
Defense Responses in Grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria Dieback Pathogens Neofusicoccum parvum and Diplodia seriata and Their Relationship with Flowering

 ,
,
Abstract
:1. Introduction
2. Results
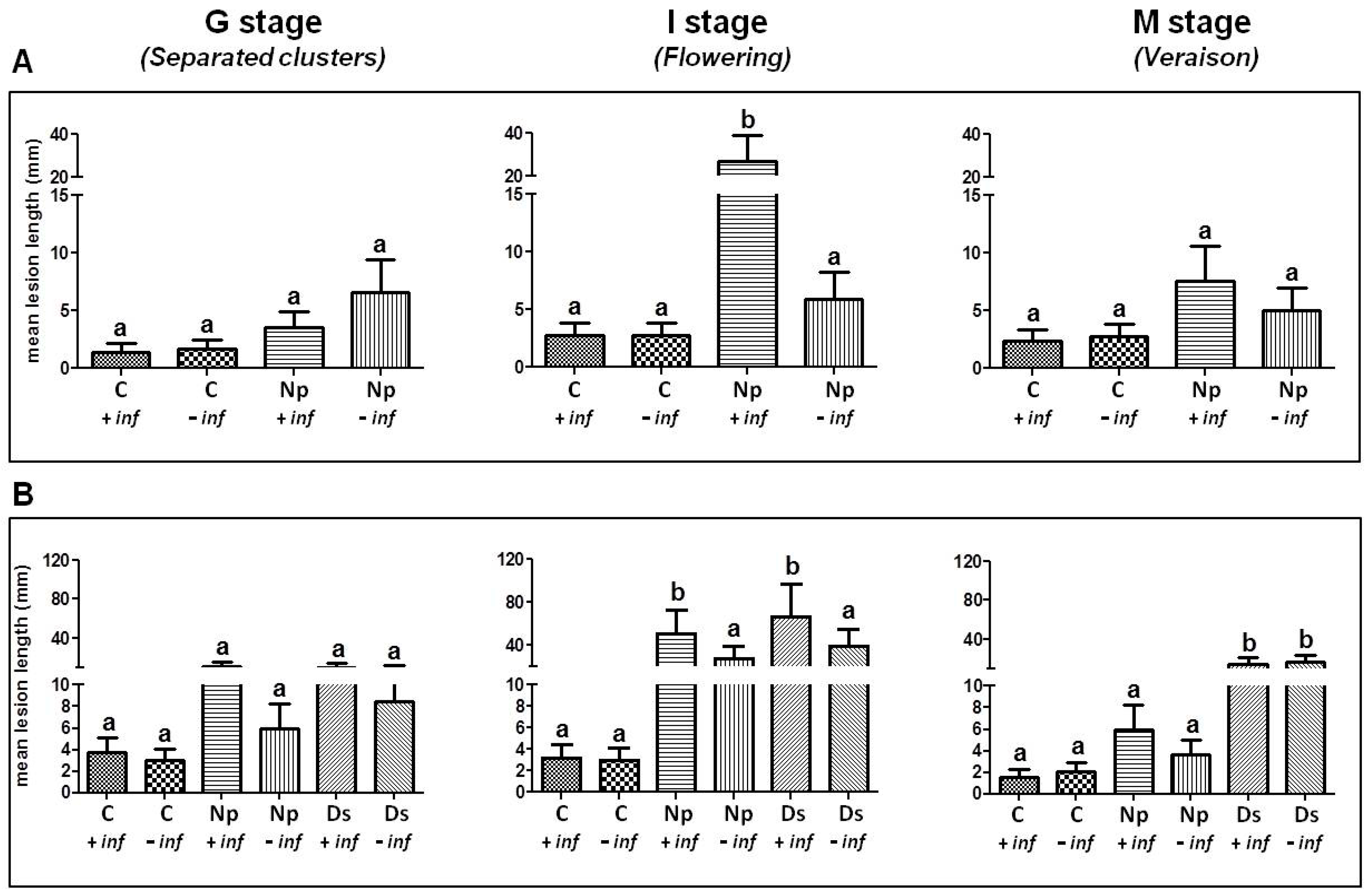
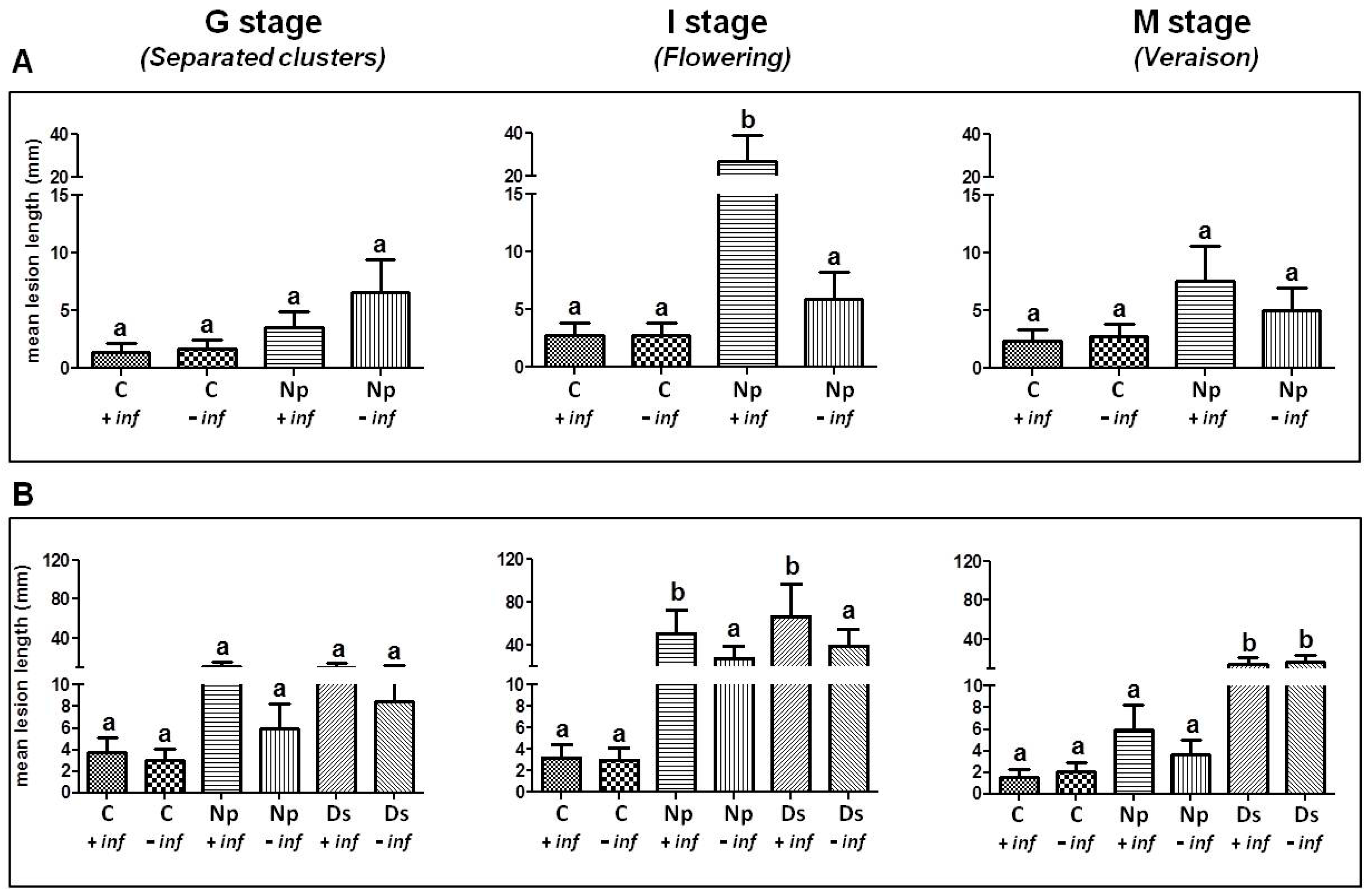
2.1. Pathogenicity Tests
2.1.1. Artificial Inoculation with N. parvum
2.1.2. Artificial Inoculation with D. seriata and N. parvum
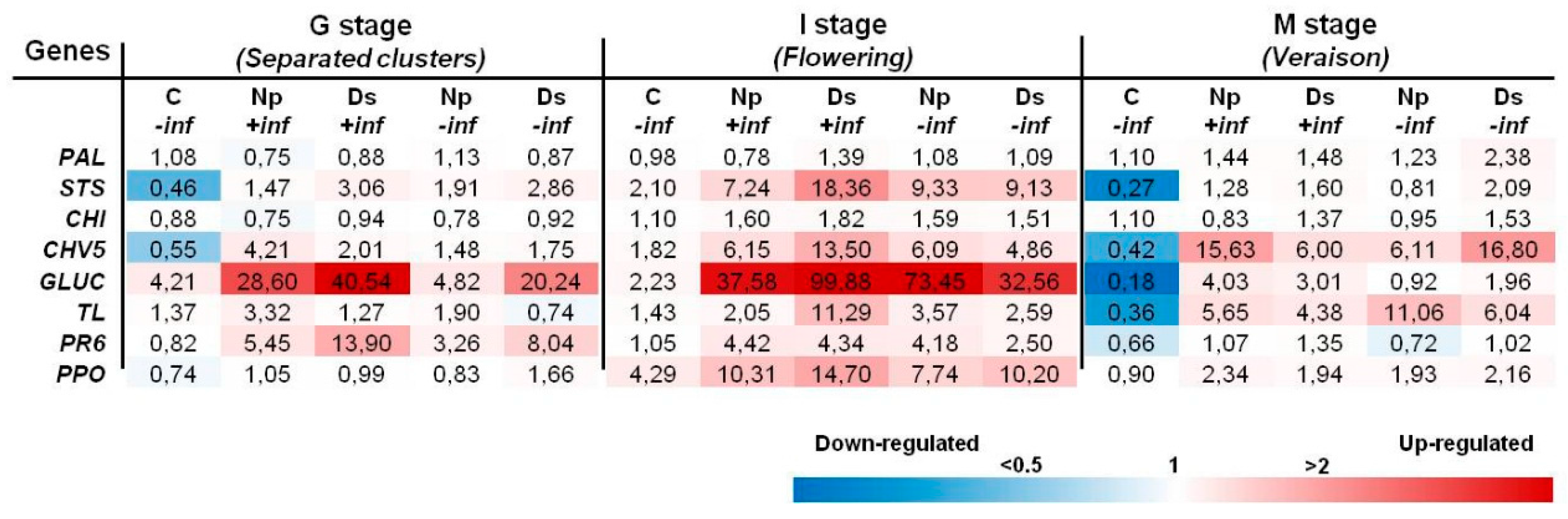
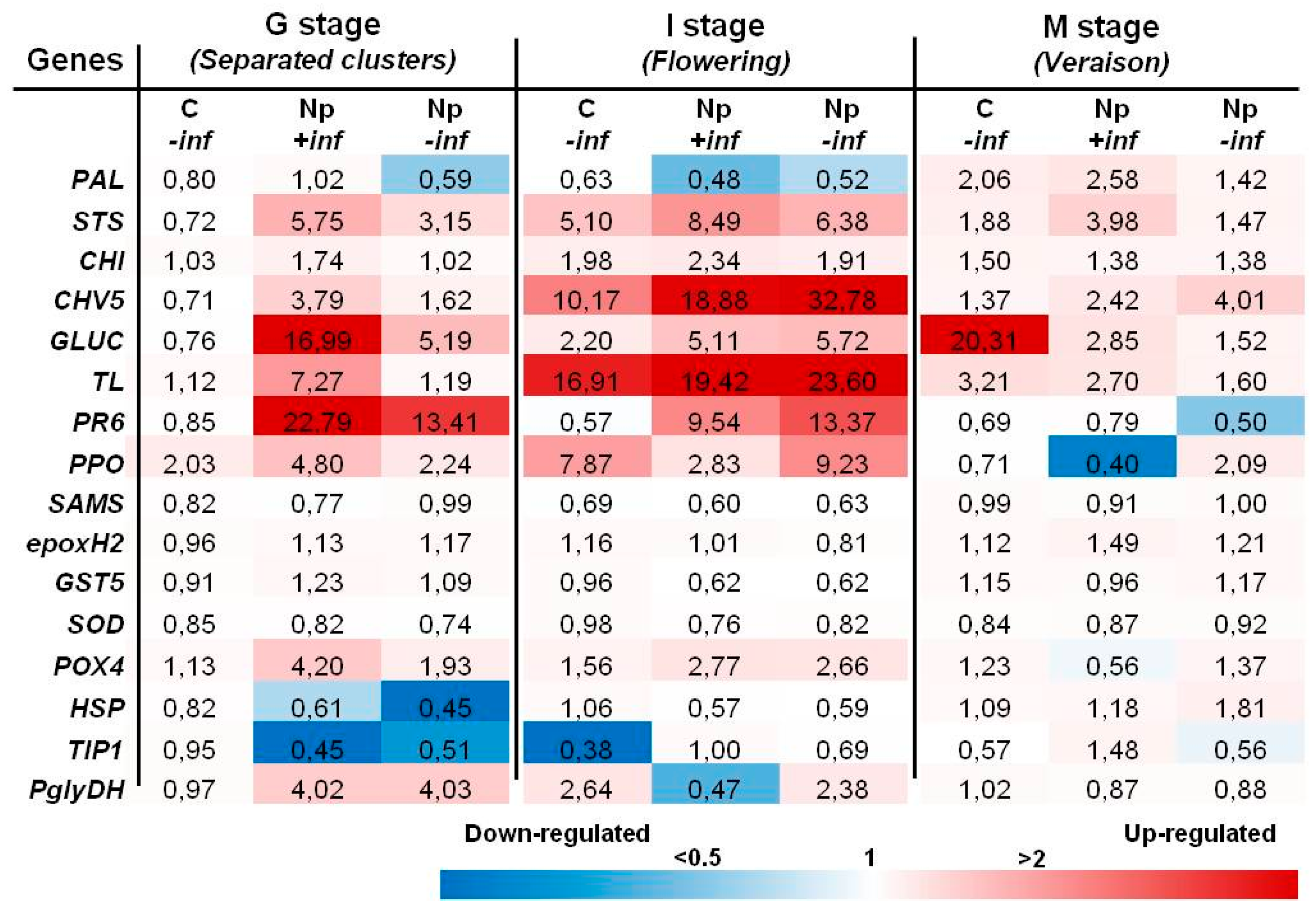
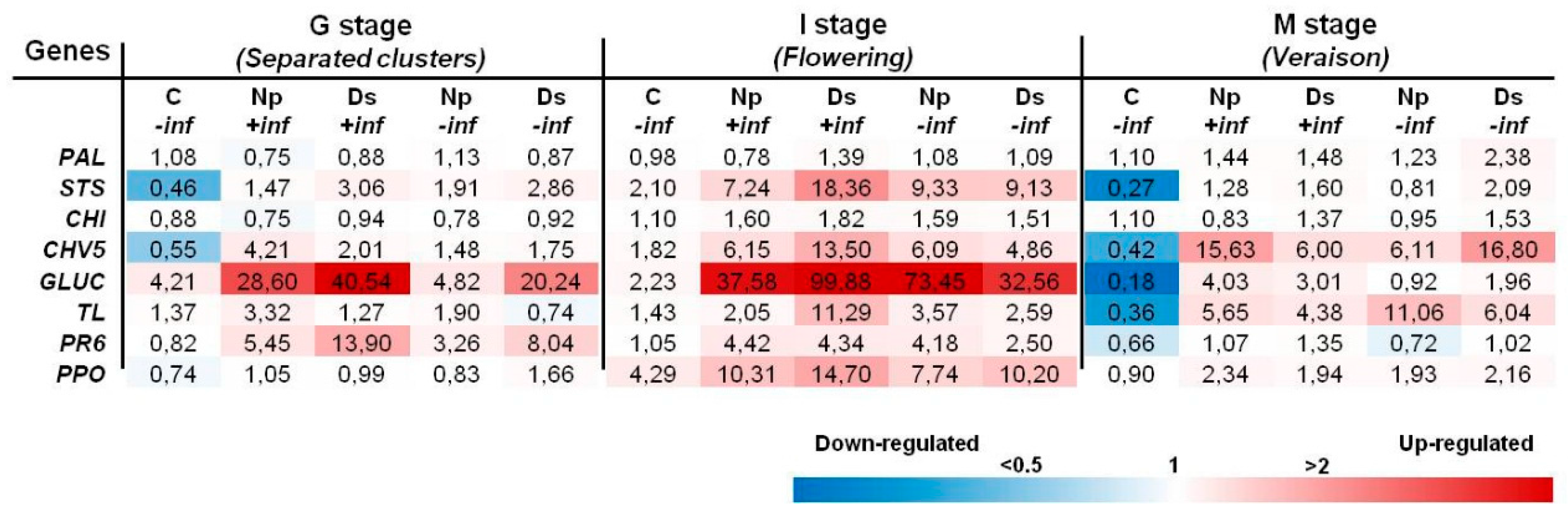
2.2. Transcript Analysis
2.2.1. Inoculations with N. parvum
2.2.2. Inoculations with D. seriata and N. parvum
3. Discussion
3.1. Pathogenicity Tests
3.2. Transcript Analyses
4. Materials and Methods
4.1. Plant Material, Fungal Strains and Artificial Inoculations
4.2. Experimental Design
4.3. Transcript Analysis
4.3.1. RNA Extraction
4.3.2. Real-Time RT-PCR Analysis of Gene Expression
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bertsch, C.; Ramirez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, C.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases. A Review, 1st ed.; OIV Publications: Paris, France, 2016. [Google Scholar]
- Di Marco, S.; Reggiori, F.; Baleani, M.; Benanchi, M.; Bossio, D.; Osti, F.; Mugnai, L. New wound infections are really relevant for grapevine leaf stripe disease? The case of Trichoderma pruning wound protection. Phytopathol. Mediterr. 2014, 53, 588–589. [Google Scholar]
- Úrbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, 5–45. [Google Scholar]
- Chethana, K.W.T.; Li, X.H.; Zhang, W.; Hyde, K.D.; Yan, J.Y. Trail of decryption of molecular research on Botryosphaeriaceae in woody plants. Phytopathol. Mediterr. 2016, 55, 147–171. [Google Scholar]
