of Plant Science, Weizmann Institute of Science, Rehovot 76100, Israel
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(6), 1306; https://doi.org/10.3390/ijms18061306
Submission received: 17 May 2017
/
Revised: 13 June 2017
/
Accepted: 14 June 2017
/
Published: 19 June 2017
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:To feed the world’s growing population, increasing the yield of crops is not the only important factor, improving crop quality is also important, and it presents a significant challenge. Among the important crops, horticultural crops (particularly fruits and vegetables) provide numerous health compounds, such as vitamins, antioxidants, and amino acids. Essential amino acids are those that cannot be produced by the organism and, therefore, must be obtained from diet, particularly from meat, eggs, and milk, as well as a variety of plants. Extensive efforts have been devoted to increasing the levels of essential amino acids in plants. Yet, these efforts have been met with very little success due to the limited genetic resources for plant breeding and because high essential amino acid content is generally accompanied by limited plant growth. With a deep understanding of the biosynthetic pathways of essential amino acids and their interactions with the regulatory networks in plants, it should be possible to use genetic engineering to improve the essential amino acid content of horticultural plants, rendering these plants more nutritionally favorable crops. In the present report, we describe the recent advances in the enhancement of essential amino acids in horticultural plants and possible future directions towards their bio-fortification.

1. Introduction
Food quality is often defined as “everything a consumer would find desirable in a food product” [1]. In developing countries, people are prone to nutritional deficiency because plants account for the majority of their food. Therefore, improving the nutritional status of plants will help meet the dietary and micronutrient requirements of people in these countries. Conversely, in developed countries, consumers with higher income place a greater emphasis on healthy foods, such as fruits and vegetables, which provide various health compounds, such as vitamins and antioxidants, etc. Hence, improvement of the nutritional quality of horticultural crops is a relevant goal for overall sustainability.
