
NGF and Its Receptors in the Regulation of Inflammatory Response

1
Division of Rheumatology and Immuno-Rheumatology Research Laboratories, Bambino Gesù Children’s Hospital, 00146 Rome, Italy
2
Institute of Translational Pharmacology, Consiglio Nazionale delle Ricerche (CNR), 00133 Rome, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(5), 1028; https://doi.org/10.3390/ijms18051028
Submission received: 17 February 2017
/
Revised: 31 March 2017
/
Accepted: 3 May 2017
/
Published: 11 May 2017
(This article belongs to the Special Issue Neurotrophic Factors—Historical Perspective and New Directions)
Abstract
:There is growing interest in the complex relationship between the nervous and immune systems and how its alteration can affect homeostasis and result in the development of inflammatory diseases. A key mediator in cross-talk between the two systems is nerve growth factor (NGF), which can influence both neuronal cell function and immune cell activity. The up-regulation of NGF described in inflamed tissues of many diseases can regulate innervation and neuronal activity of peripheral neurons, inducing the release of immune-active neuropeptides and neurotransmitters, but can also directly influence innate and adaptive immune responses. Expression of the NGF receptors tropomyosin receptor kinase A (TrkA) and p75 neurotrophin receptor (p75NTR) is dynamically regulated in immune cells, suggesting a varying requirement for NGF depending on their state of differentiation and functional activity. NGF has a variety of effects that can be either pro-inflammatory or anti-inflammatory. This apparent contradiction can be explained by considering NGF as part of an endogenous mechanism that, while activating immune responses, also activates pathways necessary to dampen the inflammatory response and limit tissue damage. Decreases in TrkA expression, such as that recently demonstrated in immune cells of arthritis patients, might prevent the activation by NGF of regulatory feed-back mechanisms, thus contributing to the development and maintenance of chronic inflammation.
1. Introduction
The discovery of nerve growth factor (NGF) more than 60 years ago is associated with embryonal development and the differentiation of peripheral neuronal cells. Studies by Levi-Montalcini in chick embryos clearly demonstrated that the production of this diffusible factor in the target organ was responsible for the survival of peripheral neurons during development. The purification of NGF, its biochemical characterization and the production of neutralizing antibodies made it possible to demonstrate in a number of animal models, both in vitro and in vivo, that NGF is essential for the survival of sympathetic and sensory neurons during nervous system differentiation [1]. The release of NGF in the innervation field not only guarantees the survival of neurons [2] but also regulates axon growth and synapse formation [3], in addition to influencing neurotransmitter and neuropeptide synthesis [4,5,6,7]. The activity of NGF is not restricted to embryonal life and although sensory neurons do not continue to be dependent on NGF for their survival, as sympathetic neurons do, the density of innervation [8,9], the expression of ion channels [10,11,12] and the synthesis of neurotransmitters and neuropeptides [13,14] are dynamically regulated by NGF. During adult life there is a basal production of NGF in the innervation field that is essential in regulating neuronal plasticity. There is a close correlation between the constitutive synthesis of NGF and the innervation density of adult tissues. The local production of NGF regulates cell body size, axonal sprouting and dendritic arborization [15,16]. In agreement with this, transgenic mice overexpressing NGF in epithelial structures show changes in neuronal phenotype associated with a striking increase in the number of axons, an altered distribution and enhanced branching of fibers in target organs [17,18,19,20].
So why refer to inflammation and immune responses in a review dedicated to nerve growth factor? The production of NGF is tightly controlled in all regions of the organism innervated by sensory and sympathetic neurons (i.e., the mucosae, skin and derma, internal organs) because its alteration profoundly modifies the organism’s physiology. Nevertheless, a significant increase in NGF synthesis in inflamed tissues has been described in patients and animal models of inflammatory diseases [21,22,23,24,25,26]. One of the effects of this enhanced production of NGF is the inflammatory pain associated with the NGF-induced expression of transient receptor potential vanilloid 1 (TRPV1) channels [27] and sodium channels [12,28] and the alteration of peripheral innervation [29,30]. Changes in NGF synthesis have a profound effect on neuron physiology [31] but, as the present review will show, they may also influence immune cell activity. This dual action of NGF on nervous and immune cells should not be surprising. The nervous and immune systems have identical functions: they are both responsible for maintaining homeostasis and for adapting the body to the environment [32]. In order to orchestrate strictly integrated responses, they need to maintain close anatomical connections and to share common chemical signals and specific receptors [33,34]. This direct bidirectional communication pathway enables a functional interaction between the two systems. Considering the well-known effects of NGF on peripheral neuron survival and its dynamic control of innervation and neuropeptide synthesis, together with the fact that NGF receptors are expressed in immune cells, it is intuitive that NGF can influence the activity of both systems. Indeed, a growing body of data indicates that NGF is a key molecule in the complex network of bidirectional signals between the nervous and immune systems.
2. NGF Is Enhanced during the Inflammatory Response
Studies on inflammatory and autoimmune diseases, which are characterized by an abnormal activation of immune cells and increased production of cytokines, have revealed a localized increase in NGF at the sites of inflammation. Enhanced NGF levels were initially found in the cerebrospinal fluid of multiple sclerosis patients, and it was shown that an increase in NGF closely follows the course of the disease [21]. The synovial fluids of patients with rheumatoid arthritis [22,23] are also characterized by an increased concentration of NGF, and its up-regulation in inflamed synovia was confirmed in studies on different animal models of induced arthritis [35,36]. Systemic Lupus Erythematosus (SLE) patients exhibit a significant increase in NGF concentration in the sera [24,25], which correlates with disease activity, and similar findings were obtained in studies of NZB/W mice, a spontaneous model of human SLE [37]. The list of inflammatory diseases characterized by an enhanced production of NGF in the inflamed tissues or in the blood is fairly long and includes diseases with different pathogenic mechanisms [38,39,40,41,42,43,44,45].
Thus, the normally low basal production of NGF is enormously up-regulated during inflammatory response. It is reasonable to hypothesize that, during the inflammatory process and because of tissue damage, there is a release of mediators that modify the local concentration of NGF. A number of studies have shown that cytokines involved in inflammation, such as IL-1β, TNF-α and IL-6, are promoters of NGF synthesis in a variety of cell types [46,47,48,49,50,51,52]. Inflammatory cytokines can induce the synthesis of NGF in neuronal and glial cells, as well as in epithelial, endothelial, connective and muscle cells (Figure 1). There are probably also other molecules that can up-regulate the basal production of NGF in tissue. For example, NGF production can be induced by prostaglandins [53,54] and histamine [55,56] in cultures of adipocytes, astrocytes and keratinocytes. Although the local alteration in NGF production seems to be correlated with inflammatory mediators, there are not many data demonstrating in vivo the specific cell types and mediators responsible for the enhanced production of NGF during inflammatory response in patients and in animal models of inflammatory diseases [57,58,59,60].
3. Expression of NGF Receptors in the Immune System
The distribution of NGF receptors in different areas of lymphoid organs suggests that local NGF production and its accumulation in afferent lymphoid vessels not only regulate the innervation of peripheral neurons but can also have an immunomodulatory activity. The expression of NGF and of the Tropomyosin Receptor Kinase A (TrkA) seems to be relevant for the differentiation of lymphoid organs, since structural abnormalities have been described in NGF receptor-deficient mouse lymphoid organs [61,62]. These findings suggest that NGF can regulate the differentiation of immune cells. The discovery of patients with mutations in the gene encoding TrkA [63] that cause an autosomal sensory neuropathy defined as congenital insensitivity to pain with anhidrosis (CIPA) provides further evidence that NGF can be involved in the regulation of the immune response in vivo. CIPA patients are characterized by neurological alterations and the absence of pain and sensation, but they also show reduced wound healing and modified functions of immune cells. They have altered immune mechanisms that are responsible for recurrent infections and inflammatory complications [64,65] that lead to chronic inflammatory responses [66].
3.1. Expression of NGF Receptors in Primary and Secondary Lymphoid Organs
The biological effects of NGF are mediated by its binding to two classes of receptors: p75 Neurotrophin Receptor (p75NTR), a 75 kDa glycoprotein that is a member of the TNF-receptor superfamily [67], and TrkA, a transmembrane tyrosine kinase of 140 kDa that is phosphorylated on tyrosine residues after binding to NGF [68]. TrkA is considered the NGF-specific receptor for its greater affinity and specificity for NGF compared with p75NTR which, in contrast, can also bind other neurotrophins with affinity similar to NGF. The majority of studies that have characterized TrkA and p75NTR expression and signaling in physiological and pathological conditions focused on neuronal cells and the peripheral and central nervous systems during development and in adult life [1,3,31,69,70].
The expression of these receptors is not restricted to the nervous system, but a number of studies have shown NGF receptor distribution in primary lymphoid organs (bone marrow, thymus and bursa of Fabricius), where the differentiation of immune cells from multipotent stem cells occurs (Table 1).
The thymus is a primary lymphoid organ in which T-cells differentiate, and it is active throughout childhood until puberty [71]. The thymus is composed of different types of cells (thymic epithelial cells, macrophages and dendritic cells), and thymocytes at various stages of T-cell differentiation move from the cortical to the medullar area. Both mRNA and protein levels of p75NTR have been detected in the thymus of mammals [72,73,74] as well as the expression of TrkA, although different TrkA isoforms have been described in the thymus [75,76]. The expression of TrkA specifically characterizes rat and human thymocytes [77,78,79]. TrkA is also expressed in epithelial subcapsular and medullar cells [73,75,80,81], in peripheral epithelial cells of Hassal’s bodies and in interdigitating reticular cells of the medulla [79]. The latter cells also express p75NTR, which is also expressed in periarteriolar macrophages, endothelial sinusal cells and nerve endings [79,80,81,82]. TrkA expression seems to be necessary for the normal development of the thymus, as demonstrated in TrkA-deficient mice, in which the thymus is smaller, with no clear delimitation between the cortex and the medulla, and the thymocyte density is lower [61].
The expression of TrkA and p75NTR is modulated in the rat embryo with mRNA levels of TrkA decreasing and of p75NTR increasing in the stroma of the thymus [78] during rat development. In the final stages of embryo development and in the early post-natal days the expression of TrkA and p75NTR in the rat thymus is maximal, and then declines with age [81] in parallel with age-dependent changes in medullar epithelial cells [82]. Interestingly, in normal human thymuses the epithelial cells exhibit a TrkA positive-p75NTR negative phenotype but a switch to a TrkA negative-p75NTR positive phenotype has been described in malignant epithelial cell tumors [73,83].
In bone marrow, a heterogeneous population of stromal cells provides the correct microenvironment for hematopoietic stem cell survival and differentiation in myeloid and lymphoid lineages. In human and rat, stromal cells with dendritic features expressing both TrkA and p75NTR appear in the fetal bone marrow before the hematopoietic activity begins [77,84]. CD34 positive hemopoietic stem cells also express NGF, p75NTR and TrkA, and in vitro this expression is maintained even in the absence of inducing factors [85,86,87].
Similarly to what has been described in bone marrow, there is also a gradient of expression of NGF receptors during embryo development in the bursa of Fabricius [88,89], the specific avian organ where the maturation and differentiation of B-cells occurs. During the post-hatching period, TrkA is expressed in epithelial cells of the follicle, in the interfollicular epithelium of the bursa of Fabricius and in blood vessels [90,91]. The local production of NGF seems to be involved in follicle differentiation in vivo, since NGF administration accelerates follicle formation in the chick embryo [91] and influences the survival of bursal cells [92].
The expression of NGF receptors also characterizes secondary lymphoid organs (spleen, lymph nodes and mucosa-associated lymphoid tissues) where antigen-presenting cells activate lymphocyte responses and effector cell differentiation.
The spleen is composed of two different tissues: the white pulp, responsible for the production and growth of immune and blood cells, and the red pulp, which filters microorganisms and cellular debris from the blood and removes older erythrocytes from circulation [93]. The expression of p75NTR and TrkA is localized primarily in the stroma of the spleen, with some expression in splenocytes [78] and in spleen mononuclear immunocompetent cells [77]. In rats, during spleen development, the level of p75NTR mRNA increases [78] and is expressed mainly by splenic nerve fibers and in a subpopulation of dendritic cells [62].
