
Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences

Abstract
:1. Introduction
2. Transcriptomics and the NF-κB Pathway
3. Effects of Microgravity on NF-κB in the Immune System
4. Effects of Microgravity on NF-κB in Muscle
5. Effects of Microgravity on NF-κB in Bone
6. Effects of Microgravity on NF-κB in Cardiovascular System
7. Effects of Microgravity on NF-κB in Brain
8. Effects of Space Radiation Exposure on NF-κB
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Cucinotta, F.A.; Durante, M. Cancer risk from exposure to galactic cosmic rays: Implications for space exploration by human beings. Lancet Oncol. 2006, 7, 431–435. [Google Scholar] [CrossRef]
- National Council on Radiation Protection and Measurements. Guidance on Radiation Received in Space Activities; Report 98; National Council on Radiation Protection and Measurements: Bethesda, MD, USA, 1989. [Google Scholar]
- Hughes-Fulford, M.; Tjandrawinata, R.; Fitzgerald, J.; Gasuad, K.; Gilbertson, V. Effects of microgravity on osteoblast growth. Gravit. Space Biol. Bull. 1998, 11, 51–60. [Google Scholar] [PubMed]
- Sibonga, J.D.; Spector, E.R.; Johnston, S.L.; Tarver, W.J. Evaluating Bone Loss in ISS Astronauts. Aerosp. Med. Hum. Perform. 2015, 86 (Suppl. S12), A38–A44. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G.; Butel, J.S.; Shearer, W.T. Effects of the space flight environment on the immune system. Rev. Environ. Health 2003, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. npj Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef]
- Peterson, L.E.; Pepper, L.J.; Hamm, P.B.; Gilbert, S.L. Longitudinal study of astronaut health: Mortality in the years 1959–1991. Radiat. Res. 1993, 133, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.A.; Goodwin, T.J. Personalized medicine in human space flight: Using omics based analyses to develop individualized countermeasures that enhance astronaut safety and performance. Metabolomics 2013, 9, 1134–1156. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Hargens, A.R.; Vico, L. Long-duration bed rest as an analog to microgravity. J. Appl. Physiol. 2016, 120, 891–903. [Google Scholar] [CrossRef] [PubMed]
- La Tessa, C.; Sivertz, M.; Chiang, I.H.; Lowenstein, D.; Rusek, A. Overview of the NASA space radiation laboratory. Life Sci. Space Res. 2016, 11, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. NF-κB, the first quarter-century: Remarkable progress and outstanding questions. Genes Dev. 2012, 26, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Hellweg, C.E. The Nuclear factor κB pathway: A link to the immune system in the radiation response. Cancer Lett. 2015, 368, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Verma, I.M.; Stevenson, J.K.; Schwarz, E.M.; Van Antwerp, D.; Miyamoto, S. Rel/NF-κB/IκB family: Intimate tales of association and dissociation. Genes Dev. 1995, 9, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Grilli, M.; Chiu, J.J.; Lenardo, M.J. NF-κB and Rel: Participants in a multiform transcriptional regulatory system. Int. Rev. Cytol. 1993, 143, 1–62. [Google Scholar] [PubMed]