- Spagnolo, A.; Larignon, P.; Magnin-Robert, M.; Hovasse, A.; Cilindre, C.; van Dorsselaer, A.; Clément, C.; Schaeffer-Reiss, C.; Fontaine, F. Flowering as the most highly sensitive period of grapevine (Vitis vinifera L. cv. Mourvèdre) to the Botryosphaeria dieback agents Neofusicoccum parvum and Diplodia seriata infection. Int. J. Mol. Sci. 2014, 15, 9644–9669. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, J.M.; Calitz, F.J.; Halleen, F.; Fourie, P.H. Temporal spore dispersal patterns of grapevine trunk pathogens in South Africa. Eur. J. Plant Pathol. 2010, 127, 375–390. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Bruez, E.; Hurtado, J.; Gubler, W.D. Effect of temperature on conidial germination of Botryosphaeriaceae species infecting grapevines. Plant Dis. 2010, 94, 1476–1484. [Google Scholar] [CrossRef]
- Larignon, P.; Dubos, B. Fungi associated with esca disease in grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Kuntzmann, P.; Villaume, S.; Larignon, P.; Bertsch, C. Esca, BDA and Eutypiosis: Foliar symptoms, trunk lesions and fungi observed in diseased vinestocks in two vineyards in Alsace. Vitis 2010, 49, 71–76. [Google Scholar]
- Zufferey, V.; Murisier, F.; Vivin, P.; Belcher, S.; Lorenzini, F.; Spring, J.L.; Viret, O. Carbohydrate reserves in grapevine (Vitis vinifera L. “Chasselas”) the influence of the leaf to fruit ratio. Vitis 2012, 51, 103–110. [Google Scholar]
- Jermini, M.; Blaise, P.; Gessler, C. Quantification of the influence of the downy mildew (Plasmopara viticola) epidemics on the compensatory capacities of Vitis vinifera “Merlot” to limit the qualitative yield damage. Vitis 2010, 49, 153–160. [Google Scholar]
