
Regulatory miRNAs in Colorectal Carcinogenesis and Metastasis


1
Department of Pathology and Institute of Precision Medicine, Jining Medical University, 16 Hehua Road, Jining 272067, China
2
Department of Pathology, University of Illinois at Chicago, Chicago, IL 60612, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(4), 890; https://doi.org/10.3390/ijms18040890
Submission received: 30 March 2017
/
Revised: 15 April 2017
/
Accepted: 20 April 2017
/
Published: 22 April 2017
(This article belongs to the Collection Regulation by Non-coding RNAs)
Abstract
:Colorectal cancer is one of the most common malignancies and is the second-leading cause of cancer-related death world-wide, which is linked to genetic mutations, epigenetic alterations, and oncogenic signaling activation. MicroRNAs, one of the categories of epigenetics, have been demonstrated significant roles in carcinogenesis and progression through regulating of oncogenic signaling pathways, stem cells, epithelial-mesenchymal transition, and metastasis. This review summarizes the roles of microRNAs in the regulating of Wnt, Ras, TGF-β, and inflammatory signaling pathways, stemness, and epithelial-mesenchymal transition, for carcinogenesis and metastasis in colorectal cancer. Improving our understanding of the mechanisms of regulatory interactions of microRNAs with signaling pathways in colorectal cancer formation and progression will aid in determining the genes responsible for colorectal cancer initiation, progression, metastasis, and recurrence and, finally, in developing personalized approaches for cancer prevention and therapy.
{kind=link}
{kind=link}
1. Introduction
Colorectal cancer (CRC) is one of the most common malignancies and is the second-leading cause of cancer-related death worldwide [1]. Although several decades of effort have been made, the underlying mechanisms are still largely unknown. However, more and more evidence has demonstrated that colorectal carcinogenesis is linked to the activation of oncogenic signaling pathways and inactivation of tumor-suppressive signaling, resulting mostly from genetic mutations and epigenetic alterations, the latter including DNA methylation, histone acetylation, and non-coding RNAs (e.g., microRNAs, long non-coding RNAs, etc.). Among them, microRNAs (miRNAs) have shown critical biological functions according to the increasing evidence from clinical epidemiology and experimental studies [2,3,4,5,6,7]. Interestingly, these miRNAs are differentially expressed in colorectal cancer tissues, serum and plasma, and body fluids and, therefore, some miRNAs have been potentially used as biomarkers for diagnosis and therapeutic targets, exhibiting clinical importance [8,9,10,11,12]. Moreover, molecular mechanistic studies have revealed that these miRNAs participate in carcinogenesis and progression. In colorectal cancers, miRNAs have shown involvement in, or directly regulating, oncogenic signaling pathways, such as Wnt, Ras, TGF-β, and NF-κB/AKT/STAT3 signaling pathways [2,3,9,10]. In addition, these miRNAs are also involved in the regulating of the stemness of cancer stem cells, epithelial-mesenchymal transition (EMT), and metastasis [13,14,15]. Based on biological functions and regulatory interaction, we named these groups of miRNAs as regulatory miRNAs. Herein, we reviewed the regulatory miRNAs in the regulating of oncogenic signaling pathways in colorectal cancer formation and metastasis.
2. Clinical Significance of miRNAs in Colorectal Cancer
The studies from animal models and clinical epidemiology have shown differential expression of miRNAs in CRC or even pre-cancerous lesions. Using a colorectal cancer mouse model Muc2 knockout mice and miRNA array analysis on colonic epithelial cells, we have found that miRNAs were differentially expressed in mouse colonic epithelial cells [16], including 20 downregulated and 71 upregulated miRNAs. To confirm the accuracy, we chose the 15 most-changed miRNAs (six upregulated and nine downregulated) for validation by quantitative RT-PCR in mouse intestinal epithelial cells and, further, the mostly-changed miRNAs were validated in human colonoscopy biopsies of colitis and colorectal cancers. Bioinformatic analysis and functional studies have shown that these miRNAs mostly target the genes associated with cytokines and chemokines that are linked to inflammatory pathways and colitis-associatedcolorectal cancer [16]. Increasing studies have demonstrated the differential levels of miRNAs in human colorectal cancer tissues and serum, some are downregulated, exhibiting tumor suppressive functions, and some are upregulated, exhibiting oncogenic functions. To date, about 50 miRNAs have been found up- or downregulated in CRC cells as compared to non-tumor normal cell [12]. Some of miRNAs, such as miR-106, miR-31, miR-21, miR-25, miR-20a, miR-93, miR-183, and miR-203, are upregulated in CRC, but miR-1, miR-126, miR-30a, miR-143, miR-145, miR-191, and miR-192 are downregulated in CRC, whereas some miRNAs are also upregulated in one report and downregulated in another report in CRC, showing controversial functions. For example, miR-27a was found downregulated and showed tumor-suppressive functions in CRC, targeting Stat3 and Smad2 [16], and in other cancers [17,18] it was also found upregulated and showed oncogenic functions in CRC [19]. Moreover, cancer cells release miRNA into peripheral blood [20]. Therefore, these miRNAs could be detected from serum and plasma, andcould be used as biomarkers for diagnosis. Interestingly, the circulating miRNAs in blood have been recently found to be covered into a complexes, known as exosomes, which are protected from RNase degradation and become more stable [21].As well as detection in peripheral blood, CRC-specific miRNA could also be detected in stool. The study from Link et al. has shown that higher levels of miR-21 and miR-106a were detected in the feces of CRC and colorectal adenoma patients compared with healthy controls [22].In addition, the plasma levels of miRNAs could be used as biomarkers monitoring cancer progression or treatment. For instance, plasma levels of miR-17-3p and miR-92a were found to be reduced after surgical removal of colon cancers [22].
Therapeutic roles of miRNAs have been proven in in vitro and in vivoexperiments. As addressed above, miRNAs are aberrantly expressed in colorectal cancer tissues and cancer cell lines. Therefore, tumor suppressive miRNAs, or their mimics, can beused as novel agents for targeted therapy; in contrast, the oncogenic miRNAs can be used as targets for personalized therapy, or correcting of the aberrant expression of miRNAs by either blocking or restoring miRNA levels and functions as therapeutic strategiesfor CRC treatment. Several lines of evidence have shown that anti-cancer miRNA mimics could inhibit CRC cancer cell proliferation and migration, induce cancer cell apoptosis in vitro, and inhibit cancer cell growth in nude mice (e.g., miRNA-27a showed anti-cancer functions in colorectal cancer cells, and miR-27a mimics could inhibit CRC cell proliferation and tumor growth in nude mice [23]); restoration of miR-195 in CRC cell lines reduced cell viability, promoted cell apoptosis, and suppressed tumorigenesis [24]; and miR-342 could induce colon cancer cell cycle arrest at G0/G1 phase [25].Recent studies have also shown that some miRNAs (e.g., miR-137, miR-139-5p, miR-143, miR-409-3p, miR-494, etc.) enhanced CRC chemo-sensitivity, but some miRNAs (e.g., miR-587, miR-133a, miR-492, miR-192, miR-215, etc.) are associated with chemo-resistance in colorectal cancers [26].Furthermore, studies have also demonstrated that several miRNAs (e.g., miR-21, miR-200, miR-215, miR-143) have shown prognostic potential in colorectal cancers [27,28]. For example, miR-143 expression has been identified as an independent predictor of patient survival. Colorectal cancer patients with low levels of miR-143 expression have a significantly higher risk of having shorter cancer-specific survival and progression-free survival [29]. Taken above, miRNAs have critical clinical significance on colorectal cancer diagnosis, targeted therapy, and outcome prediction.