- Fullard, N.; Moles, A.; O’Reilly, S.; van Laar, J.M.; Faini, D.; Diboll, J.; Reynolds, N.J.; Mann, D.A.; Reichelt, J.; Oakley, F. The c-Rel subunit of NF-κB regulates epidermal homeostasis and promotes skin fibrosis in mice. Am. J. Pathol. 2013, 182, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Grimm, S.; Baeuerle, P.A. The inducible transcription factor NF-κB: Structure-function relationship of its protein subunits. Biochem. J. 1993, 290, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Aradhya, S.; Nelson, D.L. NF-κB signaling and human disease. Curr. Opin. Genet. Dev. 2001, 11, 300–306. [Google Scholar] [CrossRef]
- Franzoso, G.; Carlson, L.; Xing, L.; Poljak, L.; Shores, E.W.; Brown, K.D.; Leonardi, A.; Tran, T.; Boyce, B.F.; Siebenlist, U. Requirement for NF-κB in osteoclast and B-cell development. Genes Dev. 1997, 11, 3482–3496. [Google Scholar] [CrossRef] [PubMed]
- Pacios, S.; Xiao, W.; Mattos, M.; Lim, J.; Tarapore, R.S.; Alsadun, S.; Yu, B.; Wang, C.Y.; Graves, D.T. Osteoblast lineage cells play an essential role in periodontal bone loss through activation of nuclear factor-κB. Sci. Rep. 2015, 15, 16694. [Google Scholar] [CrossRef] [PubMed]
- Iotsova, V.; Caamaño, J.; Loy, J.; Yang, Y.; Lewin, A.; Bravo, R. Osteopetrosis in mice lacking NF-κB1 and NF-κB2. Nat. Med. 1997, 3, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Tranter, M.; Jones, W.K.; Sankovic, J.M.; Banerjee, R.K. Differential translocation of nuclear factor-κB in a cardiac muscle cell line under gravitational changes. J. Biomech. Eng. 2009, 131, 064503. [Google Scholar] [CrossRef] [PubMed]
- Mangala, L.S.; Zhang, Y.; He, Z.; Emami, K.; Ramesh, G.T.; Story, M.; Rohde, L.H.; Wu, H. Effects of simulated microgravity on expression profile of microRNA in human lymphoblastoid cells. J. Biol. Chem. 2011, 286, 32483–32490. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Walther, I.; Li, C.F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-κB pathway and transcription of immediate early genes in T cell activation are inhibited by microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Boonyaratanakornkit, J.B.; Cogoli, A.; Li, C.F.; Schopper, T.; Pippia, P.; Galleri, G.; Meloni, M.A.; Hughes-Fulford, M. Key gravity-sensitive signaling pathways drive T cell activation. FASEB J. 2005, 19, 2020–2022. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.; Thiel, C.; Timm, J.; Schmidt, P.M.; Huber, K.; Tauber, S.; Hemmersbachm, R.; Seibt, D.; Kroll, H.; Grote, K.H.; et al. Microgravity-induced alterations in signal transduction in cells of the immune system. Acta Astronaut. 2010, 67, 1116–1125. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, T.; Wong, M.; Wang, X.; Stodieck, L.; Karouia, F.; Story, M.; Wu, H. Transient gene and microRNA expression profile changes of confluent human fibroblast cells in spaceflight. FASEB J. 2016, 30, 2211–2224. [Google Scholar] [CrossRef] [PubMed]
- Grosse, J.; Wehland, M.; Pietsch, J.; Schulz, H.; Saar, K.; Hübner, N.; Eilles, C.; Bauer, J.; Abou-El-Ardat, K.; Baatout, S.; et al. Gravity-sensitive signaling drives 3-dimensional formation of multicellular thyroid cancer spheroids. FASEB J. 2012, 26, 5124–5140. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Timmerman, K.L.; Markofski, M.M.; Walker, D.K.; Dickinson, J.M.; Jamaluddin, M.; Brasier, A.R.; Rasmussen, B.B.; Volpi, E. Short-term bed rest increases TLR4 and IL-6 expression in skeletal muscle of older adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R216–R223. [Google Scholar] [CrossRef] [PubMed]