In lymph nodes and in mucosa-associated lymphoid tissues, TrkA is present in follicular dendritic cells, in blood vessel walls, in cryptic tonsillar epithelium and in several monocyte-derived cells including epithelioid and multinucleated Langhans’ cells and interdigitated reticular cells [79,94,95]. The expression of p75NTR has been described in follicular dendritic cells of lymphoid follicles, interdigitated reticular cells, periarteriolar macrophages, endothelial sinusal cells and nerve endings, and is also found in moderate levels in dendritic cells of tonsillar follicles [79,95,96].
The picture emerging from all these studies is that the expression of NGF receptors characterizes both stromal and immune progenitors and is finely regulated during development and post-natal life. Thus, the local production of NGF not only directly and actively modulates innervation patterns and fiber density in lymphoid organs [97], but can also be involved in the creation of the correct microenvironment that regulates the differentiation of myeloid and lymphoid precursors [98]. A few in vitro studies have focused specifically on the response of stromal cells to NGF. After treatment of stromal cells derived from human bone marrow with NGF, microarray analysis demonstrated changes in cytokine gene expression in response to NGF that can influence hematopoiesis [99]. In thymic epithelial cells, the addition of NGF enhanced the expression of adhesion molecules needed for thymocyte-thymic epithelia interaction and up-regulated thymopoietic factor expression (SDF-1, IL-7, GM-CSF) [100].
3.2. Expression of NGF Receptors in Immune Cells
The expression of TrkA characterizes hemopoietic stem cells and is highest in the more undifferentiated cells, declining during lineage differentiation [85,86]. These stem cells can also produce their own NGF in an autocrine fashion, which seems necessary to increase the long-term survival and regulate differentiation of human hemopoietic cells [101,102,103,104]. As demonstrated in human cord blood cells, a gradient of TrkA and NGF expression exists that is highest in CD34-positive cells, reduced in cord blood mononuclear cells and minimal in mononuclear cells isolated from adult peripheral blood, and further declines with age [86,105]. TrkA expression is maintained during lineage differentiation and has been demonstrated in purified blood mononuclear cells, thymocytes, human B- and T-lymphocytes, monocytes, mast cells, basophils, eosinophils and neutrophils [86,105,106,107,108,109,110,111,112].
In B- and T-lymphocytes, undifferentiated monocytes and macrophages, the basal expression of TrkA is strongly up-regulated after antigenic or inflammatory stimulation when functional activity is necessary [106,107,108,113,114]. TrkA is poorly expressed in normal B- and T-lymphocytes, but is up-regulated in several B-cell lymphoma subtypes, anaplastic large cell lymphomas and Reed-Sternberg cells [79].
The expression of p75NTR has been described for many immune cells but there is still little information concerning changes in p75NTR during immune cell differentiation and activation [115,116]. A recent study on monocytes demonstrated that p75NTR expression is fairly low compared with TrkA in a normal resting condition [117]. Monocytes lose TrkA expression and only p75NTR expression is maintained [118] when these cells differentiate in dendritic cells, the antigen-presenting cells that interact closely with T-cells to form the immunological synapse and activate T-cell effector responses. This dynamic regulation of NGF receptor expression during immune cell differentiation and response suggests a differential need for NGF of immune cell populations, depending on their state of maturity and functional activity.
4. NGF and Its Direct and Indirect Effects on Immune Response
4.1. Indirect Action
The innervation of primary and secondary lymphoid organs constitutes the anatomical link between the nervous and immune systems. The integrity of this connection is essential to maintain a regulatory function of the nervous system on immune response: experiments of chemical sympathectomy, denervation of lymphoid organs or vagotomy [33,34,119] have clearly demonstrated alterations in the proliferation and responsiveness of immune cells and a modified migration. Nerve fibers end in the parenchyma in close contact with immune cells, and in this “neuroimmune junction” [120] the release of neurotransmitters from nerve endings can stimulate specific receptors present on immune cells affecting their functions. Muscarinic, nicotinic and adrenergic receptors are expressed on immune cells [121,122,123,124], and the release of neurotransmitters from nerve endings can finely tune the innate and adaptive immune responses by activating either pro- or anti-inflammatory pathways [33,34,119,120,121,122,123,124]. Sensory neuropeptides can also activate specific responses by binding to their specific receptors expressed on immune cells. The release of neuropeptides is a pivotal event in neurogenic inflammation, a well-studied neuroimmune mechanism involved in the development of inflammatory diseases such as rheumatoid arthritis, psoriasis, allergy and asthma [125,126]. Neuropeptides, including substance P (SP) and calcitonin gene-related peptide (CGRP), released from nerve endings induce vasodilatation and plasma extravasation, promote leukocyte chemotaxis and phagocytosis, and can directly affect the release of cytokines and inflammatory mediators from immune cells [127]. Thus different types of immune cell populations express “neuronal” receptors and respond to neuropeptide and neurotransmitters, indicating that peripheral neurons are an integral part of the local effector systems involved in the inflammatory responses to tissue irritation and injury.
Through its effects on neuronal cells, NGF can indirectly regulate the immune response in a variety of manners. It can affect neuronal plasticity in the inflamed tissue by increasing innervation density, axonal terminal sprouting and dendritic arborization.
In inflamed tissues there is a marked alteration in nerve fiber distribution, with an increased terminal sprouting leading to hyper-innervation of the injured tissues.
Inflammation leads to an increased sensitivity to pain and intense stimulation and firing of the nociceptors, which induces the release of neuropeptides and neurotransmitters from nerve endings in close contact with the cell membranes of keratinocytes, mast cells, macrophages, Langerhans cells and endothelial cells [33,34,119,120,121,122,123,124,125,126,127,128,129] (Figure 1).
NGF not only influences fiber density and dendritic sprouting in the inflamed tissues, but may also affect the local immune response by directly regulating the production of neurotransmitters and neuropeptides, which, as a growing body of findings is showing, have direct effects on immune cells and are involved in the development of many inflammatory diseases [4,5,6,7,8,9,13,14,15,16,19,20,29,30,97] (Figure 1). The increased concentration of NGF in inflamed tissues can regulate the expression of neuropeptides and neurotransmitters in peripheral neurons by directly acting on their promoters or by regulating the activity of neurotransmitter-producing enzymes. For example, in sympathetic neurons the production of norepinephrine is controlled by NGF through induction of the transcription of tyrosine hydroxylase, the rate-limiting enzyme of the catecholamine biosynthetic pathway [4]. In sensory neurons, however, NGF directly controls the expression of SP and CGRP. Thus, peripheral neurons exposed to enhanced concentrations of NGF in the inflamed target tissue modify the synthesis of neuropeptides and neurotransmitters and can alter their phenotype so that different types of neurotransmitters can be produced and stored in nerve endings. [6,13,14,29,130,131]. Many recent studies have focused on the release of acetylcholine and the activation of parasympathetic nerves because of their ability to inhibit inflammation and activate anti-inflammatory pathways [132]. Cholinergic neurons in the central nervous system are dependent on NGF for their survival and phenotype maintenance. Although NGF can regulate acetylcholine synthesis by increasing choline acetyltransferase activity [133], it is not clear whether it can influence parasympathetic neuron functions. Recent studies have shown that NGF can activate a parasympathetic tone. In mouse airway parasympathetic ganglia, the cholinergic neurons expressed TrkA and exposure to NGF potentiated synaptic transmission, enhancing their excitability [134,135]. In vivo administration of NGF altered dendritic length and sprouting in the bronchial ganglionic neurons of guinea pigs [134]. All together, these data suggest that NGF can influence parasympathetic neuron activity and contribute to activate regulatory cholinergic anti-inflammatory pathways responsible for the inhibition of pro-inflammatory cytokine release in the “inflammatory reflex” [136].
4.2. Direct Action
The presence of NGF receptors on cells other than neuronal cells opens new perspectives regarding the range of action of NGF. The first indication of an immunoregulatory effect of NGF was observed in 1977 by Aloe and Levi-Montalcini, who showed that injecting neonatal rats with NGF resulted in an increase in the number and size of mast cells [137]. It has now been amply demonstrated that TrkA and p75NTR are present on the surface of immune cells, as well as how their expression is modulated depending on the state of activation of the cells [108,111,114]. In vitro studies using purified immune cell populations have demonstrated a number of actions that can be ascribed to NGF and are summarized in Table 2.
NGF regulates the survival of different immune cells in a similar manner to its effect in neurons. It promotes the survival of hematopoietic stem cells [85], eosinophils [151], neutrophils [152,153], mast cells [149,150], B-cells [107] and monocytes [118]. The activation of TrkA pathways seems to inhibit apoptosis through regulation of the expression of Bcl-2, Bcl-xl and Bfl-1 [101,107,149,150]. Using different cell populations, it has been shown in vitro that NGF alone does not induce the synthesis of chemokines or cytokines. On the contrary, after inflammatory stimulation, the addition of NGF potentiates endogenous responses, inducing the release of cytokines and inflammatory mediators [138,139,140,141,142,143,144,145,146,147,151,153,154,162,166]. NGF also induces in immune cells the synthesis of neuropeptides with immunomodulatory functions: i.e., neuropeptide Y in T-cells [168], calcitonin gene-related peptide in monocytes [169], and B-cells [170]. Cells of the innate immune system respond to NGF by activating a variety of responses: NGF activates chemotaxis [148,164,165,171], stimulates the phagocytosis of neutrophils [153] and macrophages [164,165], enhances the cytotoxic activity of eosinophils [151], and induces the degranulation of mast-cells [146,149].
5. In Vivo Inflammatory and Anti-Inflammatory Mechanisms and the Roles of NGF and TrkA
Taken together, the available in vitro data suggest that NGF potentiates the activity of innate immune cells and influences B- and T-cell mediated responses. However, the mechanism by which NGF modulates the immune response in vivo is still not fully understood.
At present, despite the well-documented increase in NGF concentrations measured in the inflamed tissues of many inflammatory diseases, little is known of the effects of increased levels of NGF or of the activation of its receptors during inflammatory response. Currently available data suggest that the action of NGF is more complex in vivo than it is in vitro.
In animal models and in humans, the administration of NGF alone without a more specific inflammatory stimulus, while inducing activation of local sensory hypersensitivity, does not activate any inflammatory response [172,173,174,175].
However, the intraventricular administration of NGF after the induction of experimental autoimmune encephalomyelitis (EAE) in marmoset [176] or mouse models [177,178] delayed disease onset and decreased disease severity, preventing the full development of brain lesions. Reduced inflammatory infiltrate and demyelination were found in the brains of NGF-treated EAE animals, associated with a reduced production of interferon-gamma but with an enhanced production of IL-10 [176,178]. Similarly, when myelin basic protein-specific CD4+ T-cell clones were used to induce EAE, the contemporary injection of transfected T-clones encoding for NGF resulted in mild disease with minor symptoms [177]. Consistent with this observation, in vivo NGF deprivation in rats with EAE resulted in an increased brain inflammation and more severe clinical features [179]. Interestingly, it has been demonstrated in some animal models that the well-known increase in NGF production associated with inflammation remained elevated until the inflammatory response subsided [180]. Neutralization with anti-NGF antibodies of the enhanced production of NGF increased the disease symptoms with a higher number of infiltrating neutrophils and macrophages and more extended gut lesions in animal models of colitis [181]. Also, in contact hypersensitivity animal models, the neutralization of endogenous NGF resulted in a loss of inhibition of the systemic response [60].
Taken all together, the results obtained in different models of inflammation indicate that in vivo NGF administration ameliorates tissue inflammation, suggesting a direct influence of NGF on antigen presentation, down-regulation of inflammatory cytokines and up-regulation of the anti-inflammatory cytokine IL-10.
Although some discrepancies exist in the effects of NGF in vitro and in vivo, it seems reasonable to hypothesize that NGF is part of an endogenous mechanism that, while activating innate immune responses, also activates pathways necessary to dampen inflammatory response and limit tissue damage.
