The Roles of Long Non-Protein-Coding RNAs in Osteo-Adipogenic Lineage Commitment

Department of Calcified Tissue Biology, Hiroshima University Institute of Biomedical and Health Sciences, 734-8553 Hiroshima, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(6), 1236; https://doi.org/10.3390/ijms18061236
Submission received: 15 May 2017
/
Revised: 5 June 2017
/
Accepted: 6 June 2017
/
Published: 9 June 2017
(This article belongs to the Special Issue Advances in Bone and Cartilage Research)
Abstract
:Osteoblasts and adipocytes share a common mesenchymal progenitor in the bone marrow. This implies that a reciprocal relationship exists between osteogenic and adipogenic differentiation. Further, cells of osteoblast lineage transdifferentiate into adipocytes under some circumstances. Dysregulation of osteo-adipogenic fate-determination leads to bone diseases such as osteoporosis, accompanied by an increase in bone marrow adipose tissue. Thus, the fine-tuning of osteo-adipogenesis is necessary for bone homeostasis. Osteo-adipogenic progression is governed by a complex crosstalk of extrinsic signals, transcription factors, and epigenetic factors. Long non-protein-coding RNAs (lncRNAs) act in part as epigenetic regulators in a broad range of biological activities, such as chromatin organization, transcriptional regulation, post-translational modifications, and histone modification. In this review, we highlight the roles of epigenetic regulators, particularly lncRNAs, in the osteo-adipogenic lineage commitment of bone marrow mesenchymal stem cells and the adipogenic transdifferentiation of osteoblasts.
1. Introduction
Bone marrow mesenchymal stem cells (BMSCs) differentiate into multiple different cell types such as osteoblasts, adipocytes, and chondrocytes. BMSCs are also known to play an essential role in maintaining the hematopoietic stem cell niche by providing modulatory signals [1,2]. The bone marrow cavity is filled with active hematopoietic cells (red marrow) in newborns, and these cells are replaced with BMSC-derived adipocytes with age, resulting in the formation of marrow adipose tissue (MAT) [3]. MAT also accumulates in reaction to chemotherapy, glucocorticoid therapy, and radiotherapy [4]. Increased MAT is associated with bone pathologies including decreased cortical bone, low bone mineral density, decreased bone volume, and decreased bone formation rate [5]. Conversely, lipodystrophic mice that carry a hypomorphic allele of PPARγ, a master regulator of adipogenesis [6], show increased bone mass, with a reduction in bone marrow cavity [7]. The current understanding of the mechanism underlying MAT accumulation is that it occurs due to the increased expression of PPARγ which causes the commitment of BMSCs to the adipogenic lineage [8,9]. Thus, an inverse correlation exists between osteogenesis and adipogenesis, and the fine-tuning of the osteo-adipogenic fate-choice is crucial for bone and bone marrow health. Recent discoveries of non-protein-coding RNAs (ncRNAs) have opened an exciting new field of the epigenetic mechanisms involved in fate-choice decisions and differentiation processes in many different cell types [10,11]. For many years, ncRNAs had been considered as accessory components to aid protein activities. Non-coding XIST RNA was first identified to be directly involved in the inactivation of the X chromosome for dosage compensation [12]. Thereafter, multiple approaches, especially computational biology and RNomics, have led to the identification of numerous ncRNAs, such as microRNAs (miRNAs), lncRNAs, small nucleolar RNA, enhancer RNAs, and promoter-associated RNAs [10,11]. In this review, we highlight recent studies describing the epigenetic processes by which lncRNAs regulate osteogenic versus adipogenic differentiation in the bone marrow.
2. Epigenetics and ncRNAs
Lineage commitment of stem cells proceeds via multi-step, hierarchical processes under the control of transcription factors. With the onset of differentiation, stem cell plasticity becomes restricted due to acquisition of epigenetic modifications, such as DNA methylation and histone modifications, which impose cellular identity [13]. DNA methylation is a reversible biochemical process involving the addition of a methyl (–CH3) group to the C-5 position of cytosine in CpG dinucleotides [14]. This process is catalyzed by a family of DNA methyltransferases, including the maintenance methyltransferase DNMT1 and de novo methyltransferases DNMT3A and DNMT3B [14]. Histone modifications are also reversible processes where the N-terminal tails of histone proteins are subjected to post-translational modifications, such as acetylation, methylation, phosphorylation, and ubiquitylation [15]. These modifications are catalyzed by specific enzymes such as histone acetyltransferases, histone deacetylases (HDACs), histone methyltransferases, histone demethylases, and so on [15]. DNA methylation and histone modifications influence one another to form and stabilize higher-order chromatin structure [16].
Recent studies have uncovered the important role of ncRNA transcripts in epigenetic gene regulation. ncRNAs are primarily classified into two categories, short ncRNAs (<200 nucleotides) and long ncRNAs (lncRNAs, >200 nucleotides) [10,11]. Short ncRNAs include miRNAs that are approximately 22 nucleotides in length and usually bind to 3′ untranslated regions of target mRNAs to repress transcription and translation (see [17,18,19,20] for review). The targets of miRNAs are not only protein-coding RNAs but also lncRNAs [21]. DNA methylation regulates the expression of miRNAs, such as miR-10a and miR-200, in human HCT116 cells [22]. In the opposite direction, the miR-290 family of miRNAs controls de novo DNA methylation in mouse embryonic stem cells [23]. LncRNAs regulate gene expression either in a cis- or trans-acting manner through multiple mechanisms (Figure 1): (1) serve as reservoirs of miRNAs [24,25], (2) act as miRNA sponges (competitive inhibitors of miRNAs) [26], (3) bind to complementary DNA sequences and form RNA-DNA triplex structures that can block the transcriptional process [27], (4) bind to pre-mRNA or mRNA and modulate posttranscriptional events such as splicing, degradation, and translation [28], and (5) recruit chromatin remodeling complexes to specific DNA loci [26,29,30]. (See other excellent reviews for more details, [31,32,33]) Thus, ncRNAs, DNA methylation, and histone modifications are mutually required to regulate their modifiers and chromatin signature, leading to a cascade of complex gene regulation events.
3. Epigenetics in Osteo-Adipogenesis
Lineage commitment of BMSCs is controlled by various extracellular cues such as transforming growth factor-β (TGF-β), WNT, Notch, Hedgehog, fibroblast growth factors, and insulin-like growth factors (see [34,35,36,37] for review). These molecules have dual roles in regulating osteoblastogenesis vs. adipogenesis. For instance, TGF-β acts as a bidirectional regulator of osteoblastogenesis by activating the SMAD-dependent pathways and the mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) pathways [37]. The former recruits HDAC4 and HDAC5 to repress expression of the osteoblast master regulator RUNX2 in rat osteogenic ROS17/2.8 cells [38,39]. The latter phosphorylates RUNX2 to promote its transcriptional activity [40]. Overall, TGF-β acts on multiple steps of osteogenesis not only by promoting osteoprogenitor cell proliferation, early differentiation, and chemotaxis, but also by inhibiting osteoblast maturation, matrix mineralization, and osteocyte differentiation [37]. Concomitant with reduced adipogenesis in mouse 3T3-L1 and 3T3-F442A cells treated with TGF-β [41,42,43], transgenic mice overexpressing TGF-β show impaired development of adipose tissue [44]. This happens via the ERK pathway that phosphorylates PPARγ to inhibit its transcriptional activity and suppresses adipocyte differentiation [45,46,47]. In the case of WNT signaling, the β-catenin-dependent pathway activates osteoblastogenesis by directly stimulating RUNX2 gene expression and by suppressing the adipogenic transcription factor genes CEBPA and PPARG in mouse stromal ST2 cells, as well as in 3T3-L1 cells [48,49,50,51]. KDM2A and KDM2B, demethylases of the trimethylation of histone H3 at lysine 4 (H3K4me3) and lysine 36 (H3K36me3), mediate the degradation of non-phosphorylated β-catenin in the nucleus of human embryonic kidney cells (HEK293T), suggesting that they have an inhibitory effect on osteogenesis [52]. Indeed, KDM2A/2B demethylases inhibit osteoblast differentiation by demethylating H3K4me3 and H3K36me3 in the promoter of EREG and TFAP2A in human mesenchymal stem cells (MSCs) [53,54]. On the other hand, β-catenin-independent WNT4/5A/5B signaling stimulates adipogenesis in 3T3-L1 cells [55,56,57]. Emerging evidence points to crosstalk between these signaling pathways and miRNAs [58]. MiR-181a targets genes involved in TGF-β signaling, such as TGFBI and TGFBR1, which are negative regulators of osteoblast differentiation [59]. MiR-346 promotes osteoblast differentiation via suppression of GSK3B to prevent the degradation of β-catenin in human BMSCs [60]. MiR-23a/b regulates the balance of osteo-adipogenic differentiation in human BMSCs by targeting TMEM64, which modulates WNT/β-catenin signaling [61,62]. These signaling pathways culminate in the activation of either RUNX2 or PPARγ [6,38]. RUNX2 and PPARγ reciprocally regulate one another during osteogenic and adipogenic differentiation, respectively, of BMSCs and adipose tissue-derived stem cells [47,63,64,65,66].