- Bolton, M.D. Primary metabolism and plant defense—Fuel for the fire. Mol. Plant Microbe Interact. 2009, 22, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, F. A global view of defense gene expression regulation—A highly interconnected signaling network. Curr. Opin. Plant Biol. 2004, 7, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, C.; Gomes, A.C.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur. J. Plant Pathol. 2016, 144, 707–721. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Spagnolo, A.; Boulanger, A.; Joyeux, C.; Clément, C.; Abou-Mansour, E.; Fontaine, F. Changes in plant metabolism and accumulation of fungal metabolites in response to esca proper and apoplexy expression in the whole grapevine. Phytopathology 2016, 106, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Magnin-Robert, M.; Letousey, P.; Spagnolo, A.; Rabenoelina, F.; Jacquens, L.; Mercier, L.; Clément, C.; Fontaine, F. Leaf strip form of esca induces alteration of photosynthesis and defence reactions in presymptomatic leaves. Funct. Plant Biol. 2011, 38, 856–866. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Spagnolo, A.; Alayi, T.D.; Cilindre, C.; Mercier, L.; Schaeffer-Reiss, C.; van Dorsselaer, A.; Clément, C.; Fontaine, F. Proteomic insights into changes in wood of Vitis vinifera L. in response to esca proper and apoplexy. Phytopathol. Mediterr. 2014, 53, 173–192. [Google Scholar]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Mercier, L.; Schaeffer-Reiss, C.; van Dorsselaer, A.; Clément, C.; Fontaine, F. Physiological changes in green stems of Vitis vinifera L. cv. Chardonnay in response to esca proper and apoplexy revealed by proteomic and transcriptomic analyses. J. Proteome Res. 2012, 11, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Lebon, G.; Wojnarowiez, G.; Holzapfel, B.; Fontaine, F.; Vaillant-Gaveau, N.; Clément, C. Sugars and flowering in the grapevine (Vitis vinifera L.). J. Exp. Bot. 2008, 59, 2565–2578. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.N.; Baillieul, F.; Vaillant-Gaveau, N.; Jacquens, L.; Conreux, A.; Jeandet, P.; Clément, C.; Fontaine, F. Low responsiveness of grapevine flowers and berries at fruit set to UV-C irradiation. J. Exp. Bot. 2009, 60, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Vaillant-Gaveau, N.; Maillard, P.; Wojnarowiez, G.; Gross, P.; Clement, C.; Fontaine, F. Inflorescence of grapevine (Vitis vinifera L.): A high ability to distribute its own assimilates. J. Exp. Bot. 2011, 62, 4183–4190. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vučetić, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of grapevine phenology to water availability, temperature and CO2 concentration. Front. Environ. Sci. 2016, 4. [Google Scholar] [CrossRef]
- Paolinelli-Alfonso, M.; Villalobos-Escobedo, J.M.; Rolshausen, P.; Herrera-Estrella, A.; Galindo-Sánchez, C.; López-Hernández, J.F.; Hernandez-Martinez, R. Global transcriptional analysis suggests Lasiodiplodia theobromae pathogenicity factors involved in modulation of grapevine defensive response. BMC Genom. 2016, 17, 615. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, B.P.; Smith, J.P. Developmental stage and climatic factors impact more on carbohydrate reserve dynamics of shiraz than cultural practice. Am. J. Enol. Vitic. 2012, 63, 333–342. [Google Scholar] [CrossRef]
- Pellegrino, A.; Clingeleffer, P.; Cooley, N.; Walker, R. Management practices impact vine carbohydrate status to a greater extent than vine productivity. Front. Plant Sci. 2014, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lu, J.; Xu, X.F.; Hu, R.L.; Pan, Y.J. pH-switched HRP-catalyzed dimerization of resveratrol: A selective biomimetic synthesis. Green Chem. 2012, 14, 3281–3284. [Google Scholar] [CrossRef]