- Zwart, S.R.; Pierson, D.; Mehta, S.; Gonda, S.; Smith, S.M. Capacity of omega-3 fatty acids or eicosapentaenoic acid to counteract weightlessness-induced bone loss by inhibiting NF-κB activation: From cells to bed rest to astronauts. J. Bone Miner. Res. 2010, 25, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.L.; Bandstra, E.R.; Harrison, B.C.; Thorng, S.; Stodieck, L.S.; Kostenuik, P.J.; Morony, S.; Lacey, D.L.; Hammond, T.G.; Leinwand, L.L.; et al. Effects of spaceflight on murine skeletal muscle gene expression. J. Appl. Physiol. (1985) 2009, 106, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Gridley, D.S.; Mao, X.W.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Moldovan, M.; Cunningham, C.E.; Jones, T.A.; Slater, J.M.; Pecaut, M.J. Changes in mouse thymus and spleen after return from the STS-135 mission in space. PLoS ONE 2013, 8, e75097. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.L.; Kandarian, S.C.; Jackman, R.W. Identification of genes that elicit disuse muscle atrophy via the transcription factors p50 and Bcl-3. PLoS ONE 2011, 6, e16171. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Aoki, K.; Masuda, W.; Alles, N.; Nagano, K.; Fukushima, H.; Osawa, K.; Yasuda, H.; Nakamura, I.; Mikuni-Takagaki, Y.; et al. Disruption of NF-κB1 prevents bone loss caused by mechanical unloading. J. Bone Miner. Res. 2013, 28, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Wise, K.C.; Manna, S.K.; Yamauchi, K.; Ramesh, V.; Wilson, B.L.; Thomas, R.L.; Sarkar, S.; Kulkarni, A.D.; Pellis, N.R.; Ramesh, G.T. Activation of nuclear transcription factor-κB in mouse brain induced by a simulated microgravity environment. In Vitro Cell. Dev. Biol. Anim. 2005, 41, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Hellweg, C.E.; Baumstark-Khan, C.; Schmitz, C.; Lau, P.; Meier, M.M.; Testard, I.; Berger, T.; Reitz, G. Activation of the nuclear factor κB pathway by heavy ion beams of different linear energy transfer. Int. J. Radiat. Biol. 2011, 87, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, M.; Aravindan, N.; Meltz, M.L.; Herman, T.S. Post-translational modification of I-κBα activates NF-κB in human monocytes exposed to 56Fe ions. Radiat. Environ. Biophys. 2002, 41, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Baumstark-Khan, C.; Hellweg, C.E.; Arenz, A.; Meier, M.M. Cellular monitoring of the nuclear factor κB pathway for assessment of space environmental radiation. Radiat. Res. 2005, 164, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Hellweg, C.E.; Baumstark-Khan, C.; Schmitz, C.; Lau, P.; Meier, M.M.; Testard, I.; Berger, T.; Reitz, G. Carbon-ion-induced activation of the NF-κB pathway. Radiat. Res. 2011, 175, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K.; Sarma, A.; Krishna, M.; Verma, N.C. Expression of NF-κB and ERK following heavy ion irradiation. J. Environ. Pathol. Toxicol. Oncol. 2004, 23, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Rithidech, K.N.; Reungpatthanaphong, P.; Honikel, L.; Rusek, A.; Simon, S.R. Dose-rate effects of protons on in vivo activation of nuclear factor-κB and cytokines in mouse bone marrow cells. Radiat. Environ. Biophys. 