
The Effect of Chinese Herbal Medicine Formula mKG on Allergic Asthma by Regulating Lung and Plasma Metabolic Alternations

Abstract
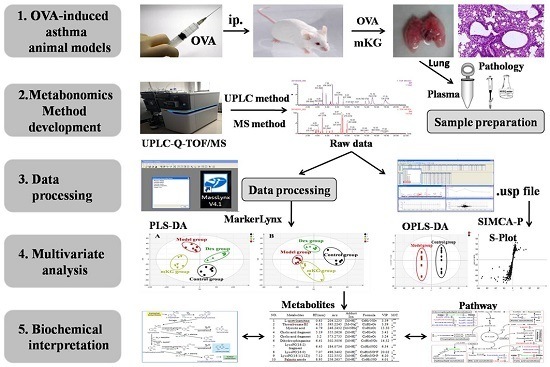
:
1. Introduction
2. Results
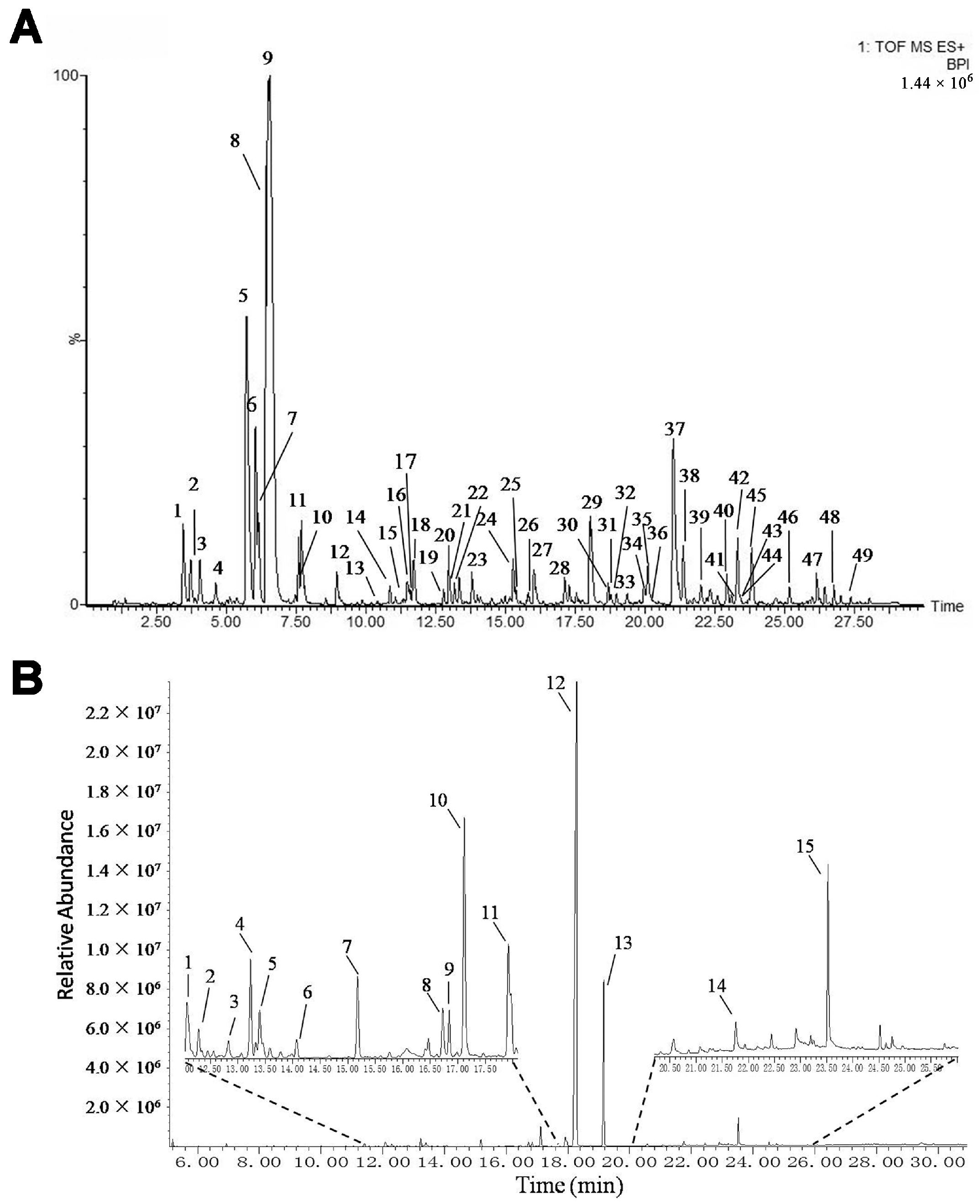
2.1. Identification of the Chemical Constituents in mKG Extract
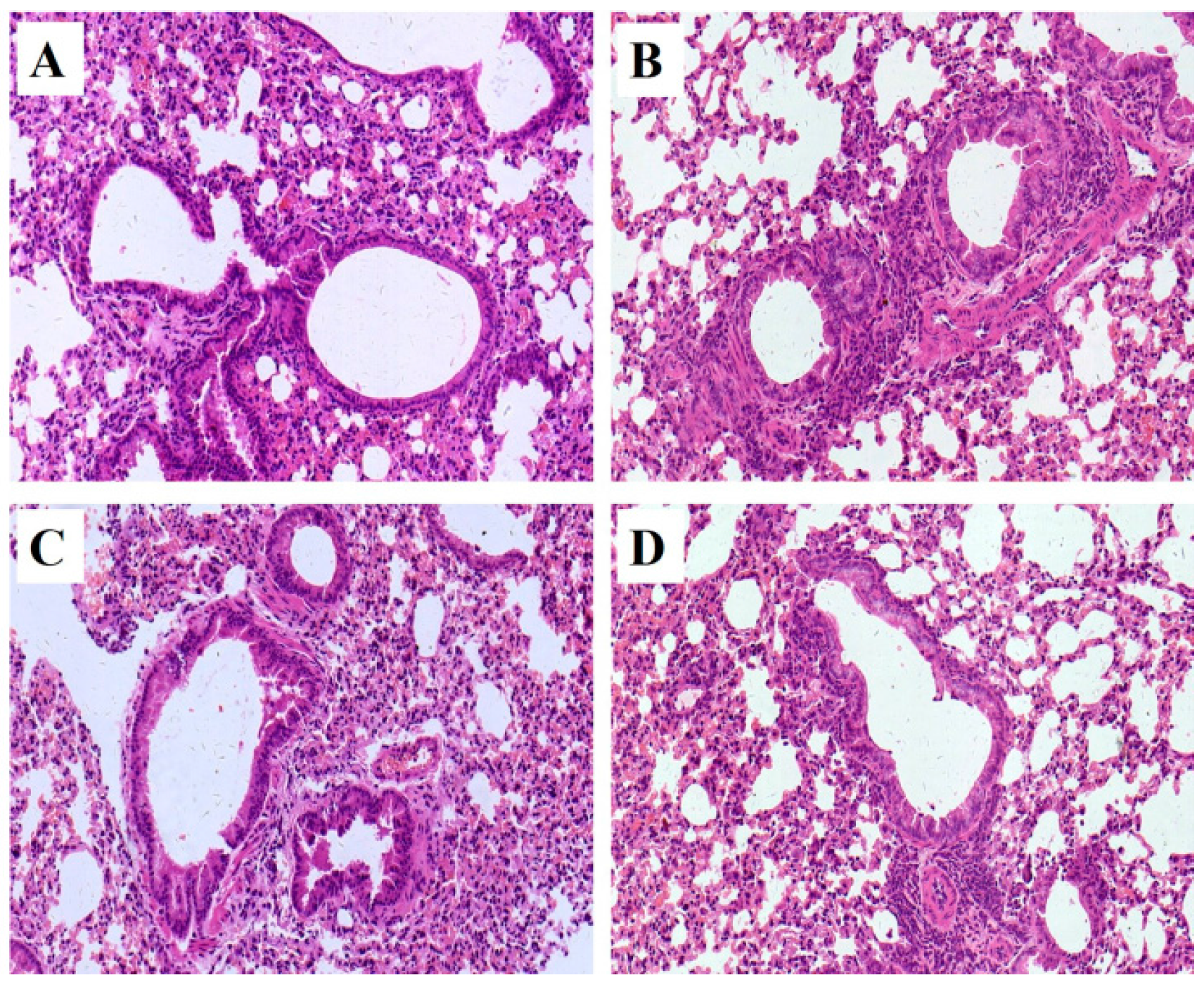
2.2. Histopathology
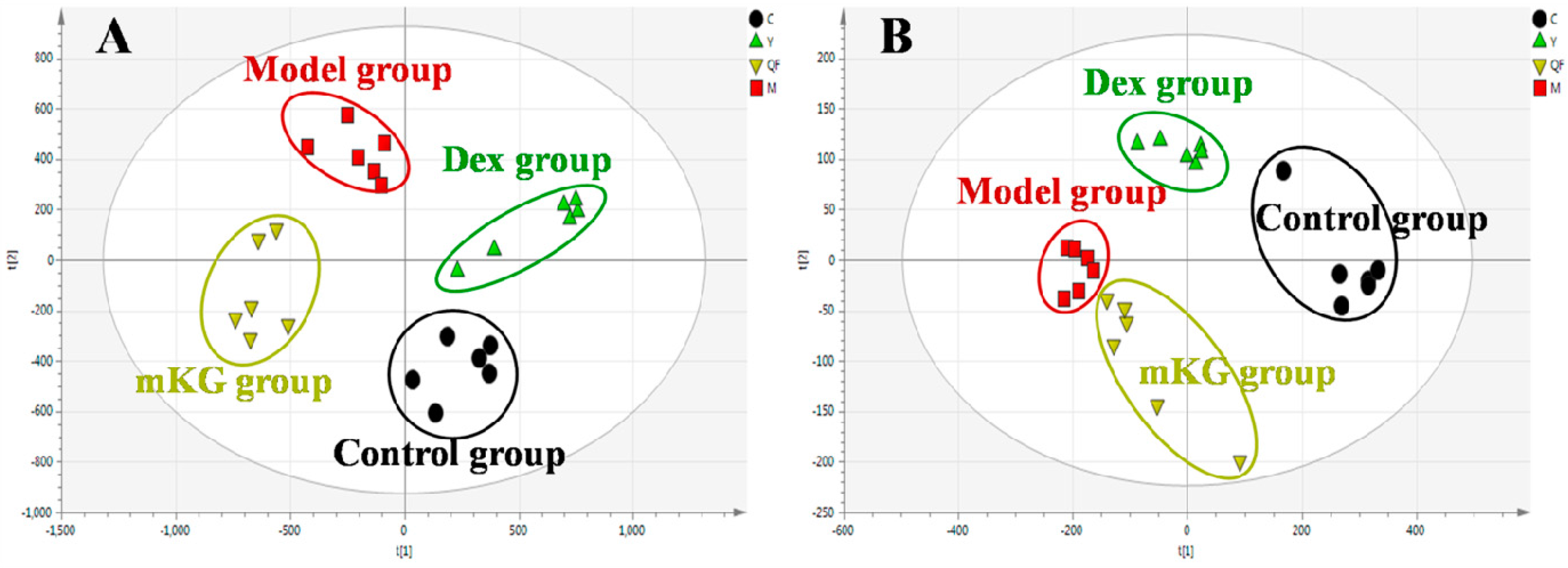
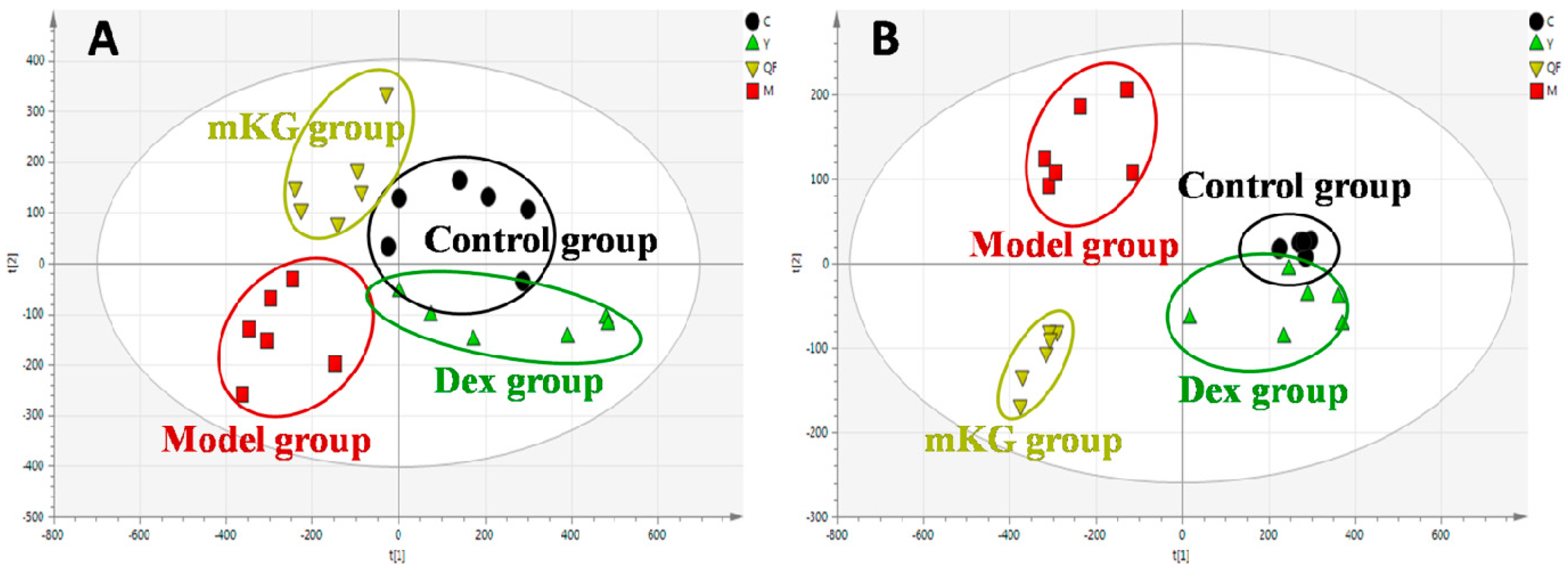
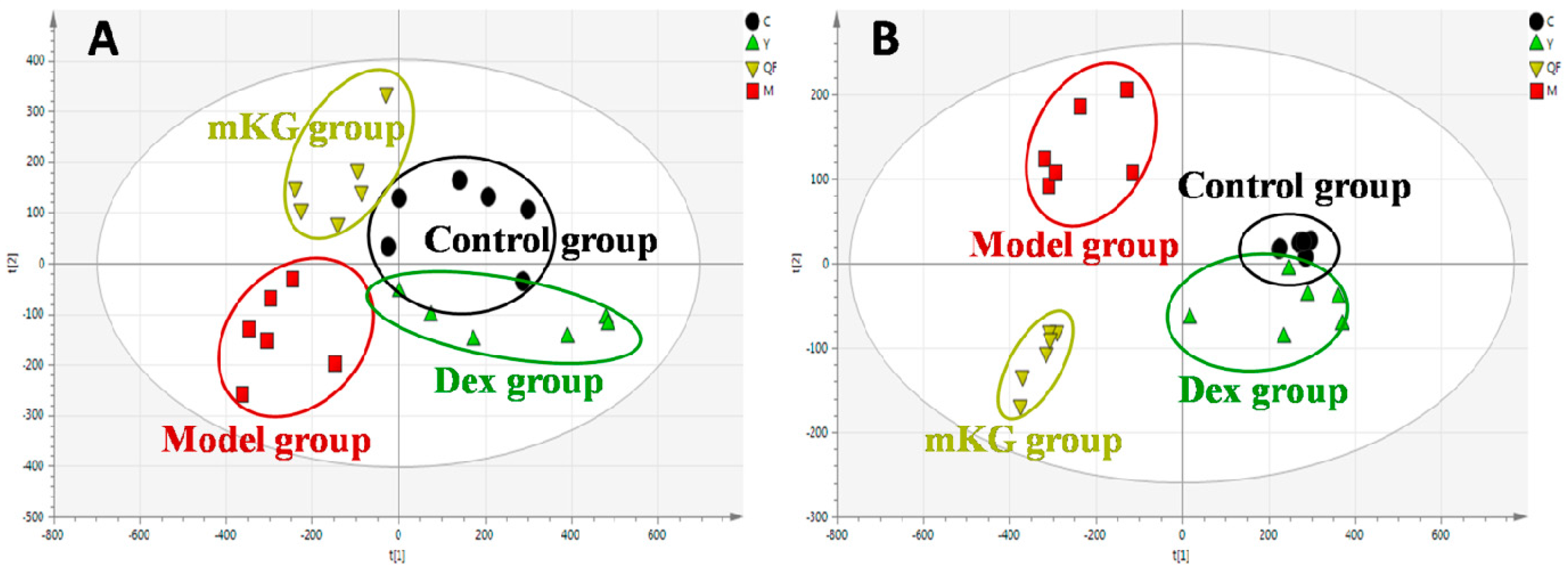
2.3. Lung Tissue Metabolic Profiles of OVA-Induced Allergic Asthma with mKG Treatment
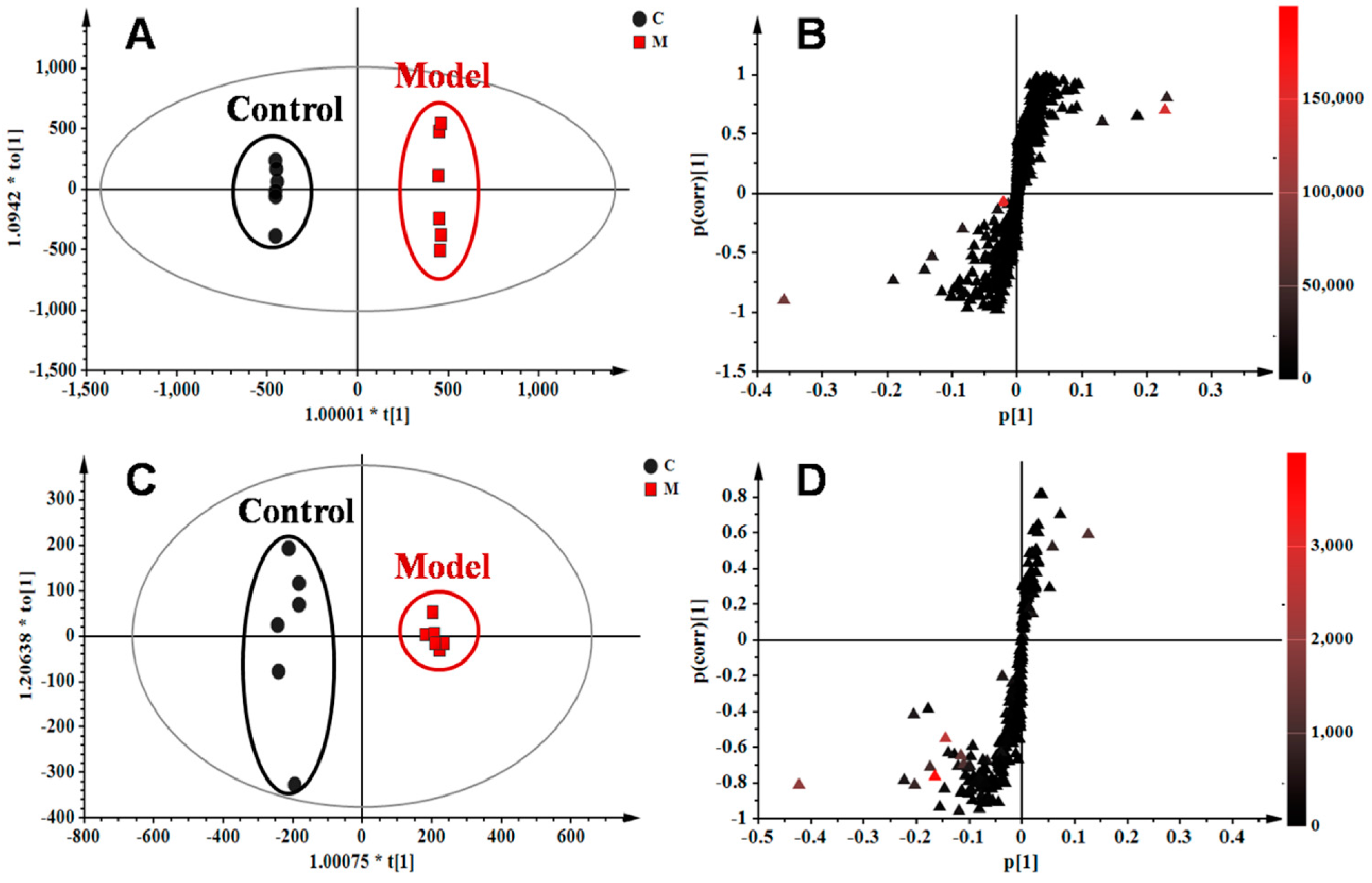
2.4. Identification of Potential Lung Tissue Biomarkers Associated with OVA-Induced Asthma
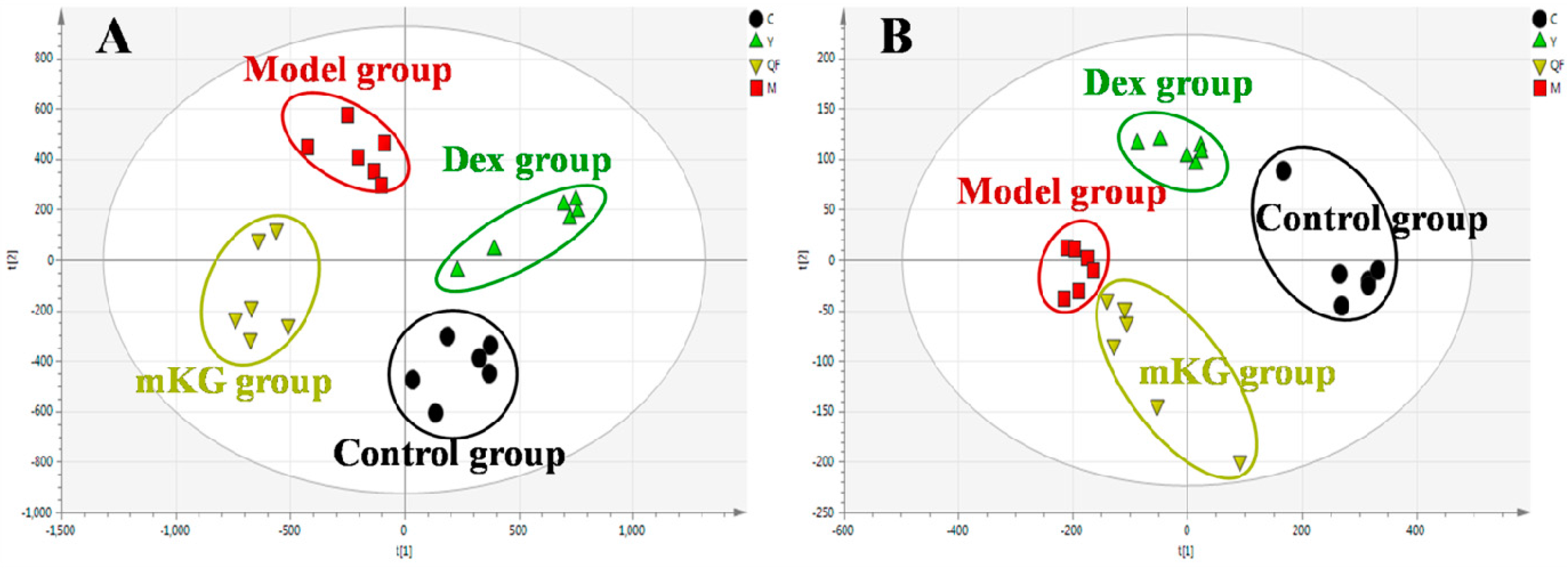
2.5. Plasma Metabolic Profiles of OVA-Induced Allergic Asthma with mKG Treatment
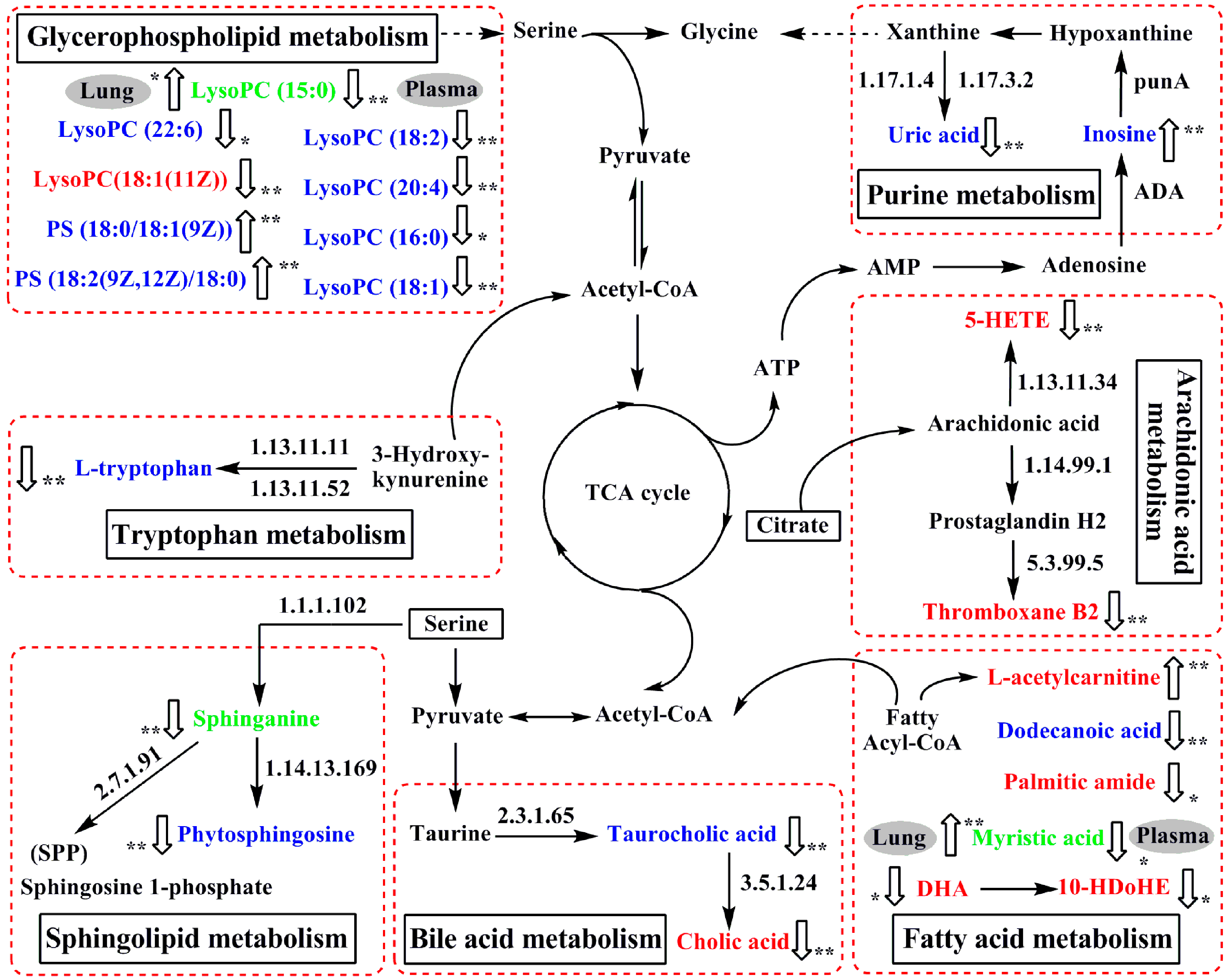
2.6. Perturbed Metabolic Pathways in Response to Allergic Asthma and mKG Treatment
2.6.1. Fatty Acid Metabolism
2.6.2. Sphingolipid Metabolism
2.6.3. Glycerophospholipid Metabolism
2.6.4. Purine Metabolism
2.6.5. Tryptophan Metabolism
2.6.6. Bile Acid Metabolism
2.6.7. Arachidonic Acid Metabolism
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Raw Herbal Medicines and mKG Extract
4.3. Qualitative Characteristics of Chemical Constituents in mKG Extract
4.4. Animal Treatment and Sample Collection
4.5. Histopathology
4.6. Sample Preparation for Metabolomics
4.7. Data Acquisition
4.8. Multivariate Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| mKG | Modified Kushen Gancao |
| TCM | traditional Chinese medicines |
| OVA | ovalbumin |
| UPLC-Q-TOF/MS | ultra high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry |
| PLS−DA | partial least squares–discriminate analysis |
| OPLS−DA | orthogonal partial least squares–discriminate analysis |
| Ach | acetylcholine |
| ASM | airway smooth muscle |
| Dex | dexamethasone |
| HE | hematoxylin-eosin |
| BPI | base peak intensity |
| PCA | principal component analysis |
| VIP | variable importance of project |
| KEGG | kyoto encyclopedia of genes and genomes |
| HMDB | human metabolome database |
| DHA | docosahexaenoic acid |
| MAP | mitogen activated protein |
| DSR | dehydrosphinganine reductase |
| MPT | mitochondrial permeability transitionpore |