The cell-intrinsic epigenetic mechanisms underlying osteo-adipogenic lineage commitment have also been well studied. HDACs, key factors for bone formation [67] (see above), deacetylate N-terminal tails of histones and RUNX2 [68], thus mediating transcriptional repression and stability respectively. HDAC3, HDAC7, and HDAC8 interact with RUNX2 and act as a co-repressor of osteoblast-specific genes, such as OCN, BSP, and RUNX2, in mouse MC3T3-E1 and C2C12 cell lines, and rat BMSCs [69,70,71]. Osteochondral lineage specific HDAC3 knockout mice show severe reductions in trabecular bone, bone formation rates, and osteoblast numbers, accompanied with increased MAT [72]. Mice lacking HDAC8 in neural crest progenitor cells display ossification defects in frontal and interparietal bones [73]. Further studies are needed to resolve discrepancies between in vivo gene knockout studies and in vitro studies. HDACs also play a vital role in adipocyte differentiation. Inhibition of HDAC activity using sodium butyrate in 3T3-L1 cells induces adipogenic gene expression and adipocyte differentiation [74]. Preadipocytes isolated from HDAC9-knockout mice exhibit accelerated differentiation [75]. Besides, Jumonji C domain-containing histone lysine demethylases (e.g., the KDM family, NO66, RBP2, and PHF2) and SET domain-containing histone lysine methyltransferases (e.g., EZH2, SETDB1, and SETD8) are involved in osteo-adipogenic differentiation (see above for KDMs). For instance, KDM4B/6B demethylates the trimethylation of histone H3 at lysines 9 (H3K9me3) and 27 (H3K27me3), activating RUNX2, SP7, DLX5, BMP, and HOX genes. This results in enhanced osteoblast differentiation as demonstrated in MC3T3-E1 cells and human MSCs [76,77]. MiR-20a promotes adipocyte differentiation of ST2 cells by targeting KDM6B directly [78]. Preadipocytes isolated from mouse knockouts of EZH2, a gene that methylates H3K27, exhibit severe defects in adipogenic activity, concomitant with the derepression of WNT1/6/10A/10B genes, and with the activation of WNT/β-catenin signaling [79]. Deletion of EZH2 in mesenchymal progenitor cells causes malformation of the growth plate, cranial suture, and the trabecular bone, accompanied by accumulation of MAT [80]. Thus, many epigenetic modifiers contribute to the balance of osteo-adipogenic differentiation (for more details see [81]).
4. Functional Roles of lncRNAs in TGF-β- and WNT-Dependent Osteo-Adipogenesis
As mentioned above, TGF-β and WNT signaling pathways are involved in osteo-adipogenesis [37,41,42,43,44,48,49,50,51,55,56,57]. These signaling pathways are modulated by lncRNAs and associated regulators, including miRNAs and histone modifiers (Table 1). The maternally expressed imprinted gene H19 (2.3-kb lncRNA) acts as a primary miRNA precursor of miR-675. Its expression increases during osteoblast differentiation but decreases during adipocyte differentiation in human MSCs and BMSCs [24,25,26]. MiR-675 decreases the levels of mRNAs and proteins of TGF-β1 and class II histone deacetylases (HDAC4/5/6) in human MSCs [24]. HDAC4/5 repressor complexes are recruited by TGF-β1-activated SMAD2/3 to the RUNX2 promoter region to suppress its transcription in ROS17/2.8 cells [39]. Consistent with these roles, miR-675 may indirectly increase RUNX2 expression and osteoblast differentiation in human MSCs [24]. Overexpression of miR-675 in human BMSCs inhibits adipogenic differentiation through the downregulation of class II HDACs [25,82]. H19 also acts as a miRNA sponge that captures miR-141, miR-22, miR-200a, and let-7 to inhibit their respective functions (in human BMSCs, HT-29 cells, and HEK293 cells [26,83,84]). MiR-141, miR-22, and miR-200a inactivate the WNT/β-catenin signaling pathway by suppressing β-catenin expression in several cell types, including human BMSCs, SGC7901 cells, and U251 cells [26,85]. MiR-141 and miR-200a suppress the expression of DLX5, a negative regulator of adipogenesis [86], with a resultant increase in osteoblast differentiation of MC3T3-E1 cells [87]. The role of let-7 in osteoblasts remains unknown, though this miRNA promotes adipogenesis in 3T3-L1 cells [88]. Thus, the regulatory effects of H19 on osteo-adipogenesis are determined by its partner miRNAs.
The lncRNA MEG3 has been shown to interact with chromatin and act as a repressive regulator in human breast cancer cells by RNA immunoprecipitation-coupled high-throughput sequencing [30]. MEG3 interacts with the PRC2 complex and suppresses the expression of genes involved in the TGF-β pathway, such as TGFB2, TGFBR1, and SMAD2, in human breast cancer cells [30], thus inhibiting the SMAD-dependent signaling pathway. In human BMSCs, increasing the expression of MEG3 activates BMP4 transcription and promotes osteogenic differentiation [93]. The DNA methylation status of MEG3 affects its expression in human leukocytes [94], and therefore, miR-29-mediated suppression of the DNA methyltransferases DNMT1 and DNMT3b elevates the expression of MEG3, as demonstrated in hepatocellular cancer [95]. TGF-β1-mediated SMAD2/3 signaling negatively regulates the expression of miR-29 in mouse C2C12 myoblasts and human pancreatic stellate cells [96,97]. In human osteoblasts, canonical WNT signaling induces the expression of miR-29 which attenuates the effect of WNT signaling by targeting the WNT antagonists Dickkopf-1, Kremen2, and secreted frizzled related protein 2 [98]. It has also been shown that miR-29 promotes osteoblast differentiation by downregulating anti-osteogenic factors, such as HDAC4, TGFB3, ACVR2A, CTNNBIP1, and DUSP2, in MC3T3-E1 cells [99]. Thus, the MEG3-miR-29 regulatory circuitry may promote osteoblast differentiation, but its function on adipogenic differentiation remains unclear.
A recent, comprehensive survey of lncRNAs expressed differentially between bovine preadipocytes and differentiated adipocytes identified a novel lncRNA, ADNCR, that inhibits adipogenic differentiation by acting as a sponge of miR-204 [89]. Reduced expression of ADNCR with the onset of adipogenic differentiation increases free-functioning miR-204 that downregulates the target genes RUNX2 and SIRT1 [89,90]. Disruption of RUNX2 in knockout mice shows maturation arrest of osteoblasts [38]. In addition, the suppression of RUNX2 is indispensable for adipogenesis in 3T3-L1 cells [100]. SIRT1 is an adipogenic inhibitor; therefore, downregulation of SIRT1 in C3H10T1/2 cells may promote adipogenic differentiation [101]. In addition, miR-204 suppresses WNT/β-catenin signaling by modulating DVL3 expression, which promotes adipogenic differentiation of human adipose-derived MSCs [91].