2010, 49, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, A.; Pecaut, M.J.; Slater, J.M.; Subramaniam, S.; Gridley, D.S. Low-dose γ-rays modify CD4+ T cell signalling response to simulated solar particle event protons in a mouse model. Int. J. Radiat. Biol. 2011, 87, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Tungjai, M.; Whorton, E.B.; Rithidech, K.N. Persistence of apoptosis and inflammatory responses in the heart and bone marrow of mice following whole-body exposure to 28Silicon (28Si) ions. Radiat. Environ. Biophys. 2013, 52, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Jangiam, W.; Tungjai, M.; Rithidech, K.N. Induction of chronic oxidative stress, chronic inflammation and aberrant patterns of DNA methylation in the liver of titanium-exposed CBA/CaJ mice. Int. J. Radiat. Biol. 2015, 91, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Crucian, B.; Huff, J.L.; Klein, S.L.; Morens, D.; Murasko, D.; Nickerson, C.A.; Sonnenfeld, G. Effects of sex and gender on adaptation to space: Immune system. J. Womens Health 2014, 23, 956–958. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G. Use of animal models for space flight physiology studies, with special focus on the immune system. Gravit. Space Biol. Bull. 2005, 18, 31–35. [Google Scholar] [PubMed]
- Belay, T.; Aviles, H.; Vance, M.; Fountain, K.; Sonnenfeld, G. Effects of the hindlimb-unloading model of spaceflight conditions on resistance of mice to infection with Klebsiella pneumoniae. J. Allergy Clin. Immunol. 2002, 110, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Bakos, A.; Varkonyi, A.; Minarovits, J.; Batkai, L. Effect of simulated microgravity on human lymphocytes. J. Gravit. Physiol. 2001, 8, 69–70. [Google Scholar]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Ott, C.M.; Pierson, D.L. Changes in monocyte functions of astronauts. Brain Behav. Immun. 2005, 19, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Meloni, M.A.; Galleri, G.; Pippia, P.; Cogoli-Greuter, M. Cytoskeleton changes and impaired motility of monocytes at modelled low gravity. Protoplasma 2006, 229, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Buravkova, L.B.; Rykova, M.P.; Grigorieva, V.; Antropova, E.N. Cell interactions in microgravity: Cytotoxic effects of natural killer cells in vitro. J. Gravit. Physiol. 2004, 11, 177–180. [Google Scholar]
- Hughes-Fulford, M.; Chang, T.T.; Martinez, E.M.; Li, C.F. Spaceflight alters expression of microRNA during T-cell activation. FASEB J. 2015, 29, 4893–4900. [Google Scholar] [CrossRef] [PubMed]
- Nogami, M.; Huang, J.T.; James, S.J.; Lubinski, J.M.; Nakamura, L.T.; Makinodan, T. Mice chronically exposed to low dose ionizing radiation possess splenocytes with elevated levels of HSP70 mRNA, HSC70 and HSP72 and with an increased capacity to proliferate. Int. J. Radiat. Biol. 1993, 63, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ni, H.; Li, M.; Sanzari, J.K.; Diffenderfer, E.S.; Lin, L.; Kennedy, A.R.; Weissman, D. Effect of solar particle event radiation and hindlimb suspension on gastrointestinal tract bacterial translocation and immune activation. PLoS ONE 2012, 7, e44329. [Google Scholar] [CrossRef] [PubMed]