| AMP | adenosine monophosphate |
| HDM | house dust mite |
| AA | Arachidonic acid |
| PUFA | polyunsaturated fatty acid |
| Lo | lipoxygenase |
| HETE | hydroxyeicosatetraenoic acid |
| BALF | bronchoalveolar lavage fluid |
References
- Busse, W.W.; Lemanske, R.F., Jr. Asthma. N. Engl. J. Med. 2001, 344, 350–362. [Google Scholar] [PubMed]
- Masoli, M.; Fabian, D.; Holt, S.; Beasley, R. The global burden of asthma: Executive summary of the GINA Dissemination Committee report. Allergy 2004, 59, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, A.; Tantisira, K.G. Integrative systems biology approaches in asthma pharmacogenomics. Pharmacogenomics 2012, 13, 1387–1404. [Google Scholar] [CrossRef] [PubMed]
- Drazen, J.M.; Silverman, E.K.; Lee, T.H. Heterogeneity of therapeutic responses in asthma. Br. Med. Bull. 2000, 56, 1054–1070. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Huang, C.K.; Zhang, T.F.; Teper, A.A.; Srivastava, K.; Schofield, B.H.; Sampson, H.A. The Chinese herbal medicine formula MSSM-002 suppresses allergic airway hyperreactivity and modulates TH1/TH2 responses in a murine model of allergic asthma. J. Allergy Clin. Immunol. 2000, 106, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yao, L.F.; Zhao, Y.; Wei, L.M.; Guo, P.; Yu, M.; Cao, B.; Li, T.; Chen, H.; Zou, Z.M. The Chinese Herbal Medicine Formula mKG Suppresses Pulmonary Fibrosis of Mice Induced by Bleomycin. Int. J. Mol. Sci. 2016, 17, 238. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Fang, J.; Huang, L.; Wang, J.; Huang, X. Sophora flavescens Ait.: Traditional usage, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2015, 172, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, N.; Song, Y.; Wang, L.; Zi, J.; Zhang, S.; Dunkin, D.; Busse, P.; Weir, D.; Tversky, J.; et al. Ganoderic acid C1 isolated from the anti-asthma formula, ASHMI suppresses TNF-α production by mouse macrophages and peripheral blood mononuclear cells from asthma patients. Int. Immunopharmacol. 2015, 27, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Wang, J.; Ma, Z.; Ma, S. Anti-asthmatic effects of matrine in a mouse model of allergic asthma. Fitoterapia 2014, 94, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Z.; Fu, Q.; Chen, T.; Ma, S.P. Anti-asthmatic effects of oxymatrine in a mouse model of allergic asthma through regulating CD40 signaling. Chin. J. Nat. Med. 2015, 13, 368–374. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, L.; Shan, L.; Fan, G.; Gao, X. Liquorice, a unique “guide drug” of traditional Chinese medicine: A review of its role in drug interactions. J. Ethnopharmacol. 2013, 150, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.W.; Bae, E.A.; Lee, B.; Lee, S.H.; Kim, J.A.; Kim, Y.S.; Kim, D.H. In vitro and in vivo antiallergic effects of Glycyrrhiza glabra and its components. Planta Med. 2007, 73, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Ram, A.; Mabalirajan, U.; Das, M.; Bhattacharya, I.; Dinda, A.K.; Gangal, S.V.; Ghosh, B. Glycyrrhizin alleviates experimental allergic asthma in mice. Int. Immunopharmacol. 2006, 6, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Tang, Y.; Hu, X.; Wang, Q.; Lei, W.; Zhou, L.; Huang, J. Regulation of Th1/Th2 balance through OX40/OX40L signalling by glycyrrhizic acid in a murine model of asthma. Respirology 2016, 21, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Liang, B.; Srivastava, K.; Zeng, J.; Zhan, J.; Brown, L.; Sampson, H.; Goldfarb, J.; Emala, C.; Li, X.M. The Sophora flavescens flavonoid compound trifolirhizin inhibits acetylcholine induced airway smooth muscle contraction. Phytochemistry 2013, 95, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Zhao, K.; Huang, Q.; Xu, C.; Shang, P. Isolation, structure and bioactivities of the polysaccharides from Angelica sinensis (Oliv.) Diels: A review. Carbohydr. Polym. 2012, 89, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.W.; Lin, B.F. Bioactivities of major constituents isolated from Angelica sinensis (Danggui). Chin. Med. 2011, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Comhair, S.A.; McDunn, J.; Bennett, C.; Fettig, J.; Erzurum, S.C.; Kalhan, S.C. Metabolomic Endotype of Asthma. J. Immunol. (Baltim. Md. 1950) 2015, 195, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Drexler, D.M.; Reily, M.D.; Shipkova, P.A. Advances in mass spectrometry applied to pharmaceutical metabolomics. Anal. Bioanal. Chem. 2011, 399, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Chen, X.; Liang, Q.; Zhang, H.; Hu, P.; Wang, Y.; Luo, G. Metabonomic study of Chinese medicine Shuanglong formula as an