The Role of Non-Coding RNAs in Cytoplasmic Male Sterility in Flowering Plants

Institute of Experimental Botany of the Czech Academy of Sciences, Rozvojová 263, 16502 Prague, Czech Republic
Int. J. Mol. Sci. 2017, 18(11), 2429; https://doi.org/10.3390/ijms18112429
Submission received: 1 November 2017
/
Revised: 13 November 2017
/
Accepted: 14 November 2017
/
Published: 16 November 2017
(This article belongs to the Special Issue Plant Mitochondria)
Abstract
:The interactions between mitochondria and nucleus substantially influence plant development, stress response and morphological features. The prominent example of a mitochondrial-nuclear interaction is cytoplasmic male sterility (CMS), when plants produce aborted anthers or inviable pollen. The genes responsible for CMS are located in mitochondrial genome, but their expression is controlled by nuclear genes, called fertility restorers. Recent explosion of high-throughput sequencing methods enabled to study transcriptomic alterations in the level of non-coding RNAs under CMS biogenesis. We summarize current knowledge of the role of nucleus encoded regulatory non-coding RNAs (long non-coding RNA, microRNA as well as small interfering RNA) in CMS. We also focus on the emerging data of non-coding RNAs encoded by mitochondrial genome and their possible involvement in mitochondrial-nuclear interactions and CMS development.
1. Introduction
In plants, male sterility refers to the inability to generate viable pollen. It is encoded by nuclear genes leading to the genic male sterility (GMS) or by mitochondrial genes interacting with nuclear genes resulting in the development of cytoplasmic male sterility (CMS). Both kinds of male sterility are broadly utilized in agriculture for the production of hybrid crops providing higher yield than inbred parents [1]. The existence of male-sterile lines eliminates the need for laborious sterilization in a long array of crops including rice (Oryza sativa), maize (Zea mays), wheat (Triticum aestivum), sorghum (Sorghum bicolor), sunflower (Helianthus annuus) and sugar beet (Beta vulgaris). Despite more than two centuries of research and high economic importance, CMS mechanisms remain poorly understood. CMS represents a special case of mitochondrial-nuclear interaction, which is regulated at multiple levels. New discoveries highlight the contributions of non-coding RNAs—a genomic “dark matter” [2]—to the complex regulatory network controlling CMS. In this review, I discuss recent observations and evidence for the action of various classes of plant non-coding RNAs in CMS biogenesis and pollen development.
2. Mitochondrial CMS Genes and Their Mode of Action
The mitochondrial and nuclear genes involved in CMS biogenesis or associated with CMS are very diverse [3,4]. Mitochondrial CMS genes are often chimeric, comprised of pieces of essential genes or unknown open reading frames) (ORF) [5,6,7,8]. Chimeric CMS genes can be generated by intramolecular recombination events. For example, maize male-sterile Texas (CMS-T), one of the first CMS lines used in agriculture, possesses T-urf13 gene [9], derived from at least seven recombination events involving atp6 and rrn26 mitochondrial genes. Functional copies of atp6 and rrn26 remain in another part of the mitochondrial genome [9]. Another example of a mitochondrial CMS gene is a mutation in cox2 encoding the cytochrome c oxidase subunit 2 truncated protein appearance [10]. CMS genes are very diverse not only across angiosperms, but also within species. For example, numerous CMS systems were described in rice [11,12], maize [9,13], and sugar beet [10,14]. Accordingly, we may expect them to employ similarly diverse modes of action.
The precise details how CMS gene expression impairs mitochondria and the pollen development is not known, but several models have been proposed. Mitochondrial CMS genes may code for cytotoxic proteins like URF13 in maize CMS-T [9], or they may cause energy deficiency during energetically highly energetically demanding male (but not female) reproductive development. Many CMS proteins are hydrophobic and could interfere with oxidative phosphorylation (OXPHOS) complexes within inner mitochondrial membrane [14,15,16], which may decrease the ATP production. Another mechanism can be a premature or delayed programmed cell death (PCD) of the tapetum, the innermost cell layer of the anther wall [17], crucial for the pollen development [18]. PCD of the tapetum, which provides nutrients for the pollen maturation, must be properly timed. CMS genes are often transcribed both in vegetative tissues and anthers, but the respective proteins are produced only at a specific time and tissue. Rice CMS-WA [19] provides a clear example: the WA352 protein accumulates only in tapetal cells and only at the microspore mother cell stage, although WA352 transcripts are constitutively present in all tissues. Accurate spatiotemporal patterning of CMS-associated mitochondrial genome expression requires fine-tuned regulation at transcriptional, post-transcriptional, translational and post-translational levels. This is achieved by employing plethora of transcription factors and regulatory non-coding RNAs which affect transcript longevity and translation efficiency [2,20].
3. Restoration of Fertility by Nuclear Genes
The sterility effects of the mitochondrial CMS genes may be inhibited by the nuclear Restorer of fertility (Rf) genes, which re-enable the development of functional anthers, fertile pollen, and hermaphroditic flowers [21]. The Rf genes suppress CMS genes’ sterilizing effect by degrading or cleaving their mRNAs [22,23], or by post-transcriptional modification including the RNA editing [24]. Alternatively, either the translation of CMS-associated transcript may be blocked [19,25], or the CMS protein degraded [26].