It should be kept in mind that inflammation is an adaptive response whose main aim is to restore homeostasis [182]. One key aspect of the inflammatory response and of its resolution is the parallel activation of pathways leading to inflammatory cytokine production that activates innate and adaptive immune responses, and of pathways that induce anti-inflammatory cytokines such as IL-10 and IL-1 receptor antagonists in order to avoid tissue destruction [183,184]. The possibility that NGF may be involved in these physiological mechanisms is supported by recent findings. In monocytes, inflammatory stimuli, while activating pro-inflammatory responses through Toll-like receptors (TLR), up-regulate the expression of the NGF receptor TrkA [108,114,117,118]. In TLR-activated monocytes, the binding of NGF to TrkA influences TLR signaling, decreasing NF-κB nuclear translocation and inhibiting glycogen synthase kinase 3 (GSK3) activity, which leads to a decreased production of inflammatory cytokines [117,185,186]. Moreover, TrkA activation further enhances the TLR-induced activation of the PI3K/Akt pathway. This pathway is one of the main anti-inflammatory pathways activated to reduce TLR ligand-induced inflammatory responses [184]. The activation of TrkA that followed TLR activation resulted in an increased production of anti-inflammatory cytokines [117,187]. Thus, the binding of NGF to TrkA can activate distinct pathways that influence the intracellular signaling activated after the recognition of inflammatory stimuli with an overall anti-inflammatory action, thus influencing the production of cytokines that are pivotal in orchestrating innate and adaptive immune responses (Figure 2). The activation of TrkA might be one of the signals that, when the immune system is activated, is involved in endogenous regulatory feed-back mechanisms enhancing the activation of anti-inflammatory pathways. This might explain the in vivo findings, in which the addition of NGF tilted the balance of the pro- and anti-inflammatory pathways that are simultaneously activated during inflammatory response, potentiating the release of anti-inflammatory mediators such as IL-10 [183,184,187]. Consequently, changes in TrkA expression levels may affect the physiological anti-inflammatory mechanism mediated by NGF. Consistent with this hypothesis, patients with chronic arthritis have a marked down-regulation of TrkA expression in blood and synovial mononuclear cells when compared with healthy donors [117]. In ex vivo experiments, the addition of NGF to TLR-activated mononuclear cells of arthritis patients, which are characterized by a significant decrease in TrkA expression, failed to reduce the production of IL-6 that was instead observed after NGF treatment in healthy donor cells expressing high TrkA levels [117]. The TrkA imbalance found in arthritis patients might prevent NGF from activating the regulatory anti-inflammatory feed-back mechanisms, thus contributing to the development and maintenance of chronic inflammation. Further studies aimed at evaluating TrkA expression in other inflammatory diseases may help us to understand whether this is a general mechanism in chronic inflammatory diseases.
6. Conclusions
A growing body of data suggests that NGF is a key regulator of the cross-talk between the immune and nervous systems. While the effects of NGF on neuronal cell survival, differentiation and phenotype maintenance are fairly well-known, the effects of NGF on immune cells have still not been completely defined. Further investigations are needed to establish why NGF is synthesized in vivo in the inflamed tissue and how the activation of its receptors can influence the cellular and molecular mechanisms triggered during the inflammatory response. What has already been demonstrated in neuronal cells and is now emerging in studies on immune cells and inflammatory diseases is that NGF definitely controls a multitude of effects, some of them even contradictory. The diverse biological activities of NGF on immune cell populations can be explained by considering the differential expression of NGF receptors during different states of cell differentiation and activation. The complexity of NGF action and its signaling can be better understood if we consider them as being correlated with the need for differing types of regulation in a variety of immune cell types in order to maintain correct homeostasis. Alteration in NGF synthesis and modification of the expression of its receptors may influence physiological responses and be involved in the pathogenesis of inflammatory diseases.
A better understanding of the physiological role of NGF and its receptors in regulating immune responses could be of groundbreaking importance with translational implications for human diseases.
Acknowledgments
This study was supported by research funding from the IRCCS Bambino Gesù Children’s Hospital “Ricerca Corrente” to Fabrizio De Benedetti.
Author Contributions
Conflicts of Interest
The authors declare no conflict of interest.
References
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Riccio, A.; Ahn, S.; Davenport, C.M.; Blendy, J.A.; Ginty, D.D. Mediation by a CREB family transcription factor of NGF-dependent survival of sympathetic neurons. Science 1999, 286, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Harrington, A.W.; Ginty, D.D. Long-distance retrograde neurotrophic factor signalling in neurons. Nat. Rev. Neurosci. 2013, 14, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Levi-Montalcini, R.; Aloe, L.; Mugnaini, E.; Oesch, F.; Thoenen, H. Nerve growth factor induces volume increase and enhances tyrosine hydroxylase synthesis in chemically axotomized sympathetic ganglia of newborn rats. Proc. Natl. Acad. Sci. USA 1975, 72, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, R.M.; Harmar, A.J. Nerve growth factor regulates expression of neuropeptide genes in adult sensory neurons. Nature 1989, 337, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Leslie, T.A.; Emson, P.C.; Dowd, P.M.; Woolf, C.J. Nerve growth factor contributes to the up-regulation of growth-associated protein 43 and preprotachykinin A messenger RNAs in primary sensory neurons following peripheral inflammation. Neuroscience 1995, 67, 753–761. [Google Scholar] [CrossRef]
- Zigmond, R.E.; Sun, Y. Regulation of neuropeptide expression in sympathetic neurons. Paracrine and retrograde influences. Ann. N. Y. Acad. Sci. 1997, 814, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Korsching, S.; Thoenen, H. Nerve growth factor in sympathetic ganglia and corresponding target organs of the rat: Correlation with density of sympathetic innervation. Proc. Natl. Acad. Sci. USA 1983, 80, 3513–3516. [Google Scholar] [CrossRef] [PubMed]
- Thrasivoulou, C.; Cowen, T. Regulation of rat sympathetic nerve density by target tissues and NGF in maturity and old age. Eur. J. Neurosci. 1995, 7, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, J.; McNaughton, P.A. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 2005, 24, 4211–4223. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, L.G.; White, G. Effects of nerve growth factor on TTX- and capsaicin-sensitivity in adult rat sensory neurons. Brain Res. 1992, 570, 61–66. [Google Scholar] [CrossRef]
- Gould, H.J.; Gould, T.N.; England, J.D.; Paul, D.; Liu, Z.P.; Levinson, S.R. A possible role for nerve growth factor in the augmentation of sodium channels in models of chronic pain. Brain Res. 2000, 854, 19–29. [Google Scholar] [CrossRef]
- Verge, V.M.; Richardson, P.M.; Wiesenfeld-Hallin, Z.; Hökfelt, T. Differential influence of nerve growth factor on neuropeptide expression in vivo: A novel role in peptide suppression in adult sensory neurons. J. Neurosci. 1995, 15, 2081–2096. [Google Scholar] [PubMed]
- Donnerer, J.; Schuligoi, R.; Stein, C. Increased content and transport of substance P and calcitonin gene-related peptide in sensory nerves innervating inflamed tissue: Evidence for a regulatory function of nerve growth factor in vivo. Neuroscience 1992, 49, 693–698. [Google Scholar] [CrossRef]
- Bennett, D.L.; Koltzenburg, M.; Priestley, J.V.; Shelton, D.L.; McMahon, S.B. Endogenous nerve growth factor regulates the sensitivity of nociceptors in the adult rat. Eur. J. Neurosci. 1998, 10, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Spillane, M.; Ketschek, A.; Donnelly, C.J.; Pacheco, A.; Twiss, J.L.; Gallo, G. Nerve growth factor-induced formation of axonal filopodia and collateral branches involves the intra-axonal synthesis of regulators of the actin-nucleating ARP2/3 complex. J. Neurosci. 2012, 32, 17671–17689. [Google Scholar] [CrossRef] [PubMed]
- Albers, K.M.; Wright, D.E.; Davis, B.M. Overexpression of nerve growth factor in epidermis of transgenic mice causes hypertrophy of the peripheral nervous system. J. Neurosci. 1994, 14, 1422–1432. [Google Scholar] [PubMed]
- Takami, S.; Getchell, M.L.; Yamagishi, M.; Albers, K.M.; Getchell, T.V. Enhanced extrinsic innervation of nasal and oral chemosensory mucosae in keratin 14-NGF transgenic mice. Cell Tissue Res. 1995, 282, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Goodness, T.P.; Albers, K.M.; Davis, F.E.; Davis, B.M. Overexpression of nerve growth factor in skin increases sensory neuron size and modulates Trk receptor expression. Eur. J. Neurosci. 1997, 9, 1574–1585. [Google Scholar] [CrossRef] [PubMed]
- Schnegelsberg, B.; Sun, T.T.; Cain, G.; Bhattacharya, A.; Nunn, P.A.; Ford, A.P.; Vizzard, M.A.; Cockayne, D.A. Overexpression of NGF in mouse urothelium leads to neuronal hyperinnervation, pelvic sensitivity, and changes in urinary bladder function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R534–R547. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Levi-Montalcini, R.; Buttinelli, C.; Schilter, D.; Gillessen, S.; Otten, U. Multiple sclerosis patients express increased levels of B-nerve growth factor in cerebrospinal fluid. Neurosci. Lett. 1992, 147, 9–12. [Google Scholar] [CrossRef]
- Aloe, L.; Tuveri, M.A.; Carcassi, U.; Levi-Montalcini, R. Nerve growth factor in the synovial fluid of patients with chronic arthritis. Arthritis Rheumatol. 1992, 35, 351–355. [Google Scholar] [CrossRef]
- Falcini, F.; Matucci Cerinic, M.; Lombardi, A.; Generini, S.; Pignone, A.; Tirassa, P.; Ermini, M.; Lepore, L.; Partsch, G.; Aloe, L. Increased circulating nerve growth factor is directly correlated with disease activity in juvenile chronic arthritis. Ann. Rheum. Dis. 1996, 55, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Levi-Montalcini, R.; Galeazzi, M.; Schilter, D.; Scully, J.L.; Otten, U. Increased levels of NGF in sera of systemic lupus erythematosus patients. Neuro. Report 1992, 4, 563–565. [Google Scholar]