In contrast to the above-mentioned lncRNAs, lncRNA HOXA-AS3 has been identified as a promoter of adipogenesis [92]. The number of HOXA-AS3 transcripts is found to increase gradually during adipogenesis of human MSCs. It interacts with EZH2 to recruit it to the RUNX2 promoter and causes subsequent suppression of RUNX2 and osteogenesis [92]. EZH2 catalyzes the methylation of H3K27 to form a repressive chromatin structure [81], and to repress WNT genes to facilitate adipogenesis [79]. HOXA-AS3 may act on this process.
Thus, complex crosstalk among signaling molecules, ncRNAs, DNA methylases, and histone modifiers is crucial for achieving a balance between osteoblast and adipocyte lineage commitment.
5. Epigenetics in Osteo-Adipogenic Transdifferentiation
It was believed that differentiation was a unidirectional process and a terminal event to yield a stable cellular identity. However, as demonstrated by induced pluripotent stem cells, terminally differentiated cells can be experimentally reprogramed to function as pluripotent cells [102]. This conversion of cell fate is also observed in vivo during the healing process of tissues, such as intestine, liver, and so on [103]. Bone injury in zebrafish induces mature osteoblasts to dedifferentiate into osteoprogenitor cells that become a source of new reparative bone [104]. Mammalian mature osteoblasts appear not to behave as undifferentiated cells during bone healing processes [105]. However, rat calvaria-derived osteoblasts show a marked plasticity of phenotypes even at the maturation stage [106], suggesting that osteoblasts have the potential to switch their cell fates to other cell lineages by inductive extracellular cues. Indeed, we and other groups have shown fate shifts of osteoblast-lineage cells into adipocytes under certain conditions [107,108,109,110,111]. For instance, a subset of relatively mature rat osteoblasts expressing PPARγ become adipocytes when cultured with the synthetic PPARγ agonist rosiglitazone [108]. Loss of osteogenic WNT/β-catenin signaling in early osteoblast-lineage cells in vivo increases the number of adipocytes [109]. Recently, a subpopulation of CXC chemokine ligand 12-abundant reticular (CAR) cells that expresses mature osteoblast markers was identified [112]. CAR cells are mesenchymal progenitor cells having osteo-adipogenic potential [113]. Thus, a subpopulation of osteoblasts may transdifferentiate to the adipogenic lineage. These findings raise a question about how cell-intrinsic epigenetic programs participate in osteo-adipogenic transdifferentiation.
Epigenetic modifications undoubtedly serve as important regulators of cell plasticity and transdifferentiation potential [114]. Among multiple histone modifications, bivalent modifications of histone H3, through both active and repressive marking by H3K4me3 and H3K27me3, respectively, are thought to play a vital role in keeping genes poised for rapid activation in response to appropriate signals [115,116,117,118]. The presence of these modifications in a subset of genes is thought to correlate with cell plasticity [115,116,117,118]. Recent genome-wide analyses of histone modifications in mouse BMSCs reveal that the most highly expressed genes at stages of osteogenic commitment allow bivalent modifications before expressing them [119]. H3K27me3 removal activates specific genes that bestow osteogenic identities upon cells and reduce their plasticity [119]. Consistent with this and as mentioned above, histone demethylases of H3K27me3 such as KDM4B/5B/6A/6B play a pivotal role in osteogenic commitment [76,77,120,121]. Interestingly, the genes downregulated during osteogenesis rarely gain H3K27me3 [119], suggesting the absence of active mechanisms for repression during osteogenic differentiation. In contrast, EZH2 mediated H3K27me3 is vital for adipogenic lineage specification [120]. Genome-wide comparison of chromatin profiles during the osteo-adipogenic differentiation of BMSCs suggests a default preference for osteogenic differentiation [111]. Gene activation-associated histone modifications in BMSCs are also found in osteogenic differentiated cells, but not in adipogenic differentiated cells, indicating that osteoblasts encode innate plasticity through the epigenome [111]. Thus, the chromatin state of osteoblasts may allow them to switch their cell fate relatively easily.
Further studies are warranted to understand the mechanisms by which epigenetic modifiers, including ncRNAs, regulate the plasticity of osteoblasts. There is an increasing body of evidence suggesting that histone variants and nuclear architectures are involved in cell plasticity [114,122]. The epigenetic program may be even more complicated than has been discovered so far for determining osteo-adipogenic commitment.
6. Concluding Remarks and Outlook
Many epigenetic factors, including ncRNAs, play an important role in the fate choice of BMSCs and enable crosstalk among multiple signaling pathways. Genome-wide studies show aging-associated depletion of DNA methylation in humans and mice [123,124,125,126]. Since DNMT1 haploinsufficiency promotes age-related health problems, including decreased bone mineral density [127], the genome-wide age-related decrease in DNA methylation may be attributed to a reduction of DNMT1 activity [128]. Similarly, aging-associated alterations are observed in the context of chromatin organization [129]. The histone deacetylase SIRT1 is known to function as a longevity factor, and its deficiency leads to premature aging, in parallel with global enrichment of acetylation at lysine 16 of histone H4 [130]. SIRT1 osteoblast-specific knockout mice show a significant decrease in trabecular bone volume and trabecular thickness [131]. Downregulation of HDAC1 and HDAC2 is also observed during senescence of human MSCs, followed by downregulation of EZH2 and upregulation of KDM6B [132]. These age-related alterations of histone-modifying enzymes contribute to perturbation in heterochromatin structure with aging [128]. Changes in the expression of ncRNAs are also believed to be associated with age-associated diseases and degenerative diseases [133,134]. Cell identity is mediated by epigenetics, so epigenetic aberrations may affect the plasticity of both stem and differentiated cells [135]. This idea is supported by recent evidence that biological senescence and tissue injury promotes epigenetic cellular reprogramming in vivo [136]. Therefore, age-related increases in PPARγ and/or adipogenic miRNA expression may affect not only the fate-choice decisions of BMSCs, but also the plasticity of the osteoblast population [8,137]. Whether osteoblasts convert their fates into adipocytes in vivo still remains controversial. Lineage tracing studies demonstrate the potential of cells in the mesenchymal lineage to transdifferentiate, for instance, hypertrophic chondrocytes transdifferentiate into osteoblasts during the endochondral bone formation and fracture-healing processes [138,139]. Forced expression of transcription factors and ncRNAs bestows the ability upon cells to directly convert from one cell type to another in vivo [140,141]. Given these observations, epigenetic alterations with age may induce transdifferentiation of osteoblasts into adipocytes, which may explain age-associated MAT accumulation. We believe that functional studies of ncRNAs and their interacting partners during osteo-adipogenesis may open new prospects for the prevention of MAT accumulation and bone loss in the future.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| lncRNAs | Long non-coding RNAs |