The majority of Rf genes belong to the large family of Pentatricopeptide Repeat (PPR) genes [27]. This gene family, highly expanded in flowering plants, controls multiple aspects of the organellar gene expression, including RNA editing, RNA stabilization and processing, and translation initiation. All PPR proteins contain P-type 35 amino acid domains, each of which recognizes a single nucleotide of RNA [28]. The Rf genes constitute a specific PPR subfamily called Restorer of fertility-like (RFL), which shows an accelerated evolutionary rate [29], and numerous domain-level recombination events [30]. Mitochondrial CMS genes and RFL genes may co-evolve similarly to a host-pathogen system [31].
In keeping with their tremendous diversity, not all Rf genes code for PPR proteins. The Rf2 gene in maize CMS-T encodes mitochondrial aldehyde dehydrogenase and restores male fertility at a metabolic level [32,33]. Another example is the restoration factor Rf17 in rice, bearing protein sequence similarity with acyl-carrier proteins [34], which restores male fertility by retrograde mitochondrial-nuclear signaling pathway.
Whereas CMS has been well studied in agricultural plants, its occurrence in the remaining species has been under the less attention. CMS forms the basis for the widespread plant reproduction system-gynodioecy, characterized by the co-occurrence of male-sterile (female) and hermaphrodite individuals in the same populations [35]. However, only a handful CMS systems from natural populations were studied at the molecular level [36,37,38,39]. Domestication is associated with a strong selection for beneficial features which also results in the loss of genetic diversity. We may therefore expect an even more diverse collection of CMS-associated genes in the wild than in agricultural species investigated so far.
4. Non-Coding RNAs in Pollen Development and CMS
CMS as a specific case of mitochondrial-nuclear interaction is a complex phenomenon which has to be tightly regulated at multiple levels. Its intricacy became apparent with the recent onset of high throughput methods which enabled complete genome and global transcriptome sequencing [40,41,42,43]. Besides long studied transcription factors [44,45], non-coding RNAs (ncRNAs) became to draw attention.
Non-coding RNAs contain no large open reading frame (ORF) and are therefore presumed not to encode proteins. They comprise well-studied structural RNAs, such as rRNA, tRNA, snoRNA, snRNA etc., and regulatory RNA. The latter are either longer than 200 nt (long non-coding RNA-lncRNA), or shorter (small RNA–sRNA) [20,46]. Among sRNAs, microRNAs (miRNAs) are the known regulators of gene expression at the post-transcriptional level [47,48]. Another subclass of ncRNAs is represented by small-interfering RNAs (siRNAs), which are involved in the defense against viruses and mobile elements [49,50,51]. Next, trans-acting small interfering RNAs (ta-siRNAs) are endogenous regulatory elements participating in complex signaling networks [52]. Of all the classes of regulatory RNAs, only the function of miRNAs in CMS biogenesis has been investigated in detail.
5. miRNAs
miRNAs are small RNAs (about 21 nt) that guide the RNA-induced silencing complex (RISC) to the target transcripts, inducing their cleavage or translational inhibition [47]. They are present in most eukaryotic organisms, but their biogenesis and signaling pathways notably differ between plants and animals [53]. miRNAs are encoded by their own genes at various genomic loci. RNA polymerase II generates a long primary miRNA transcript (pri-miRNA), which contains a hairpin with the miRNA sequence. Plant pri-miRNAs are cut in the nucleus by the complex comprised of RNase DICER LIKE1 (DCL1) and additional proteins e.g., HYPONASTIC LEAVES1 (HYL1) and SERRATE (SE) [54], producing the miRNA duplex. This duplex is transported to the cytoplasm, where it interacts with ARGONAUTE1 (AGO1) to form RISC and to guide it to target genes (Figure 1). Some miRNAs are evolutionary conserved, but many are species-specific [55].
Plant miRNAs control multiple aspects of plant development and stress response, including shoot and root apical development, leaf and trichome development, floral transition and fruit size as well as nutrition-, drought-, salinity- and heat-stress responses [48]. They are also prominent regulators of the pollen development [56]. In the last decade, numerous studies have compared the microtranscriptomes of CMS and fertile lines of agricultural species. (Table 1 and Table S1).
Hundreds of miRNAs have been identified by these studies, many of them belonging to the novel ones. Some miRNAs were differentially expressed between sterile and fertile lines (e.g., 47 in Brassica juncea [57]; 42 in cybrid pummelo (Citrus grandis) [58]; 87 in Brassica rapa CMS-Ogura [59]). Evolutionarily conserved miR156/7a targeting SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL), which regulates flowering, leaf shape and also tapetum development, was frequently present among differentially expressed miRNAs. Other examples of miRNAs involved in pollen development are miR166 targeting the transcription factor HD-ZIPIII [60], or miR167 targeting AUXIN RESPONSE FACTOR (ARF) genes, which control anther dehiscence [57]. A broad array of metabolic processes-catabolism of fatty acids [61], sugar transport [61] or inorganic phosphate homeostasis [58] occurring in anthers were affected by the differentially expressed miRNAs. Differentially expressed genes identified by the comparison between the cytoplasmic mRNA-derived transcriptomes of CMS and fertile lines belonged to the similar functional categories as differentially expressed miRNAs. They were involved in starch and sucrose metabolism, amino acid and sulphur metabolism, flavonoid biosynthesis, or pollen development [62,63].