- Tauber, S.; Hauschild, S.; Paulsen, K.; Gutewort, A.; Raig, C.; Hürlimann, E.; Biskup, J.; Philpot, C.; Lier, H.; Engelmann, F.; et al. Signal transduction in primary human T lymphocytes in altered gravity during parabolic flight and clinostat experiments. Cell. Physiol. Biochem. 2015, 35, 1034–1051. [Google Scholar] [CrossRef] [PubMed]
- Brungs, S.; Kolanus, W.; Hemmersbach, R. Syk phosphorylation—A gravisensitive step in macrophage signalling. Cell Commun. Signal. 2015, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Mourkioti, F.; Rosenthal, N. NF-κB signaling in skeletal muscle: Prospects for intervention in muscle diseases. J. Mol. Med. 2007, 86, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005, 19, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.D.; Khanna, S.; Kaminski, H.J.; Rao, J.S.; Merriam, A.P.; Richmonds, C.R.; Leahy, P.; Li, J.; Guo, W.; Andrade, F.H. A chronic inflammatory response dominates the skeletal muscle molecular signature in dystrophin-deficient mdx mice. Hum. Mol. Genet. 2002, 11, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Bakkar, N.; Guttridge, D.C. NF-κB signaling: A tale of two pathways in skeletal myogenesis. Physiol. Rev. 2010, 90, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Na, S.Y.; Choi, J.E.; Kim, H.J.; Jhun, B.H.; Lee, Y.C.; Lee, J.W. Bcl3, an IκB protein, stimulates activating protein-1 transactivation and cellular proliferation. J. Biol. Chem. 1999, 274, 28491–28496. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B. Functions of NF-κB in Bone. Ann. N. Y. Acad. Sci. 2010, 1192, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Veis Novack, D. Role of NF-κB in the skeleton. Cell Res. 2011, 21, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Antonutto, G.; di Prampero, P.E. Cardiovascular deconditioning in microgravity: Some possible countermeasures. Eur. J. Appl. Physiol. 2003, 90, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Van der Heiden, K.; Cuhlmann, S.; Luong le, A.; Zakkar, M.; Evans, P.C. Role of nuclear factor κB in cardiovascular health and disease. Clin. Sci. 2010, 118, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.W.; Shaw, J.A.; Kirshenbaum, L.A. Multiple facets of NF-κB in the heart: To be or not to NF-κB. Circ. Res. 2011, 108, 1122–1132. [Google Scholar] [CrossRef] [PubMed]
- Boerma, M.; Nelson, G.A.; Sridharan, V.; Mao, X.W.; Koturbash, I.; Hauer-Jensen, M. Space radiation and cardiovascular disease risk. World J. Cardiol. 2015, 7, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Lawley, J.S.; Petersen, L.G.; Howden, E.J.; Sarma, S.; Cornwell, W.K.; Zhang, R.; Whitworth, L.A.; Williams, M.A.; Levine, B.D. Effect of gravity and microgravity on intracranial pressure. J. Physiol. 2017, 595, 2115–2127. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Sarkar, S.; Ramesh, V.; Kim, H.; Barnes, S.; Kulkarni, A.; Hall, J.C.; Wilson, B.L.; Thomas, R.L.; Pellis, N.R.; et al. Proteomic analysis of mouse hypothalamus under simulated microgravity. Neurochem. Res. 2008, 33, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Meffert, M.K. Roles for NF-κB in nerve cell survival, plasticity, and disease. Cell Death Differ. 