| BMSCs | Bone marrow mesenchymal stem cells |
| MAT | Marrow adipose tissue |
| ncRNAs | Non-protein-coding RNAs |
| miRNAs | MicroRNAs |
| HDACs | Histone deacetylases |
| TGF-β | Transforming growth factor-β |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated kinase |
| H3K4me3 | Trimethylation of histone H3 at lysine 4 |
| H3K36me3 | Trimethylation of histone H3 at lysine 36 |
| MSCs | Mesenchymal stem cells |
| H3K9me3 | Trimethylation of histone H3 at lysine 9 |
| H3K27me3 | Trimethylation of histone H3 at lysine 27 |
| CAR cells | CXC chemokine ligand 12-abundant reticular cells |
References
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Scheller, E.L.; Cawthorn, W.P.; Burr, A.A.; Horowitz, M.C.; MacDougald, O.A. Marrow adipose tissue: Trimming the fat. Trends Endocrinol. Metab. 2016, 27, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, K.R.; Hui, S.K.; Xian, C.J. Regulatory pathways associated with bone loss and bone marrow adiposity caused by aging, chemotherapy, glucocorticoid therapy and radiotherapy. Am. J. Stem Cells 2012, 1, 205–224. [Google Scholar] [PubMed]
- Scheller, E.L.; Rosen, C.J. What’s the matter with MAT? Marrow adipose tissue, metabolism, and skeletal health. Ann. N. Y. Acad. Sci. 2014, 1311, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Cock, T.-A.; Back, J.; Elefteriou, F.; Karsenty, G.; Kastner, P.; Chan, S.; Auwerx, J. Enhanced bone formation in lipodystrophic PPARγhyp/hyp mice relocates haematopoiesis to the spleen. EMBO Rep. 2004, 5, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Moerman, E.J.; Teng, K.; Lipschitz, D.A.; Lecka-Czernik, B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: The role of PPAR-γ2 transcription factor and TGF-β/BMP signaling pathways. Aging Cell 2004, 3, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Brennan, T.A.; Russell, E.; Kim, J.-H.; Chen, Q.; Johnson, F.B.; Pignolo, R.J. Aging alters bone-fat reciprocity by shifting in vivo mesenchymal precursor cell fate towards an adipogenic lineage. Bone 2016, 85, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Kaikkonen, M.; Lam, M.; Glass, C. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Peschansky, V.J.; Wahlestedt, C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 2013, 9, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Panning, B.; Jaenisch, R. RNA and the epigenetic regulation of X chromosome inactivation. Cell 1998, 93, 305–308. [Google Scholar] [CrossRef]
- Mohn, F.; Schübeler, D. Genetics and epigenetics: Stability and plasticity during cellular differentiation. Trends Genet. 2009, 25, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.D.; Meissner, A. DNA methylation: Roles in mammalian development. Nat. Rev. Genet. 2013, 14, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Zentner, G.E.; Henikoff, S. Regulation of nucleosome dynamics by histone modifications. Nat. Struct. Mol. Biol. 2013, 20, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Ivey, K.; Srivastava, D. MicroRNAs as regulators of differentiation and cell fate decisions. Cell Stem Cell 2010, 7, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Taipaleenmäki, H.; Hokland, L.; Chen, L.; Kauppinen, S.; Kassem, M. Micro-RNAs: Targets for enhancing osteoblast differentiation and bone formation. Eur. J. Endocrinol. 2012, 166, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, N.V. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Hamam, D.; Ali, D.; Kassem, M.; Aldahmash, A.; Alajez, N. MicroRNAs as regulators of adipogenic differentiation of mesenchymal stem cells. Stem Cells Dev. 2015, 24, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Zhang, C.-Y.; Zen, K. miRNA regulates noncoding RNA: A noncanonical function model. Trends Biochem. Sci. 2012, 37, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Witmer, P.D.; Casey, E.; Valle, D.; Sukumar, S. DNA methylation regulates microRNA expression. Cancer Biol. Ther. 2007, 6, 1290–1294. [Google Scholar] [CrossRef]
- Sinkkonen, L.; Hugenschmidt, T.; Berninger, P.; Gaidatzis, D.; Mohn, F.; Artus-Revel, C.; Zavolan, M.; Svoboda, P.; Filipowicz, W. MicroRNAs control de novo DNA methylation through regulation of transcriptional repressors in mouse embryonic stem cells. Nat. Struct. Mol. Biol. 2008, 15, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zheng, Y.; Jia, L.; Li, W. Long noncoding RNA H19 promotes osteoblast differentiation via TGF-β1/Smad3/HDAC signaling pathway by deriving miR-675. Stem Cells 2015, 33, 3481–3492. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zheng, Y.; Jin, C.; Li, X.; Jia, L.; Li, W. Long non-coding RNA H19 inhibits adipocyte differentiation of bone marrow mesenchymal stem cells through epigenetic modulation of histone deacetylases. Sci. Rep. 2016, 6, 28897. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.-C.; Fu, W.-M.; Wang, Y.-B.; Sun, Y.-X.; Xu, L.-L.; Wong, C.-W.; Chan, K.-M.; Li, G.; Waye, M.M.; Zhang, J.-F. H19 activates Wnt signaling and promotes osteoblast differentiation by functioning as a competing endogenous RNA. Sci. Rep. 2016, 6, 20121. [Google Scholar] [CrossRef] [PubMed]
- Martianov, I.; Ramadass, A.; Barros, A.; Chow, N.; Akoulitchev, A. Repression of the human dihydrofolate reductase gene by a non-coding interfering transcript. Nature 2007, 445, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Mondal, T.; Mohammad, F.; Enroth, S.; Redrup, L.; Komorowski, J.; Nagano, T.; Mancini-DiNardo, D.; Kanduri, C. Kcnq1ot1 antisense noncoding RNA mediates lineage-specific transcriptional silencing through chromatin-level regulation. Mol. Cell 2008, 32, 232–246. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA-DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chang, H. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L. Linking long noncoding RNA localization and function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- James, A. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica 2013, 2013, 684736. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, A.D.; Olsen, B.R. Osteoblast-adipocyte lineage plasticity in tissue development, maintenance and pathology. Cell. Mol. Life Sci. 2014, 71, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.-H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef]
- Kang, J.; Alliston, T.; Delston, R.; Derynck, R. Repression of Runx2 function by TGF-β through recruitment of class II histone deacetylases by Smad3. EMBO J. 2005, 24, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Yang, Q.; Zhao, G.; Yu, H.; Kirkwood, K.; Franceschi, R. Interactions between extracellular signal-regulated kinase 1/2 and P38 Map kinase pathways in the control of RUNX2 phosphorylation and transcriptional activity. J. Bone Miner. Res. 2012, 27, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N.; Tremblay, E.; McAdam, L.; Roberts, A.; Elliott, B. Autocrine secretion of TGF-β1 and TGF-β2 by pre-adipocytes and adipocytes: A potent negative regulator of adipocyte differentiation and proliferation of mammary carcinoma cells. Vitro Cell. Dev. Biol. 1998, 34, 412–420. [Google Scholar] [CrossRef]
- Choy, L.; Skillington, J.; Derynck, R. Roles of autocrine TGF-β receptor and Smad signaling in adipocyte differentiation. J. Cell Biol. 2000, 149, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Choy, L.; Derynck, R. Transforming growth factor-β inhibits adipocyte differentiation by Smad3 interacting with CCAAT/enhancer-binding protein (C/EBP) and repressing C/EBP transactivation function. J. Biol. Chem. 2003, 278, 9609–9619. [Google Scholar] [CrossRef] [PubMed]