3. Regulatory miRNAs in Oncogenic Signaling Pathways
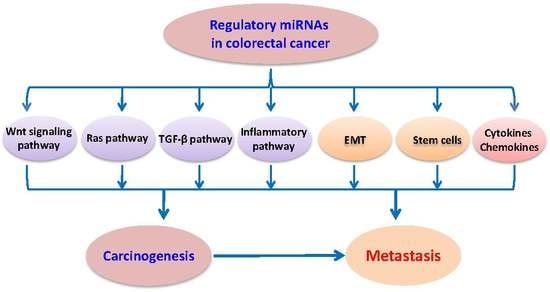
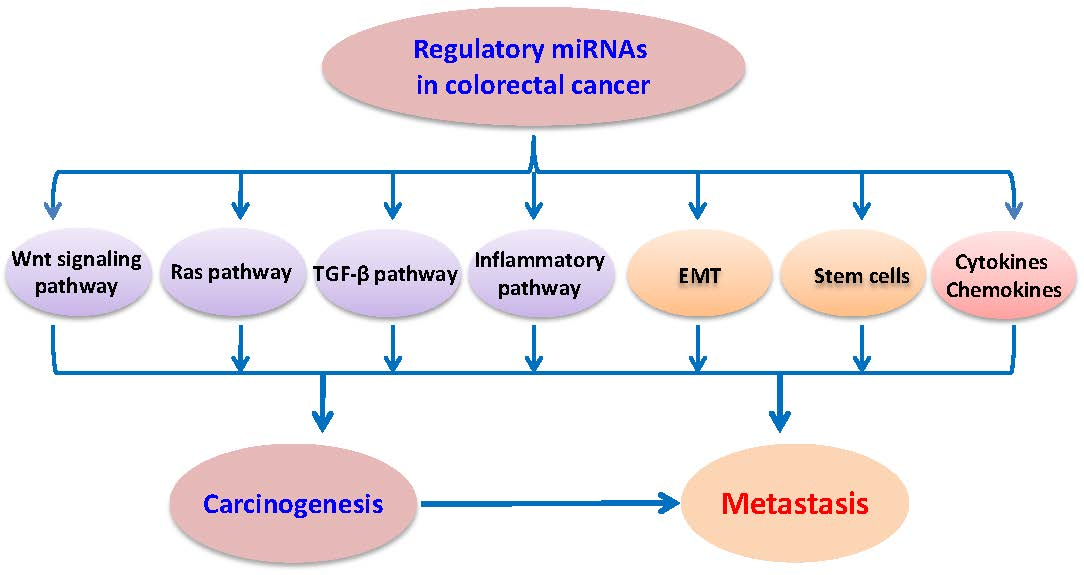
Colorectal cancers are mainly caused by the activation of driven genes in the oncogenic signaling pathways, such as Wnt, Ras, TGF-β, and inflammatory signaling pathways, and these signaling pathways are regulated by individual miRNA or a clusters/groups of miRNAs (Figure 1).
3.1. miRNAs and Wnt Signaling
The deregulation of Wnt signaling is one of the most frequently-changed events in colorectal cancer, resulting from APC (adenomatosis polyposis coli) gene mutations that have been observed in about 75% of sporadic CRC [30,31,32]. APC, AXIN, and GSK-3β form a destruction complex, and this complex is essential for the phosphorylation and degradation of β-catenin. APC mutation and Wnt ligands present could cause the loss of the destroying function and cytoplastic accumulation of β-catenin, a key effector of Wnt signaling, resulting in β-catenin nuclear translocation and interacting with TCF4 (transcription factor 4), and activating the transcription of target genes, such as cyclin D1 and c-myc.
Recent studies have revealed that miRNAs can also regulate the Wnt signaling pathway by targeting the key elements of the Wnt pathway. For example, miR-135a/b is overexpressed in CRC and is able to directly target APC, leading to the repression of APC expression and to the upregulation of Wnt signaling [33]. miR-135a/b is also predicted to target and inhibit secreted frizzled-related protein 4 (SFRP4), the latter is a Wnt/β-catenin inhibitor through binding and repressing extracellular Wnt proteins [34].On the other hand, miR-135b can be transcriptionally activated by β-catenin/TCF4, which shows that miR-135b is significantly upregulated in human CRC and in the tumors of an Apc mutation mouse model [35]. Unlike miR-135, miR-21 can increase β-catenin nuclear translocation and promote tumorigenesis in colorectal cancer [36]. Vice versa, miRNA-21 can also target the β-catenin signaling pathway and enhance Wnt-driven epithelial carcinogenesis [37]. Like miR-21, miR-155 is also a Wnt/β-catenin stimulator through targeting the Wnt signaling inhibitor HMGB1 and indirectly increasing Wnt/β-catenin expression [38,39].
Another group of miRNAs have tumor suppressive properties and can directly target and repress Wnt signaling. For example, the miR-34 family (miR-34a/b/c) can directly target Wnt ligands WNT1, WNT3, and LRP6, and β-catenin and LEF1, transcription factors that interact with β-catenin [40]. In addition, miR-29b, miR-29c, and miR-93 can inhibit Wnt ligands and β-catenin-mediated functions, e.g., miR-29c targets GNA13 and PTP4A that are negative regulators of GSK3β, a kinase that phosphorylates β-catenin and triggers its degradation; miR-93 targets SMAD7, which promotes nuclear accumulation of β-catenin [41,42].
3.2. miRNAs and RAS Pathway
The RAS gene family is well characterized and plays important roles in regulating cell proliferation, apoptosis, differentiation, and migration and, therefore, acts as an oncogene [43]. There are three human isoforms, NRAS, HRAS, and KRAS, and KRAS is frequently mutated in colorectal cancers with 30%–40% mutation rates, moreover, KRAS mutations have been shown to be well-associated with poorer outcomes, in terms of shorter survival times, and being more aggressive and drug-resistant [44,45,46].
Since the RAS genes have several miRNA let-7 binding sites at the 3′-UTR, let-7 targets and regulates KRAS gene expression, and the reduction of let-7 in cancer tissues is correlated with higher KRAS mRNA expression, suggesting the regulatory roles of let-7 miRNAs in KRAS [47,48,49]. Indeed, the in vitro study showed that let-7 miRNA suppressed colon cancer growth and proliferation [50], in contrast, transfection with let-7a precursor miRNA significantly inhibited cancer cell growth and reduced the expression of KRAS and c-MYC [50].
miR-143 has been shown downregulation in colorectal cancer tissues to bind to the 3′-UTR of the KRAS gene [28,51], and reduced expression of miR-143 led to cell proliferation in vitro, which is linked to the increased expression of KRAS [51], thus, miR-143, like let-7, acts as tumor suppressor in KRAS-driven colorectal carcinogenesis [29], whereas treating colorectal cancer cells with a miR-143 mimic or overexpressing miR-143 resulted in cell proliferation and downregulation of KRAS and ERK1/2 [51]. Unlike let-7 and miR-143, miR-31 has been shown to negatively regulate KRAS inhibitor RASA1; thus, miR-31 could be a potent enhancer of KRAS in colorectal cancer [52,53,54].
3.3. miRNAs and TGF-β Pathway
Transforming growth factor-β (TGF-β) is a multitasking cytokine and TGF-β signaling pathway plays important physical and pathological roles in regulating cell proliferation, differentiation, apoptosis, migration, invasion, and modification of the microenvironment and cancer metastasis [55,56,57]. TGF-β binds two distinct receptor serine/threonine kinases, the type I receptors (TβRI) and type II receptors (TβRII), and activates SMADs and non-SMADs signaling pathways [56,57,58]. Interestingly, the TGF-β signaling pathway has a paradoxical effect on cancer biology: it maintains proliferation and differentiation in normal cells and early-stage cancer cells, but promotes cancer cell invasion and metastasis in late-stage cancers [59]. The TGF-β signaling pathway can regulate, and be regulated by, a series of molecular and signaling pathways, where miRNAs have been shown to play important roles, and among them, the miR-17 family seems to have crosstalk with the TGF-β signaling pathway [60,61].
The miR-17 family has eight miRNAs, including miR-17, miR-18a/b, miR-20a/b, miR-93, and miR-106a/b, and three of them (i.e., miR-17, miR-18a, and miR-20a) are transcribed from the miR-17-92 locus.MiR-17 targets and inhibits PTEN [62] and RHOE (RND3) [63], a tumor suppressor that is downregulated in CRC and exhibits inhibition of cancer cell invasion [64].
Several lines of evidence have also shown that miR-20a, another member of the miR-17 family, promotes cancer progression by facilitating CRC cell line migration and invasion and upregulating the expression of epithelial-mesenchymal transition (EMT) markers, neutralizes the growth-repressive properties of TGF-β, and further enhances the ability of TGF-β to drive cancer cell migration, invasion, and metastasis [55,65,66]. Like miR-20a, miR-106a/b seems also to enhance EMT and metastasis by targeting TGF-β receptorTGFBR2. miR-106a is highly expressed in metastatic CRC cell lines, and promotes cancer cell migration and invasion, but miR-106b has been reported to exert stimulatory and inhibitory effects on the migration and EMT of CRC cell lines [67,68,69]. Considering theabove, the studies to date have suggested that the miR-17 family promotes CRC metastasis through interaction with TGF-β signaling, as well as other pathways that modulate EMT.