- Aalto, K.; Korhonen, L.; Lahdenne, P.; Pelkonen, P.; Lindholm, D. Nerve growth factor in serum of children with systemic lupus erythematosus is correlated with disease activity. Cytokine 2002, 20, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Fauchais, A.L.; Boumediene, A.; Lalloue, F.; Gondran, G.; Loustaud-Ratti, V.; Vidal, E.; Jauberteau, M.O. Brain-derived neurotrophic factor and nerve growth factor correlate with T-cell activation in primary Sjogren’s syndrome. Scand. J. Rheumatol. 2009, 38, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Samad, T.A.; Jin, S.X.; Schmoll, R.; Woolf, C.J. p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron 2002, 36, 57–68. [Google Scholar] [CrossRef]
- Mamet, J.; Lazdunski, M.; Voilley, N. How nerve growth factor drives physiological and inflammatory expressions of acid-sensing ion channel 3 in sensory neurons. J. Biol. Chem. 2003, 278, 48907–48913. [Google Scholar] [CrossRef] [PubMed]
- Ghilardi, J.R.; Freeman, K.T.; Jimenez-Andrade, J.M.; Coughlin, K.A.; Kaczmarska, M.J.; Castaneda-Corral, G.; Bloom, A.P.; Kuskowski, M.A.; Mantyh, P.W. Neuroplasticity of sensory and sympathetic nerve fibers in a mouse model of a painful arthritic joint. Arthritis Rheumatol. 2012, 64, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Reinert, A.; Kaske, A.; Mense, S. Inflammation-induced increase in the density of neuropeptide-immunoreactive nerve endings in rat skeletal muscle. Exp. Brain Res. 1998, 121, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Howe, C.L.; Mobley, W.C. Nerve growth factor signaling, neuroprotection, and neural repair. Annu. Rev. Neurosci. 2001, 24, 1217–1281. [Google Scholar] [CrossRef] [PubMed]
- Kioussis, D.; Pachnis, V. Immune and nervous systems: More than just a superficial similarity? Immunity 2009, 31, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Madden, K.S.; Felten, D.L. Experimental basis for neuronal-immune interactions. Physiol. Rev. 1995, 75, 77–106. [Google Scholar] [PubMed]
- Elenkov, I.J.; Wilder, R.L.; Chrousos, G.P.; Vizi, E.S. The sympathetic nerve—An integrative interface between two supersystems: The brain and the immune system. Pharmacol. Rev. 2000, 52, 595–638. [Google Scholar] [PubMed]
- Aloe, L.; Probert, L.; Kollias, G.; Bracci-Laudiero, L.; Spillantini, M.G.; Levi-Montalcini, R. The synovium of transgenic arthritic mice expressing human tumor necrosis factor contains a high level of nerve growth factor. Growth Factors 1993, 9, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Manni, L.; Aloe, L. Role of IL-1β and TNF-α in the regulation of NGF in experimentally induced arthritis in mice. Rheumatol. Int. 1998, 18, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Lundeberg, T.; Stenfors, C.; Theodorsson, E.; Tirassa, P.; Aloe, L. Modification of lymphoid and brain nerve growth factor levels in systemic lupus erythematosus mice. Neurosci. Lett. 1996, 204, 13–16. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Smaldone, M.C.; Tyagi, V.; Philips, B.J.; Jackman, S.V.; Leng, W.W.; Tyagi, P. Increased nerve growth factor in neurogenic overactive bladder and interstitial cystitis patients. Can. J. Urol. 2010, 17, 4989–4994. [Google Scholar] [PubMed]
- Bonini, S.; Lambiase, A.; Bonini, S.; Angelucci, F.; Magrini, L.; Manni, L.; Aloe, L. Circulating nerve growth factor levels are increased in humans with allergic diseases and asthma. Proc. Natl. Acad. Sci. USA 1996, 93, 10955–10960. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Bonini, S.; Bonini, S.; Micera, A.; Magrini, L.; Bracci-Laudiero, L.; Aloe, L. Increased plasma levels of nerve growth factor in vernal keratoconjunctivitis and relationship to conjunctival mast cells. Investig. Ophthalmol. Vis. Sci. 1995, 36, 2127–2132. [Google Scholar]
- Di Mola, F.F.; Friess, H.; Zhu, Z.W.; Koliopanos, A.; Bley, T.; di Sebastiano, P.; Innocenti, P.; Zimmermann, A.; Büchler, M.W. Nerve growth factor and Trk high affinity receptor (TrkA) gene expression in inflammatory bowel disease. Gut 2000, 46, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Stanzel, R.D.; Lourenssen, S.; Blennerhassett, M.G. Inflammation causes expression of NGF in epithelial cells of the rat colon. Exp. Neurol. 2008, 211, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Fantini, F.; Magnoni, C.; Bracci-Laudiero, L.; Pincelli, C.T.E. Nerve growth factor is increased in psoriatic skin. J. Investig. Dermatol. 1995, 105, 854–855. [Google Scholar] [PubMed]
- Raychaudhuri, S.P.; Jiang, W.Y.; Farber, E.M. Psoriatic keratinocytes express high levels of nerve growth factor. Acta Dermato. Venereol. 1998, 78, 84–86. [Google Scholar]
- Toyoda, M.; Nakamura, M.; Makino, T.; Hino, T.; Kagoura, M.; Morohashi, M. Nerve growth factor and substance P are useful plasma markers of disease activity in atopic dermatitis. Br. J. Dermatol. 2002, 147, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J.; Allchorne, A.; Safieh-Garabedian, B.; Poole, S. Cytokines, nerve growth factor and inflammatory hyperalgesia: The contribution of tumour necrosis factor α. Br. J. Pharmacol. 1997, 121, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Steiner, P.; Pfeilschifter, J.; Boeckh, C.; Radeke, H.; Otten, U. Interleukin-1β and tumor necrosis factor-α synergistically stimulate nerve growth factor synthesis in rat mesangial cells. Am. J. Physiol. 1991, 261, 792–798. [Google Scholar]
- März, P.; Heese, K.; Dimitriades-Schmutz, B.; Rose-John, S.; Otten, U. Role of interleukin-6 and soluble IL-6 receptor in region-specific induction of astrocytic differentiation and neurotrophin expression. Glia 1999, 26, 191–200. [Google Scholar] [CrossRef]
- Freund, V.; Pons, F.; Joly, V.; Mathieu, E.; Martinet, N.; Frossard, N. Upregulation of nerve growth factor expression by human airway smooth muscle cells in inflammatory conditions. Eur. Respir. J. 2002, 20, 458–463. [Google Scholar] [CrossRef] [PubMed]
- Manni, L.; Lundeberg, T.; Fiorito, S.; Bonini, S.; Vigneti, E.; Aloe, L. Nerve growth factor release by human synovial fibroblasts prior to and following exposure to tumor necrosis factor-α, interleukin-1β and cholecystokinin-8: The possible role of NGF in the inflammatory response. Clin. Exp. Rheumatol. 2003, 21, 617–624. [Google Scholar] [PubMed]
- Wang, B.; Jenkins, J.R.; Trayhurn, P. Expression and secretion of inflammation-related adipokines by human adipocytes differentiated in culture: Integrated response to TNF-α. Am. J. Physiol. Endocrinol. Metab. 2005, 288, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Von Boyen, G.B.; Steinkamp, M.; Reinshagen, M.; Schäfer, K.H.; Adler, G.; Kirsch, J. Nerve growth factor secretion in cultured enteric glia cells is modulated by proinflammatory cytokines. J. Neuroendocrinol. 2006, 18, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Bulló, M.; Peeraully, M.R.; Trayhurn, P. Stimulation of NGF expression and secretion in 3T3-L1 adipocytes by prostaglandins PGD2, PGJ2, and δ12-PGJ2. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E62–E67. [Google Scholar] [CrossRef] [PubMed]
- Toyomoto, M.; Ohta, M.; Okumura, K.; Yano, H.; Matsumoto, K.; Inoue, S.; Hayashi, K.; Ikeda, K. Prostaglandins are powerful inducers of NGF and BDNF production in mouse astrocyte cultures. FEBS Lett. 2004, 562, 211–215. [Google Scholar] [CrossRef]
- Lipnik-Stangelj, M.; Carman-Krzan, M. Activation of histamine H1-receptor enhances neurotrophic factor secretion from cultured astrocytes. Inflamm. Res. 2004, 53, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Watanabe, S. Histamine enhances the production of nerve growth factor in human keratinocytes. J. Investig. Dermatol. 2003, 121, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Iannone, F.; de Bari, C.; Dell’Accio, F.; Covelli, M.; Patella, V.; Lo Bianco, G.; Lapadula, G. Increased expression of nerve growth factor (NGF) and high affinity NGF receptor (p140 TrkA) in human osteoarthritic chondrocytes. Rheumatology 2002, 41, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Rihl, M.; Kruithof, E.; Barthel, C.; de Keyser, F.; Veys, E.M.; Zeidler, H.; Yu, D.T.; Kuipers, J.G.; Baeten, D. Involvement of neurotrophins and their receptors in spondyloarthritis synovitis: Relation to inflammation and response to treatment. Ann. Rheum. Dis. 2005, 64, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Barthel, C.; Yeremenko, N.; Jacobs, R.; Schmidt, R.E.; Bernateck, M.; Zeidler, H.; Tak, P.P.; Baeten, D.; Rihl, M. Nerve growth factor and receptor expression in rheumatoid arthritis and spondyloarthritis. Arthritis Res. Ther. 2009, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Townley, S.L.; Grimbaldeston, M.A.; Ferguson, I.; Rush, R.A.; Zhang, S.H.; Zhou, X.F.; Conner, J.M.; Finlay-Jones, J.J.; Hart, P.H. Nerve growth factor, neuropeptides, and mast cells in ultraviolet-B-induced systemic suppression of contact hypersensitivity responses in mice. J. Investig. Dermatol. 2002, 118, 396–401. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Germanà, A.; Hannestad, J.; Ciriaco, E.; Laurà, R.; Naves, J.; Esteban, I.; Silos-Santiago, I.; Vega, J.A. TrkA is necessary for the normal development of the murine thymus. J. Neuroimmunol. 2000, 108, 11–21. [Google Scholar] [CrossRef]
- Pérez-Pérez, M.; García-Suárez, O.; Esteban, I.; Germanà, A.; Fariñas, I.; Naves, F.J.; Vega, J.A. p75NTR in the spleen: Age-dependent changes, effect of NGF and 4-methylcatechol treatment, and structural changes in p75NTR-deficient mice. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2003, 270, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Indo, Y.; Tsuruta, M.; Hayashida, Y.; Karim, M.A.; Ohta, K.; Kawano, T.; Mitsubuchi, H.; Tonoki, H.; Awaya, Y.; Matsuda, I. Mutations in the TrkA/NGF receptor gene in patients with congenital insensitivity to pain with anhidrosis. Nat. Genet. 1996, 13, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Melamed, I.; Levy, J.; Parvari, R.; Gelfand, E.W. A novel lymphocyte signaling defect: Trk A mutation in the syndrome of congenital insensitivity to pain and anhidrosis (CIPA). J. Clin. Immunol. 2004, 24, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Tsuboi, Y.; Kurosawa, H.; Sugita, K.; Eguchi, M. Anti-apoptotic effect of nerve growth factor is lost in congenital insensitivity to pain with anhidrosis (CIPA) B lymphocytes. J. Clin. Immunol. 2004, 24, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Barone, R.; Lempereur, L.; Anastasi, M.; Parano, E.; Pavone, P. Congenital insensitivity to pain with Anhidrosis (NTRK1 mutation) and early onset renal disease: Clinical report on three sibs with a 25-year follow-up in one of them. Neuropediatrics 2005, 36, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Hehlgans, T.; Pfeffer, K. The intriguing biology of the tumour necrosis factor/tumour necrosis factor receptor superfamily: Players, rules and the games. Immunology 2005, 115, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, M. Recent advances in understanding neurotrophin signaling. F1000 Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Deinhardt, K.; Chao, M.V. Trk receptors. Handb. Exp. Pharmacol. 2014, 220, 103–119. [Google Scholar] [PubMed]