- Clouthier, D.E.; Comerford, S.A.; Hammer, R.E. Hepatic fibrosis, glomerulosclerosis, and a lipodystrophy-like syndrome in PEPCK-TGF-β1 transgenic mice. J. Clin. Investig. 1997, 100, 2697–2713. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.; Kim, J.B.; Sarraf, P.; Spiegelman, B.M. Inhibition of adipogenesis through MAP kinase–mediated phosphorylation of PPARγ. Science 1999, 274, 2100–2103. [Google Scholar] [CrossRef]
- Beekum, O.; Fleskens, V.; Kalkhoven, E. Posttranslational modifications of PPAR-γ: Fine-tuning the metabolic master regulator. Obesity 2009, 17, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Cawthorn, W.P.; Li, Y.; Zhao, G.; MacDougald, O.A.; Franceschi, R.T. Reciprocal control of osteogenic and adipogenic differentiation by ERK/MAP kinase phosphorylation of Runx2 and PPARγ transcription factors. J. Cell. Physiol. 2016, 231, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar] [CrossRef] [PubMed]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.N.; Komm, B.S.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [PubMed]
- Kennell, J.A.; MacDougald, O.A. Wnt signaling inhibits adipogenesis through β-catenin-dependent and -independent mechanisms. J. Biol. Chem. 2005, 280, 24004–24010. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Bennett, C.N.; Gerin, I.; Rapp, L.A.; Hankenson, K.D.; MacDougald, O.A. Wnt signaling stimulates osteoblastogenesis of mesenchymal precursors by suppressing CCAAT/enhancer-binding protein α and peroxisome proliferator-activated receptor γ. J. Biol. Chem. 2007, 282, 14515–14524. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Gao, Y.; Zhang, Z.; Cao, Q.; Zhang, X.; Zou, J.; Cao, Y. Kdm2a/b lysine demethylases regulate canonical Wnt signaling by modulating the stability of nuclear β-catenin. Dev. Cell 2015, 33, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Yamaza, T.; Lee, J.; Yu, J.; Wang, S.; Fan, G.; Shi, S.; Wang, C.-Y. BCOR regulates mesenchymal stem cell function by epigenetic mechanisms. Nat. Cell Biol. 2009, 11, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ma, Y.; Ma, P.; Wang, S.; Fan, Z. Demethylation of epiregulin gene by histone demethylase FBXL11 and BCL6 corepressor inhibits osteo/dentinogenic differentiation. Stem Cells 2013, 31, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, A.; Tsukada, S.; Kamiyama, M.; Yanagimoto, T.; Nakajima, M.; Maeda, S. Wnt5b partially inhibits canonical Wnt/beta-catenin signaling pathway and promotes adipogenesis in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 2005, 330, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, M.; Koyanagi, A.; Osada, S.; Imagawa, M. Wnt4 and Wnt5a promote adipocyte differentiation. FEBS Lett. 2008, 582, 3201–3205. [Google Scholar] [CrossRef] [PubMed]
- Van Tienen, F.H.J.; Laeremans, H.; Kallen, C.J.H.; Smeets, H.J.M. Wnt5b stimulates adipogenesis by activating PPARγ, and inhibiting the β-catenin dependent Wnt signaling pathway together with Wnt5a. Biochem. Biophys. Res. Commun. 2009, 387, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, R.; Grünhagen, J.; Becker, J.; Robinson, P.; Ott, C.-E.; Knaus, P. miR-181a promotes osteoblastic differentiation through repression of TGF-β signaling molecules. Int. J. Biochem. Cell Biol. 2013, 45, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cai, J.; Cai, X.; Chen, L. MiR-346 regulates osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by targeting the Wnt/β-catenin pathway. PLoS ONE 2013, 8, e72266. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.-C.; Kim, T.; Kim, H.; Lee, S.-H.; Choi, Y. Transmembrane protein 64 reciprocally regulates osteoblast and adipocyte differentiation by modulating Wnt/β-catenin signaling. Bone 2015, 78, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Chen, Y.; Guo, L.; Jiang, T.; Lin, Z. miR-23a/b regulates the balance between osteoblast and adipocyte differentiation in bone marrow mesenchymal stem cells. Bone Res 2016, 4, 16022. [Google Scholar] [CrossRef] [PubMed]
- Lecka-Czernik, B.; Gubrij, I.; Moerman, E.J.; Kajkenova, O.; Lipschitz, D.A.; Manolagas, S.C.; Jilka, R.L. Inhibition of Osf2/Cbfa1 expression and terminal osteoblast differentiation by PPARγ2. J. Cell. Biochem. 1999, 74, 357–371. [Google Scholar] [CrossRef]
- Jeon, M.J.; Kim, J.A.; Kwon, S.H.; Kim, S.W.; Park, K.S.; Park, S.W.; Kim, S.Y.; Shin, C.S. Activation of peroxisome proliferator-activated receptor-γ inhibits the Runx2-mediated transcription of osteocalcin in osteoblasts. J. Biol. Chem. 2003, 278, 23270–23277. [Google Scholar] [CrossRef] [PubMed]
- Akune, T.; Ohba, S.; Kamekura, S.; Yamaguchi, M.; Chung, U.; Kubota, N.; Terauchi, Y.; Harada, Y.; Azuma, Y.; Nakamura, K.; et al. PPARγ insufficiency enhances osteogenesis through osteoblast formation from bone marrow progenitors. J. Clin. Investig. 2004, 113, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, M.; Lin, L.; Chen, P.; Ma, K.; Zhou, C.; Ao, Y. Runx2 overexpression enhances osteoblastic differentiation and mineralization in adipose-derived stem cells in vitro and in vivo. Calc. Tissue Int. 2006, 79, 169–178. [Google Scholar] [CrossRef] [PubMed]
- McGee-Lawrence, M.; Westendorf, J. Histone deacetylases in skeletal development and bone mass maintenance. Gene 2011, 474, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.-J.; Lee, K.-Y.; Choi, N.-S.; Lee, M.-H.; Kim, H.-N.; Jin, Y.-H.; Ryoo, H.-M.; Choi, J.-Y.; Yoshida, M.; Nishino, N.; et al. Bone morphogenetic protein-2 stimulates Runx2 acetylation. J. Biol. Chem. 2006, 281, 16502–16511. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, T.; Kahler, R.; Li, X.; Westendorf, J. Histone deacetylase 3 interacts with Runx2 to repress the osteocalcin promoter and regulate osteoblast differentiation. J. Biol. Chem. 2004, 279, 41998–42007. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.; Schroeder, T.; Bailey, J.; Gopalakrishnan, R.; Westendorf, J. Histone deacetylase 7 associates with Runx2 and represses its activity during osteoblast maturation in a deacetylation-independent manner. J. Bone Miner. Res. 2008, 23, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, P.; Ge, J.; Cheng, J.; Dong, W.; Yuan, H.; Du, Y.; Yang, M.; Sun, R.; Jiang, H. Histone deacetylase 8 suppresses osteogenic differentiation of bone marrow stromal cells by inhibiting histone H3K9 acetylation and RUNX2 activity. Int. J. Biochem. Cell Biol. 2014, 54, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Razidlo, D.; Whitney, T.; Casper, M.; McGee-Lawrence, M.; Stensgard, B.; Li, X.; Secreto, F.; Knutson, S.; Hiebert, S.; Westendorf, J. Histone deacetylase 3 depletion in osteo/chondroprogenitor cells decreases bone density and increases marrow fat. PLoS ONE 2010, 5, e11492. [Google Scholar] [CrossRef] [PubMed]
- Haberland, M.; Mokalled, M.; Montgomery, R.; Olson, E. Epigenetic control of skull morphogenesis by histone deacetylase 8. Genes Dev. 2009, 23, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Yoo, E.; Chung, J.-J.; Choe, S.; Kim, K.; Kim, J. Down-regulation of histone deacetylases stimulates adipocyte differentiation. J. Biol. Chem. 2006, 281, 6608–6615. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, T.; Idelman, G.; Blanco, V.; Blomkalns, A.; Piegore, M.; Weintraub, D.; Kumar, S.; Rajsheker, S.; Manka, D.; Rudich, S.; et al. Histone deacetylase 9 is a negative regulator of adipogenic differentiation. J. Biol. Chem. 2011, 286, 27836–27847. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Fan, Z.; Yu, B.; Chang, J.; Hezaimi, K.; Zhou, X.; Park, N.-H.; Wang, C.-Y. Histone demethylases KDM4B and KDM6B promotes osteogenic differentiation of human MSCs. Cell Stem Cell 2012, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Okamura, H.; Nakashima, Y.; Haneji, T. Histone demethylase Jmjd3 regulates osteoblast differentiation via transcription factors Runx2 and Osterix. J. Biol. Chem. 2013, 288, 33530–33541. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Guo, F.; Wang, G.; Wang, J.; Zheng, F.; Guan, X.; Chang, A.; Zhang, X.; Dai, C.; Li, S.; et al. MiR-20a regulates adipocyte differentiation by targeting lysine-specific demethylase 6b and transforming growth factor-β signaling. Int. J. Obes. 2015, 39, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jin, Q.; Lee, J.-E.; Su, I.; Ge, K. Histone H3K27 methyltransferase Ezh2 represses Wnt genes to facilitate adipogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 7317–7322. [Google Scholar] [CrossRef] [PubMed]