3.4. miRNAs and Inflammatory Pathway
Epidemiology and experimental studies have strongly suggested that inflammatory signaling pathways are also key drivers of CRC [5,70,71,72], and the upregulation of chemokines and cytokinesis the major characteristic of inflammation-associated colorectal cancer [73,74]. The studies from us and others have demonstrated that the increase of cytokines (e.g., COX2, NF-κB, TNFα, IL-1β, IL6, etc.), C-X-C Motif Chemokine Ligand (CXCL) family members (CXCL1, CXCL2, CXCL6, CXCL8, and CXCL12), and CXC receptors (CXCRs) could be the cause of the malignant transformation of chronic colitis, resulting from gut microbiota disorder and mucosa barrier deficiency, leading to genetic and epigenetic alterations and oncogenic signaling activation (e.g., Wnt, Ras, PI3K/AKT/STAT3, etc.) [5,16,75,76], whereas the inflammatory signaling pathway could be regulated by miRNAs.
COX2 is a prostaglandin-endoperoxide synthase and is responsible for generating PGE2, a pro-inflammatory prostaglandin that also activates the Wnt signaling pathway and is frequently overexpressed in CRC [77,78]. More studies have demonstrated that programmed cell death 4 (PDCD4) is reduced during CRC tumorigenesis, along with the transformation of normal tissue to adenocarcinoma COX2/PGE2-mediated repression of PDCD4 occurring via the induction of miR-21 [79], and the reduction of PDCD4 is well-associated with a shorter survival time of CRC patients [80]. miR-21 is one of the most prominent oncogenic miRNAs in colorectal cancer and has pro-tumorigenic properties in many other cancers [81].
miR-21 is also involved in the regulation of NF-κB and MyD88, an adapter of Toll-like receptors (TLRs) needed for NF-κB activation by TLR ligands [82,83]. In the azoxymethane (AOM)/DSS mouse model of CRC, genetic inactivation of miR-21 reduced the tumor burden and decreased the expression of pro-inflammatory cytokines, and the loss of miR-21 in tumors also increased PDCD4 expression and apoptosis, but reduced the expression of activated STAT3 and BCL2 [81]. Similar tomiR-21, miR-221 and miR-222 can also positively activate NF-κB and STAT3 by indirectly modulating their protein stability through miR-221/222-mediatedpositive feedback loops to increase expression levels of RelA and STAT3 [84].Therefore, the miR-21 family acts as a key modulator in oncogenic and inflammatory signaling pathways in which the miR-21 family maintains a positive loop with the modulation factors PDCD4, NF-κB, and STAT3.
miR-34a is a regulator of IL-6/STAT3 signaling in colorectal cancer. Rokavec et al. treated human colorectal cancer cells with cytokine IL-6and found that IL-6 activated the oncogenic STAT3 transcription factor, which directly represses miR-34a via a conserved STAT3-binding site in the first intron [85]. Repression of miR-34a was required for IL-6-induced EMT and invasion. An active IL-6R/STAT3/miR-34aloop was necessary for EMT, invasion, and metastasis of CRC cell lines and was associated with nodal and distant metastasis in CRC patients [85].
Our recent studies have also revealed tumor-suppressive functions of miRNAs (miR-138, miR-145, miR-146a, and miR-150) that were significantly reduced in colitis and colitis-associated CRC [16]. As to their regulatory functions, several studies have demonstrated that miR-138 activates the central cytokine NF-κB, promotes lipid raft formation in esophageal squamous cell carcinoma [86], and induction of miR-138 by pro-inflammatory cytokines causes endothelial cell dysfunction [87]; miR-145 targets the SOX9/ADAM17 axis to regulate tumor-initiating cells’ properties in head and neck cancers and suppresses the IL-6-mediated paracrine signaling pathway in the tumor microenvironment [88]. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) can induce miR-146a expression and the later suppresses CXCR4-mediated cancer cell migration [89]; and miR-150 interacts with cytokines and is downregulated by inflammation in cytotoxic T lymphocytes. Moreover, miR-150 acts with the miRNA network and controls perforin, eomesodermin, and IL-2Rα expression in differentiating CTLs and whose activity is modulated by IL-2, inflammation, and antigenic stimulation [90]. In addition, miR-150 plays a critical role in the development and function of invariant NKT cells via regulationof IFN-γ expression [91]. As reported by us [16], cytokines were significantly increased and miR-138, 145, 146a, and miR-150 were significantly decreased in Muc2-/- mouse colon and human colitis and colorectal cancer. Taken together, miR-138, 145, 146a, and miR-150 are well regulatorily interacted with cytokines and inflammatory factors in the development of chronic colitis and its malignant transformation.
4. Regulatory miRNAs in Cancer Stem Cells, Epithelial-Mesenchymal Transition (EMT), and Metastasis
4.1. miRNAs and Cancer Stem Cells
Emerging data have suggested that miRNAs have significant roles in regulating the function of normal cells and cancer stem cells (CSCs) through their interaction with various signaling pathways (Figure 1). The CSCs are a special population of cells that have two main properties of heterogeneity and plasticity [92], in terms of showing distinct characteristics compared to normal cancer cells, such as tumor maintenance, progression, invasion, recurrence, and chemo-resistance [92,93,94]. Therefore, CSCs are clinically important.
Six major factors are required for stem cell pluripotency maintenance. They are Nanog, Sox2, Oct4, KLF4, Lin28, and c-Myc. In addition to the regulation of CSCs by Wnt, TGF-β, Ras, and inflammatory signaling pathways, the CSCs are also regulated by miRNAs [95,96,97]. It has been observed that several miRNAs, such as miR-470, miR-296, and miR-134 may inhibit the self-renewing factors Oct4, Sox2, and Nanog [98]. In contrast, miR-145 helps in cell differentiation through targeting of KLF4, Sox2, and Oct4 [99]. Moreover, Hwang et al. have reported that miRNA-146a regulates snail-dependent symmetric division of colorectal CSCs [100], and the snail-miR-146a-β-catenin loop plays significant roles in the symmetric division of colorectal CSCs. Xu et al. have reported that expression of miR-328 was reduced in colorectal CSCs, and that increased expression of miR-328 suppressed cancer cells invasiveness and sensitized chemotherapy [101]. Therefore, miR-328 might be a potential target for CRC therapy. Bitarte et al. have found that miR-451 suppressed the self-renewal, tumor malignancy, and recurrence of colorectal CSCs [102]. Bu et al. have reported that miR-34a, like miR-451, is also a tumor suppressor and is differentially expressed in differentiating and self-renewing colon progenitor cells [103]. Moreover, the expression levels of miR-34a can regulate differentiation and self-renewalin vitroand in vivo by targeting Notch signaling. In fact, numerous studies have demonstrated that the Notch signaling pathway is an important regulator in asymmetric division and plays essential roles in promoting self-renewal of gastrointestinal stem cells and in colonic cell lineage differentiation [13,104,105]. This is also supported by experimental studies showing that colorectal CSCs might share common properties with normal colon stem cells because the CSCs in colorectal cancer xenografts exhibit similar morphology heterogeneity and histopathology as the parental tumor [106,107].
4.2. miRNAs and EMT and Metastasis
Epithelial-mesenchymal transition (EMT) is a specific physiological and/or pathological event of transformation from epithelial cells to mesenchymal cells. Numerous studies have demonstrated that EMT is an early event of cancer metastasis [108,109], in which epithelial cells lose their polarities, intercellular junctions and epithelial-like characteristics, and acquire less-differentiation and spindle-like phenotypes. EMT includes cell morphology and genotype changes; for instance, the round and less-aggressive epithelial cells transform into spindle-like and more-aggressive cells, and the cells also encounter molecular reprogramming, including the loss of cell adhesion molecules, such as E-cadherin, and gain of the expression of vimentin, N-cadherin, snail, slug, and other interstitial or stromal proteins [109,110]. Thus, EMT cells exert higher capabilities of migration and invasion and, therefore, the EMT plays critical roles during the early stages of cancer invasion and metastasis.
It is well known that the EMT is regulated by oncogenic signaling pathways, including Wnt/β-catenin, Notch/snail, Hedgehog/MMPs, TGF-β/Smad, and EGFR/MAPK/JNK signaling pathways [109,110]. Recent findings have shown that miRNA expression is also important in regulating the EMT process, such as the miR-200 family, miR-34 family, let-7, and miR-15a/miR-16-1, etc., act via the EMT signaling pathway and enhances metastasis (Figure 1).