2006, 13, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, B.; Uherek, M.; Volk, B.; Baeuerle, P.A.; Kaltschmidt, C. Transcription factor NF-κB is activated in primary neurons by amyloid β peptides and in neurons surrounding early plaques from patients with Alzheimer disease. Proc. Natl. Acad. Sci. USA 1997, 94, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Baluchamy, S.; Ravichandran, P.; Ramesh, V.; He, Z.; Zhang, Y.; Hall, J.C.; Jejelowo, O.; Gridley, D.S.; Wu, H.; Ramesh, G.T. Reactive oxygen species mediated tissue damage in high energy proton irradiated mouse brain. Mol. Cell. Biochem. 2012, 360, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Bielinski, D.F.; Carrihill-Knoll, K.; Rabin, B.M.; Shukitt-Hale, B. Exposure to 16O-particle radiation causes aging-like decrements in rats through increased oxidative stress, inflammation and loss of autophagy. Radiat. Res. 2011, 176, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Fazio, G.G.; Jelley, J.V.; Charman, W.N. Generation of Cherenkov light flashes by cosmic radiation within the eyes of the Apollo astronauts. Nature 1970, 228, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, L.S.; Osborne, W.Z.; Hoffman, R.A.; Bailey, J.V. Light flashes observed by astronauts on skylab 4. Science 1975, 188, 928–930. [Google Scholar] [CrossRef] [PubMed]
- George, K.; Durante, M.; Wu, H.; Willingham, V.; Badhwar, G.; Cucinotta, F.A. Chromosome aberrations in the blood lymphocytes of astronauts after space flight. Radiat. Res. 2001, 156, 731–738. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Manuel, F.K.; Jones, J.; Iszard, G.; Murrey, J.; Djojonegro, B.; Wear, M. Space radiation and cataracts in astronauts. Radiat. Res. 2001, 156, 460–466. [Google Scholar] [CrossRef]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Liu, J.; Liu, X.; Zhao, S.; Li, H.; Tan, Y.; Xu, J. GABAB R/GSK-3β/NF-κB signaling pathway regulates the proliferation of colorectal cancer cells. Cancer Med. 2016, 5, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, G.V.; Bhanumurthy, L.; Balasubramanian, A.; Ramanathan, A. A novel mutation in the DNA binding domain of NFKB is associated with speckled leukoplakia. Asian Pac. J. Cancer Prev. 2016, 17, 3627–3629. [Google Scholar] [PubMed]
- Habraken, Y.; Piette, J. NF-κB activation by double-strand breaks. Biochem. Pharmacol. 2006, 72, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Matsumoto, H.; Funayama, T.; Yokota, Y.; Otsuka, K.; Maeda, M.; Kobayashi, Y. Nitric oxide-mediated bystander signal transduction induced by heavy-ion microbeam irradiation. Life Sci. Space Res. 2015, 6, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Ivanov, V.N.; Hong, M.; Lieberman, H.B.; Brenner, D.J.; Amundson, S.A.; Geard, C.R. Mechanism of radiation-induced bystander effects: A unifying model. J. Pharm. Pharmacol. 2008, 60, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.M.; Li, J.J. ATM-NF-κB connection as a target for tumor radiosensitization. Curr. Cancer Drug Targets 2007, 7, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.M.; Li, J.J. NF-κB-mediated adaptive resistance to ionizing radiation. Free Radic. Biol. Med. 2008, 44, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.S.; Sarkar, S.; Periyakaruppan, A.; Ravichandran, P.; Sadanandan, B.; Ramesh, V.; Thomas, R.; Hall, J.C.; Wilson, B.L.; Ramesh, G.T. Simulated microgravity activates apoptosis and NF-κB in mice testis. Mol. Cell. Biochem. 2008, 313, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Panday, A.; Inda, M.E.; Bagam, P.; Sahoo, M.K.; Osorio, D.; Batra, S. Transcription Factor NF-κB: An Update on Intervention Strategies. Arch. Immunol. Ther. Exp. 2016, 64, 463–483. [Google Scholar] [CrossRef] [PubMed]