Essential amino acids are those that are not produced in humans and farm animals; thus, they must be obtained from external sources, which are generally plants. There are nine essential amino acids, lysine (Lys), methionine (Met),threonine (Thr), phenylalanine (Phe), tryptophan (Trp), valine (Val), isoleucine (Ile), leucine (Leu), and histidine (His) [2]. The level of four of these amino acids, Lys, Met, Thr, and Trp, are the most limiting essential amino acids in plants, i.e., they are present in low quantities compared to the levels required for the optimum growth of humans and other animals [3]. Large populations in developing countries, whose diets are largely plant-derived, have insufficient levels of these essential amino acids, which can lead to major, devastating diseases. Therefore, to prevent such diseases, enriching the essential amino acid content of crops and horticultural plants (particularly fruits and vegetables), which are the major sources of human food and livestock feed in these countries is essential.
The knowledge obtained from genetic engineering research has been successfully applied to increase the content of some essential amino acids in crop plants. However, their application in horticultural crops is extremely limited. In the present report, we describe some of the approaches used to increase essential amino acid levels in horticultural crops, and evaluate the efficacy of these efforts towards the bio-fortification of horticultural plants.
2. Metabolism of Essential Amino Acids in Plants
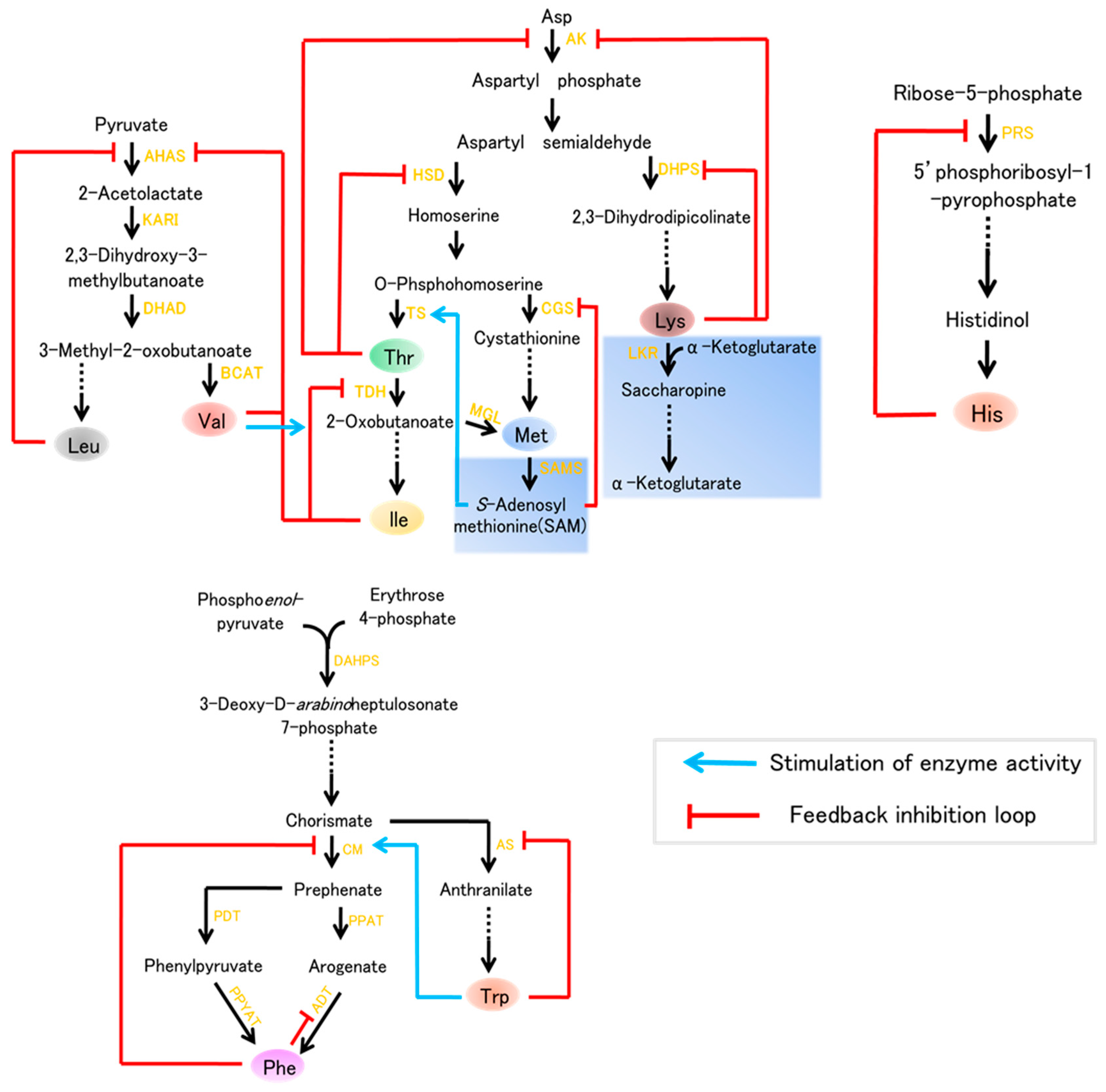
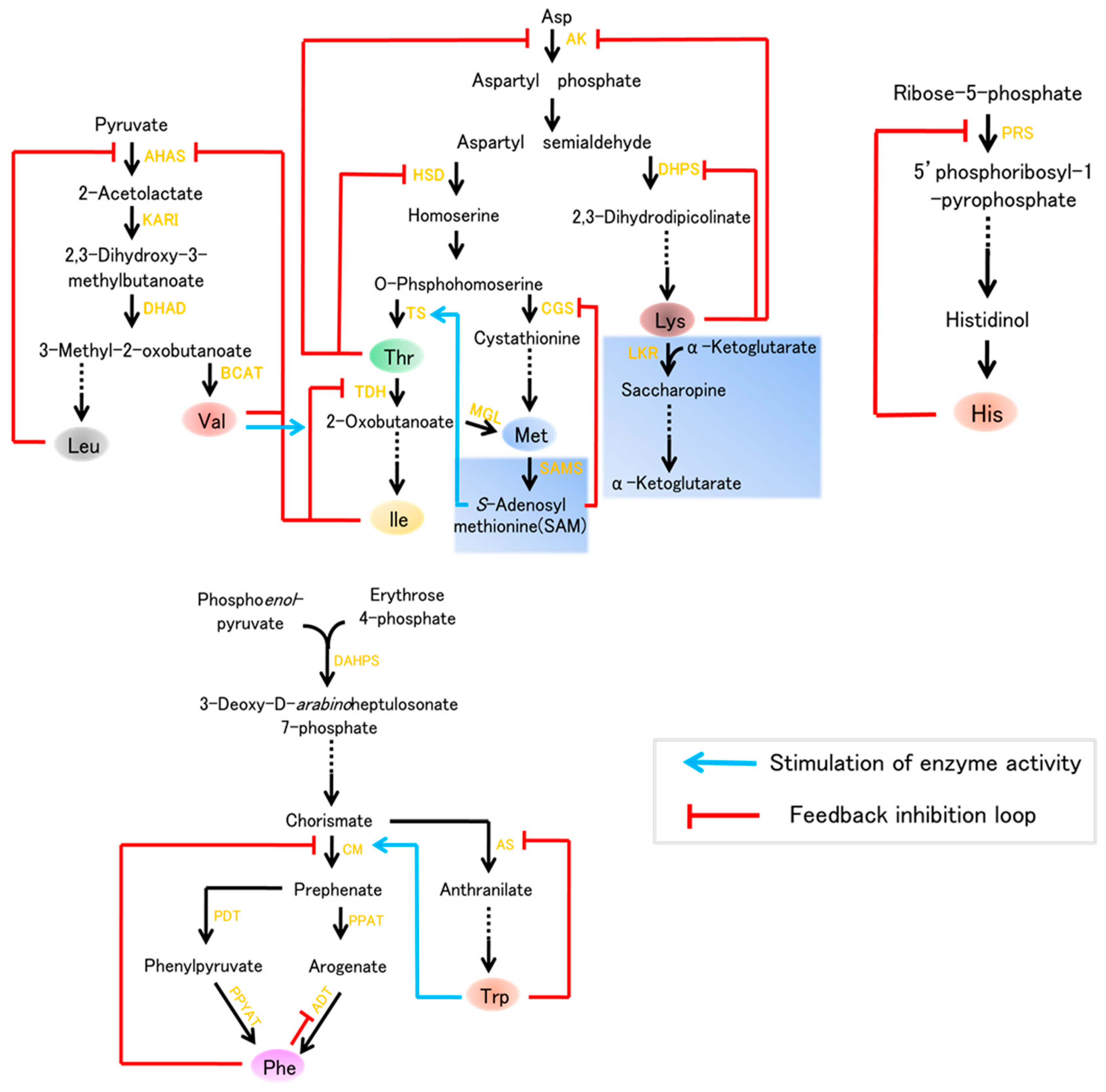
The Aspartate (Asp) family of amino acids and the biosynthetic pathways for Lys, Thr, Met, and Ile in plants are shown in Figure 1. Lysine is considered to be the most limiting essential amino acid in cereals and legumes, i.e., it is present in the smallest quantity [4]. Thus, extensive efforts have been made to improve the Lys content in plants, especially in seeds, which are the major sources of human food and livestock feed in developing countries. The pathway of lysine is primarily regulated by two key enzymes, namely aspartate kinase (AK), which is the first enzyme of Asp family pathway, and the feedback-insensitive dihydrodipicolinate synthase (DHPS), which is the first enzyme specific for lysine synthesis (Figure 1). However, lysine is efficiently degraded by its catabolism in to the tricarboxylic (TCA) cycle, a pathway initiated by the bi-functional enzyme lysine-ketoglutarate reductase (LKR)/saccharopine dehydrogenase (SDH) (Figure 1). Our understanding of lysine metabolism in plants dates to the 1960s, since the discovery of the maize high-lysine mutant opaque-2(o2) which contains a low content of lysine seed storage proteins (zeins) and, consequently, increased lysine [5]. With a deep understanding of biosynthesis of lysine, a number of studies aiming to improve the content concentrated on expressing feedback-insensitive DHPS or preventing lysine degrade into the TCA cycle.