As described above, miRNAs regulate EMT in colorectal cancer, partly by regulating the expression of tumor suppressors and oncogenes, and partly by functioning as tumor suppressors or oncogenes themselves [14,111], promoting or repressing CRC, EMT, and metastasis. The miR-200 family, including miR-200a, miR-200b, miR-200c, miR-141, and miR-429, was found to target the complementary sites in the 3-UTR of Zeb1 and Zeb2 (repressors of E-cadherin and key regulators of EMT progression), resulting indirectly in the upregulated expression of E-cadherin. EMT activator TGF-β is produced by tumor cells and can trigger the expression of Zeb1/2. Interestingly, the expression of miR-141 and miR-200c could be suppressed by Zeb1; for instance, knockdown of Zeb1 leads to increases of miR-141, miR-200c, and E-cadherin expression, increases cell-cell adhesion, induces epithelial phenotype, and reduces cell migration and invasion [112].In contrast, overexpressing Zeb1 can facilitate EMT progression and promote cancer cell invasion via triggering a miRNA-mediated feed-forward loop. Vice versa, induction of miR-200 promotes the differentiation and inhibition of epithelial-mesenchymal-specific gene expression by downregulating the expression of Zeb1 and Zeb2. Thus, the ZEB/miR-200 feedback loop is the molecular motor of cellular plasticity in the development of, and in particular is a driving force for, cancer progression towards metastasis by controlling the state of cancer stem cells [113].
The miR-34 family (miR-34a, miR-34b, and miR-34c) exerts physiological functions involved in cell cycle progression, senescence, and apoptosis. The members target snail, one of the EMT-inducing transcription factors, and leads to the downregulation of Snail expression [114,115]. Furthermore, miR-34a can suppress tumor progression by inhibiting the IL-6R/STAT3/miR-34a feedback loop and by inhibiting IL-6-induced colorectal cancer cell EMT, invasiveness, and metastasis [85].
Transcription factor AP4 is a downstream target of p53. It can directly repress E-cadherin via a non-canonical AP4-binding motif and induces N-cadherin-mediated EMT in colorectal cancer. Recent studies have shown that miR-15a/16-1 targets AP4 3'-UTR, attenuates EMT progression, and metastasis [116,117]. Recent findings have shown that miRNA let-7 is associated with cancer EMT and metastasis by targeting HMGA2 [118], and that let-7 could be repressed by Lin28, in conjunction with OCT4, SOX2, and KLF4, to promote colorectal cancer progression and metastasis [119]. Moreover, the TGF-β signaling pathway promotes cancer cell migration and invasion, which could be enhanced by miR-21 and miR-31 via suppressing TIAM1, a guanidine exchange factor of Rac GTPase and a direct target of both miR-21 and miR-31 [120].
5. Conclusions and Perspectives
As summarized in this review and in Figure 1, miRNAs represent a novel category of critical regulators in modulating tumor suppressors and oncogenes, or acting as tumor suppressors or oncogenes themselves, in colorectal cancer, participating in the regulation of colorectal cancer initiation, progression, stemness, EMT, metastasis, and chemotherapy response, they also represent promising biomarkers for CRC diagnosis and therapeutic targets for precision oncology. Better understanding of the regulatory roles of miRNAs in colorectal cancer initiation and progression may provide new insights of developing mini-invasive diagnostic tools for CRC screening and personalized therapy. Despite the numerous studies of miRNAs and extensive analyses of their expression, the roles and functions of many individual miRNAs in CRC remain poorly understood. Therefore, the integrated analysis of multiple miRNA targets for a given miRNA, and the integrated bioinformatic analysis of mRNAs, proteins, copy number variants, and mutations from the available public online databases (e.g., The Cancer Genome Atlas database and Oncomine database), are strongly needed. Improving our understanding of the mechanisms of regulatory interactions of mRNAs with the signaling pathways in colorectal cancer stem cells will aid in determining the genes responsible for progression, metastasis, and recurrence and, finally, in developing personalized prevention and therapy.
Acknowledgments
This work was supported in part by the grants from the Innovation Team of Science and Technology (grant 13IRTSTHN013 to Wancai Yang) of Henan Province, China, and the Startup Fund from Jining Medical University (to Yongchen Guo and Yonghua Bao).
Author Contributions
Wancai Yang conceived and designed the study; Yongchen Guo and Yonghua Bao collected and analyzed the data; and Yonghua Bao and Wancai Yang wrote the paper.
Conflicts of Interest
The authors declared no conflict of interest.
Abbreviations
| miRNA | microRNA |
| CRC | colorectal cancer |
| CSC | cancer stem cells |
| EMT | epithelial-mesenchymal transition |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Punt, C.J.; Koopman, M.; Vermeulen, L. From tumour heterogeneity to advances in precision treatment of colorectal cancer. Nat. Rev. Clin. Oncol. 2017, 14, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Dienstmann, R.; Vermeulen, L.; Guinney, J.; Kopetz, S.; Tejpar, S.; Tabernero, J. Consensus molecular subtypes and the evolution of precision medicine in colorectal cancer. Nat. Rev. Cancer 2017, 17, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Pommergaard, H.C.; Burcharth, J.; Rosenberg, J. Colorectal carcinogenesis-update and perspectives. World J. Gastroenterol. 2014, 20, 18151–18164. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I. Inflammation and colorectal cancer: Colitis-associated neoplasia. Semin. Immunopathol. 2013, 35, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet 2013, 383, 1490–1502. [Google Scholar] [CrossRef]
- Schetter, A.J.; Harris, C.C. Alterations of microRNAs contribute to colon carcinogenesis. Semin. Oncol. 2011, 38, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.V.; Galbraith, N.J.; Yang, D.; Burton, J.F.; Walker, S.P.; Galandiuk, S. Blood-based microRNAs as biomarkers for the diagnosis of colorectal cancer: A systematic review and meta-analysis. Br. J. Cancer 2017, 116, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Strubberg, A.M.; Madison, B.B. MicroRNAs in the etiology of colorectal cancer: Pathways and clinical implications. Dis. Models Mech. 2017, 10, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Marmol, I.; Sanchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Moridikia, A.; Mirzaei, H.; Sahebkar, A.; Salimian, J. MicroRNAs: Potential Candidates for Diagnosis and Treatment of Colorectal Cancer. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Okugawa, Y.; Grady, W.M.; Goel, A. Epigenetic Alterations in Colorectal Cancer: Emerging Biomarkers. Gastroenterology 2015. [Google Scholar] [CrossRef] [PubMed]