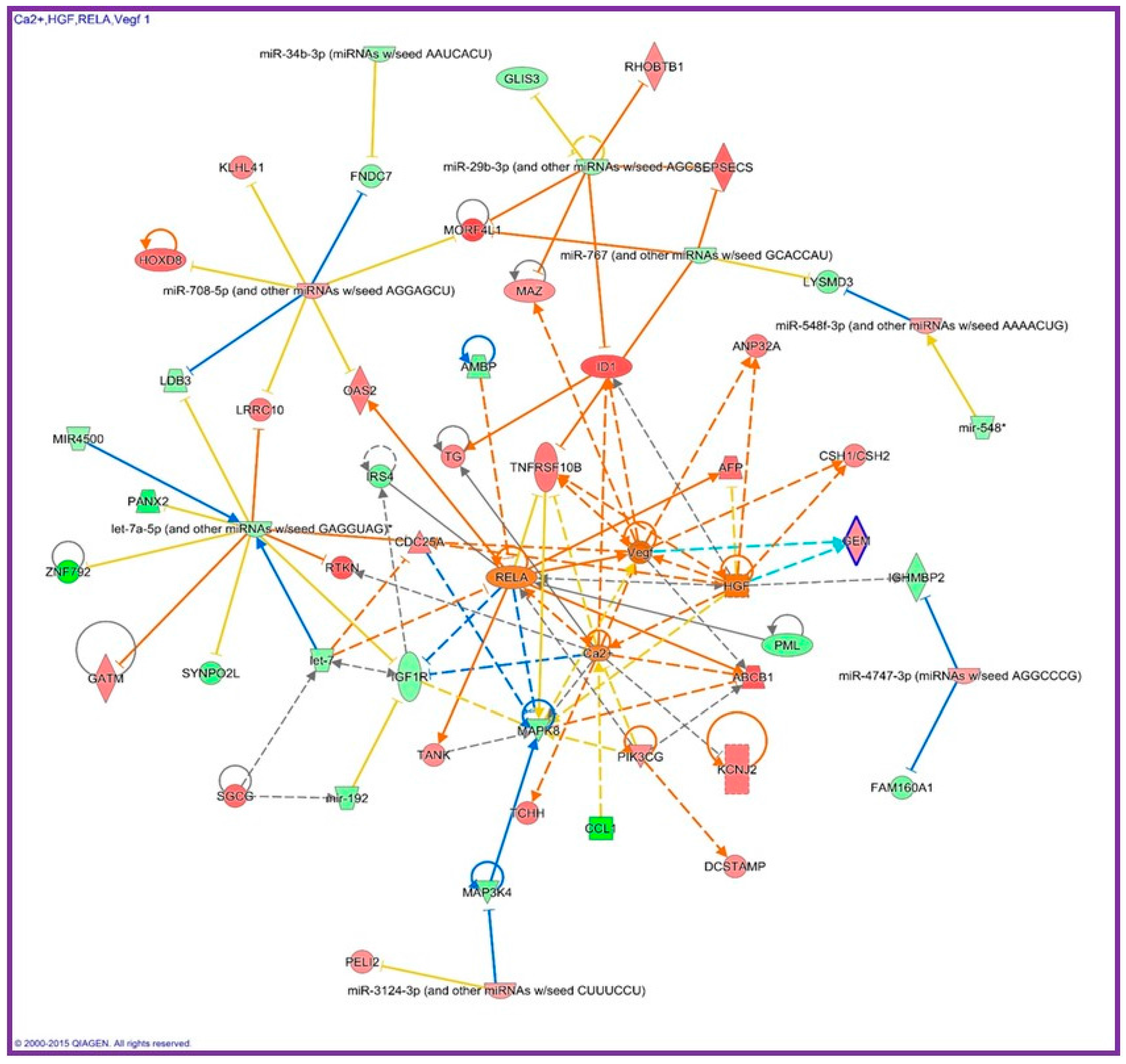

{kind=link}
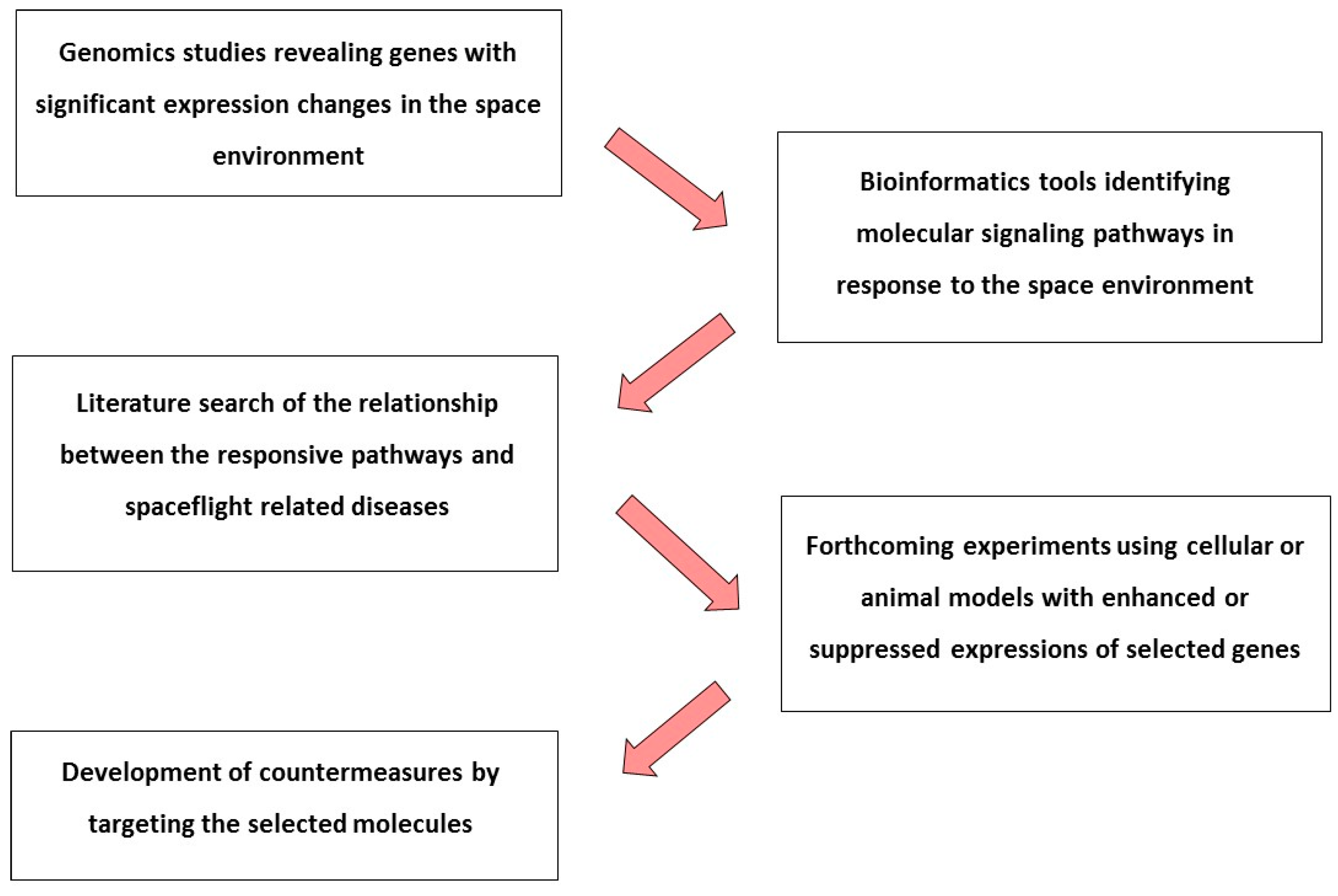
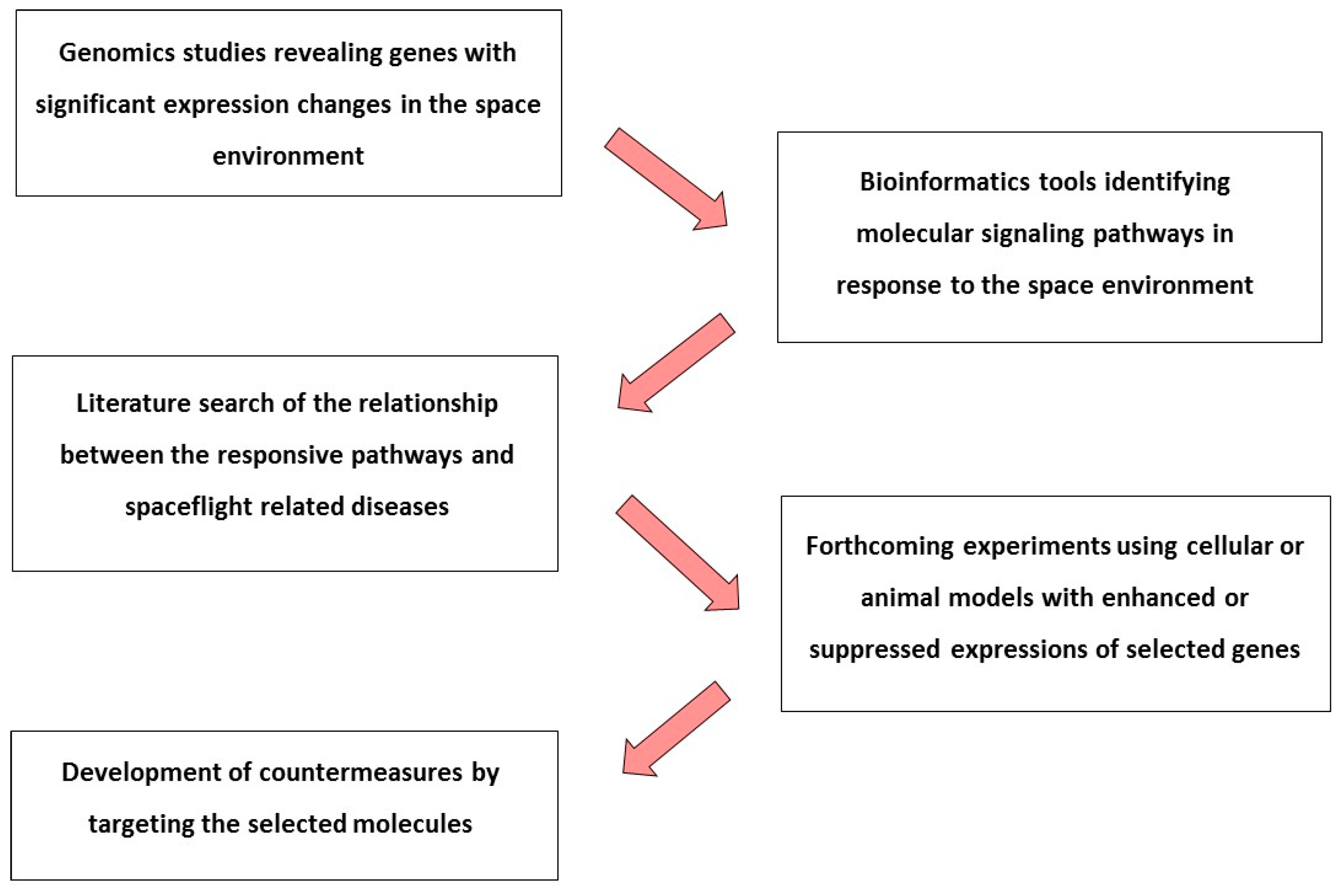{kind=link}
| Cells/Organism | Type of Microgravity | Type of Analysis | Length of Exposure | Result | Author |
|---|---|---|---|---|---|
| Cell Line | |||||
| Rat cardiac cells (H9c2) | RWV | Western blot; ELISA on nuclear lysates | 3 h | Increased NF-κB p65 DNA binding activity | Kwon et al. [25] |
| Human lymphoblastoid cells (TK6) | RWV | Microarray; PCR array on whole cell lysates | 72 h | Differential expression profile of genes and miRNAs identifying activation of the NF-κB pathway | Mangala et al. [26] |
| Activated human T cells | ISS | Microarray on whole cell lysates | 1.5 h | Suppressed expression of cREL/NF-κB gene targets | Chang et al. [27] |
| Activated human T cells | RPM | RT-PCR on whole cell lysates | 4 h | Suppressed expression of NF-κB gene targets | Boonyaratnakornkit et al. [28] |
| Human Jurkat T cells | RWV | Western blot on nuclear lysates | 5 min | Decreased translocation of NF-κB p65 protein | Paulsen et al. [29] |
| Human fibroblasts (AG1522) | ISS | Microarray; PCR array on whole cell lysates | 72 h | Differential expression profile of genes and miRNAs identifying activation of the NF-κB pathway | Zhang et al. [30] |
| Human Thyroid cancer cells (FTC-133) | RPM | Microarray; Western blot on whole cell lysates | 24 h | Increased NF-κB p65 protein level | Grosse et al. [31] |
| Human Study/Mouse Cells | |||||
| Human muscle tissue | Bed-rest | RT-PCR; Western blot on tissue sample | 7 days | Increased expression of NFKB1, No change in total and phosphorylated NF-κB p65 protein level | Drummond et al. [32] |
| Human PBMCs; Murine monocyte/macrophage cells (RAW264.7) | RWV for RAW264.7 cells; Space Shuttle for PBMCs | Western blot and ELISA on nuclear lysates for RAW264.7 cells; Immunocytochemical method for PBMCs | 24 h for RAW264.7 cells; 12–16 days for PBMCs | Increased NF-κB p65 DNA binding activity and increased p65 protein level in RAW264.7 cells; Increased NF-κB p65 protein level after spaceflight; Omega-3 fatty acids or eicosapentaenoic acid reduced NF-κB p65 protein level | Zwart et al. [33] |
| Mouse Study | |||||
| Mouse Gastrocnemius (C57BL/6) | STS-108; HU | Microarray; RT-PCR on tissue sample | 11 days 19 h | Increased expression of Nfkbia/Iκbα | Allen et al. [34] |
| Moue spleen (C57BL/6) | STS-135 | PCR array on tissue sample | 13 days | Suppressed expression of Nfκb1 | Gridley et al. [35] |
| WT and Nfkb1−/− or Bcl-3−/− mouse gastrocnemius and plantaris muscles (B6129PF2/) | HU | Microarray; RT-PCR on tissue sample | 10 days | Reduced muscle atrophy in Nfkb1−/− or Bcl-3−/− mice; Increased expression of NF-κB gene targets in WT mice | Wu et al. [36] |