- Dudakovic, A.; Camilleri, E.; Xu, F.; Riester, S.; McGee-Lawrence, M.; Bradley, E.; Paradise, C.; Lewallen, E.; Thaler, R.; Deyle, D.; et al. Epigenetic control of skeletal development by the histone methyltransferase Ezh2. J. Biol. Chem. 2015, 290, 27604–27617. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Chen, Q.-M.; Hong, C.; Wang, C.-Y. Histone methyltransferases and demethylases: Regulators in balancing osteogenic and adipogenic differentiation of mesenchymal stem cells. Int. J. Oral Sci. 2015, 7, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, A.; Dell’Aversana, C.; Bugge, A.; Sarno, R.; Valente, S.; Rotili, D.; Manzo, F.; Teti, D.; Mandrup, S.; Ciana, P.; et al. HDACs class II-selective inhibition alters nuclear receptor-dependent differentiation. J. Mol. Endocrinol. 2010, 45, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Kallen, A.N.; Zhou, X.-B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.-S.; Zhang, H.; et al. The Imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.-C.; Fu, W.-M.; Wong, C.-W.; Wang, Y.; Wang, W.-M.; Hu, G.-X.; Zhang, L.; Xiao, L.-J.; Wan, D.; Zhang, J.-F.; et al. The lncRNA H19 promotes epithelial to mesenchymal transition by functioning as miRNA sponges in colorectal cancer. Oncotarget 2015, 6, 22513–22525. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, A.; Shi, Z.; Ma, F.; Pu, P.; Wang, T.; Zhang, J.; Kang, C.; Zhang, Q. MicroRNA-200a suppresses the WNT/β-catenin signaling pathway by interacting with β-catenin. Int. J. Oncol. 2012, 40, 1162–1170. [Google Scholar] [PubMed]
- Lee, H.; Woo, K.; Ryoo, H.; Baek, J. Distal-less homeobox 5 inhibits adipogenic differentiation through the down-regulation of peroxisome proliferator-activated receptor γ expression. J. Cell. Physiol. 2013, 228, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Nozawa, Y.; Akao, Y. MicroRNA-141 and -200a are involved in bone morphogenetic protein-2-induced mouse pre-osteoblast differentiation by targeting distal-less homeobox 5. J. Biol. Chem. 2009, 284, 19272–19279. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Fu, M.; Bookout, A.; Kliewer, S.; Mangelsdorf, D. MicroRNA let-7 regulates 3T3-L1 adipogenesis. Mol. Endocrinol. 2009, 23, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, X.; Cai, H.; Sun, Y.; Plath, M.; Li, C.; Lan, X.; Lei, C.; Lin, F.; Bai, Y.; et al. Long non-coding RNA ADNCR suppresses adipogenic differentiation by targeting miR-204. Biochim. Biophys. Acta 2016, 1859, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, L.; Xing, L.; Chen, D. MicroRNA-204 regulates Runx2 protein expression and mesenchymal progenitor cell differentiation. Stem Cells 2010, 28, 357–364. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Chen, K.; Wang, F.; Zhao, L.; Wan, X.; Wang, L.; Mo, Z. miR-204-5p promotes the adipogenic differentiation of human adipose-derived mesenchymal stem cells by modulating DVL3 expression and suppressing Wnt/β-catenin signaling. Int. J. Mol. Med. 2015, 35, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-X.; Yan, Y.-W.; Chen, D.; Ai, C.-Z.; Lu, X.; Xu, S.-S.; Jiang, S.; Zhong, G.-S.; Chen, D.-B.; Jiang, Y.-Z. Long non-coding RNA HoxA-AS3 interacts with EZH2 to regulate lineage commitment of mesenchymal stem cells. Oncotarget 2016, 7, 63561–63570. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Ge, X.; Yang, S.; Huang, M.; Zhuang, W.; Chen, P.; Zhang, X.; Fu, J.; Qu, J.; Li, B. Upregulation of lncRNA MEG3 promotes osteogenic differentiation of mesenchymal stem cells from multiple myeloma patients by targeting BMP4 transcription. Stem Cells 2015, 33, 1985–1997. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; O’Sullivan, M.; Green, A.; Watabe, Y.; Arisaka, O.; Masawa, N.; Matsuoka, K.; Fukami, M.; Matsubara, K.; Kato, F.; et al. The IG-DMR and the MEG3-DMR at human chromosome 14q32.2: Hierarchical interaction and distinct functional properties as imprinting control centers. PLoS Genet. 2010, 6, e1000992. [Google Scholar] [CrossRef] [PubMed]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.; Patel, T. microRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, L.; Lu, L.; Jiang, P.; Sun, H.; Wang, H. Inhibition of miR-29 by TGF-β-Smad3 signaling through dual mechanisms promotes transdifferentiation of mouse myoblasts into myofibroblasts. PLoS ONE 2012, 7, e33766. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Nabinger, S.; Vega, Z.; Sahu, S.; Alluri, R.; Abdul-Sater, Z.; Yu, Z.; Gore, J.; Nalepa, G.; Saxena, R.; et al. Pathophysiological role of microRNA-29 in pancreatic cancer stroma. Sci. Rep. 2015, 5, 11450. [Google Scholar] [CrossRef] [PubMed]
- Kapinas, K.; Kessler, C.; Ricks, T.; Gronowicz, G.; Delany, A. MiR-29 modulates Wnt signaling in human osteoblasts through a positive feedback loop. J. Biol. Chem. 2010, 285, 25221–25231. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hassan, M.; Jafferji, M.; Aqeilan, R.; Garzon, R.; Croce, C.; Wijnen, A.; Stein, J.; Stein, G.; Lian, J. Biological functions of miR-29b contribute to positive regulation of osteoblast differentiation. J. Biol. Chem. 2009, 284, 15676–15684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.; Qian, S.; Guo, L.; Huang, H.; He, Q.; Liu, Y.; Ma, C.; Tang, Q.-Q. Down-regulation of type I Runx2 mediated by dexamethasone is required for 3T3-L1 adipogenesis. Mol. Endocrinol. 2012, 26, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Bäckesjö, C.; Li, Y.; Lindgren, U.; Haldosén, L. Activation of Sirt1 decreases adipocyte formation during osteoblast differentiation of mesenchymal stem cells. J. Bone Miner. Res. 2006, 21, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Merrell, A.J.; Stanger, B.Z. Adult cell plasticity in vivo: De-differentiation and transdifferentiation are back in style. Nat. Rev. Mol. Cell Biol. 2016, 17, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Geurtzen, K.; Knopf, F.; Wehner, D.; Huitema, L.F.; Schulte-Merker, S.; Weidinger, G. Mature osteoblasts dedifferentiate in response to traumatic bone injury in the zebrafish fin and skull. Development 2014, 141, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Spencer, J.A.; Koh, B.; Kobayashi, T.; Fujisaki, J.; Clemens, T.L.; Lin, C.P.; Kronenberg, H.M.; Scadden, D.T. Endogenous bone marrow MSCs are dynamic, fate-restricted participants in bone maintenance and regeneration. Cell Stem Cell 2012, 10, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Malaval, L.; Aubin, J. The mature osteoblast phenotype is characterized by extensive plasticity. Exp. Cell Res. 1997, 232, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Schiller, P.C.; D’Ippolito, G.; Brambilla, R.; Roos, B.A.; Howard, G.A. Inhibition of gap-junctional communication induces the trans-differentiation of osteoblasts to an adipocytic phenotype in vitro. J. Biol. Chem. 2001, 276, 14133–14138. [Google Scholar] [CrossRef] [PubMed]