- Mamoori, A.; Gopalan, V.; Smith, R.A.; Lam, A.K. Modulatory roles of microRNAs in the regulation of different signalling pathways in large bowel cancer stem cells. Biol. Cell 2016, 108, 51–64. [Google Scholar] [CrossRef] [PubMed]
- De Krijger, I.; Mekenkamp, L.J.; Punt, C.J.; Nagtegaal, I.D. MicroRNAs in colorectal cancer metastasis. J. Pathol. 2011, 224, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Du, Y.; Liu, X.; Cho, W.C.; Yang, Y. MicroRNAs as Regulator of Signaling Networks in Metastatic Colon Cancer. BioMed Res. Int. 2015, 2015, 823620. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Guo, Y.; Li, Z.; Fang, W.; Yang, Y.; Li, X.; Li, Z.; Xiong, B.; Chen, Z.; Wang, J.; et al. MicroRNA profiling in Muc2 knockout mice of colitis-associated cancer model reveals epigenetic alterations during chronic colitis malignant transformation. PLoS ONE 2014, 9, e99132. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wang, Z.; Fan, Q.; Wang, R.; Sun, Y. microRNA-27a functions as a tumor suppressor in esophageal squamous cell carcinoma by targeting KRAS. Oncol. Rep. 2014, 31, 280–286. [Google Scholar] [PubMed]
- Scheibner, K.A.; Teaboldt, B.; Hauer, M.C.; Chen, X.; Cherukuri, S.; Guo, Y.; Kelley, S.M.; Liu, Z.; Baer, M.R.; Heimfeld, S.; et al. miR-27a functions as a tumor suppressor in acute leukemia by regulating 14-3-3θ. PLoS ONE 2012, 7, e50895. [Google Scholar] [CrossRef] [PubMed]
- Chintharlapalli, S.; Papineni, S.; Abdelrahim, M.; Abudayyeh, A.; Jutooru, I.; Chadalapaka, G.; Wu, F.; Mertens-Talcott, S.; Vanderlaag, K.; Cho, S.D.; et al. Oncogenic microRNA-27a is a target for anticancer agent methyl 2-cyano-3,11-dioxo-18β-olean-1,12-dien-30-oate in colon cancer cells. Int. J. Cancer 2009, 125, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.L.; Schmittgen, T.D.; et al. Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Mansoori, B.; Baradaran, B. The role of microRNAs in colorectal cancer. Biomed. Pharmacother. 2016, 84, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Link, A.; Balaguer, F.; Shen, Y.; Nagasaka, T.; Lozano, J.J.; Boland, C.R.; Goel, A. Fecal MicroRNAs as novel biomarkers for colon cancer screening. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1766–1774. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Chen, Z.; Guo, Y.; Feng, Y.; Li, Z.; Han, W.; Wang, J.; Zhao, W.; Jiao, Y.; Li, K.; et al. Tumor suppressor microRNA-27a in colorectal carcinogenesis and progression by targeting SGPP1 and Smad2. PLoS ONE 2014, 9, e105991. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, L.; Xu, Y.; Li, R.; Du, X. microRNA-195 promotes apoptosis and suppresses tumorigenicity of human colorectal cancer cells. Biochem. Biophys. Res. Commun. 2010, 400, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, J.; Meng, X.; Ying, X.; Zuo, Y.; Liu, R.; Pan, Z.; Kang, T.; Huang, W. MicroRNA-342 inhibits colorectal cancer cell proliferation and invasion by directly targeting DNA methyltransferase 1. Carcinogenesis 2011, 32, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Hollis, M.; Nair, K.; Vyas, A.; Chaturvedi, L.S.; Gambhir, S.; Vyas, D. MicroRNAs potential utility in colon cancer: Early detection, prognosis, and chemosensitivity. World J. Gastroenterol. 2015, 21, 8284–8292. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ma, D.; Fesler, A.; Zhai, H.; Leamniramit, A.; Li, W.; Wu, S.; Ju, J. Expression analysis of microRNA as prognostic biomarkers in colorectal cancer. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Song, W.; Chen, Z.H.; Wei, J.H.; Liao, Y.J.; Lei, J.; Hu, M.; Chen, G.Z.; Liao, B.; Lu, J.; et al. Prognostic and predictive value of a microRNA signature in stage II colon cancer: A microRNA expression analysis. Lancet 2013, 14, 1295–1306. [Google Scholar] [CrossRef]
- Pichler, M.; Winter, E.; Stotz, M.; Eberhard, K.; Samonigg, H.; Lax, S.; Hoefler, G. Down-regulation of KRAS-interacting miRNA-143 predicts poor prognosis but not response to EGFR-targeted agents in colorectal cancer. Br. J. Cancer 2012, 106, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Taketo, M.M. Adenomatous polyposis coli (APC): A multi-functional tumor suppressor gene. J. Cell Sci. 2007, 120 Pt 19, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Nathke, I. Cytoskeleton out of the cupboard: Colon cancer and cytoskeletal changes induced by loss of APC. Nat. Rev. Cancer 2006, 6, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Segditsas, S.; Tomlinson, I. Colorectal cancer and genetic alterations in the Wnt pathway. Oncogene 2006, 25, 7531–7537. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; le Sage, C.; Diosdado, B.; van der Waal, M.; Oude Vrielink, J.A.; Bolijn, A.; Meijer, G.A.; Agami, R. Regulation of the adenomatous polyposis coli gene by the miR-135 family in colorectal cancer. Cancer Res. 2008, 68, 5795–5802. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signalling pathway. J. Cell Sci. 2003, 116 Pt 13, 2627–2634. [Google Scholar] [CrossRef] [PubMed]
- Valeri, N.; Braconi, C.; Gasparini, P.; Murgia, C.; Lampis, A.; Paulus-Hock, V.; Hart, J.R.; Ueno, L.; Grivennikov, S.I.; Lovat, F.; et al. MicroRNA-135b promotes cancer progression by acting as a downstream effector of oncogenic pathways in colon cancer. Cancer Cell 2014, 25, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.L.; Wu, D.W.; Huang, C.C.; He, T.Y.; Chou, M.C.; Sheu, G.T.; Lee, H. MicroRNA-21 promotes tumour malignancy via increased nuclear translocation of β-catenin and predicts poor outcome in APC-mutated but not in APC-wild-type colorectal cancer. Carcinogenesis 2014, 35, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Yue, X.; Han, L.; Shi, Z.; Yang, Y.; Pu, P.; Yao, Z.; Kang, C. Genome-wide identification of TCF7L2/TCF4 target miRNAs reveals a role for miR-21 in Wnt-driven epithelial cancer. Int. J. Oncol. 2012, 40, 519–526. [Google Scholar] [PubMed]
- Wan, J.; Xia, L.; Xu, W.; Lu, N. Expression and Function of miR-155 in Diseases of the Gastrointestinal Tract. Int. J. Mol. Sci. 2016, 17, 709. [Google Scholar] [CrossRef] [PubMed]