Thr, Met, and Ile, three additional essential amino acids, which are generated via another branch of the Asp family biosynthetic pathway (Figure 1), also play critical roles in plant growth and human nutrition. In plants, aspartate kinase (AK), and homoserine dehydrogenase (HSD), two key enzymes of Asp pathway, occur as either mono- or bifunctional proteins which feedback regulated by Thr. Moreover, the level of Thr is regulated by its catabolism (Figure 1). The synthesis of Met initiates from an intermediate of the Asp pathway, O-phosphohomoserine. Additionally, the level of Met is also regulated by its catabolic enzymes. In particular, Met is the most limiting essential amino acid in cereal and legume crops, this limitation lead to nonspecific signs of protein deficiencies in humans, such as reduced resistance to diseases [3].
The Val biosynthetic pathway starts with pyruvate, and Leu biosynthesis starts with 3-methyl-2-oxobutanoate. There are four enzymes in Val and Leu biosynthesis: acetohydroxyacid synthase (AHAS), ketol acid reductoisomerase (KARI), dihydroxyaciddehydratase (DHAD), and branched-chain aminotransferase (BCAT). Val and Leu levels are also elevated by environmental stresses and are similarly elevated by drought stress, high light, and heat stress [6,7,8]. Two essential amino acids, Trp and Phe, are important aromatic amino acids (AAA) required for protein biosynthesis that are also the precursors of various natural products in plants, such as pigments, hormones, and alkaloids [9,10]. Trp is a precursor of alkaloids, phytoalexins, and the plant hormone auxin, whereas Phe is a common precursor of numerous phenolic compounds, such as flavonoids, condensed tannins, and phenylpropanoid/benzenoid volatiles [10,11,12]. All AAAs are derived from the conversion of phosphoenolpyruvate and erythrose 4-phosphate into chorismate via the shikimate pathway; individual postchorismate pathways then lead to the synthesis of Trp and Phe (Figure 1). These pathways are present in bacteria, fungi, and plants, but are absent in animals [13]. Thus, it is important to enhance the level of these AAAs in plants which are used as human food and livestock feed.
The research on His biosynthesis in plants is far behind studies in fungi and bacteria. The His biosynthetic pathway includes nine enzymes (Figure 1), which were identified in Arabidopsis, and increasing evidence has implicated the critical role of ATP-phosphorilbosyl transferase in the regulation of His biosynthesis.
Although the regulation of some essential amino acid biosynthetic pathways has been extensively studied in model plants, such as Lys and Met, those in horticultural plants are not fully understood, limiting breeding and engineering efforts to improve the levels of most essential amino acids.
3. Fortifying Horticultural Crops with the Essential Amino Acids Lys, Met, Thr, and Trp to Improve Their Nutritional Quality
In recent decades, traditional breeding methods and mutagenesis have been applied to enhance the essential amino acid content of crop plants, and in recent years, we have obtained a detailed understanding of the enzymes involved in essential amino acid biosynthesis, degradation, and regulation in Arabidopsis and other model plants. These studies have made it possible to apply genetic engineering approaches for improvements of essential amino acid levels in horticultural plants. However, increasing the levels of these essential amino acids in plants, especially in horticultural plants is still difficult because (i) the synthetic pathways of some essential amino acids, such as Lys, Leu, lle, Val, Phe, Trp, are strongly regulated by a negative feedback loop; and (ii) the targeted essential amino acids are efficiently degraded by catabolism, e.g., Lys, which is degraded in the tricarboxylic (TCA) cycle [14].