effective treatment for myocardial infarction in rats. J. Proteome Res. 2011, 10, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Cui, F.X.; Jia, H.M.; Zhou, C.; Yang, Y.; Zhang, H.W.; Ding, G.; Zou, Z.M. Aberrant purine metabolism in allergic asthma revealed by plasma metabolomics. J. Pharm. Biomed. Anal. 2016, 120, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Stanley, W.C.; Recchia, F.A.; Lopaschuk, G.D. Myocardial substrate metabolism in the normal and failing heart. Physiol. Rev. 2005, 85, 1093–1129. [Google Scholar] [CrossRef] [PubMed]
- Al-Biltagi, M.; Isa, M.; Bediwy, A.S.; Helaly, N.; El Lebedy, D.D. l-carnitine improves the asthma control in children with moderate persistent asthma. J. Allergy 2012, 2012, 509730. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, X.; Wang, M.; Liu, L.; Sun, X.; Ma, L.; Xie, W.; Wang, C.; Tang, S.; Wang, D.; et al. Lysophosphatidylcholine and amide as metabolites for detecting alzheimer disease using ultrahigh-performance liquid chromatography-quadrupole time-of-flight mass spectrometry-based metabonomics. J. Neuropathol. Exp. Neurol. 2014, 73, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Shang, V.C.; O’Sullivan, S.E.; Kendall, D.A.; Roberts, R.E. The endogenous cannabinoid anandamide increases human airway epithelial cell permeability through an arachidonic acid metabolite. Pharmacol. Res. 2016, 105, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Miyata, J.; Arita, M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases. Allergol. Int. Off. J. Jpn. Soc. Allergol. 2015, 64, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Tsai, T.H.; Chuang, L.T.; Li, Y.Y.; Zouboulis, C.C.; Tsai, P.J. Anti-bacterial and anti-inflammatory properties of capric acid against Propionibacterium acnes: A comparative study with lauric acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, E.; Perea, J.M.; Jimenez, A.I.; Rodriguez-Rodriguez, P.; Lopez-Sobaler, A.M.; Ortega, R.M. Fat intake and asthma in Spanish schoolchildren. Eur. J. Clin. Nutr. 2010, 64, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.J.; Hackett, S.R.; Harshman, L.G.; Clark, A.G. Environmental and genetic perturbations reveal different networks of metabolic regulation. Mol. Syst. Biol. 2011, 7, 563. [Google Scholar] [CrossRef] [PubMed]