- Itou, J.; Taniguchi, N.; Oishi, I.; Kawakami, H.; Lotz, M.; Kawakami, Y. HMGB factors are required for posterior digit development through integrating signaling pathway activities. Dev. Dyn. 2011, 240, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, H.S.; Kim, N.G.; Lee, I.; Choi, H.S.; Li, X.Y.; Kang, S.E.; Cha, S.Y.; Ryu, J.K.; Na, J.M.; et al. p53 and microRNA-34 are suppressors of canonical Wnt signaling. Sci. Signal. 2011, 4, ra71. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, M.; Rao, S.R.; Thacker, P.; Chatterjee, S.; Karunagaran, D. miR-29b downregulates canonical Wnt signaling by suppressing coactivators of β-catenin in human colorectal cancer cells. J. Cell. Biochem. 2014, 115, 1974–1984. [Google Scholar] [PubMed]
- Zhang, J.X.; Mai, S.J.; Huang, X.X.; Wang, F.W.; Liao, Y.J.; Lin, M.C.; Kung, H.F.; Zeng, Y.X.; Xie, D. miR-29c mediates epithelial-to-mesenchymal transition in human colorectal carcinoma metastasis via PTP4A and GNA13 regulation of β-catenin signaling. Ann. Oncol. 2014, 25, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Fang, B. RAS signaling and anti-RAS therapy: Lessons learned from genetically engineered mouse models, human cancer cells, and patient-related studies. Acta Biochim. Biophys. Sin. 2016, 48, 27–38. [Google Scholar] [CrossRef] [PubMed]
- De Roock, W.; Piessevaux, H.; de Schutter, J.; Janssens, M.; de Hertogh, G.; Personeni, N.; Biesmans, B.; van Laethem, J.L.; Peeters, M.; Humblet, Y.; et al. KRAS wild-type state predicts survival and is associated to early radiological response in metastatic colorectal cancer treated with cetuximab. Ann. Oncol. 2008, 19, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.M.; Scott, N.; Cross, D.; Sagar, P.; Lewis, F.A.; Blair, G.E.; Taylor, G.R.; Dixon, M.F.; Quirke, P. Prognostic value of p53 overexpression and c-Ki-ras gene mutations in colorectal cancer. Gastroenterology 1993, 104, 57–64. [Google Scholar] [CrossRef]
- Vogelstein, B.; Fearon, E.R.; Hamilton, S.R.; Kern, S.E.; Preisinger, A.C.; Leppert, M.; Nakamura, Y.; White, R.; Smits, A.M.; Bos, J.L. Genetic alterations during colorectal-tumor development. N. Engl. J. Med. 1988, 319, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.K.; Law, P.T.; Lee, C.W.; Cho, C.H.; Fan, D.; Wu, K.; Yu, J.; Sung, J.J. MicroRNA in colorectal cancer: From benchtop to bedside. Carcinogenesis 2011, 32, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Saridaki, Z.; Weidhaas, J.B.; Lenz, H.J.; Laurent-Puig, P.; Jacobs, B.; de Schutter, J.; de Roock, W.; Salzman, D.W.; Zhang, W.; Yang, D.; et al. A let-7 microRNA-binding site polymorphism in KRAS predicts improved outcome in patients with metastatic colorectal cancer treated with salvage cetuximab/panitumumab monotherapy. Clin. Cancer Res. 2014, 20, 4499–4510. [Google Scholar] [CrossRef] [PubMed]
- Sha, D.; Lee, A.M.; Shi, Q.; Alberts, S.R.; Sargent, D.J.; Sinicrope, F.A.; Diasio, R.B. Association study of the let-7 miRNA-complementary site variant in the 3′ untranslated region of the KRAS gene in stage III colon cancer (NCCTG N0147 Clinical Trial). Clin. Cancer Res. 2014, 20, 3319–3327. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Nakagawa, Y.; Naoe, T. let-7 microRNA functions as a potential growth suppressor in human colon cancer cells. Biol. Pharm. Bull. 2006, 29, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, X.; Zhang, H.; Xiang, Y.; Chen, J.; Yin, Y.; Cai, X.; Wang, K.; Wang, G.; Ba, Y.; et al. Role of miR-143 targeting KRAS in colorectal tumorigenesis. Oncogene 2009, 28, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Kent, O.A.; Mendell, J.T.; Rottapel, R. Transcriptional Regulation of miR-31 by Oncogenic KRAS Mediates Metastatic Phenotypes by Repressing RASA1. Mol. Cancer Res. 2016, 14, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yu, F.; Ma, Y.; Zhao, R.; Chen, X.; Zhu, J.; Zhang, C.Y.; Chen, J.; Zhang, J. MicroRNA-31 activates the RAS pathway and functions as an oncogenic MicroRNA in human colorectal cancer by repressing RAS p21 GTPase activating protein 1 (RASA1). J. Biol. Chem. 2013, 288, 9508–9518. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.D.; Boyd, K.L.; Moyo, T.; Mitra, R.; Duszynski, R.; Arrate, M.P.; Chen, X.; Zhao, Z.; Blackwell, T.S.; Andl, T.; et al. MicroRNA-31 initiates lung tumorigenesis and promotes mutant KRAS-driven lung cancer. J. Clin. Investig. 2016, 126, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Oft, M.; Heider, K.H.; Beug, H. TGFβ signaling is necessary for carcinoma cell invasiveness and metastasis. Curr. Biol. 1998, 8, 1243–1252. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Padgett, R.W. Matters of context guide future research in TGFβ superfamily signaling. Sci. Signal. 2015, 8, re10. [Google Scholar] [CrossRef] [PubMed]
- Dumont, N.; Arteaga, C.L. Targeting the TGF β signaling network in human neoplasia. Cancer Cell 2003, 3, 531–536. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.B.; Wakefield, L.M. The two faces of transforming growth factor β in carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 8621–8623. [Google Scholar] [CrossRef] [PubMed]
- Dews, M.; Fox, J.L.; Hultine, S.; Sundaram, P.; Wang, W.; Liu, Y.Y.; Furth, E.; Enders, G.H.; El-Deiry, W.; Schelter, J.M.; et al. The myc-miR-17~92 axis blunts TGFβ signaling and production of multiple TGFβ-dependent antiangiogenic factors. Cancer Res. 2010, 70, 8233–8246. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Bostrom, A.K.; Impens, F.; Fredlund, E.; van Peer, G.; de Antonellis, P.; von Stedingk, K.; Ghesquiere, B.; Schulte, S.; Dews, M.; et al. The miR-17–92 microRNA cluster regulates multiple components of the TGF-β pathway in neuroblastoma. Mol. Cell 2010, 40, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Li, H.; Wang, L.; Hu, J.; Jin, T.; Wang, J.; Yang, B.B. MicroRNA-17–5p promotes chemotherapeutic drug resistance and tumour metastasis of colorectal cancer by repressing PTEN expression. Oncotarget 2014, 5, 2974–2987. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zou, J.; Dong, Z.; Zeng, Q.; Wu, D.; Liu, L. Up-regulated miR-17 promotes cell proliferation, tumour growth and cell cycle progression by targeting the RND3 tumour suppressor gene in colorectal carcinoma. Biochem. J. 2012, 442, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Paysan, L.; Piquet, L.; Saltel, F.; Moreau, V. RND3 in Cancer: A Review of the Evidence for Tumor Promoter or Suppressor. Mol. Cancer Res. 2016, 14, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, V.; Fiorino, A.; Zoni, E.; Crippa, E.; Reid, J.F.; Gariboldi, M.; Pierotti, M.A. The Effects of miR-20a on p21: Two Mechanisms Blocking Growth Arrest in TGF-β-Responsive Colon Carcinoma. J. Cell. Physiol. 2015, 230, 3105–3114. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Liu, C.; Yang, J.; Liu, G.; Feng, F.; Tang, J.; Hu, L.; Li, L.; Jiang, F.; Chen, C.; et al. miR-20a triggers metastasis of gallbladder carcinoma. J. Hepatol. 2013, 59, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, Y.; Lin, S.; Sun, A.; Chen, R.; Ding, Y.; Ding, Y. Down-regualtion of miR-106b induces epithelial-mesenchymal transition but suppresses metastatic colonization by targeting Prrx1 in colorectal cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 10534–10544. [Google Scholar] [PubMed]