Whereas the impact of mitochondrial CMS genes on the global transcriptome is well described in many crops, the retrograde signal which communicates the mitochondrial impairments to the nucleus of tapetal or pollen cells and triggers diverse cascades of regulatory elements is still elusive. The recently described NAC transcription factor ANAC017 [64] is not the only factor responsible for the mitochondrial retrograde signaling [65] and the mediators communicating between mitochondria and nucleus in CMS are yet to be discovered. In parallel with animal mitochondria [2], we may assume that not only proteins, but also ncRNAs may convey retrograde signals (Figure 1).
6. siRNAs and ta-si RNAs
Unlike miRNAs, siRNAs mediate the silencing of the same genes from which they originate [53]. They are typically 20–24 nt long and are cleaved from a long precursor dsRNA, which may be derived from viruses, transposons or the combination of a sense and antisense transcript. They guide RISC complex to degrade complementary RNA of viral or endogenous origin, or inhibit translation of respective mRNAs. Alternatively, siRNAs mediate de novo modification to form transcriptionally inactive chromatin by recruiting DNA- and histone-modifying enzymes to the specific chromosomal targets [66]. Plant siRNAs may move from cell to cell, but also at longer distance through plasmodesmata or the vascular phloem tissue [53]. They facilitate the communication among individual organs, fine-tuning the response to environmental cues.
The complex interplay between siRNAs and miRNAs is illustrated by the action of ta-si RNAs. They are produced from the transcripts of TRANS-ACTING SIRNA (TAS) genes which are initially cleaved by the specific miRNAs. Instead being degraded, TAS cleavage products are transcribed by the RNA-dependent polymerase and subsequently diced by the DCL4 complex. The resulting siRNAs have a phased pattern, starting at the miRNA cleavage site. They may target the parental or different genes, frequently the members of large gene families [67,68].
The miR173-TAS1/2-PPR ta-si pathway has been described in Arabidopsis thaliana [68,69]. It is triggered by miR173 which initiates the cleavage of TAS1/2 transcripts and the subsequent production of ta-si RNAs targeting selected PPR genes. This pathway is highly conserved across angiosperms [70]. Considering the pervasive influence of PPR genes on mitochondrial metabolism, the participation of ta-si RNA in CMS biogenesis is highly plausible, but has not yet been demonstrated. Another example of ta-si pathway which may play a role in the CMS biogenesis is the miR390-TAS3 module. It may influence pollen development by modulating ARF gene expression [68].
7. lncRNA
The field of lncRNAs research has expanded and accelerated recently [71,72]. lncRNAs represent the most diverse class of regulatory ncRNA. They are capped and polyadenylated [73,74]; some of them, however, do not contain poly(A) tail [75]. Their modes of action are very diverse. They may be produced as antisense transcripts and to inhibit sense transcription (e.g., COOLAIR, a cold-induced antisense transcript of the floral inhibitor FLOWERING LOCUS C (FLC) in A. thaliana [76]).
Some lncRNAs influence alternative splicing. They interact with the nuclear speckle RNA-binding protein (NSR), which forms complexes with pre-mRNAs. At least two lncRNAs compete with target pre-mRNAs for NSR and modify their alternative splicing during lateral root formation [77].
The chromatin remodeling caused by lncRNA was also described. The lncRNA COLDAIR is transcribed from the intron of the FLC gene under cold temperature. It interacts with Polycomb Repressive Complex 2 (PRC2), recruits it to FLC and induces FLC repression. Silencing the floral inhibitor FLC activates flowering in the course of vernalization [78].
lncRNAs often affect the male fertility and the pollen development. Ma and coworkers [79] reported that lnc transcript zm401 was essential for the tapetum and pollen development in maize. Long Day Specific Male Fertility Associated RNA (LDMAR) encodes a 1236 nt lncRNA necessary for male fertility under long days. A mutation reducing LDMAR transcript levels leads to premature PCD in anthers and male sterility. This example refers to GMS and not to CMS, as no mitochondrial genes are involved in male sterility.
Some lncRNAs harboring miRNA binding sites function as endogenous target mimics (eTMs) to reduce the repression imposed by miRNAs [80] and to affect the reproductive development–e.g., osa-eTM160 and ath-eTM160 in rice and A. thaliana, respectively. They attenuate the repression imposed by miR160 on ARFs, which leads to the failure of pollen production [80,81].