Lys levels are particularly low in crops, and extensive efforts have been made to improve Lys content in Arabidopsis and various crops, particularly rice and maize, leading to significantly higher levels of Lys [14,15,16,17,18,19]. Since Lys biosynthesis and catabolism are well characterized in plants, it was possible to achieve desirable results in horticultural plants. A number of the earliest studies aimed to improve lysine levels in horticultural plants concentrated on expressing the lysine feedback-intensive DHPS enzymes, such as expression in potato, soybean, and canola led to a notable increase in free lysine [20,21] (Table 1). Meanwhile, plants overexpressing bacterial DHPS often exhibited the typical abnormal phenotype, such as a partial loss of apical dominance, delayed flowering, and abnormal leaf morphology [15]. Furthermore, Hacham et al. crossed homozygous tobacco plants overexpressing both feedback-insensitive DHPS and AtCGS, the result showed the level of lysine is similar with those expressing only DHPS. Unexpectedly, the level of methionine was significantly increased in plants whose co-expression of both transgenes compared with the expression of AtCGS alone [22]. De nova expression of α-helical coiled-coil protein also improved lysine accumulation in tobacco seeds [23]. Lys is efficiently degraded by catabolism; therefore, another effective approach for increasing Lys levels is to prevent its degradation via the TCA cycle. However, aside from the results obtained in Arabidopsis, maize, and rice [14,16,17,18,19], none of these approach have been successful in horticultural plants.
Met is the most limiting essential amino acid in plants, and a number of traditional breeding methods, mutagenesis, and genetic engineering approaches have been applied to enhance Met levels. However, traditional plant breeding methods have been generally met with very limited success [3]; therefore, most current efforts are focused on using genetic engineering approaches in Arabidopsis and horticultural plants. Similar to lysine, most attempts to elevate content have been focused on enhancing the synthesis or reducing the catabolism of Met. Overexpression of cystathionine γ-synthase (CGS), which is the first enzyme in the Met biosynthesis pathway (Figure 1), led to a 6.5-, 12.8-, and 32.7-fold elevation in soluble Met content in transgenic potato, tobacco, and alfalfa leaves, respectively, revealing the regulatory role of CGS in Met accumulation in horticultural plants. Another effective approach for increasing Met content in horticultural plants involves the expression of sulfur-rich proteins, such as 2S albumin from Brazil nut (Bertholletia excelsa) and sunflower (Helianthus annuus). Transgenic expression of Brazil nut 2S albumin gene resulted in higher levels of Met in canola, tobacco, and Vicia [24,25,26,27].
Similar to Lys, Thr is synthesized through a branch of the Asp family pathway (Figure 1), and the first (Asp kinase, AK) and third (homoserine dehydrogenase, HSD) enzymes in the pathway are inhibited by Thr. Accumulating evidence has shown that AK is the major rate-limiting enzyme in Thr biosynthesis in plants. Previous studies demonstrated that mutant transgenic tobacco and alfalfa plants possessing AK or overexpressing AK showed a marked accumulation of Thr. [28,29]. Additionally, transgenic tobacco plants expressing Thr synthase (TS), the last enzyme in Thr biosynthesis, showed a five-fold accumulation of Thr [30]. Moreover, given that Thr and Met diverge from the same branch of the Asp synthesis pathway, their biosynthetic pathways compete, to some extent, for the same carbon substrate. Thus, CGS, the first unique enzyme in Met biosynthesis, likely plays an important role in Thr accumulation in horticultural plants [31].
Trp synthesis in plants is strongly regulated by feedback inhibition through the biosynthetic enzyme anthranilate synthase (AS), which catalyzes the first step in Trp biosynthesis, the conversion of chorismate to anthranilate (Figure 1). Some success has been achieved in overexpressing feedback insensitive AS to improve Trp content in Arabidopsis and crop plants, leading to significantly higher levels of Trp [32,33,34,35]. In horticultural plants, overexpression of AS is also an effective approach for enhancing Trp accumulation. Overexpression of AS led to a 10-fold elevation in free Trp content in transgenic tobacco and Astragalus sinicus, and a much greater increase (431-fold) in potato [36]. Moreover, recent studies showed that Trp levels are generally upregulated by some environmental conditions, such as light, water, and dark-induced senescence [37,38].
4. Conclusions
Traditional breeding has mostly failed to increase the levels of essential amino acids in plants, especially in horticultural plants, due to limited availability of genetic resources and mutants. In-depth research on the pathways of essential amino acids and their interactions with the regulatory networks in plants suggest that genetic engineering methods may be more promising. In general, three main approaches have been used to increase the levels of amino acids in plants, (i) enhance the efficiency of amino acid synthesis; (ii) prevent amino acid degradation (catabolism); and (iii) combine (i) and (ii) in the same plant. Although these approaches have been considerably successful in model and crop plants, in horticultural plants these studies are still in their infancy due to various reasons, such as the availability of efficient transformation technologies, incomplete genomic information, etc. So far, although biotechnology has been used to improve quality in crops and horticultural plants over 20 years, commercial cultivation of genetically modified (GM) crops, such as maize and cotton, has been approved in some countries, including in America, Brazil, India, and Africa [52] recently, Genome editing technology, represented by the clustered regularly interspaced short palindromic repeat(CRISPR)/Cas9 system, attain various types of genetic modification, subsequently providing a number of agricultural benefits in horticultural crops, including improvement of essential amino acids. However, as with all other GM plants, the opportunities for using such GM or genome-edited horticultural crops is dependent upon the conclusion of the public debate regarding the safety of GM food.
Acknowledgments
Base funds supported by Guangzhou Science and Technology Project key project (2014J4500033); Guangdong Science and Technology Project (2014B020202007).