- Ammit, A.J.; Hastie, A.T.; Edsall, L.C.; Hoffman, R.K.; Amrani, Y.; Krymskaya, V.P.; Kane, S.A.; Peters, S.P.; Penn, R.B.; Spiegel, S.; et al. Sphingosine 1-phosphate modulates human airway smooth muscle cell functions that promote inflammation and airway remodeling in asthma. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2001, 15, 1212–1214. [Google Scholar] [CrossRef]
- Shevchenko, A.; Simons, K. Lipidomics: Coming to grips with lipid diversity. Nat. Rev. Mol. Cell Biol. 2010, 11, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.R. Lipidomics: New tools and applications. Cell 2010, 143, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatse, S.; De Clercq, E.; Balzarini, J. Role of antimetabolites of purine and pyrimidine nucleotide metabolism in tumor cell differentiation. Biochem. Pharmacol. 1999, 58, 539–555. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; Coakley, R.D.; Henderson, A.G.; Zhou, Y.H.; Wright, F.A.; Boucher, R.C. Metabolomic Evaluation of Neutrophilic Airway Inflammation in Cystic Fibrosis. Chest 2015, 148, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Van der Sluijs, K.F.; van de Pol, M.A.; Kulik, W.; Dijkhuis, A.; Smids, B.S.; van Eijk, H.W.; Karlas, J.A.; Molenkamp, R.; Wolthers, K.C.; Johnston, S.L.; et al. Systemic tryptophan and kynurenine catabolite levels relate to severity of rhinovirus-induced asthma exacerbation: A prospective study with a parallel-group design. Thorax 2013, 68, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.E.; Xu, Y.J.; Cheng, C.; Peh, H.Y.; Tannenbaum, S.R.; Wong, W.S.; Ong, C.N. Metabolomics Reveals Inflammatory-Linked Pulmonary Metabolic Alterations in a Murine Model of House Dust Mite-Induced Allergic Asthma. J. Proteome Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.S.; Rokach, J. Biosynthesis, biological effects, and receptors of hydroxyeicosatetraenoic acids (HETEs) and oxoeicosatetraenoic acids (oxo-ETEs) derived from arachidonic acid. Biochim. Biophys. Acta 2015, 1851, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Dev, A.; Leong, A.B. Zileuton, a 5-lipoxygenase inhibitor, increases production of thromboxane A2 and platelet aggregation in patients with asthma. Am. J. Hematol. 