- Berger, S.; Sinha, A.K.; Roitsch, T. Plant physiology meets phytopathology: Plant primary metabolism and plant–pathogen interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant–pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.; Magnin-Robert, M.; Nascimento, T.; Spagnolo, A.; Abou-Mansour, E.; Fioretti, C.; Clément, C.; Rego, C.; Fontaine, F. Reproducing Botryosphaeria dieback foliar symptoms in a simple model system. Plant Dis. 2016, 100, 1071–1079. [Google Scholar] [CrossRef]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, T.D.; Cilindre, C.; Schaeffer-Reiss, C.; van Dorsselaer, A.; Clément, C.; Larignon, P.; Ramirez-Suero, M.; Chong, J.; et al. Differential responses of three grapevine cultivars to Botryosphaeria dieback. Phytopathology 2014, 104, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.S.; Wang, L.J. The phenylpropanoid pathway and plant defence—A genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Bisson, J.; Waffo-Teguo, P.; Papastamoulis, Y.; Richard, T.; Corio-Costet, M.F.; Merillon, J.M.; Cluzet, S. Phenolics and their antifungal role in grapevine wood decay: Focus on the Botryosphaeriaceae family. J. Agric. Food Chem. 2012, 60, 11859–11868. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.R.M.; Dias, A.C.P. Phaeomoniella chlamydospora-induced oxidative burst in Vitis vinifera cell suspensions: Role of NADPH oxidase and Ca2+. J. Phytopathol. 2012, 160, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.L.; Heene, E.; Qiao, F.; Nick, P. The phytoalexin resveratrol regulates the initiation of hypersensitive cell death in Vitis cell. PLoS ONE 2011. [Google Scholar] [CrossRef] [PubMed]