Author Contributions
Wenyi Wang: drafted the manuscript and critically revised it; Mengyun Xu: revised the figures; Guoping Wang and Gad Galili: conceived the manuscript and critically revised it; All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| T | Transgene |
| WT | Wild type |
| SS | Seed-specific promoter |
| Sup | Suppression the gene by antisense or RNAi |
| M | Mutant |
| NR | Not reported |
| NS | Not significant |
| SSP | Seed storage protein |
| AtCGS | ARABIDOPSIS thaliana cystathionine γ-synthase |
| AtD-CGS | Feedback insensitive form of AtCGS |
| EcSAT | E. coli serine acetyl transferase |
| SAMS | S-adenosyl Met synthase |
| MGL | Met γ-lyase |
| TS | Thr synthase |
| 2S SSA | 2S sunflower seed albumin |
| 2S BN | 2S brazil nut albumin |
| bAK | Bacterial feedback-insensitive Asp kinase |
| bDHDPS | Bacterial feedback-insensitive dihydrodipicolinate synthase |
| LKR | Lys-ketoglutarate reductase |
| Bip | Lys-rich binding protein |
| sb401 | Lys-rich pollen-specific protein |
| RLRH | Rice Lys-rich histone proteins |
| OASA1D | Rice mutated feedback-resistant a subunit of rice anthranilate synthase |
| NtASA2 | Tobacco feedback-insensitive anthranilate synthase 2 |
References
- Grunert, K.G.; Jeppesen, L.F.; Jespersen, K.R.; Sonne, A.M.; Hansen, K.; Trondsen, T.; Young, J.A. Market orientation of value chains: A conceptual framework based on four case studies from the food industry. Eur. J. Mark. 2005, 39, 428–455. [Google Scholar] [CrossRef]
- Galili, G.; Höfgen, R. Metabolic engineering of amino acids and storage proteins in plants. Metab. Eng. 2002, 4, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R. Fortifying plants with the essential amino acids lysine and methionine to improve nutritional quality. Plant Biotechnol. J. 2013, 11, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Karchi, H.; Shaul, O.; Perl, A.; Cahana, A.; Tzchori, I.B.T.; Galili, S. Production of transgenic plants containing elevated levels of lysine and threonine. Biochem. Soc. Trans. 1994, 22, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science 1964, 145, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Mourad, G.; King, J. LO-methylthreonine-resistant mutant of Arabidopsis defective in isoleucine feedback regulation. Plant Physiol. 1995, 107, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Halgand, F.; Wessel, P.M.; Laprévote, O.; Dumas, R. Biochemical and mass spectrometric evidence for quaternary structure modifications of plant threonine deaminase induced by isoleucine. Biochemistry 2002, 41, 13767–13773. [Google Scholar] [CrossRef] [PubMed]
- Binder, S. Branched-chain amino acid metabolism in Arabidopsis thaliana. Arabidopsis Book 2010, 8, e0137. [Google Scholar] [CrossRef] [PubMed]
- Razal, R.A.; Ellis, S.; Singh, S.; Lewis, N.G.; Towers, G.N. Nitrogen recycling in phenylpropanoid metabolism. Phytochemistry 1996, 41, 31–35. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in Arabidopsis. Arabidopsis Book 2011, 9, e0152. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, P.F. Tetrahydropterin-dependent amino acid hydroxylases. Annu. Rev. Biochem. 1999, 68, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Galili, G. Transgenic high-lysine rice—A realistic solution to malnutrition? J. Exp. Bot. 2016, 67, 4009–4011. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O.; Galili, G. Concerted regulation of lysine and threonine synthesis in tobacco plants expressing bacterial feedback-insensitive aspartate kinase and dihydrodipicolinate synthase. Plant Mol. Biol. 1993, 23, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Tzchori, I.B.T.; Perl, A.; Galili, G. Lysine and threonine metabolism are subject to complex patterns of regulation in Arabidopsis. Plant Mol. Boil. 1996, 32, 727–734. [Google Scholar] [CrossRef]
- Zhu, X.; Galili, G. Increased lysine synthesis coupled with a knockout of its catabolism synergistically boosts lysine content and also transregulates the metabolism of other amino acids in Arabidopsis seeds. Plant Cell 2003, 15, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Galili, G. Lysine metabolism is concurrently regulated by synthesis and catabolism in both reproductive and vegetative tissues. Plant Physiol. 2004, 135, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Perl, A.; Shaul, O.; Galili, G. Regulation of lysine synthesis in transgenic potato plants expressing a bacterial dihydrodipicolinate synthase in their chloroplasts. Plant Mol. Biol. 1992, 19, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Falco, S.C.; Guida, T.; Locke, M.; Mauvais, J.; Sanders, C.; Ward, R.T.; Webber, P. Transgenic canola and soybean seeds with increased lysine. Nat. Biotechnol. 1995, 13, 577–582. [Google Scholar] [CrossRef]