- Yoshiko, Y.; Oizumi, K.; Hasegawa, T.; Minamizaki, T.; Tanne, K.; Maeda, N.; Aubin, J.E. A subset of osteoblasts expressing high endogenous levels of PPARγ switches fate to adipocytes in the rat calvaria cell culture model. PLoS ONE 2010, 5, e11782. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Liu, M.; Ono, N.; Bringhurst, R.; Kronenberg, H.; Guo, J. Loss of Wnt/β-catenin signaling causes cell fate shift of preosteoblasts from osteoblasts to adipocytes. J. Bone Miner. Res. 2012, 27, 2344–2358. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Seong, S.; Kim, K.; Kim, I.; Jeong, B.-C.C.; Kim, N. Downregulation of Runx2 by 1,25-dihydroxyvitamin D3 induces the transdifferentiation of osteoblasts to adipocytes. Int. J. Mol. Sci. 2016, 17, 770. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.B.; Benkusky, N.A.; Sen, B.; Rubin, J.; Pike, J.W. Epigenetic plasticity drives adipogenic and osteogenic differentiation of marrow-derived mesenchymal stem cells. J. Biol. Chem. 2016, 291, 17829–17847. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Link, D. Targeting of mesenchymal stromal cells by cre-recombinase transgenes commonly used to target osteoblast lineage cells. J. Bone Miner. Res. 2016, 31, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, Y.; Sugiyama, T.; Kohara, H.; Kondoh, G.; Fujii, N.; Kohno, K.; Nagasawa, T. The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity 2010, 33, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Perino, M.; Veenstra, G.J.C. Chromatin control of developmental dynamics and plasticity. Dev. Cell 2016, 38, 610–620. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.E.; Mikkelsen, T.S.; Xie, X.; Kamal, M.; Huebert, D.J.; Cuff, J.; Fry, B.; Meissner, A.; Wernig, M.; Plath, K. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 2006, 125, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, T.; Ku, M.; Jaffe, D.; Issac, B.; Lieberman, E.; Giannoukos, G.; Alvarez, P.; Brockman, W.; Kim, T.-K.; Koche, R.; et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 2007, 448, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Rugg-Gunn, P.J.; Cox, B.J.; Ralston, A.; Rossant, J. Distinct histone modifications in stem cell lines and tissue lineages from the early mouse embryo. Proc. Natl. Acad. Sci. USA 2010, 107, 10783–10790. [Google Scholar] [CrossRef] [PubMed]
- Voigt, P.; Tee, W.-W.; Reinberg, D. A double take on bivalent promoters. Genes Dev. 2013, 27, 1318–1338. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Gordon, J.; Whitfield, T.; Tai, P.; Wijnen, A.; Stein, J.; Stein, G.; Lian, J. Chromatin dynamics regulate mesenchymal stem cell lineage specification and differentiation to osteogenesis. Biochim. Biophys. Acta 2017, 1860, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Hemming, S.; Cakouros, D.; Isenmann, S.; Cooper, L.; Menicanin, D.; Zannettino, A.; Gronthos, S. EZH2 and KDM6A act as an epigenetic switch to regulate mesenchymal stem cell lineage specification. Stem Cells 2014, 32, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Aguilar, R.; Henriquez, B.; Lian, J.; Stein, J.; Stein, G.; Wijnen, A.; Zundert, B.; Allende, M.; Montecino, M. Epigenetic control of the bone-master Runx2 gene during osteoblast-lineage commitment by the histone demethylase JARID1B/KDM5B. J. Biol. Chem. 2015, 290, 28329–28342. [Google Scholar] [CrossRef] [PubMed]
- Santoro, S.; Dulac, C. Histone variants and cellular plasticity. Trends Genet. 2015, 31, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.L.; Jones, P.A. DNA methylation decreases in aging but not in immortal cells. Science 1983, 220, 1055–1057. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.L.; Smith, R.A.; Ma, S.; Cutler, R.G. Genomic 5-methyldeoxycytidine decreases with age. J. Biol. Chem. 1987, 262, 9948–9951. [Google Scholar] [PubMed]
- Fuke, C.; Shimabukuro, M.; Petronis, A.; Sugimoto, J.; Oda, T.; Miura, K.; Miyazaki, T.; Ogura, C.; Okazaki, Y.; Jinno, Y. Age related changes in 5-methylcytosine content in human peripheral leukocytes and placentas: An HPLC-based study. Ann. Hum. Genet. 2004, 68, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Maegawa, S.; Hinkal, G.; Kim, H.; Shen, L.; Zhang, L.; Zhang, J.; Zhang, N.; Liang, S.; Donehower, L.; Issa, J.-P. Widespread and tissue specific age-related DNA methylation changes in mice. Genome Res. 2010, 20, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Groen, T.; Kadish, I.; Li, Y.; Wang, D.; James, S.; Karpf, A.; Tollefsbol, T. Insufficient DNA methylation affects healthy aging and promotes age-related health problems. Clin. Epigenet. 2011, 2, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tyler, J. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed]
- Feser, J.; Tyler, J. Chromatin structure as a mediator of aging. FEBS Lett. 2011, 585, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-H.; Zhao, T.; Cui, K.; Hu, G.; Chen, Q.; Chen, W.; Wang, X.-W.; Soto-Gutierrez, A.; Zhao, K.; Deng, C.-X. Negative reciprocal regulation between Sirt1 and Per2 modulates the circadian clock and aging. Sci. Rep. 2016, 6, 28633. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Perrien, D.; Fleming, N.; Nyman, J.; Ono, K.; Connelly, L.; Moore, M.; Lwin, S.; Yull, F.; Mundy, G.; et al. Silent information regulator (Sir) T1 inhibits NF-κB signaling to maintain normal skeletal remodeling. J. Bone Miner. Res. 2013, 28, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-W.; Lee, S.; Seo, M.-S.; Park, S.-B.; Kurtz, A.; Kang, S.-K.; Kang, K.-S. Histone deacetylase controls adult stem cell aging by balancing the expression of polycomb genes and jumonji domain containing 3. Cell. Mol. Life Sci. 2010, 67, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Nicotera, P. MicroRNAs in age-related diseases. EMBO Mol. Med. 2013, 5, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, K.; Noh, J.; Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Long noncoding RNAs in diseases of aging. Biochim. Biophys. Acta 2016, 1859, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Issa, J.-P. Aging and epigenetic drift: A vicious cycle. J. Clin. Investig. 2014, 124, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Mosteiro, L.; Pantoja, C.; Alcazar, N.; Marión, R.; Chondronasiou, D.; Rovira, M.; Fernandez-Marcos, P.; Muñoz-Martin, M.; Blanco-Aparicio, C.; Pastor, J.; et al. Tissue damage and senescence provide critical signals for cellular reprogramming in vivo. Science 2016, 354, aaf4445. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Cheng, P.; Liang, M.-K.; Chen, Y.-S.; Lu, Q.; Wang, J.-Y.; Xia, Z.-Y.; Zhou, H.-D.; Cao, X.; Xie, H. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. J. Clin. Investig. 2015, 125, 1509–1522. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tsang, K.; Tang, H.; Chan, D.; Cheah, K. Hypertrophic chondrocytes can become osteoblasts and osteocytes in endochondral bone formation. Proc. Natl. Acad. Sci. USA 2014, 111, 12097–12102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; von der Mark, K.; Henry, S.; Norton, W.; Adams, H.; de Crombrugghe, B. Chondrocytes transdifferentiate into osteoblasts in endochondral bone during development, postnatal growth and fracture healing in mice. PLoS Genet. 2014, 10, e1004820. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Zhang, W.; Zhang, W.; Liu, G.-H.; Belmonte, J. Switching cell fate, ncRNAs coming to play. Cell Death Dis. 2013, 4, e464. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.; DeWitt, N. In vivo cellular reprogramming: The next generation. Cell 2016, 166, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Long non-protein-coding RNAs (LncRNAs) play multiple roles in gene expression.