The crosstalk between lncRNA and siRNA biogenesis has been reported in rice. lncRNAs serve as the source of siRNAs associated with the MEIOSIS ARRESTED AT LEPTOTENE1 (MEL1) transcript [82]. MEL1 protein is necessary for the meiotic progress and its loss of function results in aberrant vacuolation of spore mother cells and impaired male fertility.
Global analyses of long non-coding transcriptomes have greatly expanded our understanding of lncRNA function. The regulatory role of lncRNA in the course of fruit development of hot peppers is particularly known [83]. However, no comprehensive study of lncRNA participation in CMS biogenesis has been published.
8. Non-Coding RNAs Encoded by the Mitochondrial Genome
Whereas knowledge about the regulatory functions of plant nuclear-encoded ncRNAs has been steadily accumulating [47,53,71,72], evidence about the role of ncRNAs encoded by organellar genomes remains sparse. Dietrich and coworkers [84] provided an overview of the functions of organellar ncRNA in plants and animals, including the well-documented regulatory roles of both small and long ncRNAs in animal mitochondria and plant chloroplasts, but they provide only a few candidates of plant mitochondrial ncRNA. Additional examples of mitochondrial ncRNA were described in the comprehensive review on plant organelle biogenesis by Rurek [85]. Small RNAs may be the products of degradation of longer transcripts [86], or may arise due to the relaxed transcription of intergenic regions [75]. As mitochondrial DNA is often transferred to the nucleus [87], a nuclear origin for the already reported mitochondrial ncRNA cannot be excluded.
Ruwe and coworkers [88] described sRNA clusters near the 3′ ends of mitochondrial transcripts in A. thaliana. They can stabilize the transcripts or compete for PPR proteins.
A non-coding mitochondrial transcript about 500 nt long was reported by Holec and coworkers [89] in A. thaliana. It carried short stretches of sequence homology with 18S rRNA and tRNA, and exhibited editing sites. No conclusion about its possible function was drawn.
lncRNA accumulated and edited preferentially in male sterile plants (but not in their restored siblings) was documented in bladder campion (Silene vulgaris) [90]. Its sequence was not similar to any known sequence in GenBank, it was transcribed from its own promoter. Although it cannot be determined whether this lncRNA is the molecular cause or a consequence of male sterility, it becomes a very first example of mitochondrial ncRNA molecule associated with CMS in plants.
9. Future Perspectives
The role of nucleus-encoded miRNAs in CMS has been addressed by numerous studies, as documented in Table 1. However, the function of other classes of regulatory ncRNAs requires additional investigation. In addition to performing new experiments, existing data should be reexamined for the current insights. For example, data sets utilized for the analyses of miRNAs, may be used to study siRNAs in CMS biogenesis. Similarly, CMS-related transcriptomes constructed from polyA-enriched or rRNA-depleted samples may provide information about novel lncRNAs. Given the near-ubiquity of ncRNA involvement in plant development and stress response [48,53,71], investigating their role in CMS should be a priority.
Whereas a plethora of transcription factors and miRNAs induced by CMS and influencing plant metabolism, including mitochondrial functions, have been reported (Table 1), very little is known about retrograde signaling from mitochondria to nucleus (Figure 1). The application of genome editing and the existence loss-of-function mutations in candidate genes in many species with CMS make it possible to reveal the novel candidate genes involved in retrograde signaling during CMS.
Agricultural species went through genetic bottlenecks in the course of domestication which decreased their genetic variation in many genomic regions. As the studies of CMS have been performed primarily in crops, many important aspects or features may have been missed. The investigations of plants from natural populations will likely reveal novel characteristics associated with CMS [36,37,90].
Supplementary Materials
Supplementary materials can be found at www.mdpi.com/1422-0067/18/11/2429/s1.
Acknowledgments
The author thanks James D. Stone for valuable comments and linguistic correction, and Filip Štorch for help with drawing the figure. Financial support was provided by the grant of the Grant Agency of the Czech Republic 16-09220S.