- Hacham, Y.; Song, L.; Schuster, G.; Amir, R. Lysine enhances methionine content by modulating the expression of S-adenosylmethionine synthase. Plant J. 2007, 51, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Karchi, H.; Shaul, O.; Galili, G. Lysine synthesis and catabolism are coordinately regulated during tobacco seed development. Proc. Nat. Acad. Sci. USA 1994, 91, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Pearson, K.W.; Meeker, G.; Staraci, L.C.; Sun, S.S. Enhancement of the methionine content of seed proteins by the expression of a chimeric gene encoding a methionine-rich protein in transgenic plants. Plant. Mol. Biol. 1989, 13, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Kuo, C.C.; Staraci, L.C.; Pearson, K.W.; Wainwright, C.; Georgescu, A.; Townsend, J. Accumulation of a Brazil nut albumin in seeds of transgenic canola results in enhanced levels of seed protein methionine. Plant. Mol. Biol. 1992, 18, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Molvig, L.; Tabe, L.M.; Eggum, B.O.; Moore, A.E.; Craig, S.; Spencer, D.; Higgins, T.J. Enhanced methionine levels and increased nutritive value of seeds of transgenic lupins (Lupinus angustifolius L.) expressing a sunflower seed albumin gene. Proc. Nat. Acad. Sci. USA 1997, 94, 8393–8398. [Google Scholar] [CrossRef] [PubMed]
- Tiger, T.T.; Miki, M.C.; Rolis, C.W.; Chen, L.J.; Su, R.C.; Wang, C.S.; Jason, T.C. Enhanced methionine and cysteine levels in transgenic rice seeds by the accumulation of sesame 2S albumin. Biosci. Biotechnol. Biochem. 2003, 67, 1699–1705. [Google Scholar]
- Frankard, V.; Ghislain, M.; Negrutiu, I.; Jacobs, M. High threonine producer mutant of Nicotiana sylvestris (Spegg. and Comes). Theor. Appl. Genet. 1991, 82, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Karchi, H.; Shaul, O.; Galili, G. Seed-specific expression of a bacterial desensitized aspartate kinase increases the production of seed threonine and methionine in transgenic tobacco. Plant J. 1993, 3, 721–727. [Google Scholar] [CrossRef]
- Hacham, Y.; Matityahu, I.; Schuster, G.; Amir, R. Overexpression of mutated forms of aspartate kinase and cystathionine γ-synthase in tobacco leaves resulted in the high accumulation of methionine and threonine. Plant J. 2008, 54, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Zeh, M.; Casazza, A.P.; Kreft, O.; Roessner, U.; Bieberich, K.; Willmitzer, L.; Hesse, H. Antisense inhibition of threonine synthase leads to high methionine content in transgenic potato plants. Plant Physiol. 2001, 127, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Kreps, J.A.; Ponappa, T.; Dong, W.; Town, C.D. Molecular basis of α-methyltryptophan resistance in amt-1, a mutant of Arabidopsis thaliana with altered tryptophan metabolism. Plant Physiol. 1996, 110, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Last, R.L. The Arabidopsis thaliana Trp5 mutant has a feedback-resistant anthranilate synthase and elevated soluble tryptophan. Plant Physiol. 1996, 110, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, K.; Hasegawa, H.; Nemoto, H.; Matsuda, F.; Miyazawa, H.; Tozawa, Y.; Miyagawa, H. High-level tryptophan accumulation in seeds of transgenic rice and its limited effects on agronomic traits and seed metabolite profile. J. Exp. Bot. 2006, 57, 3069–3078. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Asada, Y.; Takahashi, Y.; Yabe, N.; Komeda, Y.; Nishioka, T.; Wakasa, K. Metabolic changes in Arabidopsis thaliana expressing the feedback-resistant anthranilate synthase α subunit gene OASA1D. Phytochemistry 2006, 67, 2349–2362. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Tozawa, Y.; Hasegawa, H.; Terakawa, T.; Ohkawa, Y.; Wakasa, K. Use of a feedback-insensitive α subunit of anthranilate synthase as a selectable marker for transformation of rice and potato. Mol. Breed. 2004, 14, 363–373. [Google Scholar] [CrossRef]
- Araújo, W.L.; Ishizaki, K.; Nunes-Nesi, A.; Larson, T.R.; Tohge, T.; Krahnert, I.; Leaver, C.J. Identification of the 2-hydroxyglutarate and isovaleryl-CoA dehydrogenases as alternative electron donors linking lysine catabolism to the electron transport chain of Arabidopsis mitochondria. Plant Cell 2010, 22, 1549–1563. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R.; Fernie, A.R. The regulation of essential amino acid synthesis and accumulation in plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Shaul, O.; Galili, G. Threonine overproduction in transgenic tobacco plants expressing a mutant desensitized aspartate kinase of Escherichia coli. Plant Physiol. 1992, 100, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Guenoune, D.; Amir, R.; Badani, H.; Wolf, S.; Galili, S. Combined expression of S-VSPα in two different organelles enhances its accumulation and total lysine production in leaves of transgenic tobacco plants. J. Exp. Bot. 2002, 53, 1867–1870. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Bauw, G.; van Montagu, M.; Inzé, D. Distinct phenotypes generated by overexpression and suppression of S-adenosyl-l-methionine synthetase reveal developmental patterns of gene silencing in tobacco. Plant Cell 1994, 6, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- Saalbach, I.; Waddell, D.; Pickardt, T.; Schieder, O.; Müntz, K. Stable expression of the sulphur-rich 2S albumin gene in transgenic Vicia narbonensis increases the methionine content of seeds. J. Plant Physiol. 1995, 145, 674–681. [Google Scholar] [CrossRef]