- Shu, Y.H.; Lu, X.M.; Wei, J.X.; Xiao, L.; Wang, Y.T. Update on the role of p75NTR in neurological disorders: A novel therapeutic target. Biomed. Pharmacother. 2015, 76, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kurd, N.; Robey, E.A. T-cell selection in the thymus: A spatial and temporal perspective. Immunol. Rev. 2016, 271, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Johnson, E.M., Jr. An immunohistochemical study of the nerve growth factor receptor in developing rats. J. Neurosci. 1988, 8, 3481–3498. [Google Scholar] [PubMed]
- Parrens, M.; Labouyrie, E.; Groppi, A.; Dubus, P.; Carles, D.; Velly, J.F.; de Mascarel, A.; Merlio, J.P. Expression of NGF receptors in normal and pathological human thymus. J. Neuroimmunol. 1998, 85, 11–21. [Google Scholar] [CrossRef]
- Marinova, T.T.; Velikova, K.K.; Petrov, D.B.; Kutev, N.S.; Stankulov, I.S.; Chaldakov, G.N.; Triaca, V.; Manni, L.; Aloe, L. Structural and ultrastructural localization of NGF and NGF receptors in the thymus of subjects affected by myasthenia gravis. Autoimmunity 2004, 37, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Hannestad, J.; García-Suárez, O.; Huerta, J.J.; Esteban, I.; Naves, F.J.; Vega, J.A. TrkA neutrophin receptor protein in the rat and human thymus. Anat. Rec. 1997, 249, 373–379. [Google Scholar] [CrossRef]
- Dubus, P.; Parrens, M.; El-Mokhtari, Y.; Ferrer, J.; Groppi, A.; Merlio, J.P. Identification of novel TrkA variants with deletions in leucine-rich motifs of the extracellular domain. J. Neuroimmunol. 2000, 107, 42–49. [Google Scholar] [CrossRef]
- Laurenzi, M.A.; Barbany, G.; Timmusk, T.; Lindgren, J.A.; Persson, H. Expression of mRNA encoding neurotrophins and neurotrophin receptors in rat thymus, spleen tissue and immunocompetent cells. Regulation of neurotrophin-4 mRNA expression by mitogens and leukotriene B4. Eur. J. Biochem. 1994, 223, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Lomen-Hoerth, C.; Shooter, E.M. Widespread neurotrophin receptor expression in the immune system and other nonneuronal rat tissues. J. Neurochem. 1995, 64, 1780–1789. [Google Scholar] [CrossRef] [PubMed]
- Labouyrie, E.; Parrens, M.; de Mascarel, A.; Bloch, B.; Merlio, J.P. Distribution of NGF receptors in normal and pathologic human lymphoid tissues. J. Neuroimmunol. 1997, 77, 161–173. [Google Scholar] [CrossRef]
- Garcia-Suárez, O.; Germanà, A.; Hannestad, J.; Ciriaco, E.; Silos-Santiago, I.; Germanà, G.; Vega, J.A. Involvement of the NGF receptors (TrkA and p75LNGFR) in the development and maintenance of the thymus. Ital. J. Anat. Embryol. 2001, 106, 279–285. [Google Scholar] [PubMed]
- Aloe, L.; Micera, A.; Bracci-Laudiero, L.; Vigneti, E.; Turrini, P. Presence of nerve growth factor in the thymus of prenatal, postnatal and pregnant rats. Thymus 1997, 24, 221–231. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Germanà, A.; Hannestad, J.; Pérez-Pérez, M.; Esteban, I.; Naves, F.J.; Vega, J.A. Changes in the expression of the nerve growth factor receptors TrkA and p75LNGR in the rat thymus with ageing and increased nerve growth factor plasma levels. Cell Tissue Res. 2000, 301, 225–234. [Google Scholar] [PubMed]
- Parrens, M.; Dubus, P.; Groppi, A.; Velly, J.F.; Labouyrie, E.; de Mascarel, A.; Merlio, J.P. Differential expression of NGF receptors in human thymic epithelial tumors. Pathol. Res. Pract. 1999, 195, 549–553. [Google Scholar] [CrossRef]
- Cattoretti, G.; Schirò, R.; Orazi, A.; Soligo, D.; Colombo, M.P. Bone marrow stroma in humans: Anti-nerve growth factor receptor antibodies selectively stain reticular cells in vivo and in vitro. Blood 1993, 81, 1726–1738. [Google Scholar] [PubMed]
- Chevalier, S.; Praloran, V.; Smith, C.; MacGrogan, D.; Ip, N.Y.; Yancopoulos, G.D.; Brachet, P.; Pouplard, A.; Gascan, H. Expression and functionality of the TrkA proto-oncogene product/NGF receptor in undifferentiated hematopoietic cells. Blood 1994, 83, 1479–1485. [Google Scholar] [PubMed]
- Bracci-Laudiero, L.; Celestino, D.; Starace, G.; Antonelli, A.; Lambiase, A.; Procoli, A.; Rumi, C.; Lai, M.; Picardi, A.; Ballatore, G.; et al. CD34-positive cells in human umbilical cord blood express nerve growth factor and its specific receptor TrkA. J. Neuroimmunol. 2001, 136, 130–139. [Google Scholar] [CrossRef]
- Paczkowska, E.; Piecyk, K.; Luczkowska, K.; Kotowski, M.; Roginska, D.; Pius-Sadowska, E.; Oronowicz, K.; Ostrowski, M.; Machalinski, B. Expression of neurotrophins and their receptors in human CD34+ bone marrow cells. J. Physiol. Pharmacol. 2016, 67, 151–159. [Google Scholar] [PubMed]
- Ernfors, P.; Hallböök, F.; Ebendal, T.; Shooter, E.M.; Radeke, M.J.; Misko, T.P.; Persson, H. Developmental and regional expression of β-nerve growth factor receptor mRNA in the chick and rat. Neuron 1988, 1, 983–996. [Google Scholar] [CrossRef]
- Ciriaco, E.; Dall’Aglio, C.; Hannestad, J.; Huerta, J.J.; Laurà, R.; Germanà, G.; Vega, J.A. Localization of TrK neurotrophin receptor-like proteins in avian primary lymphoid organs: Thymus and bursa of Fabricius. J. Neuroimmunol. 1996, 69, 73–83. [Google Scholar] [CrossRef]
- Ciriaco, E.; García-Suárez, O.; Ricci, A.; Abbate, F.; Piedimonte, G.; Vega, J.A. Trk-like proteins during the post-hatching growth of the avian bursa of Fabricius. Vet. Immunol. Immunopathol. 1997, 55, 313–320. [Google Scholar] [CrossRef]
- Bracci-Laudiero, L.; Vigneti, E.; Aloe, L. In vivo and in vitro effect of NGF on bursa of Fabricius cells during chick embryo development. Int. J. Neurosci. 1991, 59, 189–198. [Google Scholar] [CrossRef]
- Bracci-Laudiero, L.; Vigneti, E.; Iannicola, C.; Aloe, L. NGF retards apoptosis in chick embryo bursal cell in vitro. Differentiation 1993, 53, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Pittet, M.J. The spleen in local and systemic regulation of immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Bronzetti, E.; Artico, M.; Pompili, E.; Felici, L.M.; Stringaro, A.; Bosco, S.; Magliulo, G.; Colone, M.; Arancia, G.; Vitale, M.; et al. Neurotrophins and neurotransmitters in human palatine tonsils: An immunohistochemical and RT-PCR analysis. Int. J. Mol. Med. 2006, 18, 49–58. [Google Scholar] [CrossRef] [PubMed]
- García-Suárez, O.; Hannestad, J.; Esteban, I.; Martínez del Valle, M.; Naves, F.J.; Vega, J.A. Neurotrophin receptor-like protein immunoreactivity in human lymph nodes. Anat. Rec. 1997, 249, 226–232. [Google Scholar] [CrossRef]
- Pezzati, P.; Stanisz, A.M.; Marshall, J.S.; Bienenstock, J.; Stead, R.H. Expression of nerve growth factor receptor immunoreactivity on follicular dendritic cells from human mucosa associated lymphoid tissues. Immunology 1992, 76, 485–490. [Google Scholar] [PubMed]
- Carlson, S.L.; Albers, K.M.; Beiting, D.J.; Parish, M.; Conner, J.M.; Davis, B.M. NGF modulates sympathetic innervation of lymphoid tissues. J. Neurosci. 1995, 15, 5892–5899. [Google Scholar] [PubMed]
- Lucas, D.; Scheiermann, C.; Chow, A.; Kunisaki, Y.; Bruns, I.; Barrick, C.; Tessarollo, L.; Frenette, P.S. Chemotherapy-induced bone marrow nerve injury impairs hematopoietic regeneration. Nat. Med. 2013, 19, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, F.; Rellick, S.L.; Piedimonte, G.; Akers, S.M.; O’Leary, H.A.; Martin, K.; Craig, M.D.; Gibson, L.F. Neurotrophins regulate bone marrow stromal cell IL-6 expression through the MAPK pathway. PLoS ONE 2010, 5, e9690. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Na, Y.J.; Jung, P.K.; Kim, M.N.; Kim, S.M.; Chung, J.S.; Kim, B.S.; Kim, J.B.; Moon, J.O.; Yoon, S. Nerve growth factor stimulates proliferation, adhesion and thymopoietic cytokine expression in mouse thymic epithelial cells in vitro. Regul. Pept. 2008, 147, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Auffray, I.; Chevalier, S.; Froger, J.; Izac, B.; Vainchenker, W.; Gascan, H.; Coulombel, L. Nerve growth factor is involved in the supportive effect by bone marrow derived stromal cells of the factor-dependent human cell line UT-7. Blood 1996, 88, 1608–1618. [Google Scholar] [PubMed]
- Matsuda, H.; Coughlin, M.D.; Bienenstock, J.; Denburg, J.A. Nerve growth factor promotes human hemapoietic colony growth and differentiation. Proc. Natl. Acad. Sci. USA 1988, 85, 6508–6512. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Wong, D.; Dolovich, J.; Bienenstock, J.; Marshall, J.; Denburg, J.A. Synergistic effects of nerve growth factor and granulocytes-macrophage colony-stimulating factor on human basophilic cell differentiation. Blood 1991, 77, 971–979. [Google Scholar] [PubMed]
- Welker, P.; Grabbe, J.; Gibbs, B.; Zuberbier, T.; Henz, B.M. Nerve growth factor-β induces mast-cell marker expression during in vitro culture of human umbilical cord blood cells. Immunology 2000, 99, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Bracci-Laudiero, L.; Aloe, L. Altered plasma nerve growth factor-like immunoreactivity and nerve growth factor-receptor expression in human old age. Gerontology 2003, 49, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Ehrhard, P.B.; Erb, P.; Graumann, U.; Otten, U. Expression of nerve growth factor and nerve growth factor receptor tyrosine kinase Trk in activated CD4-positive T-cell clones. Proc. Natl. Acad. Sci. USA 1993, 90, 10984–10988. [Google Scholar] [CrossRef] [PubMed]
- Torcia, M.; Bracci-Laudiero, L.; Lucibello, M.; Nencioni, L.; Labardi, D.; Rubartelli, A.; Cozzolino, F.; Aloe, L.; Garaci, E. Nerve growth factor is an autocrine survival factor for memory B lymphocytes. Cell 1996, 85, 345–356. [Google Scholar] [CrossRef]
- Ehrhard, P.B.; Ganter, U.; Bauer, J.; Otten, U. Expression of functional Trk protooncogene in human monocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 5423–5427. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, G.; Forsberg-Nilsson, K.; Xiang, Z.; Hallböök, F.; Nilsson, K.; Metcalfe, D.D. Human mast cells express functional TrkA and are a source of nerve growth factor. Eur. J. Immunol. 1997, 27, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Burgi, B.; Otten, U.H.; Ochensberger, B.; Rihs, S.; Heese, K.; Ehrhard, P.B.; Ibanez, C.F.; Dahinden, C.A. Basophil priming by neurotrophic factors. Activation through the Trk receptor. J. Immunol. 1996, 157, 5582–5588. [Google Scholar]