Author Contributions
Helena Štorchová is the only author of this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, R.; Marine, J.C.; Leucci, E. Non-coding RNAs: The dark side of nuclear-mitochondrial communication. EMBO J. 2017, 36, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Horn, R.; Gupta, K.J.; Colombo, N. Mitochondrion role in molecular basis of cytoplasmic male sterility. Mitochondrion 2014, 19, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Touzet, P.; Meyer, E.H. Cytoplasmic male sterility and mitochondrial metabolism in plants. Mitochondrion 2014, 19, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Bentolila, S. Interactions of mitochondrial and nuclear genes that affect male gametophyte development. Plant Cell 2004, 16, S154–S169. [Google Scholar] [CrossRef] [PubMed]
- Gillman, J.D.; Bentolila, S.; Hanson, M.R. The petunia restorer of fertility protein is part of a large mitochondrial complex that interacts with transcripts of the CMS-associated locus. Plant J. 2007, 49, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kang, J.G.; Kim, B.D. Isolation and characterization of the cytoplasmic male sterility-associated orf456 gene of chili pepper (Capsicum annuum L.). Plant Mol. Biol. 2007, 63, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Duroc, Y.; Hiard, S.; Vrielynck, N.; Ragu, S.; Budar, F. The Ogura sterility-inducing protein forms a large complex without interfering with the oxidative phosphorylation components in rapeseed mitochondria. Plant Mol. Biol. 2009, 70, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Dewey, R.E.; Timothy, D.H.; Levings, C.S. A mitochondrial protein associated with cytoplasmic male-sterility in the T-cytoplasm of maize. Proc. Natl. Acad. Sci. USA 1987, 84, 5374–5378. [Google Scholar] [CrossRef] [PubMed]
- Ducos, E.; Touzet, P.; Boutry, M. The male sterile G cytoplasm of wild beet displays modified mitochondrial respiratory complexes. Plant J. 2001, 26, 171–180. [Google Scholar] [CrossRef]
- Itabashi, E.; Kazama, T.; Toriyama, K. Characterization of cytoplasmic male sterility of rice with Lead Rice cytoplasm in comparison with that with Chinsurah Boro II cytoplasm. Plant Cell Rep. 2009, 28, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gao, F.; Ji, Y.X.; Liu, Y.; Dan, Z.W.; Yang, P.F.; Zhu, Y.G.; Li, S.Q. ORFH79 impairs mitochondrial function via interaction with a subunit of electron transport chain complex III in Honglian cytoplasmic male sterile rice. New Phytol. 2013, 198, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Zabala, G.; Gabay-Laughnan, S.; Laughnan, J.R. The nuclear gene Rf3 affects the expression of the mitochondrial chimeric sequence R implicated in S-type male sterility in maize. Genetics 1997, 147, 847–860. [Google Scholar] [PubMed]
- Yamamoto, M.P.; Kubo, T.; Mikami, T. The 5′-leader sequence of sugar beet mitochondrial atp6 encodes a novel polypeptide that is characteristic of Owen cytoplasmic male sterility. Mol. Genet. Genom. 2005, 273, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Grelon, M.; Budar, F.; Bonhomme, S.; Pelletier, G. Ogura cytoplasmic male sterility (CMS)-associated ORF138 is translated into a mitochondrial-membrane polypeptide in male-sterile Brassica cybrids. Mol. Gen. Genet. 1994, 243, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Kazama, T.; Itabashi, E.; Fujii, S.; Nakamura, T.; Toriyama, K. Mitochondrial ORF79 levels determine pollen abortion in cytoplasmic male sterile rice. Plant J. 2016, 85, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Balk, J.; Leaver, C.J. The PET1-CMS mitochondrial mutation in sunflower is associated with premature programmed cell death and cytochrome c release. Plant Cell 2001, 13, 1803–1818. [Google Scholar] [CrossRef] [PubMed]
- Twell, D. Male gametogenesis and germline specification in flowering plants. Sex. Plant Reprod. 2011, 24, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.P.; Xu, H.; Liu, Z.L.; Guo, J.X.; Li, H.Y.; Chen, L.T.; Fang, C.; Zhang, Q.Y.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, S.; Li, J.R.; Sun, Q.W. Functions of plants long non-coding RNAs. Biochim. Biophys. Acta 2016, 1859, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Wise, R.P. The molecular basis of cytoplasmic male sterility and fertility restoration. Trends Plant Sci. 1998, 3, 175–180. [Google Scholar] [CrossRef]
- Sabar, M.; Gagliardi, D.; Balk, J.; Leaver, C.J. ORFB is a subunit of F1FO-ATP synthase: Insight into the basis of cytoplasmic male sterility in sunflower. EMBO Rep. 2003, 4, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Kazama, T.; Nakamura, T.; Watanabe, M.; Sugita, M.; Toriyama, K. Suppression mechanism of mitochondrial ORF79 accumulation by Rf1 protein in BT-type cytoplasmic male sterile rice. Plant J. 2008, 55, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Mitra, J.; Bhattacharyya, J.; Pradhan, S.; Sikdar, N.; Das, S.; Chakraborty, S.; Kumar, S.; Lakhanpaul, S.; Sen, S.K. Transgenic expression of an unedited mitochondrial orfB gene product from wild abortive (WA) cytoplasm of rice (Oryza sativa L.) generates male sterility in fertile rice lines. Planta 2015, 241, 1463–1479. [Google Scholar] [CrossRef] [PubMed]