- Zheng, L.; Zhang, Y.; Liu, Y.; Zhou, M.; Lu, Y.; Yuan, L.; Zhang, C.; Hong, M.; Wang, S.; Li, X. miR-106b induces cell radioresistance via the PTEN/PI3K/AKT pathways and p21 in colorectal cancer. J. Transl. Med. 2015, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- Petrocca, F.; Vecchione, A.; Croce, C.M. Emerging role of miR-106b-25/miR-17-92 clusters in the control of transforming growth factor β signaling. Cancer Res. 2008, 68, 8191–8194. [Google Scholar] [CrossRef] [PubMed]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory networks underlying colorectal cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Terzic, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114 e5. [Google Scholar] [CrossRef] [PubMed]
- Bromberg, J.; Wang, T.C. Inflammation and cancer: IL-6 and STAT3 complete the link. Cancer Cell 2009, 15, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Wilson, J.E.; Schneider, M.; Lich, J.D.; Roberts, R.A.; Arthur, J.C.; Woodford, R.M.; Davis, B.K.; Uronis, J.M.; Herfarth, H.H.; et al. NLRP12 suppresses colon inflammation and tumorigenesis through the negative regulation of noncanonical NF-κB signaling. Immunity 2012, 36, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Roy, I.; Veldkamp, C.T.; Volkman, B.F.; Dwinell, M.B. Chemokines in colitis: MicroRNA control. Gut 2014, 63, 1202–1204. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Wu, Z.; Huang, L.; Qiu, H.; Wang, L.; Li, L.; Yao, L.; Kang, K.; Qu, J.; Wu, Y.; et al. Mulberry fruit prevents LPS-induced NF-κB/pERK/MAPK signals in macrophages and suppresses acute colitis and colorectal tumorigenesis in mice. Sci. Rep. 2015, 5, 17348. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Trinchieri, G. Innate immune mechanisms of colitis and colitis-associated colorectal cancer. Nat. Rev. Immunol. 2011, 11, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, F.G.; DuBois, R.N. Connecting COX-2 and Wnt in cancer. Cancer Cell 2006, 9, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Tsujii, M.; Kawano, S.; DuBois, R.N. Cyclooxygenase-2 expression in human colon cancer cells increases metastatic potential. Proc. Natl. Acad. Sci. USA 1997, 94, 3336–3340. [Google Scholar] [CrossRef] [PubMed]
- Peacock, O.; Lee, A.C.; Cameron, F.; Tarbox, R.; Vafadar-Isfahani, N.; Tufarelli, C.; Lund, J.N. Inflammation and miR-21 pathways functionally interact to downregulate PDCD4 in colorectal cancer. PLoS ONE 2014, 9, e110267. [Google Scholar] [CrossRef] [PubMed]
- Allgayer, H. Pdcd4, a colon cancer prognostic that is regulated by a microRNA. Crit. Rev. Oncol. Hematol. 2010, 73, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Yang, Y.; Xia, Y.; Okugawa, Y.; Yang, J.; Liang, Y.; Chen, H.; Zhang, P.; Wang, F.; Han, H.; et al. Novel evidence for an oncogenic role of microRNA-21 in colitis-associated colorectal cancer. Gut 2016, 65, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Oshima, M. The inflammatory network in the gastrointestinal tumor microenvironment: Lessons from mouse models. J. Gastroenterol. 2012, 47, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Adachi, O.; Ogawa, T.; Takeda, K.; Akira, S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity 1999, 11, 115–122. [Google Scholar] [CrossRef]
- Liu, S.; Sun, X.; Wang, M.; Hou, Y.; Zhan, Y.; Jiang, Y.; Liu, Z.; Cao, X.; Chen, P.; Liu, Z.; et al. A microRNA 221- and 222-mediated feedback loop maintains constitutive activation of NFκB and STAT3 in colorectal cancer cells. Gastroenterology 2014, 147, 847–859 e11. [Google Scholar] [CrossRef] [PubMed]
- Rokavec, M.; Oner, M.G.; Li, H.; Jackstadt, R.; Jiang, L.; Lodygin, D.; Kaller, M.; Horst, D.; Ziegler, P.K.; Schwitalla, S.; et al. IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated colorectal cancer invasion and metastasis. J. Clin. Investig. 2014, 124, 1853–1867. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Song, L.; Lin, C.; Liu, A.; Lin, X.; Wu, J.; Li, M.; Li, J. Downregulation of miR-138 sustains NF-κB activation and promotes lipid raft formation in esophageal squamous cell carcinoma. Clin. Cancer Res. 2013, 19, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Most, P.; Peppel, K. Induction of microRNA-138 by pro-inflammatory cytokines causes endothelial cell dysfunction. FEBS Lett. 2014, 588, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.C.; Tsai, L.L.; Wang, M.L.; Yu, C.H.; Lo, W.L.; Chang, Y.C.; Chiou, G.Y.; Chou, M.Y.; Chiou, S.H. miR145 targets the SOX9/ADAM17 axis to inhibit tumor-initiating cells and IL-6-mediated paracrine effects in head and neck cancer. Cancer Res. 2013, 73, 3425–3440. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, D.; Gao, J.; Liu, M.; Liu, S.; Jiang, M.; Liu, Y.; Zheng, D. TRAIL-induced miR-146a expression suppresses CXCR4-mediated human breast cancer migration. FEBS J. 2013, 280, 3340–3353. [Google Scholar] [CrossRef] [PubMed]
- Trifari, S.; Pipkin, M.E.; Bandukwala, H.S.; Aijo, T.; Bassein, J.; Chen, R.; Martinez, G.J.; Rao, A. MicroRNA-directed program of cytotoxic CD8+ T-cell differentiation. Proc. Natl. Acad. Sci. USA 2013, 110, 18608–18613. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Zhou, L.; Mi, Q.S. MicroRNA miR-150 is involved in Vα14 invariant NKT cell development and function. J. Immunol. 2012, 188, 2118–2126. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.G. Understanding cancer stem cell heterogeneity and plasticity. Cell Res. 2012, 22, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E. Let-7 and miR-200 microRNAs: Guardians against pluripotency and cancer progression. Cell Cycle 2009, 8, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Zeki, S.S.; Graham, T.A.; Wright, N.A. Stem cells and their implications for colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Martello, G.; Piccolo, S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol. 