- Noga, O.; Englmann, C.; Hanf, G.; Grützkau, A.; Guhl, S.; Kunkel, G. Activation of the specific neurotrophin receptors TrkA, TrkB and TrkC influences the function of eosinophils. Clin. Exp. Allergy 2002, 32, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Oda, M.; Ikari, S.; Matsuno, T.; Morimune, Y.; Nagahama, M.; Sakurai, J. Signal transduction mechanism involved in Clostridium perfringens α-toxin-induced superoxide anion generation in rabbit neutrophils. Infect. Immun. 2006, 74, 2876–2886. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Bracci-Laudiero, L.; Bonini, Se.; Bonini, St.; Starace, G.; D’Elios, M.M.; de Carli, M.; Aloe, L. Human CD4+ T-clones produce and release nerve growth factor and express TrkA. J. Allergy Clin. Immunol. 1997, 100, 408–414. [Google Scholar] [CrossRef]
- Caroleo, M.C.; Costa, N.; Bracci-Laudiero, L.; Aloe, L. Human monocyte/macrophages activate by exposure to LPS overexpress NGF and NGF receptors. J. Neuroimmunol. 2001, 113, 193–201. [Google Scholar] [CrossRef]
- Fischer, T.C.; Lauenstein, H.D.; Serowka, F.; Pilzner, C.; Groneberg, D.A.; Welker, P. Pan-neurotrophin receptor p75NTR expression is strongly induced in lesional atopic mast cells. Clin. Exp. Allergy 2008, 38, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.L.; Bailey, S.; Matusica, D.; Nicholson, I.; Muyderman, H.; Pagadala, P.C.; Neet, K.E.; Zola, H.; Macardle, P.; Rush, R.A. ProNGF mediates death of Natural Killer cells through activation of the p75NTR-sortilin complex. J. Neuroimmunol. 2010, 226, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Prencipe, G.; Minnone, G.; Strippoli, R.; de Pasquale, L.; Petrini, S.; Caiello, I.; Manni, L.; de Benedetti, F.; Bracci-Laudiero, L. Nerve growth factor downregulates inflammatory response in human monocytes through TrkA. J. Immunol. 2014, 192, 3345–3354. [Google Scholar] [CrossRef] [PubMed]
- La Sala, A.; Corinti, S.; Federici, M.; Saragovi, H.U.; Girolomoni, G. Ligand activation of nerve growth factor receptor TrkA protects monocytes from apoptosis. J. Leukoc. Biol. 2000, 68, 104–110. [Google Scholar] [PubMed]
- Pavlov, V.A.; Tracey, K.J. Neural circuitry and immunity. Immunol. Res. 2015, 63, 38–57. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H. Complexity of the bi-directional neuroimmune junction in the spleen. Trends Pharmacol. Sci. 2004, 25, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Fujii, T.; Moriwaki, Y.; Misawa, H. Critical roles of acetylcholine and the muscarinic and nicotinic acetylcholine receptors in the regulation of immune function. Life Sci. 2012, 91, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, D.L.; Lorton, D. Autonomic regulation of cellular immune function. Auton. Neurosci. 2001, 182, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Sanders, V.M. The β2-adrenergic receptor on T- and B-lymphocytes: Do we understand it yet? Brain Behav. Immun. 2012, 26, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Pongratz, G.; Straub, R.H. The sympathetic nervous response in inflammation. Arthritis Res. Ther. 2014, 16, 504. [Google Scholar] [CrossRef] [PubMed]
- Chiu, I.M.; von Hehn, C.A.; Woolf, C.J. Neurogenic inflammation and the peripheral nervous system in host defense and immunopathology. Nat. Neurosci. 2012, 15, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Talbot, S.; Abdulnour, R.E.; Burkett, P.R.; Lee, S.; Cronin, S.J.; Pascal, M.A.; Laedermann, C.; Foster, S.L.; Tran, J.V.; Lai, N.; et al. Silencing nociceptor neurons reduces allergic airway inflammation. Neuron 2015, 87, 341–354. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, T.M.; O’Connell, J.; O’Brien, D.I.; Goode, T.; Bredin, C.P.; Shanahan, F. The role of substance P in inflammatory disease. J. Cell. Physiol. 2004, 201, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.J.; Undem, B.J. Inflammation-induced plasticity of the afferent innervation of the airways. Environ. Health Perspect. 2001, 109, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Lourenssen, S.; Wells, R.W.; Blennerhassett, M.G. Differential responses of intrinsic and extrinsic innervation of smooth muscle cells in rat colitis. Exp. Neurol. 2005, 195, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; McGregor, G.P.; Saria, A.; Philippin, B.; Kummer, W. Induction of tachykinin gene and peptide expression in guinea pig nodose primary afferent neurons by allergic airway inflammation. J. Clin. Investig. 1996, 98, 2284–2291. [Google Scholar] [CrossRef] [PubMed]
- Galeazza, M.T.; Garry, M.G.; Yost, H.J.; Strait, K.A.; Hargreaves, K.M.; Seybold, V.S. Plasticity in the synthesis and storage of substance P and calcitonin gene-related peptide in primary afferent neurons during peripheral inflammation. Neuroscience 1995, 66, 443–458. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Tracey, K.J. Neural regulation of immunity: Molecular mechanisms and clinical translation. Nat. Neurosci. 2017, 20, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, G.; Mietelska-Porowska, A.; Mazurkiewicz, M. The cholinergic system, nerve growth factor and the cytoskeleton. Behav. Brain Res. 2011, 221, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Hazari, M.S.; Pan, J.H.; Myers, A.C. Nerve growth factor acutely potentiates synaptic transmission in vitro and induces dendritic growth in vivo on adult neurons in airway parasympathetic ganglia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Weigand, L.A.; Kwong, K.; Myers, A.C. The effects of nerve growth factor on nicotinic synaptic transmission in mouse airway parasympathetic neurons. Am. J. Respir. Cell Mol. Biol. 2015, 53, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Aloe, L.; Levi-Montalcini, R. Mast cells increase in tissues of neonatal rats injected with the nerve growth factor. Brain Res. 1977, 133, 358–366. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Dahinden, C.A. Effect of nerve growth factor on the release of inflammatory mediators by mature human basophils. Blood 1992, 79, 2662–2669. [Google Scholar] [PubMed]
- Miura, K.; Saini, S.S.; Gauvreau, G.; MacGlashan, D.W., Jr. Differences in functional consequences and signal transduction induced by IL-3, IL-5, and nerve growth factor in human basophils. J. Immunol. 2001, 167, 2282–2291. [Google Scholar] [CrossRef] [PubMed]
- Takafuji, S.; Bischoff, S.C.; de Weck, A.L.; Dahinden, C.A. Opposing effects of tumor necrosis factor-a and nerve growth factor upon leukotriene C4 production by human eosinophils triggered with N-formyl-methionyl-leuyl-phenylalanine. Eur. J. Immunol. 1992, 22, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Sin, A.Z.; Roche, E.M.; Togias, A.; Lichtenstein, L.M.; Schroeder, J.T. Nerve growth factor or IL-3 induces more IL-13 production from basophils of allergic subjects than from basophils of nonallergic subjects. J. Allergy Clin. Immunol. 2001, 108, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, B.F.; Zillikens, D.; Grabbe, J. Nerve growth factor influences IgE-mediated human basophil activation: Functional properties and intracellular mechanisms compared with IL-3. Int. Immunopharmacol. 2005, 5, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Welker, P.; Grabbe, J.; Grützkau, A.; Henz, B.M. Effects of nerve growth factor (NGF) and other fibroblast-derived growth factors on immature human mast cells (HMC-1). Immunology 1998, 94, 310–317. [Google Scholar] [PubMed]
- Murakami, M.; Tada, K.; Nakajima, K.; Kudo, I. Cyclooxygenase-2-dependent delayed prostaglandin D2 generation is initiated by nerve growth factor in rat peritoneal mast cells: Its augmentation by extracellular type II secretory phospholipase A2. J. Immunol. 1997, 159, 439–446. [Google Scholar] [PubMed]
- Marshall, J.S.; Gomi, K.; Blennerhassett, M.G.; Bienenstock, J. Nerve growth factor modifies the expression of inflammatory cytokines by mast cells via a prostanoid-dependent mechanism. J. Immunol. 1999, 162, 4271–4276. [Google Scholar] [PubMed]
- Mazurek, N.; Weskamp, G.; Erne, P.; Otten, U. Nerve growth factor induces mast cell degranulation without changing intracellular calcium levels. FEBS Lett. 1986, 198, 315–320. [Google Scholar] [CrossRef]
- Horigome, K.; Pryor, J.C.; Bullock, E.D.; Johnson, E.M., Jr. Mediator release from mast cells by nerve growth factor. Neurotrophin specificity and receptor mediation. J. Biol. Chem. 1993, 268, 14881–14887. [Google Scholar] [PubMed]
- Sawada, J.; Itakura, A.; Tanaka, A.; Furusaka, T.; Matsuda, H. Nerve growth factor functions as a chemoattractant for mast cells through both mitogen-activated protein kinase and phosphatidylinositol 3-kinase signalling pathways. Blood 2000, 95, 2052–2058. [Google Scholar] [PubMed]
- Kawamoto, K.; Okada, T.; Kannan, Y.; Ushio, H.; Matsumoto, M.; Matsuda, H. Nerve growth factor prevents apoptosis of rat peritoneal mast cells through the Trk proto-oncogene receptor. Blood 1995, 86, 4638–4644. [Google Scholar] [PubMed]
- Bullock, E.D.; Johnson, E.M. Nerve growth factor induces the expression of certain cytokines genes and Bcl2 in mast cells: Potential role in survival promotion. J. Biol. Chem. 1996, 271, 27500–27508. [Google Scholar] [CrossRef] [PubMed]
- Hamada, A.; Watanabe, N.; Ohtomo, H.; Matsuda, H. Nerve growth factor enhances survival and cytotoxic activity of human eosinophils. Br. J. Haematol. 1996, 93, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Kannan, Y.; Usami, K.; Okada, M.; Shimizu, S.; Matsuda, H. Nerve growth factor suppresses apoptosis of murine neutrophils. Biochem. Biophys. Res. Commun. 1992, 186, 1050–1056. [Google Scholar] [CrossRef]
- Kannan, Y.; Ushio, H.; Koyama, H.; Okada, M.; Oikawa, M.; Yoshihara, T.; Kaneko, M.; Matsuda, H. Nerve growth factor enhances survival, phagocytosis, and superoxide production of murine neutrophils. Blood 1991, 77, 1320–1325. [Google Scholar] [PubMed]
- Hepburn, L.; Prajsnar, T.K.; Klapholz, C.; Moreno, P.; Loynes, C.A.; Ogryzko, N.V.; Brown, K.; Schiebler, M.; Hegyi, K.; Antrobus, R.; et al. Innate immunity. A Spaetzle-like role for nerve growth factor β in vertebrate immunity to Staphylococcus aureus. Science 2014, 346, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, L.W.; Stach, R.W.; Hashim, G.A.; Marchetti, D.; Perez-Polo, J.R. Receptors for nerve growth factor on rat spleen mononuclear cells. J. Neurosci. Res. 1987, 17, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Brodie, C.; Gelfand, E.W. Functional nerve growth factor receptors on human B lymphocytes. Interaction with IL-2. J. Immunol. 1992, 148, 3492–3497. [Google Scholar] [PubMed]