- Uyttewaal, M.; Arnal, N.; Quadrado, M.; Martin-Canadell, A.; Vrielynck, N.; Hiard, S.; Gherbi, H.; Bendahmane, A.; Budar, F.; Mireau, H. Characterization of Raphanus sativus pentatricopeptide repeat proteins encoded by the fertility restorer locus for Ogura cytoplasmic male sterility. Plant Cell 2008, 20, 3331–3345. [Google Scholar] [CrossRef] [PubMed]
- Sarria, R.; Lyznik, A.; Vallejos, C.E.; Mackenzie, S.A. A cytoplasmic male sterility-associated mitochondrial peptide in common bean is post-translationally regulated. Plant Cell 1998, 10, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Gaborieau, L.; Brown, G.G.; Mireau, H. The propensity of pentatricopeptide repeat genes to evolve into restorers of cytoplasmic male sterility. Front. Plant Sci. 2016, 7, 1816. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Haertel, B.; Brennicke, A. RNA Editing in plants and its evolution. Annu. Rev. Genet. 2013, 47, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Bond, C.S.; Small, I.D. Selection patterns on restorer-like genes reveal a conflict between nuclear and mitochondrial genomes throughout angiosperm evolution. Proc. Natl. Acad. Sci. USA 2011, 108, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Melonek, J.; Stone, J.D.; Small, I. Evolutionary plasticity of restorer-of-fertility-like proteins in rice. Sci. Rep. 2016, 6, 35152. [Google Scholar] [CrossRef] [PubMed]
- Gouyon, P.H.; Couvet, D. A conflict between two sexes, females and hermaphrodites. In The Evolution of Sex and Its Consequences; Stearns, S.C., Ed.; Birkhauser Verlag: Basel, Switzerland, 1987; pp. 245–261. ISBN 978-3-0348-6273-8. [Google Scholar]
- Cui, X.; Wise, R.P.; Schnable, P.S. The rf2 nuclear restorer gene of male-sterile T-cytoplasm maize. Science 1996, 272, 1334–1336. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Cui, X.; Horner, H.T.; Weiner, H.; Schnable, P.S. Mitochondrial aldehyde dehydrogenase activity is required for male fertility in maize. Plant Cell 2001, 13, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Toriyama, K. Suppressed expression of RETROGRADE-REGULATED MALE STERILITY restores pollen fertility in cytoplasmic male sterile rice plants. Proc. Natl. Acad. Sci. USA 2009, 106, 9513–9518. [Google Scholar] [CrossRef] [PubMed]
- McCauley, D.E.; Olson, M.S. Do recent findings in plant mitochondrial molecular and population genetics have implications for the study of gynodioecy and cytonuclear conflict? Evolution 2008, 62, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Case, A.L.; Willis, J.H. Hybrid male sterility in Mimulus (Phrymaceae) is associated with a geographically restricted mitochondrial rearrangement. Evolution 2008, 62, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Darracq, A.; Varré, J.S.; Marechal-Drouard, L.; Courseaux, A.; Castric, V.; Saumitou-Laprade, P.; Oztas, S.; Lenoble, P.; Vacherie, B.; Barbe, V.; et al. Structural and content diversity of mitochondrial genome in beet: A comparative genomic analysis. Genome Biol. Evol. 2011, 3, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Mower, J.P.; Case, A.L.; Floro, E.R.; Willis, J.H. Evidence against equimolarity of large repeat arrangements and a predominant master circle structure of the mitochondrial genome from a monkeyflower (Mimulus guttatus) lineage with cryptic CMS. Genome Biol. Evol. 2012, 4, 670–686. [Google Scholar] [CrossRef] [PubMed]
- Štorchová, H.; Müller, K.; Lau, S.; Olson, M.S. Mosaic origin of a complex chimeric mitochondrial gene in Silene vulgaris. PLoS ONE 2012, 7, e30401. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Yang, J.; Pan, F.; Jin, B. Differential expression of microRNAs may regulate pollen development in Brassica oleracea. Genet. Mol. Res. 2015, 14, 15024–15034. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.D.; Štorchová, H. The application of RNA-seq to the comprehensive analysis of plant mitochondrial transcriptomes. Mol. Genet. Genom. 2015, 290, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Zhang, H.Y.; Zheng, Y.Z.; Ding, Y. Comparative expression profiling of miRNAs between the cytoplasmic male sterile line MeixiangA and its maintainer line MeixiangB during rice anther development. Planta 2015, 241, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xie, Y.; Xu, L.; Wang, Y.; Zhu, X.W.; Wang, R.H.; Zhang, Y.; Muleke, E.M.; Liu, L.W. Identification of microRNAs and Their Target Genes Explores miRNA-Mediated Regulatory Network of Cytoplasmic Male Sterility Occurrence during Anther Development in Radish (Raphanus sativus L.). Front. Plant Sci. 2016, 7, 1054. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.K.; Li, Y.; Zhang, C.W.; Duan, W.K.; Huang, F.Y.; Hou, X.L. Basic helix-loop-helix transcription factor BcbHLHpol functions as a positive regulator of pollen development in non-heading Chinese cabbage. Funct. Integr. Genom. 