- Valtaud, C.; Foyer, C.H.; Fleurat-Lessard, P.; Bourbouloux, A. Systemic effects on leaf glutathione metabolism and defence protein expression caused by esca infection in grapevines. Funct. Plant Biol. 2009, 36, 260–279. [Google Scholar] [CrossRef]
- Letousey, P.; Baillieul, F.; Perrot, G.; Rabenoelina, F.; Boulay, M.; Vaillant-Gaveau, N.; Clément, C.; Fontaine, F. Early events prior to visual symptoms in the apoplectic form of grapevine esca disease. Phytopathology 2010, 100, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Camps, C.; Kappel, C.; Lecomte, P.; Leon, C.; Gomes, E.; Coutos-Thevenot, P.; Delrot, S. A transcriptomic study of grapevine (Vitis vinifera cv. Cabernet-sauvignon) interaction with the vascular ascomycete fungus Eutypa lata. J. Exp. Bot. 2010, 61, 1719–1737. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Suero, M.; Benard-Gellon, M.; Chong, J.; Laloue, H.; Stempien, E.; Abou-Mansour, E.; Fontaine, F.; Larignon, P.; Mazet-Kieffer, F.; Farine, S.; et al. Extracellular compounds produced by fungi associated with Botryosphaeria dieback induce differential defence gene expression patterns and necrosis in Vitis vinifera cv. Chardonnay cells. Protoplasma 2014, 251, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M. Les stades repères dans le développement annuel de la vigne et leur utilisasion pratique. Rev. Romande d’Agric. d’Arboric. 1952, 8, 4–6. (In French) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Experiment | Condition | Sample Codes | ||
|---|---|---|---|---|
| G Stage (Separated Clusters) | I Stage (Flowering) | M Stage (Veraison) | ||
| Inoculation Trial No. 1 and 2 | Control | C − inf/C + inf | C − inf/C + inf | C − inf/C + inf |
| N. parvum strain Np SV | Np − inf/Np + inf | Np − inf/Np + inf | Np − inf/Np + inf | |