| WT and Nfkb1−/− mouse bone (C57BL/6) | HU | µCT on tibias and femurs bone mass; Western blot on tissue sample | 2 weeks | Reduced bone loss in Nfkb1−/− mice in comparison to WT, Increased NF-κB p50 protein level in WT HU group | Nakamura et al. [37] |
| Mouse brain (BALB/c) | HU | EMSA on nuclear lysates | 7 days | Increased NF-κB DNA binding activity | Wise et al. [38] |
| Cells/Organism | Radiation Type and Quality | Dose/Dose Rate Range | Method | Result | Reference |
|---|---|---|---|---|---|
| Cell Line | |||||
| Human embryonic kidney cells (HEK 293) | C (34 keV/µm) Ne (91 keV/µm); Ar (272 keV/µm); Ni (906 keV/µm); Pb (9674 keV/µm) | 0.1–60 Gy ~1 Gy/min | Fluorescence d2EGFP reporter gene assay | RBE for NF-κB dependent d2EGFP expression varies by particle types with peak RBE = 8.9 | Hellweg et al. [39] |
| Human monocytes (MM6) | Fe | 0.2–1.4 Gy 1 Gy/min | EMSA on nuclear lysates | NF-κB DNA binding activity increased in a dose and time dependent manner | Natarajan et al. [40] |
| Human embryonic kidney cells (HEK 293) | Ar (230 keV/µm) | 0.2–30 Gy | Fluorescence d2EGFP reporter gene assay | Increased NF-κB dependent d2EGFP expression in a dose and time dependent manner | Baumstark-Khan et al. [41] |
| Human embryonic kidney cells (HEK 293) | C (33 and 73 keV/µm) | 0.2–20 Gy ~1 Gy/min | Fluorescence d2EGFP reporter gene assay | NF-κB dependent d2EGFP expression was comparable to X-rays | Hellweg et al. [42] |
| Chinese hamster cells (V79) | O | 1 Gy | Western blot on whole cell lysates | Decreased NF-κB p65 level at 30 min post irradiation, but the level recovered at longer time points | Mitra et al. [43] |
| Mouse Study | |||||
| Mouse bone marrow (BALB/cJ) | Proton (0.7 keV/µm) | 1 Gy 5 and 10 mGy/min | ELISA on nuclear lysates | NF-κB p65 DNA binding activity was both dose rate and time dependent | Rithidech et al. [44] |
| Mouse spleen T cells (C57BL/6) | SPE protons | 1.7 Gy delivered over 36 h | ELISA on whole cell lysates | Increased total and phosphorylated form of NF-κB p65 | Rizvi et. al. [45] |
| Mouse heart and bone marrow (CBA/CaJ) | Si (77 keV/µm) | 0.1–0.5 Gy 10 mGy/min in two fractionations with 15 days apart | ELISA on nuclear lysates | Increased NF-κB p65 DNA binding activity up to 6 months post irradiation | Tungjai et al. [46] |
| Mouse liver (CBA/CaJ) | Ti (107 keV/µm) | 0.1–0.5 Gy 0.01 Gy/min | ELISA on nuclear lysates | Increased level of NF-κB p65 DNA binding activity up to 6 months post irradiation | Jangiam et al. [47] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Moreno-Villanueva, M.; Krieger, S.; Ramesh, G.T.; Neelam, S.; Wu, H. Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences. Int. J. Mol. Sci. 2017, 18, 1166. https://doi.org/10.3390/ijms18061166
Zhang Y, Moreno-Villanueva M, Krieger S, Ramesh GT, Neelam S, Wu H. Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences. International Journal of Molecular Sciences. 2017; 18(6):1166. https://doi.org/10.3390/ijms18061166
Chicago/Turabian StyleZhang, Ye, Maria Moreno-Villanueva, Stephanie Krieger, Govindarajan T. Ramesh, Srujana Neelam, and Honglu Wu. 2017. "Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences" International Journal of Molecular Sciences 18, no. 6: 1166. https://doi.org/10.3390/ijms18061166