2003, 74, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kim, S.H.; Lee, H.S.; Choi, G.S.; Jung, Y.S.; Ryu, D.H.; Park, H.S.; Hwang, G.S. Serum metabolomics reveals pathways and biomarkers associated with asthma pathogenesis. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2013, 43, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Mattarucchi, E.; Baraldi, E.; Guillou, C. Metabolomics applied to urine samples in childhood asthma; differentiation between asthma phenotypes and identification of relevant metabolites. Biomed. Chromatogr. BMC 2012, 26, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, W.E.; Xu, Y.J.; Xu, F.; Cheng, C.; Peh, H.Y.; Tannenbaum, S.R.; Wong, W.S.; Ong, C.N. Metabolomics reveals altered metabolic pathways in experimental asthma. Am. J. Respir. Cell Mol. Biol. 2013, 48, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Jia, H.M.; Chang, X.; Ding, G.; Zhang, H.W.; Zou, Z.M. The metabolic disturbances of isoproterenol induced myocardial infarction in rats based on a tissue targeted metabonomics. Mol. BioSyst. 2013, 9, 2823–2834. [Google Scholar] [CrossRef] [PubMed]



 : up-regulated,
: up-regulated,  : down-regulated; * p < 0.05, ** p < 0.01.
: up-regulated, : down-regulated; * p < 0.05, ** p < 0.01.
: down-regulated; * p < 0.05, ** p < 0.01.
: up-regulated, : down-regulated; * p < 0.05, ** p < 0.01.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Group | Components | PLS−DA (Positive Ion Mode) | PLS−DA (Negative Ion Mode) | ||||
|---|---|---|---|---|---|---|---|---|
| R2X | R2Y | Q2 (cum) | R2X | R2Y | Q2 (cum) | |||
| Plasma samples | Control vs. Model | 3 | 0.684 | 0.998 | 0.939 | 0.823 | 0.991 | 0.973 |
| Model vs. mKG | 3 | 0.579 | 0.992 | 0.891 | 0.558 | 0.963 | 0.898 | |
| Model vs. Dex | 3 | 0.679 | 0.998 | 0.943 | 0.801 | 0.989 | 0.884 | |
| Lung tissue samples | Control vs. Model | 3 | 0.417 | 0.992 | 0.894 | 0.558 | 0.990 | 0.919 |
| Model vs. mKG | 3 | 0.613 | 0.991 | 0.848 | 0.753 | 0.985 | 0.939 | |
| Model vs. Dex | 3 | 0.464 | 0.987 | 0.836 | 0.587 | 0.970 | 0.977 | |
| NO. | Metabolites | RT (min) | m/z | Formula | VIP | M/C | Y/M | mKG/M | Pathways |
|---|---|---|---|---|---|---|---|---|---|
| Positive ion mode | |||||||||
| L1 | l-Acetylcarnitine b | 0.63 | 204.1233 | C9H17NO4 | 3.59 | ↑ ** | ↓ ** | ↓ | Fatty acid metabolism |
| L2 | Thromboxane B2 b | 4.15 | 393.2245 | C20H34O6 | 3.39 | ↓ ** | ↑ * | ↑ | Arachidonicacid metabolism |