- Di, R.; Kim, J.; Martin, M.N.; Leustek, T.; Jhoo, J.; Ho, C.T.; Tumer, N.E. Enhancement of the primary flavor compound methional in potato by increasing the level of soluble methionine. J. Agric. Food Chem. 2003, 51, 5695–5702. [Google Scholar] [CrossRef] [PubMed]
- Demidov, D.; Horstmann, C.; Meixner, M.; Pickardt, T.; Saalbach, I.; Galili, G.; Müntz, K. Additive effects of the feed-back insensitive bacterial aspartate kinase and the Brazil nut 2S albumin on the methionine content of transgenic narbon bean (Vicia narbonensis L.). Mol. Breed. 2003, 11, 187–201. [Google Scholar] [CrossRef]
- Chiaiese, P.; Ohkama-Ohtsu, N.; Molvig, L.; Godfree, R.; Dove, H.; Hocart, C.; Tabe, L.M. Sulphur and nitrogen nutrition influence the response of chickpea seeds to an added, transgenic sink for organic sulphur. J. Exp. Bot. 2004, 55, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Amira, G.; Ifat, M.; Tal, A.; Hana, B.; Shmuel, G.; Rachel, A. Soluble methionine enhances accumulation of a 15 kDa zein, a methionine-rich storage protein, in transgenic alfalfa but not in transgenic tobacco plants. J. Exp. Bot. 2005, 56, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- Dancs, G.; Kondrák, M.; Bánfalvi, Z. The effects of enhanced methionine synthesis on amino acid and anthocyanin content of potato tubers. BMC Plant Biol. 2008, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, M.S.; Rahman, S.M.; Nakamoto, Y.; Fujiwara, T.; Naito, S.; Wakasa, K.; Ishimoto, M. Differential response of methionine metabolism in two grain legumes, soybean and azuki bean, expressing a mutated form of Arabidopsis cystathionine γ-synthase. J. Plant Physiol. 2013, 170, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Brotherton, J.E.; Song, H.S.; Widholm, J.M. Increasing tryptophan synthesis in a forage legume Astragalus sinicus by expressing the tobacco feedback-insensitive anthranilate synthase (ASA2) gene. Plant Physiol. 2000, 123, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Brotherton, J.E.; Widholm, J.M.; Portis, A.R. Targeting a nuclear anthranilate synthase α-subunit gene to the tobacco plastid genome results in enhanced tryptophan biosynthesis. Return of a gene to its pre-endosymbiotic origin. Plant Physiol. 2001, 127, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Nakamoto, Y.; Takahashi, M.; Kitamura, K.; Wakasa, K.; Ishimoto, M. Manipulation of amino acid composition in soybean seeds by the combination of deregulated tryptophan biosynthesis and storage protein deficiency. Plant Cell Rep. 2010, 29, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Araki, M. Consumer acceptance of food crops developed by genome editing. Plant Cell Rep. 2016, 35, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Nine essential amino acids biosynthesis leading to lysine (Lys), methionine (Met),threonine (Thr), phenylalanine (Phe), tryptophan (Trp), valine (Val), isoleucine (Ile), leucine (Leu), and histidine (His) in plants. Enzymes are indicated in yellow text. The catabolism indicated by the light blue boxes. Abbreviations: AK, Asp kinase; HSD, homoserine dehydrogenase; DHDPS, dihydrodipicolinate synthase; LKR, lysine ketoglutaric acid reductase; TS, Thr synthase; CGS, cystathionine γ-synthase; TDH, Thr dehydratase; PRS, ribose-phosphate diphosphokinase; MGL, Met γ-synthase; SAMS, S-adenosylmethionine synthase; AHAS, acetohydroxyacid synthase; KARI, ketol acid reductoisomerase; DHAD, dihydroxyacid dehydratase; BCAT, branched-chain aminotransferase; DAHPS, 3-deoxy-d-arabinoheptulosonate 7-phosphate synthase; CM, chorismate mutase; AS, anthranilate synthase; PDT, prephenate dehydratase; PPAT, prephenate aminotransferase; PPYAT, phenylpyruvate aminotransferase; ADT, arogenate dehydratase.
Figure 1.
Nine essential amino acids biosynthesis leading to lysine (Lys), methionine (Met),threonine (Thr), phenylalanine (Phe), tryptophan (Trp), valine (Val), isoleucine (Ile), leucine (Leu), and histidine (His) in plants. Enzymes are indicated in yellow text. The catabolism indicated by the light blue boxes. Abbreviations: AK, Asp kinase; HSD, homoserine dehydrogenase; DHDPS, dihydrodipicolinate synthase; LKR, lysine ketoglutaric acid reductase; TS, Thr synthase; CGS, cystathionine γ-synthase; TDH, Thr dehydratase; PRS, ribose-phosphate diphosphokinase; MGL, Met γ-synthase; SAMS, S-adenosylmethionine synthase; AHAS, acetohydroxyacid synthase; KARI, ketol acid reductoisomerase; DHAD, dihydroxyacid dehydratase; BCAT, branched-chain aminotransferase; DAHPS, 3-deoxy-d-arabinoheptulosonate 7-phosphate synthase; CM, chorismate mutase; AS, anthranilate synthase; PDT, prephenate dehydratase; PPAT, prephenate aminotransferase; PPYAT, phenylpyruvate aminotransferase; ADT, arogenate dehydratase.

{kind=link}
{kind=link}
Table 1.
Increase the levels of essential amino acids lysine (Lys), methionine (Met), threonine (Thr), and tryptophan (Trp) in transgenic horticultural plants.
Table 1.