2014, 14, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Ding, X.L.; Wang, X.; He, T.T.; Zhang, H.; Yang, L.S.; Wang, T.L.; Chen, L.F.; Gai, J.Y.; Yang, S.P. Genome-wide comparative analysis of DNA methylation between soybean cytoplasmic male-sterile line NJCMS5A and its maintainer NJCMS5B. BMC Genom. 2017, 18, 596. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Achkar, N.P.; Cambiagno, D.A.; Manavella, P.A. miRNA biogenesis: A dynamic pathway. Trends Plant Sci. 2016, 21, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Castillo-Gonzalez, C.; Yu, B.; Zhang, X.R. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Voinnet, O.; Chappell, L.; Baulcombe, D. Two classes of short interfering RNA in RNA silencing. EMBO J. 2002, 21, 4671–4679. [Google Scholar] [CrossRef] [PubMed]
- Zilberman, D.; Cao, X.F.; Jacobsen, S.E. ARGONAUTE4 control of locus-specific siRNA accumulation and DNA and histone methylation. Science 2003, 299, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, K.B.; Guang, S.; Buckley, B.A.; Wong, L.; Bochner, A.F.; Kennedy, S. A pre-mRNA-associating factor links endogenous siRNAs to chromatin regulation. PLoS Genet. 2011, 7, e1002249. [Google Scholar] [CrossRef] [PubMed]
- MacLean, D.; Elina, N.; Havecker, E.R.; Heimstaedt, S.B.; Studholme, D.J.; Baulcombe, D.C. Evidence for Large Complex Networks of Plant Short Silencing RNAs. PLoS ONE 2010, 5, e9901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamthan, A.; Chaudhuri, A.; Kamthan, M.; Datta, A. Small RNAs in plants: Recent development and application for crop improvement. Front. Plant Sci. 2015, 6, 208. [Google Scholar] [CrossRef]
- Rogers, K.; Chen, X.M. Biogenesis, Turnover, and Mode of Action of Plant MicroRNAs. Plant Cell 2013, 25, 2383–2399. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; You, C.J.; Chen, X.M. The evolution of microRNAs in plants. Curr. Opin. Plant Biol. 2017, 35, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Y.; Wu, M.; Li, L.H.; Jin, C.; Zhang, Q.L.; Chen, C.B.; Song, W.Q.; Wang, C.G. Small RNA Sequencing Reveals Differential miRNA Expression in the Early Development of Broccoli (Brassica oleracea var. italica) Pollen. Front. Plant Sci. 2017, 8, 404. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Liu, X.Y.; Xu, B.C.; Zhao, N.; Yang, X.D.; Zhang, M.F. Identification of miRNAs and their targets using high-throughput sequencing and degradome analysis in cytoplasmic male-sterile and its maintainer fertile lines of Brassica juncea. BMC Genom. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.N.; Zheng, B.B.; Wang, L.; Wu, X.M.; Xu, Q.; Guo, W.W. High-throughput sequencing and degradome analysis reveal altered expression of miRNAs and their targets in a male-sterile cybrid pummelo (Citrus grandis). BMC Genom. 2016, 17, 591. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.C.; Zhang, X.H.; Yao, Q.J.; Yuan, Y.X.; Li, X.X.; We, F.; Zhao, Y.Y.; Zhang, Q.; Wang, Z.Y.; Jiang, W.S.; et al. The miRNAs and their regulatory networks responsible for pollen abortion in Ogura-CMS Chinese cabbage revealed by high-throughput sequencing of miRNAs, degradomes, and transcriptomes. Front. Plant Sci. 2015, 6, 894. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.L.; Li, J.J.; Zhang, H.; He, T.T.; Han, S.H.; Li, Y.W.; Yang, S.P.; Gai, J.Y. Identification of miRNAs and their targets by high-throughput sequencing and degradome analysis in cytoplasmic male-sterile line NJCMS1A and its maintainer NJCMS1B of soybean. BMC Genom. 2016, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhang, Z.M.; Lin, H.J.; Liu, H.J.; Chen, J.; Peng, H.; Cao, M.J.; Rong, T.Z.; Pan, G.T. Cytoplasmic male sterility-regulated novel microRNAs from maize. Funct. Integr. Genom. 2011, 11, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Han, S.H.; Ding, X.L.; He, T.T.; Dai, J.Y.; Yang, S.P.; Gai, J.Y. Comparative Transcriptome Analysis between the Cytoplasmic Male Sterile Line NJCMS1A and Its Maintainer NJCMS1B in Soybean (Glycine max (L.) Merr.). PLoS ONE 2015, 10, e0126771. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yang, S.P.; Gai, J.Y. Transcriptome comparative analysis between the cytoplasmic male sterile line and fertile line in soybean (Glycine max (L.) Merr.). Genes Genom. 2017, 39, 1117–1127. [Google Scholar] [CrossRef]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; van Aken, O.; de Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A membrane-Bound NAC transcription factor, ANAC017, mediates mitochondrial retrograde signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef] [PubMed]