- Otten, U.; Ehrhard, P.; Peck, R. Nerve growth factor induces growth and differentiation of human B lymphocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 10059–10063. [Google Scholar] [CrossRef] [PubMed]
- Kimata, H.; Yoshida, A.; Ishioka, C.; Kusunoki, T.; Hosoi, S.; Mikawa, H. Nerve growth factor specifically induces human IgG4 production. Eur. J. Immunol. 1991, 21, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Brodie, C.; Gelfand, E.W. Regulation of immunoglobulin production by nerve growth factor: Comparison with anti-CD40. J. Neuroimmunol. 1994, 52, 87–96. [Google Scholar] [CrossRef]
- Brodie, C.; Oshiba, A.; Renz, H.; Bradley, K.; Gelfand, E.W. Nerve growth-factor and anti-CD40 provide opposite signals for the production of IgE in interleukin-4-treated lymphocytes. Eur. J. Immunol. 1996, 26, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Abram, M.; Wegmann, M.; Fokuhl, V.; Sonar, S.; Luger, E.O.; Kerzel, S.; Radbruch, A.; Renz, H.; Zemlin, M. Nerve growth factor and neurotrophin-3 mediate survival of pulmonary plasma cells during the allergic airway inflammation. J. Immunol. 2009, 182, 4705–4712. [Google Scholar] [CrossRef] [PubMed]
- Bayas, A.; Kruse, N.; Moriabadi, N.F.; Weber, F.; Hummel, V.; Wohleben, G.; Gold, R.; Toyka, K.V.; Rieckmann, P. Modulation of cytokine mRNA expression by brain-derived neurotrophic factor and nerve growth factor in human immune cells. Neurosci. Lett. 2003, 335, 155–158. [Google Scholar] [CrossRef]
- Kobayashi, H.; Mizisin, A.P. Nerve growth factor and neurotrophin-3 promote chemotaxis of mouse macrophages in vitro. Neurosci. Lett. 2001, 305, 157–160. [Google Scholar] [CrossRef]
- Samah, B.; Porcheray, F.; Gras, G. Neurotrophins modulate monocyte chemotaxis without affecting macrophage function. Clin. Exp. Immunol. 2008, 151, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Susaki, Y.; Shimizu, S.; Katakura, K.; Watanabe, N.; Kawamoto, K.; Matsumoto, M.; Tsudzuki, M.; Furusaka, T.; Kitamura, Y.; Matsuda, H. Functional properties of murine macrophages promoted by nerve growth factor. Blood 1996, 88, 4630–4637. [Google Scholar] [PubMed]
- Barouch, R.; Kazimirsky, G.; Appel, E.; Brodie, C. Nerve growth factor regulates TNF-α production in mouse macrophages via MAPkinase activation. J. Leukoc. Biol. 2001, 69, 1019–1026. [Google Scholar] [PubMed]
- Noga, O.; Peiser, M.; Altenähr, M.; Knieling, H.; Wanner, R.; Hanf, G.; Grosse, R.; Suttorp, N. Differential activation of dendritic cells by nerve growth factor and brain-derived neurotrophic factor. Clin. Exp. Allergy 2007, 37, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Stenfors, C.; Tirassa, P.; Theodorsson, E.; Lundberg, T. Nerve growth factor stimulates production of neuropeptide Y in human lymphocytes. NeuroReport 1996, 7, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Caroleo, M.C.; Buanne, P.; Costa, N.; Starace, G.; Lundeberg, T. Endogenous NGF regulates CGRP expression in human monocytes, and affects HLA-DR and CD86 expression and IL-10 production. Blood 2005, 106, 3507–3514. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Buanne, P.; Finn, A.; Stenfors, C.; Vigneti, E.; Theodorsson, E.; Lundeberg, T. NGF modulates CGRP synthesis in human B-lymphocytes: A possible anti-inflammatory action of NGF? J. Neuroimmunol. 2002, 123, 58–65. [Google Scholar] [CrossRef]
- Beigelman, A.; Levy, J.; Hadad, N.; Pinsk, V.; Haim, A.; Fruchtman, Y.; Levy, R. Abnormal neutrophil chemotactic activity in children with congenital insensitivity to pain with anhidrosis (CIPA): The role of nerve growth factor. Clin. Immunol. 2009, 130, 365–372. [Google Scholar]
- Safieh-Garabedian, B.; Poole, S.; Allchorne, A.; Winter, J.; Woolf, C.J. Contribution of interleukin-1β to the inflammation-induced increase in nerve growth factor levels and inflammatory hyperalgesia. Br. J. Pharmacol. 1995, 7, 1265–1275. [Google Scholar] [CrossRef]
- Aloe, L.; Tuveri, M.A.; Levi-Montalcini, R. Studies on carrageenan-induced arthritis in adult rats: Presence of nerve growth factor and role of sympathetic innervation. Rheumatol. Int. 1992, 12, 213–216. [Google Scholar] [CrossRef] [PubMed]
- De Vries, A.; Dessing, M.C.; Engels, F.; Henricks, P.A.; Nijkamp, F.P. Nerve growth factor induces a neurokinin-1 receptor-mediated airway hyperresponsiveness in guinea pigs. Am. J. Respir. Crit. Care Med. 1999, 159, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Rukwied, R.; Mayer, A.; Kluschina, O.; Obreja, O.; Schley, M.; Schmelz, M. NGF induces non-inflammatory localized and lasting mechanical and thermal hypersensitivity in human skin. Pain 2010, 148, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Villoslada, P.; Hauser, S.L.; Bartke, I.; Unger, J.; Heald, N.; Rosenberg, D.; Cheung, S.W.; Mobley, W.C.; Fisher, S.; Genain, C.P. Human nerve growth factor protects common marmosets against autoimmune encephalomyelitis by switching the balance of T helper cell type 1 and 2 cytokines within the central nervous system. J. Exp. Med. 2000, 191, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Flügel, A.; Matsumuro, K.; Neumann, H.; Klinkert, W.E.; Birnbacher, R.; Lassmann, H.; Otten, U.; Wekerle, H. Anti-inflammatory activity of nerve growth factor in experimental autoimmune encephalomyelitis: Inhibition of monocyte transendothelial migration. Eur. J. Immunol. 2001, 31, 11–22. [Google Scholar] [CrossRef]
- Arredondo, L.R.; Deng, C.; Ratts, R.B.; Lovett-Racke, A.E.; Holtzman, D.M.; Racke, M.K. Role of nerve growth factor in experimental autoimmune encephalomyelitis. Eur. J. Immunol. 2001, 31, 625–633. [Google Scholar] [CrossRef]
- Micera, A.; Properzi, F.; Triaca, V.; Aloe, L. Nerve growth factor antibody exacerbates neuropathological signs of experimental allergic encephalomyelitis in adult lewis rats. J. Neuroimmunol. 2000, 104, 116–123. [Google Scholar] [CrossRef]
- Barada, K.A.; Mourad, F.H.; Sawah, S.I.; Khoury, C.; Safieh-Garabedian, B.; Nassar, C.F.; Tawil, A.; Jurjus, A.; Saadé, N.E. Up-regulation of nerve growth factor and interleukin-10 in inflamed and non inflamed intestinal segments in rats with experimental colitis. Cytokine 2007, 37, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Reinshagen, M.; Rohm, H.; Steinkamp, M.; Lieb, K.; Geerling, I.; Von Herbay, A.; Flämig, G.; Eysselein, V.E.; Adler, G. Protective role of neurotrophins in experimental inflammation of the rat gut. Gastroenterology 2000, 119, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ulevitch, R.J. Limiting inflammatory responses during activation of innate immunity. Nat. Immunol. 2005, 6, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y.; Xu, D.; Brint, E.K.; O’Neill, L.A. Negative regulation of Toll-like receptor-mediated immune responses. Nat. Rev. Immunol. 2005, 5, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Michalek, S.M.; Jope, R.S. Innate and adaptive immune responses regulated by glycogen synthase kinase-3 (GSK3). Trends Immunol. 2010, 31, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Koyasu, S. PI3K and negative regulation of TLR signaling. Trends Immunol. 2003, 24, 358–363. [Google Scholar] [CrossRef]
Figure 1.
Direct and indirect effects of nerve growth factor (NGF) on inflammatory responses. At the site of inflammation, inflammatory cytokines induce (blue arrows) the production of NGF in different cell types, such as muscle cells, epithelial cells, fibroblasts, adipocytes, neurons, glia, and immune cells. The enhanced local production of NGF influences nerve fiber distribution and neuronal activity, inducing the synthesis and release of neuropeptides and neurotransmitters that have immunomodulatory effects. NGF receptors are also expressed on the membrane of immune cells and NGF can directly modulate the activity and functions of immune cells.
Figure 1.
Direct and indirect effects of nerve growth factor (NGF) on inflammatory responses. At the site of inflammation, inflammatory cytokines induce (blue arrows) the production of NGF in different cell types, such as muscle cells, epithelial cells, fibroblasts, adipocytes, neurons, glia, and immune cells. The enhanced local production of NGF influences nerve fiber distribution and neuronal activity, inducing the synthesis and release of neuropeptides and neurotransmitters that have immunomodulatory effects. NGF receptors are also expressed on the membrane of immune cells and NGF can directly modulate the activity and functions of immune cells.
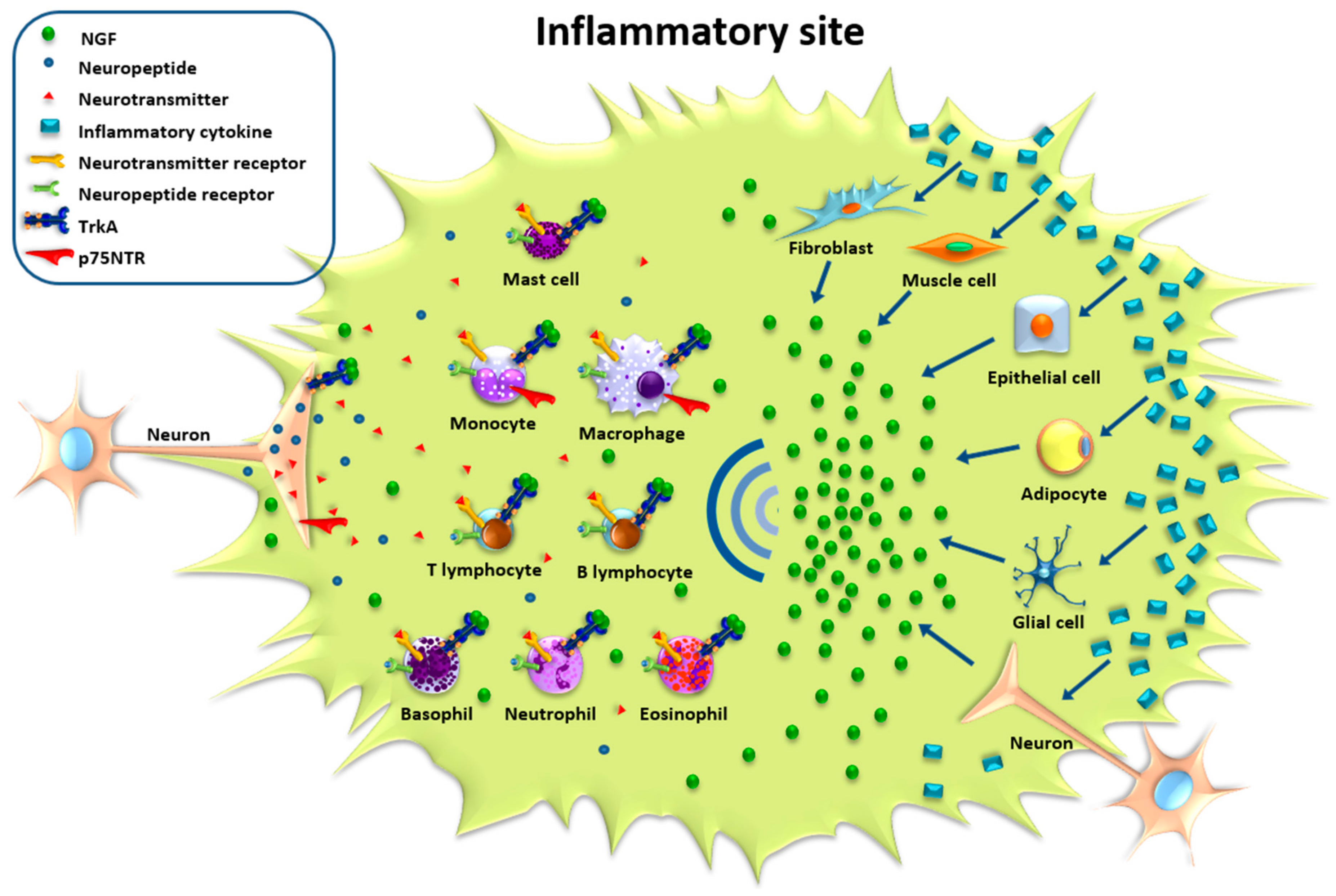
Figure 2.
TrkA activation promotes anti-inflammatory pathways. In human monocytes the expression of TrkA is enhanced when TLR4 is activated. Auto-phosphorylation of TrkA induced by NGF binding activates intracellular pathways that influence the downstream signaling of TLR4. The NGF-induced phosphorylation of Akt inhibits NF-κB translocation in the nucleus. The inhibitory phosphorylation of GSK3 induced by Akt further prevents NF-κB activation, and the NF-κB-dependent transcription of pro-inflammatory cytokine genes. Concomitantly, NGF activation of the PI3K/Akt pathway induces the expression of IL-10 and IL-1 receptor antagonist (IL-1ra), promoting a net anti-inflammatory effect. Black bold arrow shows dimerization of NGF and binding to TrkA. Red and green bold arrows show respectively decrease and increase of cytokine levels. Black slim arrows and black T bar show respectively activation and inhibition of molecular pathways. Dotted line arrow show nuclear translocation.
Figure 2.
TrkA activation promotes anti-inflammatory pathways. In human monocytes the expression of TrkA is enhanced when TLR4 is activated. Auto-phosphorylation of TrkA induced by NGF binding activates intracellular pathways that influence the downstream signaling of TLR4. The NGF-induced phosphorylation of Akt inhibits NF-κB translocation in the nucleus. The inhibitory phosphorylation of GSK3 induced by Akt further prevents NF-κB activation, and the NF-κB-dependent transcription of pro-inflammatory cytokine genes. Concomitantly, NGF activation of the PI3K/Akt pathway induces the expression of IL-10 and IL-1 receptor antagonist (IL-1ra), promoting a net anti-inflammatory effect. Black bold arrow shows dimerization of NGF and binding to TrkA. Red and green bold arrows show respectively decrease and increase of cytokine levels. Black slim arrows and black T bar show respectively activation and inhibition of molecular pathways. Dotted line arrow show nuclear translocation.