Increase the levels of essential amino acids lysine (Lys), methionine (Met), threonine (Thr), and tryptophan (Trp) in transgenic horticultural plants.
| Amino Acid | Plant | Transgene | Tissue | Free fold Increase T/WT (Souble) | Phenotype | References |
|---|---|---|---|---|---|---|
| Lys | Potato | Constitutive::DHDPS | Leaves | 2–4 | ND | [20] |
| Potato | Constitutive::DHDPS | Roots | 1.2–5 | ND | [20] | |
| Potato | Constitutive::DHDPS | Tubers | 2–3.5 | ND | [20] | |
| Tobacco | Constitutive::AK | Leaves | 1.1–2 | ND | [39] | |
| Tobacco | Constitutive::DHDPS | Leaves | 9–11 | A partial loss of apical dominance, delayed flowering and senescence, partial sterility, and abnormal leaf morphology | [15] | |
| Tobacco | Seed-specific::DHDPS | Seeds | 1–2 | ND | [23] | |
| Canola | Seed-specific::DHDPS/AK | Seeds | More than 100 | ND | [21] | |
| Soybean | Seed-specific::DHDPS/AK | Seeds | Several hundreds | Wrinkled seed with poor germinaiton | [21] | |
| Soybean | Soybean vegetative storage proteins(S-VSPs) | Leaves | NR | ND | [40] | |
| Tobacco | Constitutive::DHDPS | Leaves | 30 | Mosaic green color in newly developed leaves at the tip of the apex and partial loss of apical dominance | [22] | |
| Tobacco | Constitutive::DHDPS/Constitutive::AtCGS | Leaves | 24 | ND | [22] | |
| Thr | Tobacco | Constitutive::AK | Leaves | 2–8 | ND | [39] |
| Tobacco | Constitutive::DHDPS | Leaves | 6–8 | A partial loss of apical dominance, delayed flowering and senescence, partial sterility,and abnormal leaf morphology | [15] | |
| Tobacco | Seed-specific::DHDPS | Seeds | 1 | ND | [23] | |
| Tobacco | Supress AK | Seeds | About 5.8 | ND | [23] | |
| Tobacco | Seed-specific::DHDPS/AK | Seeds | About 3.9 | ND | [23] | |
| Tobacco | Constitutive::AtCGS | Leaves | 1.8 | ND | [22] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 173 | ND | [30] | |
| Met | Tobacco | antisense SAMS | Leaves | 433 | Vein showed dark gren | [41] |
| Vicia narbonensis | Seed-specific::2S BN/AK | Seeds | 2 | ND | [42] | |
| Lupin | Seed-specific::2S SSA | Seeds | 2 | Statistically significant increases in live weight gain, true protein digestibility, biological value, and net protein utilization | [26] | |
| Potato | Antisense TS | Leaves | 239 | Severe growth retardation, strong chlorosis, and an acute reduction in tuber yield | [31] | |
| Potato | Constitutive::AtCGS | Leaves | 2–7.1 | ND | [43] | |
| Potato | Constitutive::AtCGS | Root | 1.2–4.1 | ND | [43] | |
| Bean | Seed-specific::2S BN | Seeds | 1.25 | ND | [44] | |
| Chickpea | Seed-specific::2S SSA | Seeds | 1.9 | ND | [45] | |
| Alfalfa | Constitutive::AtCGS | Leaves | 37 | ND | [46] | |
| Tobacco | Constitutive::DHDPS | Leaves | 2 | ND | [22] | |
| Tobacco | Constitutive::AtCGS | Leaves | 5 | ND | [22] | |
| Tobacco | Constitutive::DHDPS/Constitutive::AtCGS | Leaves | 10 | ND | [22] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | [22] | ||
| Potato | Constitutive::AtCGS/zein | Tubers | 2–6 | ND | [47] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | ND | [30] | |
| Soybean | Seed -specific::AtCGS | Seeds | 2.1 | ND | [48] | |
| Azuki bean | Seed -specific::AtCGS | Seeds | 2.3 | ND | [48] | |
| Trp | Astragalus | Constitutive::ASA2 | Hariy roots | 1.3–5.5 | ND | [49] |
| Tobacoo | Constitutive::ASA2 | Leaves | 16 | ND | [50] | |
| Tobacco | Constitutive::CGS/AK | Leaves | 39 | ND | [30] | |
| Soybean | Seed -specific::OASA1D | Seeds | 50.9 | ND | [51] |
Abbreviations: DHDPS, feedback-insensitive dihydrodipicolinate synthase; AK, feedback-insensitive Asp kinase; AtCGS, Arabidopsis thaliana cystathionine γ-synthase; TS, Thr synthase; 2S SSA, 2S sunflower seed albumin; 2S BN, 2S brazil nut albumin; OASA1D, rice mutated feedback-resistant a subunit of rice anthranilate synthase; NtASA2, tobacco feedback-insensitive anthranilate synthase 2; ND, not detected.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, G.; Xu, M.; Wang, W.; Galili, G. Fortifying Horticultural Crops with Essential Amino Acids: A Review. Int. J. Mol. Sci. 2017, 18, 1306. https://doi.org/10.3390/ijms18061306
AMA Style
Wang G, Xu M, Wang W, Galili G. Fortifying Horticultural Crops with Essential Amino Acids: A Review. International Journal of Molecular Sciences. 2017; 18(6):1306. https://doi.org/10.3390/ijms18061306
Chicago/Turabian StyleWang, Guoping, Mengyun Xu, Wenyi Wang, and Gad Galili. 2017. "Fortifying Horticultural Crops with Essential Amino Acids: A Review" International Journal of Molecular Sciences 18, no. 6: 1306. https://doi.org/10.3390/ijms18061306
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.