| Inoculation Trial No. 2 | D. seriata strain Bo98.1 | Ds − inf/Ds + inf | Ds − inf/Ds + inf | Ds − inf/Ds + inf |
| Function | Gene | Primer Sequences | GenBank or TC TIGR * Accession Number |
|---|---|---|---|
| Housekeeping genes | EF1 (EF1-α elongation factor) | 5′-GAACTGGGTGCTTGATAGGC-3′ | GU585871 |
| 5′-AACCAAAATATCCGGAGTAAAAGA-3′ | |||
| 60SRP (60S ribosomal protein L18) | 5′-ATCTACCTCAAGCTCCTAGTC-3′ | XM_002270599 | |
| 5′-CAATCTTGTCCTCCTTTCCT-3′ | |||
| Phenylpropanoid metabolism | CHI (Chalcone isomerase) | 5′-GCAGAAGCCAAAGCCATTGA-3′ | NM_001281104 |
| 5′-GCCGATGATGGACTCCAGTAC-3′ | |||
| PAL (Phenylalanine ammonia lyase) | 5′-TCCTCCCGGAAAACAGCTG-3′ | X75967 | |
| 5′-TCCTCCAAATGCCTCAAATCA-3′ | |||
| POX4 (Peroxidase-like 4) | 5′-AACATCCCCCCTCCCACTT-3′ | XM_002269882 | |
| 5′-TGCATCTCGCTTGGCCTATT-3′ | |||
| STS (Stilbene synthase) | 5′-AGGAAGCAGCATTGAAGGCTC-3′ | FJ851185 | |
| 5′-TGCACCAGGCATTTCTACACC-3′ | |||
| Defense protein | CHV5 (Chitinase class v) | 5′-CTACAACTATGGCGCTGCTG-3′ | AF532966 |
| 5′-CCAAAACCATAATGCGGTCT-3′ | |||
| GLUC (β-1,3 glucanase) | 5′-TCAATGGCTGCAATGGTGC-3′ | DQ267748 | |
| 5′-CGGTCGATGTTGCGAGATTTA-3′ | |||
| PPO (Polyphenol oxidase) | 5′-TGGTCTTGCTGATAAGCCTAGTGA-3′ | XM_002727606 | |
| 5′-TCCACATCCGATCGACATTG-3′ | |||
| PR6 (Serine-protease inhibitor 6) | 5′-AGGGAACAATCGTTACCCAAG-3′ | AY156047 | |
| 5′-CCGATGGTAGGGACACTGAT-3′ | |||
| SAMS (S-adenosylmethionine synthetase) | 5′-CCTGAAATCAAAGTTCTCCTTCACA-3′ | XM_002266322 | |
| 5′-CCGGGCCTGAAATCAAAGTT-3′ | |||
| TL (Thaumatin-like) | 5′-CCTAACACCTTAGCCGAATTCGC-3′ | AF532965 | |
| 5′-GGCCATAGGCACATTAAATCCATC-3′ | |||
| Detoxification and Stress tolerance | epoxH2 (Epoxide hydrolase 2) | 5′-TCTGGATTCCGAACTGCATTG-3′ | XM_002270484 |
| 5′-ACCCATGATTAGCAGCATTGG-3′ | |||
| GST5 (Glutathione s-transferase 5) | 5′-GCAGAAGCTGCCAGTGAAATT-3′ | XM_002277883 | |