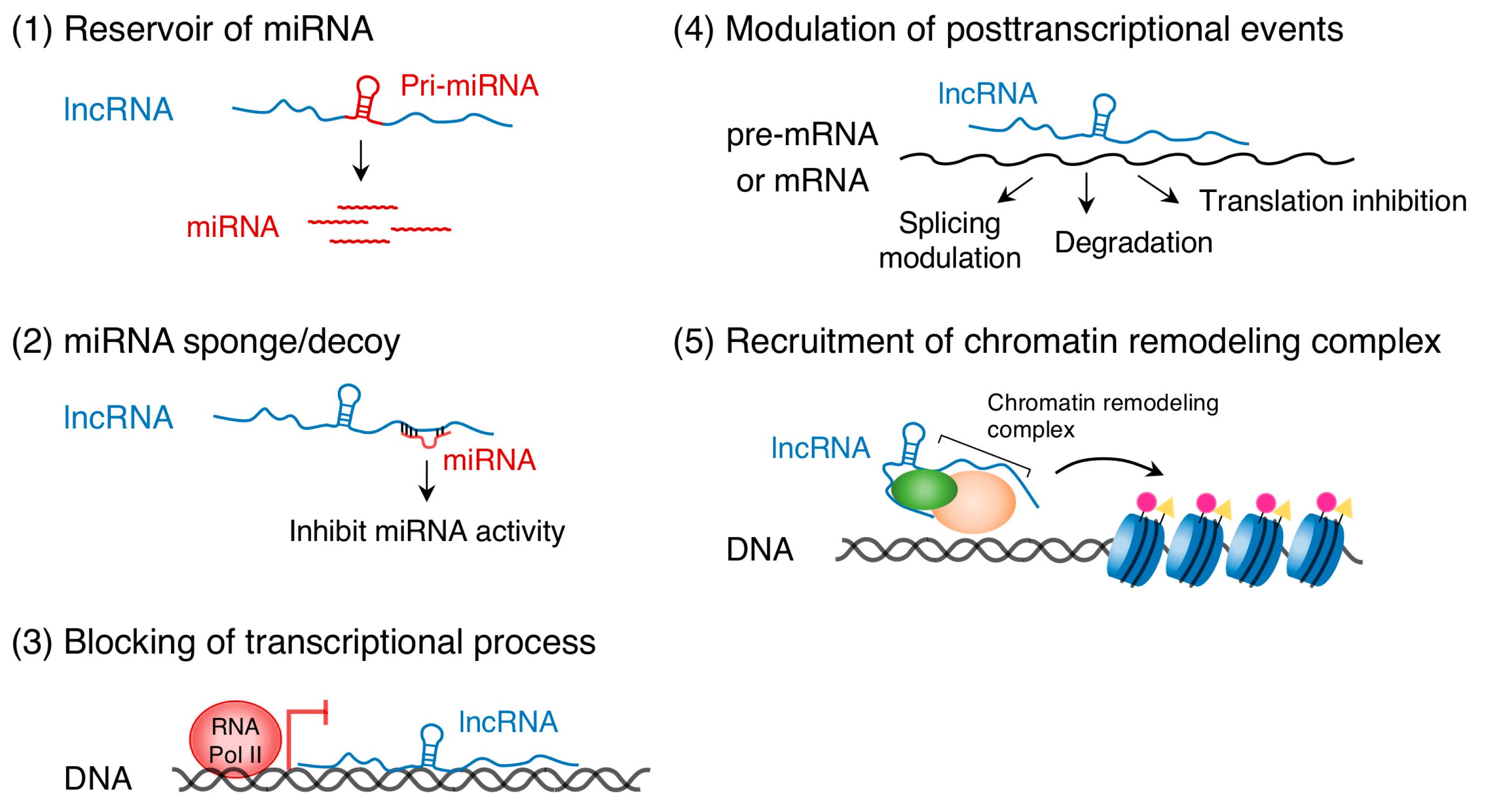

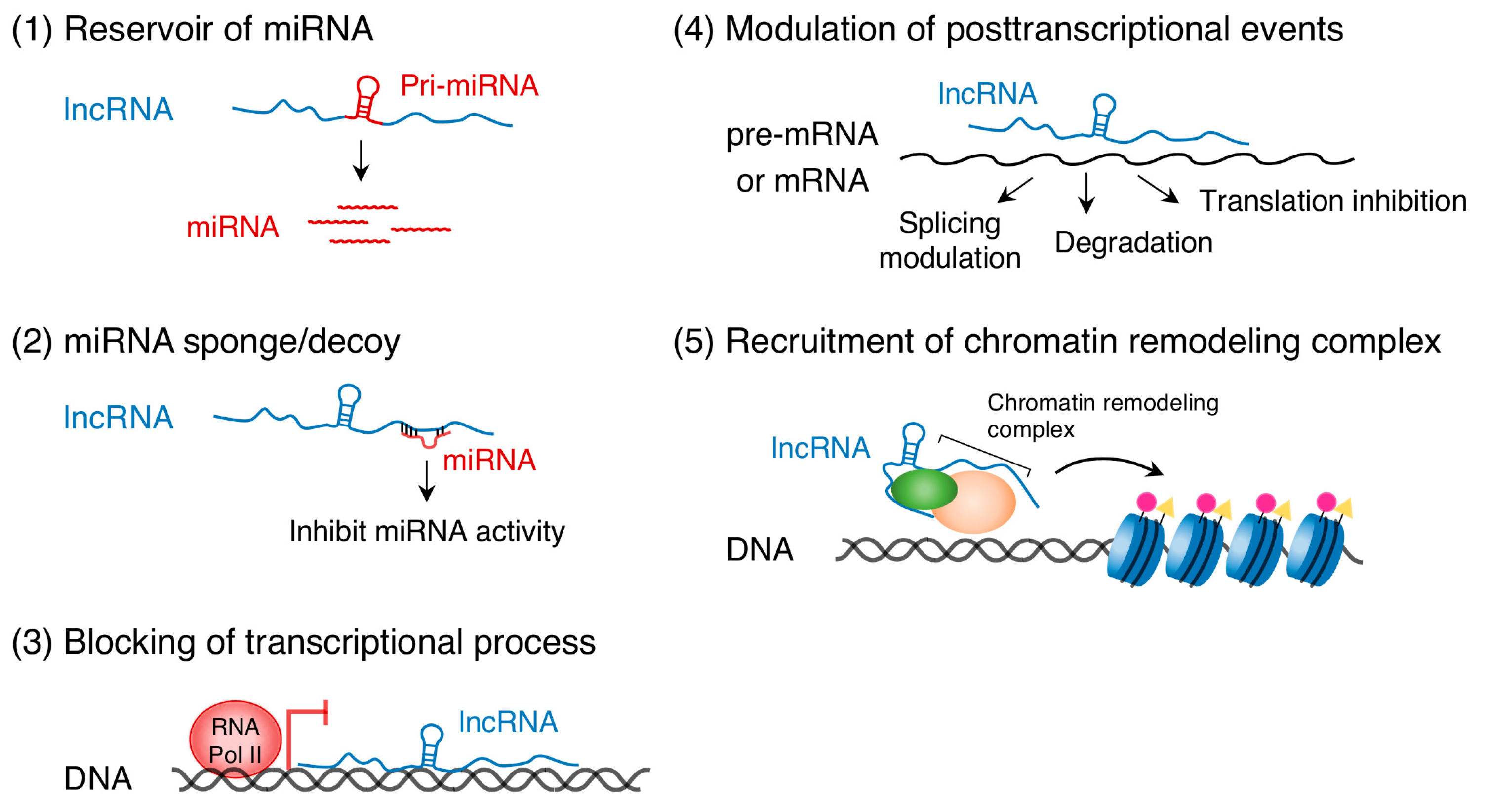{kind=link}
Table 1.
LncRNAs and osteo-adipogenic fate decision.
| LncRNAs | Expression Profiles | Functions | Effector Molecules | Target Genes | References |
|---|---|---|---|---|---|
| H19 | Osteogenic ↑ | miRNA precursor | miR-675 | TGFB1, HDAC4, HDAC5, HAC6 | [24,25] |
| Adipogenic ↓ | miRNA sponge | miR-200a, miR-141, miR-22, let-7 | DLX5, CTNNB1 | [26,83,84,85,87] | |
| MEG3 | Osteogenic ↑ | protein recruiter | PRC2 | TGFB2, TGFBR1, SMAD2 | [30] |
| Adipogenic ↓ | |||||
| ADNCR | Osteogenic ? | miRNA sponge | miR-204 | RUNX2, SIRT1, DVL3 | [89,90,91] |
| Adipogenic ↓ | |||||
| HOXA-AS3 | Osteogenic → | protein recruiter | EZH2 | RUNX2 | [92] |
| Adipogenic ↑ |
↑, upregulation; ↓, downregulation; →, no change in expression; ?, not determined.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yoshioka, H.; Yoshiko, Y. The Roles of Long Non-Protein-Coding RNAs in Osteo-Adipogenic Lineage Commitment. Int. J. Mol. Sci. 2017, 18, 1236. https://doi.org/10.3390/ijms18061236
AMA Style
Yoshioka H, Yoshiko Y. The Roles of Long Non-Protein-Coding RNAs in Osteo-Adipogenic Lineage Commitment. International Journal of Molecular Sciences. 2017; 18(6):1236. https://doi.org/10.3390/ijms18061236
Chicago/Turabian StyleYoshioka, Hirotaka, and Yuji Yoshiko. 2017. "The Roles of Long Non-Protein-Coding RNAs in Osteo-Adipogenic Lineage Commitment" International Journal of Molecular Sciences 18, no. 6: 1236. https://doi.org/10.3390/ijms18061236
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.