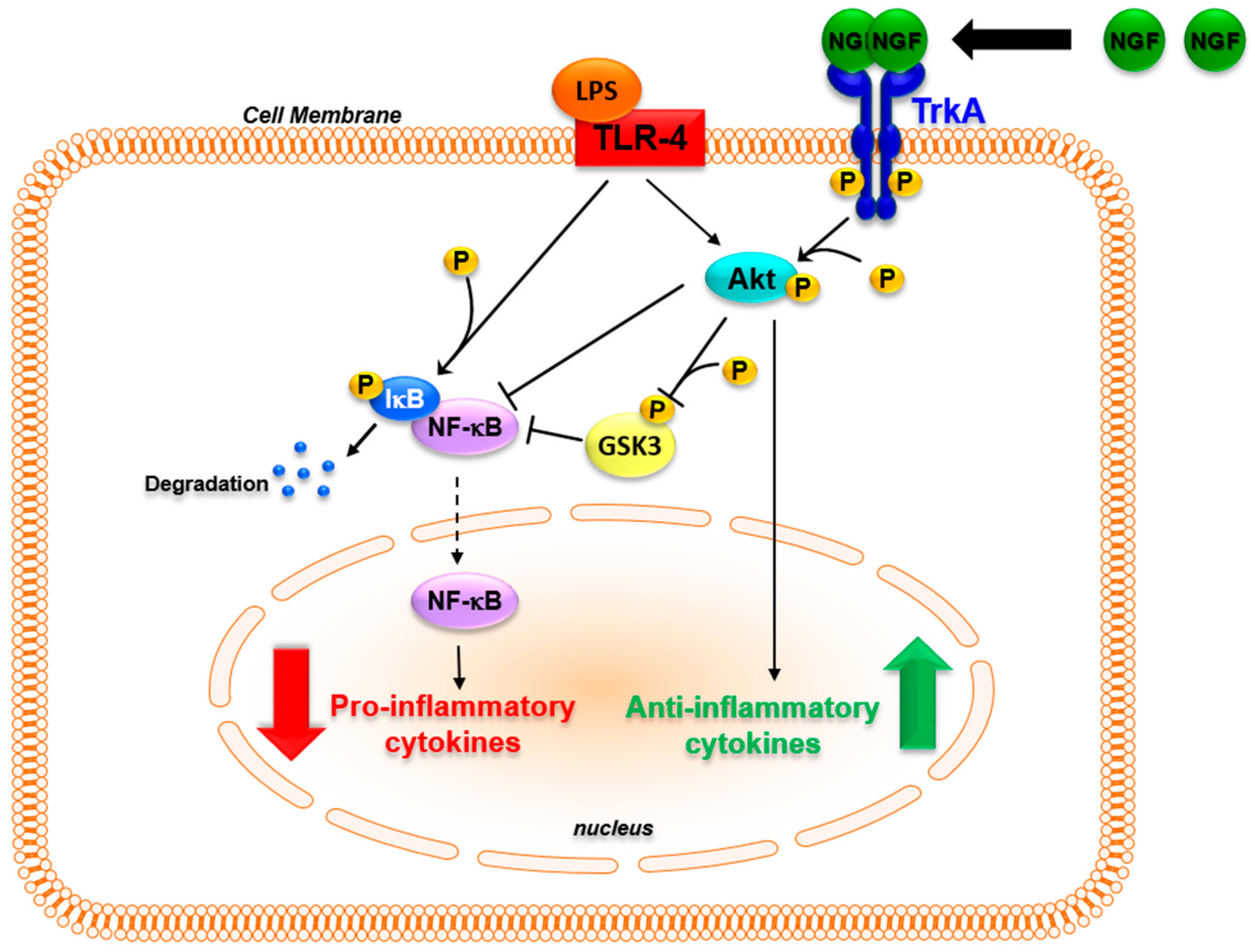

{kind=link}
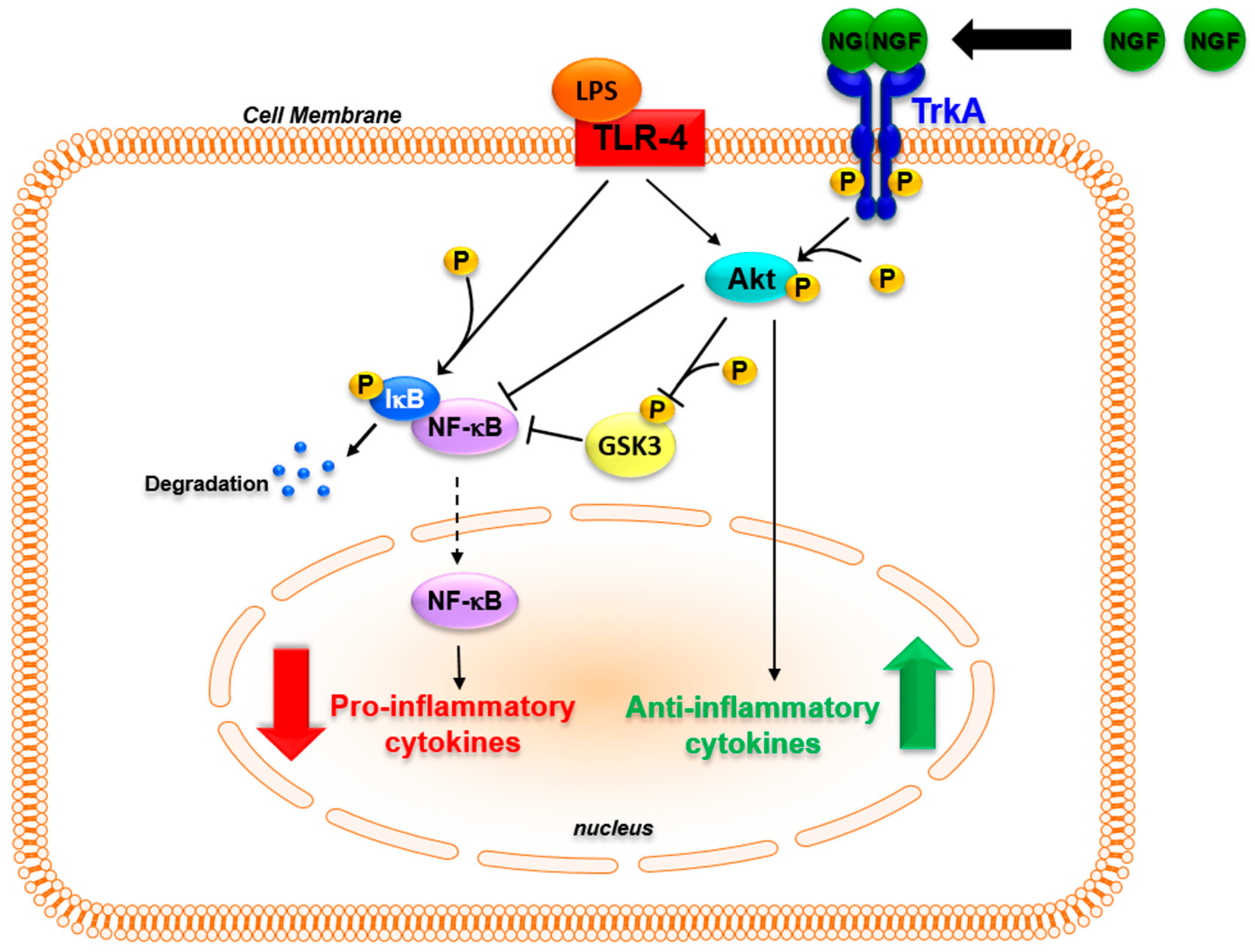{kind=link}
Table 1.
Distribution of NGF in primary and secondary lymphoid organs.
| NGF Receptor Distribution in Lymphoid Organs | |||
|---|---|---|---|
| Tissue | TrkA | p75NTR | |
| Primary Lymphoid Organs | Thymus | thymocytes [77,78,79] | interdigitating reticular cells of the medulla [79]; |
| epithelial subcapsular and medullar cells [73,75,80,81] | periarteriolar macrophages [79,80,81,82]; | ||
| peripheral epithelial cells of Hassal’s bodies [79] | endothelial sinusal cells and nerve endings [72,79,80,81,82]; | ||
| interdigitating reticular cells of the medulla [79] | - | ||
| Bone marrow | stromal cells with dendritic features [77,84] | stromal cells with dendritic features [77,84]; | |
| CD34 positive hemopoietic stem cells [85,86,87] | CD34 positive hemopoietic stem cells [85,86,87]; | ||
| Bursa of fabricius | epithelial cells of the follicle [90,91] | bursa of Fabricius of chick embryo [88]; | |
| interfollicular epithelium [90,91] | |||
| blood vessels [90,91] | |||
| Secondary Lymphoid Organs | Spleen | stroma of the spleen and splenocytes [78] | stroma of the spleen and splenocytes [78]; |
| spleen mononuclear immunocompetent cells [77] | spleen mononuclear immunocompetent cells [77]; | ||
| Lymph nodes and mucosa-associated lymphoid tissues | follicular dendritic cells [79,94,95] | follicular dendritic cells [79,94,95]; | |
| blood vessel walls [79,94,95] | blood vessel walls [79,94,95]; | ||
| cryptic tonsillar epithelium [79,94,95] | cryptic tonsillar epithelium [79,94,95]; | ||
| monocyte-derived cells [79,94,95] | monocyte-derived cells [79,94,95]; | ||
| interdigitated reticular cells [79,94,95] | interdigitated reticular cells [79,94,95]. | ||
Table 2.
Multiple effects of NGF on purified immune cells.
| Effect of NGF Stimulation | |
|---|---|
| Activated basophils | ↑ leukotriene and cytokine synthesis [110,138,139,140] |
| ↑ histamine release [141] | |
| ↑ response to IgE [141,142] | |
| Immature mast cells | ↑ tryptase and IgE receptors [143] |
| Mature mast cells | ↑ cyclooxygenase2 (COX2) and prostaglandin D2 [144] |
| ↑ IL-6 induction [145] | |
| ↑ histamine release [146,147] | |
| ↑ chemotaxis [148] | |
| ↑ survival (by suppressing apoptosis) [149,150] | |
| Eosinophils | ↓ suppression of leukotriene formation [140] |
| ↑ IL-4 production [111] | |
| ↑ peroxidase release and cytotoxic activity [151] | |
| ↑ survival (by suppressing apoptosis) [151] | |
| Neutrophils | ↑ survival (by suppressing apoptosis) [152] |
| ↑ superoxide production and phagocytosis [153,154] | |
| B-cells | ↑ proliferative response [155,156] |
| ↑ IL-2 receptors [156] | |
| influences the production of IgM and IgG [157,158,159,160] | |
| ↑ survival of memory B-cells [107] | |
| ↑differentiation of B-cells into immunoglobulin-secreting plasma cells [157] | |
| influences plasma cell survival [161] | |
| T-cells | ↑ proliferative response [155] |
| ↑ cytokine expression [162] | |
| Monocytes/macrophages | protection from apoptosis, by inducing the anti-apoptotic proteins Bcl-2, Bcl-xl and Bfl-1 [118] |
| ↑ CXCR4 expression and chemotactic response [163,164] | |
| ↑ phagocytosis, enhanced parasite-killing activity and IL-1β [165] | |
| ↑ TNF-α, IL-8 secretion [154,166] | |
| Dendritic cells | ↑ maturation of dendritic cells and secretion of inflammatory cytokines [167] |
| ↑ IL-6 release in allergic patients; | |
| ↑ IL-10 release in healthy controls [167] | |
Footnote: ↑ = increase; ↓ = decrease.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Minnone, G.; De Benedetti, F.; Bracci-Laudiero, L. NGF and Its Receptors in the Regulation of Inflammatory Response. Int. J. Mol. Sci. 2017, 18, 1028. https://doi.org/10.3390/ijms18051028
AMA Style
Minnone G, De Benedetti F, Bracci-Laudiero L. NGF and Its Receptors in the Regulation of Inflammatory Response. International Journal of Molecular Sciences. 2017; 18(5):1028. https://doi.org/10.3390/ijms18051028
Chicago/Turabian StyleMinnone, Gaetana, Fabrizio De Benedetti, and Luisa Bracci-Laudiero. 2017. "NGF and Its Receptors in the Regulation of Inflammatory Response" International Journal of Molecular Sciences 18, no. 5: 1028. https://doi.org/10.3390/ijms18051028
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.