| L3 | Myristic acid b | 4.79 | 246.2432 | C14H28O2 | 13.30 | ↑ * | ↓ ** | ↓ | Fatty acid metabolism |
| L4 | Cholic acid fragment a | 5.19 | 355.2626 | C24H34O2 | 5.41 | ↓ ** | ↑ ** | ↑ ** | Bile Acid Biosynthesis |
| L5 | Cholic acid fragment a | 5.2 | 373.2735 | C24H36O3 | 3.24 | ↓ ** | ↑ | ↑ | Bile Acid Biosynthesis |
| L6 | Sphinganine b | 6.41 | 302.3056 | C18H39NO2 | 14.52 | ↑ ** | ↓ ** | ↓ * | Sphingolipid metabolism |
| L7 | LysoPC(18:1(11Z)) b | 7.12 | 522.3552 | C26H52NO7P | 6.20 | ↓ ** | ↑ | ↑ | Glycerophospholipid metabolism |
| L8 | Palmitic amide b | 8.95 | 256.2637 | C16H33NO | 4.01 | ↓ * | ↑ * | ↑ | Fatty acid metabolism |
| Negative ion mode | |||||||||
| L9 | Cholic acid a | 5.19 | 407.279 | C24H40O5 | 4.04 | ↓ ** | ↑ ** | ↑ ** | Bile Acid Biosynthesis |
| L10 | 10-HDoHE b | 6.93 | 343.2262 | C22H32O3 | 1.71 | ↓ * | ↑ * | ↑ | Fatty acid metabolism |
| L11 | 5-HETE b | 7.04 | 319.2264 | C20H32O3 | 7.90 | ↓ ** | ↑ ** | ↑ ** | Arachidonicacid metabolism |
| L12 | LysoPC(15:0) b | 7.04 | 480.3081 | C23H48NO7P | 2.33 | ↑ * | ↑ | ↓ | Glycerophospholipid metabolism |
| L13 | Docosahexaenoic acid (DHA) b | 8.91 | 327.2315 | C21H31O2 | 3.39 | ↓ * | ↑ | ↑ | Fatty acid metabolism |
| NO. | Metabolites | RT (min) | m/z | Formula | VIP | M/C | Y/M | mKG/M | Pathways |
|---|---|---|---|---|---|---|---|---|---|
| Positive ion mode | |||||||||
| P1 | Dodecanoic acid b | 2.25 | 218.2116 | C12H24O2 | 2.60 | ↓ ** | ↑ ** | ↑ ** | Fatty acid metabolism |
| P2 | Myristic acid b | 2.87 | 246.2427 | C14H28O2 | 2.57 | ↓ ** | ↑ ** | ↑ ** | Fatty acid metabolism |
| P3 | Phytosphingosine b | 4.13 | 318.3002 | C18H39NO3 | 5.00 | ↓ ** | ↑ ** | ↑ ** | Sphingolipid metabolism |
| P4 | Sphinganine b | 6.06 | 302.3053 | C18H39NO2 | 4.70 | ↓ ** | ↑ ** | ↑ ** | Sphingolipid metabolism |
| P5 | LysoPC(22:6) b | 6.85 | 568.3402 | C30H50NO7P | 2.45 | ↓ ** | ↑ ** | ↑ | Glycerophospholipid metabolism |
| P6 | LysoPC(18:2) b | 6.90 | 520.3405 | C26H50NO7P | 8.34 | ↓ ** | ↑ ** | ↑ ** | Glycerophospholipid metabolism |
| P7 | LysoPC(20:4) b | 6.92 | 544.3402 | C28H50NO7P | 5.95 | ↓ ** | ↓ | ↑ ** | Glycerophospholipid metabolism |
| P8 | LysoPC(16:0) b | 7.85 | 496.3404 | C24H50NO7P | 5.56 | ↓ * | ↑ * | ↑ | Glycerophospholipid metabolism |