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Ashley, N. Regulation of intestinal cancer stem cells. Cancer Lett. 2013, 338, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Majumdar, A.P. Signaling in colon cancer stem cells. J. Mol. Signal. 2014, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.L.; Jiang, J.K.; Yang, S.H.; Huang, T.S.; Lan, H.Y.; Teng, H.W.; Yang, C.Y.; Tsai, Y.P.; Lin, C.H.; Wang, H.W.; et al. MicroRNA-146a directs the symmetric division of Snail-dominant colorectal cancer stem cells. Nat. Cell Biol. 2014, 16, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.T.; Xu, Q.; Tong, J.L.; Zhu, M.M.; Nie, F.; Chen, X.; Xiao, S.D.; Ran, Z.H. MicroRNA expression profiling identifies miR-328 regulates cancer stem cell-like SP cells in colorectal cancer. Br. J. Cancer 2012, 106, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Bitarte, N.; Bandres, E.; Boni, V.; Zarate, R.; Rodriguez, J.; Gonzalez-Huarriz, M.; Lopez, I.; Javier Sola, J.; Alonso, M.M.; Fortes, P.; et al. MicroRNA-451 is involved in the self-renewal, tumorigenicity, and chemoresistance of colorectal cancer stem cells. Stem Cells 2011, 29, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
- Bu, P.; Chen, K.Y.; Chen, J.H.; Wang, L.; Walters, J.; Shin, Y.J.; Goerger, J.P.; Sun, J.; Witherspoon, M.; Rakhilin, N.; et al. A microRNA miR-34a-regulated bimodal switch targets Notch in colon cancer stem cells. Cell Stem Cell 2013, 12, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Guilmeau, S. Notch signaling and intestinal cancer. Adv. Exp. Med. Biol. 2012, 727, 272–288. [Google Scholar] [PubMed]
- Fender, A.W.; Nutter, J.M.; Fitzgerald, T.L.; Bertrand, F.E.; Sigounas, G. Notch-1 promotes stemness and epithelial to mesenchymal transition in colorectal cancer. J. Cell. Biochem. 2015, 116, 2517–2527. [Google Scholar] [CrossRef] [PubMed]
- Schwitalla, S.; Fingerle, A.A.; Cammareri, P.; Nebelsiek, T.; Goktuna, S.I.; Ziegler, P.K.; Canli, O.; Heijmans, J.; Huels, D.J.; Moreaux, G.; et al. Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem-cell-like properties. Cell 2013, 152, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Seton-Rogers, S. Epithelial-mesenchymal transition: Untangling EMT’s functions. Nat. Rev. Cancer 2016, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, E.; Birchmeier, W.; Birchmeier, C.; Vande Woude, G. Targeting MET in cancer: Rationale and progress. Nat. Rev. Cancer 2012, 12, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Weinberg, R.A. Epithelial-mesenchymal transition: At the crossroads of development and tumor metastasis. Dev. Cell 2008, 14, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Burk, U.; Schubert, J.; Wellner, U.; Schmalhofer, O.; Vincan, E.; Spaderna, S.; Brabletz, T. A reciprocal repression between ZEB1 and members of the miR-200 family promotes EMT and invasion in cancer cells. EMBO Rep. 2008, 9, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, S.; Brabletz, T. The ZEB/miR-200 feedback loop—A motor of cellular plasticity in development and cancer? EMBO Rep. 2010, 11, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, H.S.; Li, X.Y.; Lee, I.; Choi, H.S.; Kang, S.E.; Cha, S.Y.; Ryu, J.K.; Yoon, D.; Fearon, E.R.; et al. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial-mesenchymal transition. J. Cell Biol. 2011, 195, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Siemens, H.; Jackstadt, R.; Hunten, S.; Kaller, M.; Menssen, A.; Gotz, U.; Hermeking, H. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle 2011, 10, 4256–4271. [Google Scholar] [CrossRef] [PubMed]
- Jackstadt, R.; Roh, S.; Neumann, J.; Jung, P.; Hoffmann, R.; Horst, D.; Berens, C.; Bornkamm, G.W.; Kirchner, T.; Menssen, A.; et al. AP4 is a mediator of epithelial-mesenchymal transition and metastasis in colorectal cancer. J. Exp. Med. 2013, 210, 1331–1350. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Jackstadt, R.; Siemens, H.; Li, H.; Kirchner, T.; Hermeking, H. p53-induced miR-15a/16–1 and AP4 form a double-negative feedback loop to regulate epithelial-mesenchymal transition and metastasis in colorectal cancer. Cancer Res. 2014, 74, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Shell, S.; Park, S.M.; Radjabi, A.R.; Schickel, R.; Kistner, E.O.; Jewell, D.A.; Feig, C.; Lengyel, E.; Peter, M.E. Let-7 expression defines two differentiation stages of cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 11400–11405. [Google Scholar] [CrossRef] [PubMed]
- King, C.E.; Cuatrecasas, M.; Castells, A.; Sepulveda, A.R.; Lee, J.S.; Rustgi, A.K. LIN28B promotes colon cancer progression and metastasis. Cancer Res. 2011, 71, 4260–4268. [Google Scholar] [CrossRef] [PubMed]
- Cottonham, C.L.; Kaneko, S.; Xu, L. miR-21 and miR-31 converge on TIAM1 to regulate migration and invasion of colon carcinoma cells. J. Biol. Chem. 2010, 285, 35293–35302. [Google Scholar] [CrossRef] [PubMed]
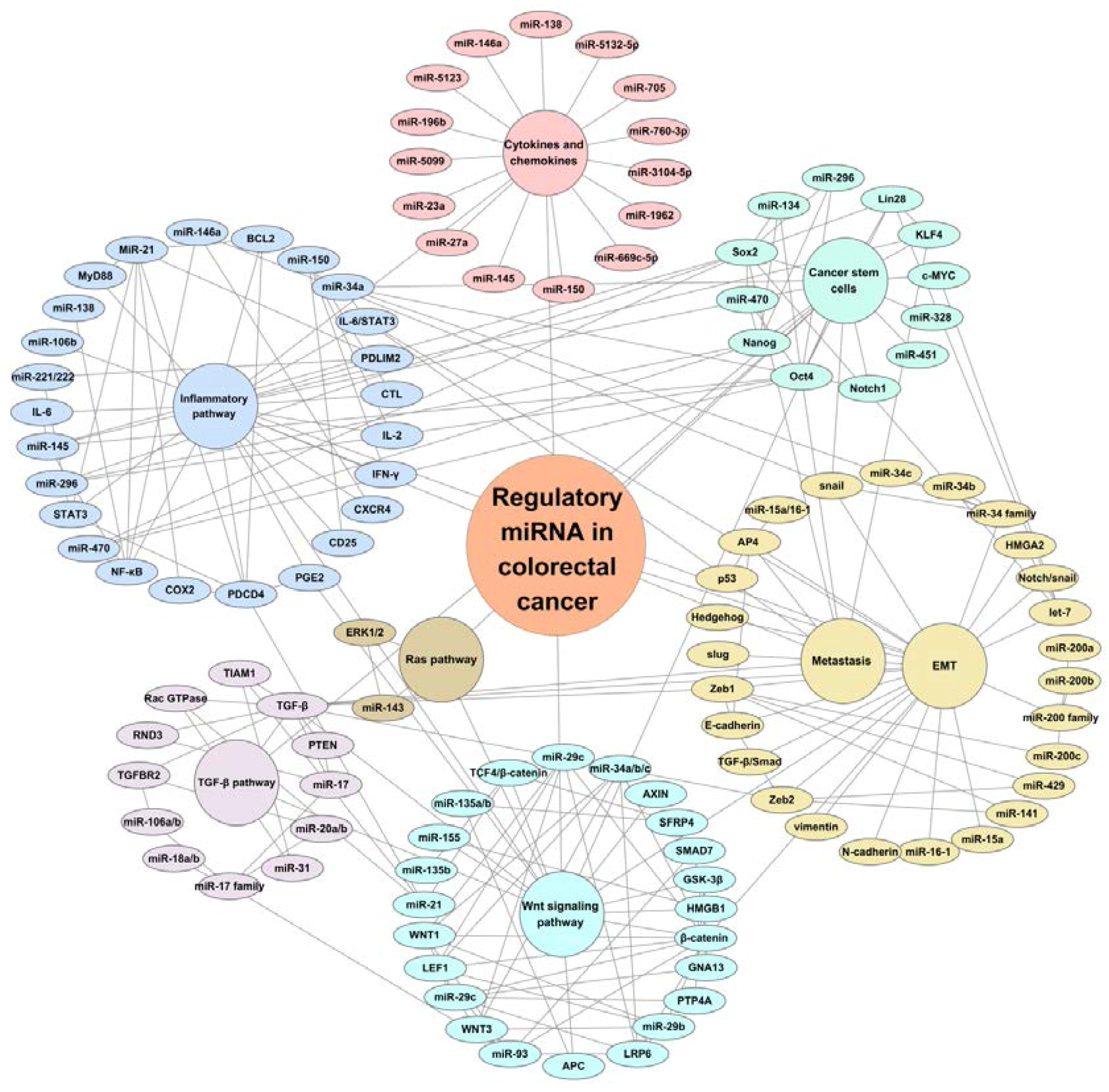
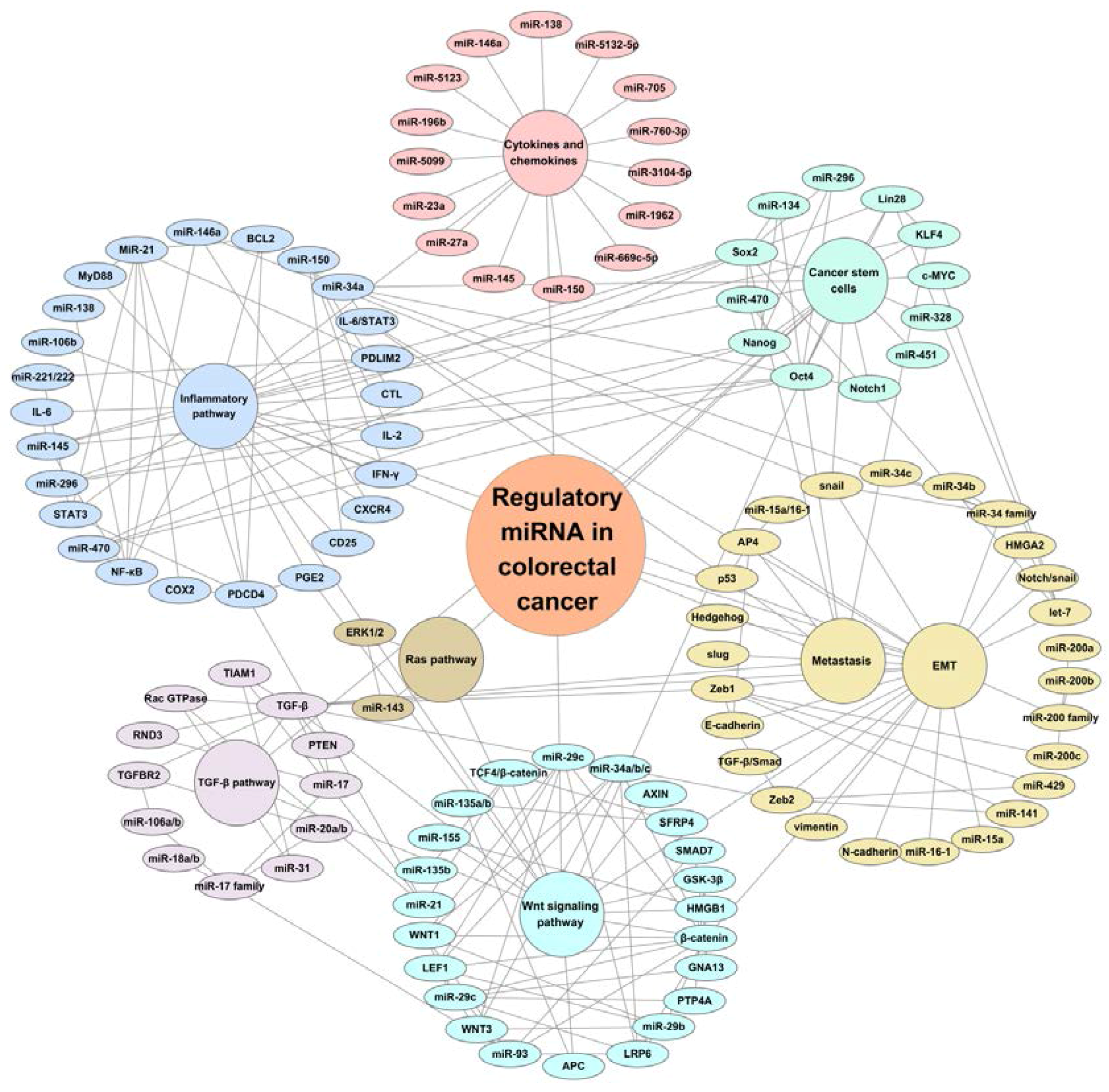
Figure 1.
Scheme of the regulatory miRNAs and signaling pathways in colorectal cancer. There is a regulatory network linked to miRNAs and their targets, such as miRNAs regulating Wnt, Ras, transforming growth factor β (TGF-β),inflammatory, cytokine and chenmokines, cancer stem cells, epithelial-mesenchymal (EMT), and metastasis signaling pathways (in different colors). In addition, these signaling pathways are linked and regulated by each other.
Figure 1.
Scheme of the regulatory miRNAs and signaling pathways in colorectal cancer. There is a regulatory network linked to miRNAs and their targets, such as miRNAs regulating Wnt, Ras, transforming growth factor β (TGF-β),inflammatory, cytokine and chenmokines, cancer stem cells, epithelial-mesenchymal (EMT), and metastasis signaling pathways (in different colors). In addition, these signaling pathways are linked and regulated by each other.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, Y.; Bao, Y.; Yang, W. Regulatory miRNAs in Colorectal Carcinogenesis and Metastasis. Int. J. Mol. Sci. 2017, 18, 890. https://doi.org/10.3390/ijms18040890
AMA Style
Guo Y, Bao Y, Yang W. Regulatory miRNAs in Colorectal Carcinogenesis and Metastasis. International Journal of Molecular Sciences. 2017; 18(4):890. https://doi.org/10.3390/ijms18040890
Chicago/Turabian StyleGuo, Yongchen, Yonghua Bao, and Wancai Yang. 2017. "Regulatory miRNAs in Colorectal Carcinogenesis and Metastasis" International Journal of Molecular Sciences 18, no. 4: 890. https://doi.org/10.3390/ijms18040890
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.