| 5′-GGCAAGCCATGAAAGTGACA-3′ | |||
| HSP (alpha crystalline heat shock protein) | 5′-TCGGTGGAGGATGACTTGCT-3′ | XM_002272382 | |
| 5′-CGTGTGCTGTACGAGCTGAAG-3′ | |||
| SOD (Superoxide dismutase) | 5′-GTGGACCTAATGCAGTGATTGGA-3′ | AF056622 | |
| 5′-TGCCAGTGGTAAGGCTAAGTTCA-3′ | |||
| Primary metabolism | PglyDH (Phosphoglycerate dehydrogenase) | 5′-CGTCGAAGATGCTCAATGATGA-3′ | XM_002285322 |
| 5′-CCCCCACGAGCAACATTAATT-3′ | |||
| Water stress | TIP1 (Tonoplast intrinsic protein) | 5′-ATCACCAACCTCATTCATATGC-3′ | AF271661 |
| 5′-GTTGTTGTCTCAACCCATTTCC-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spagnolo, A.; Mondello, V.; Larignon, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Fontaine, F. Defense Responses in Grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria Dieback Pathogens Neofusicoccum parvum and Diplodia seriata and Their Relationship with Flowering. Int. J. Mol. Sci. 2017, 18, 393. https://doi.org/10.3390/ijms18020393
Spagnolo A, Mondello V, Larignon P, Villaume S, Rabenoelina F, Clément C, Fontaine F. Defense Responses in Grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria Dieback Pathogens Neofusicoccum parvum and Diplodia seriata and Their Relationship with Flowering. International Journal of Molecular Sciences. 2017; 18(2):393. https://doi.org/10.3390/ijms18020393
Chicago/Turabian StyleSpagnolo, Alessandro, Vincenzo Mondello, Philippe Larignon, Sandra Villaume, Fanja Rabenoelina, Christophe Clément, and Florence Fontaine. 2017. "Defense Responses in Grapevine (cv. Mourvèdre) after Inoculation with the Botryosphaeria Dieback Pathogens Neofusicoccum parvum and Diplodia seriata and Their Relationship with Flowering" International Journal of Molecular Sciences 18, no. 2: 393. https://doi.org/10.3390/ijms18020393