| P9 | LysoPC(18:1) b | 8.46 | 522.3554 | C26H52NO7P | 4.01 | ↓ ** | ↑ ** | ↑ * | Glycerophospholipid metabolism |
| P10 | PS(18:0/18:1(9Z)) b | 13.57 | 790.5587 | C42H80NO10P | 1.79 | ↑ ** | ↓ * | ↓ * | Glycerophospholipid metabolism |
| P11 | PS(18:2(9Z,12Z)/18:0) b | 13.67 | 788.5428 | C42H78NO10P | 3.64 | ↑ ** | ↓ * | ↓* | Glycerophospholipid metabolism |
| Negative ion mode | |||||||||
| P12 | Uric acid a | 0.61 | 167.0123 | C5H4N4O3 | 1.50 | ↓ ** | ↑ ** | ↑ ** | Purine metabolism |
| P13 | Inosine a | 0.66 | 267.0561 | C10H12N4O5 | 1.02 | ↑ ** | ↓ ** | ↓ ** | Purine metabolism |
| P14 | l-tryptophan a | 1.28 | 203.0728 | C11H12N2O2 | 1.29 | ↓ ** | ↑ * | ↑ | Tryptophan metabolism |
| P15 | Taurocholic acid a | 2.40 | 514.2839 | C26H45NO7S | 1.74 | ↓ ** | ↑ | ↑ | Bile acid metabolism |
| P5 | LysoPC(22:6) b | 6.86 | 612.3295 | C30H50NO7P | 1.74 | ↓ ** | ↑ ** | ↑ | Glycerophospholipid metabolism |
| P6 | LysoPC(18:2) b | 6.90 | 564.3299 | C26H50NO7P | 3.08 | ↓ ** | ↑ ** | ↑ ** | Glycerophospholipid metabolism |
| P7 | LysoPC(20:4) b | 6.92 | 588.3292 | C28H50NO7P | 2.44 | ↓ ** | ↓ | ↑ ** | Glycerophospholipid metabolism |
| P8 | LysoPC(16:0) b | 7.86 | 540.3302 | C24H50NO7P | 3.37 | ↓ * | ↑ * | ↑ | Glycerophospholipid metabolism |
| P16 | LysoPC(15:0) b | 7.87 | 480.3091 | C23H48NO7P | 4.33 | ↓ ** | ↑ | ↓ | Glycerophospholipid metabolism |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Jia, H.-M.; Cui, F.-X.; Yang, Y.; Zhao, Y.; Yang, M.-H.; Zou, Z.-M. The Effect of Chinese Herbal Medicine Formula mKG on Allergic Asthma by Regulating Lung and Plasma Metabolic Alternations. Int. J. Mol. Sci. 2017, 18, 602. https://doi.org/10.3390/ijms18030602
Yu M, Jia H-M, Cui F-X, Yang Y, Zhao Y, Yang M-H, Zou Z-M. The Effect of Chinese Herbal Medicine Formula mKG on Allergic Asthma by Regulating Lung and Plasma Metabolic Alternations. International Journal of Molecular Sciences. 2017; 18(3):602. https://doi.org/10.3390/ijms18030602
Chicago/Turabian StyleYu, Meng, Hong-Mei Jia, Feng-Xia Cui, Yong Yang, Yang Zhao, Mao-Hua Yang, and Zhong-Mei Zou. 2017. "The Effect of Chinese Herbal Medicine Formula mKG on Allergic Asthma by Regulating Lung and Plasma Metabolic Alternations" International Journal of Molecular Sciences 18, no. 3: 602. https://doi.org/10.3390/ijms18030602