- Van Aken, O.; Ford, E.; Lister, R.; Huang, S.B.; Millar, A.H. Retrograde signalling caused by heritable mitochondrial dysfunction is partially mediated by ANAC017 and improves plant performance. Plant J. 2016, 88, 542–558. [Google Scholar] [CrossRef] [PubMed]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Allen, E.; Xie, Z.X.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Peragine, A.; Park, M.Y.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes Dev. 2005, 19, 2164–2175. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.D.; Fahlgren, N.; Chapman, E.J.; Cumbie, J.S.; Sullivan, C.M.; Givan, S.A.; Kasschau, K.D.; Carrington, J.C. Genome-wide analysis of the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 pathway in Arabidopsis reveals dependency on miRNA- and tasiRNA-directed targeting. Plant Cell 2007, 19, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.; Meyers, B.C.; Liu, Z.C.; Beers, E.P.; Ye, S.Q.; Liu, Z.R. MicroRNA Superfamilies Descended from miR390 and Their Roles in Secondary Small Interfering RNA Biogenesis in Eudicots. Plant Cell 2013, 25, 1555–1572. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Lee, Y.S. Molecular functions of long noncoding transcripts in plants. J. Plant Biol. 2015, 58, 361–365. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Chua, N.H. Long noncoding RNA transcriptome of plants. Plant Biotechnol. J. 2015, 13, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Zhu, C.; Lu, G.; Guo, Y.; Zhou, Y.; Zhang, Z.; Zhao, Y.; Li, W.; Lu, Y.; Tang, W.; et al. Strand-specific RNA-seq reveals widespread occurrence of novel cis-natural antisense transcripts in rice. BMC Genom. 2012, 13, 721. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.H.; Liu, M.; Downie, B.; Liang, C.; Ji, G.; Li, Q.Q.; Hunt, A.G. Genome-wide landscape of polyadenylation in Arabidopsis provides evidence for extensive alternative polyadenylation. Proc. Natl. Acad. Sci. USA 2011, 108, 12533–12538. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Deng, W.; Fan, X.; Liu, T.T.; He, G.; Chen, R.; Terzaghi, W.; Zhu, D.; Deng, X.W. Genomic Features and Regulatory Roles of Intermediate-Sized Non-Coding RNAs in Arabidopsis. Mol. Plant 2014, 7, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 2009, 462, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.X.; Yan, B.X.; Qu, Y.Y.; Qin, F.F.; Yang, Y.T.; Hao, X.J.; Yu, J.J.; Zhao, Q.; Zhu, D.Y.; Ao, G.M. Zm401, a short-open reading-frame mRNA or noncoding RNA, is essential for tapetum and microspore development and can regulate the floret formation in maize. J. Cell Biochem. 2008, 105, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, H.J.; Fang, J.; Chu, C.C.; Wang, X.J. A long noncoding RNA involved in rice reproductive development by negatively regulating osa-miR160. Sci. Bull. 2017, 62, 470–475. [Google Scholar] [CrossRef]
- Wu, H.J.; Wang, Z.M.; Wang, M.; Wang, X.J. Widespread long noncoding RNAs as endogenous target mimics for microRNAs in plants. Plant Physiol. 2013, 161, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Nonomura, K.I. Isolation and bioinformatic analyses of small RNAs interacting with germ cell-specific Argonaute in rice. In PIWI-Interacting RNAs. Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2014; Volume 1093, pp. 1–249. [Google Scholar] [CrossRef]
- Ou, L.J.; Liu, Z.B.; Zhang, Z.Q.; Wei, G.; Zhang, Y.P.; Kang, L.Y.; Yang, B.Z.; Yang, S.; Lv, J.H.; Liu, Y.H. Noncoding and coding transcriptome analysis reveals the regulation roles of long noncoding RNAs in fruit development of hot pepper (Capsicum annuum L.). Plant Growth Regul. 2017, 83, 141–156. [Google Scholar] [CrossRef]
- Dietrich, A.; Wallet, C.; Iqbal, R.K.; Gualberto, J.M.; Lotfi, F. Organellar non-coding RNAs: Emerging regulation mechanisms. Biochimie 2015, 117, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Rurek, M. Participation of non-coding RNAs in plant organelle biogenesis. Acta Biochim. Pol. 2016, 63, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.Q.; Stone, J.D.; Štorchová, H.; Sloan, D.B. High transcript abundance, RNA editing, and small RNAs originating from intergenic regions in the massive mitochondrial genome of the angiosperm Silene noctiflora. BMC Genom. 2015, 16, 938. [Google Scholar] [CrossRef] [PubMed]
- Richly, E.; Leister, D. NUMTs in sequenced eukaryotic genomes. Mol. Biol. Evol. 2004, 21, 1081–1084. [Google Scholar] [CrossRef] [PubMed]
- Ruwe, H.; Wang, G.W.; Gusewski, S.; Schmitz-Linneweber, C. Systematic analysis of plant mitochondrial and chloroplast small RNAs suggests organelle-specific mRNA stabilization mechanisms. Nucleic Acids Res. 2016, 44, 7406–7417. [Google Scholar] [CrossRef] [PubMed]
- Holec, S.; Lange, H.; Kuhn, K.; Alioua, M.; Borner, T.; Gagliardi, D. Relaxed transcription in Arabidopsis mitochondria is counterbalanced by RNA stability control mediated by polyadenylation and polynucleotide phosphorylase. Mol. Cell. Biol. 2006, 26, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.D.; Koloušková, P.; Sloan, D.B.; Štorchová, H. Non-coding RNA may be associated with cytoplasmic male sterility in Silene vulgaris. J. Exp. Bot. 2017, 68, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
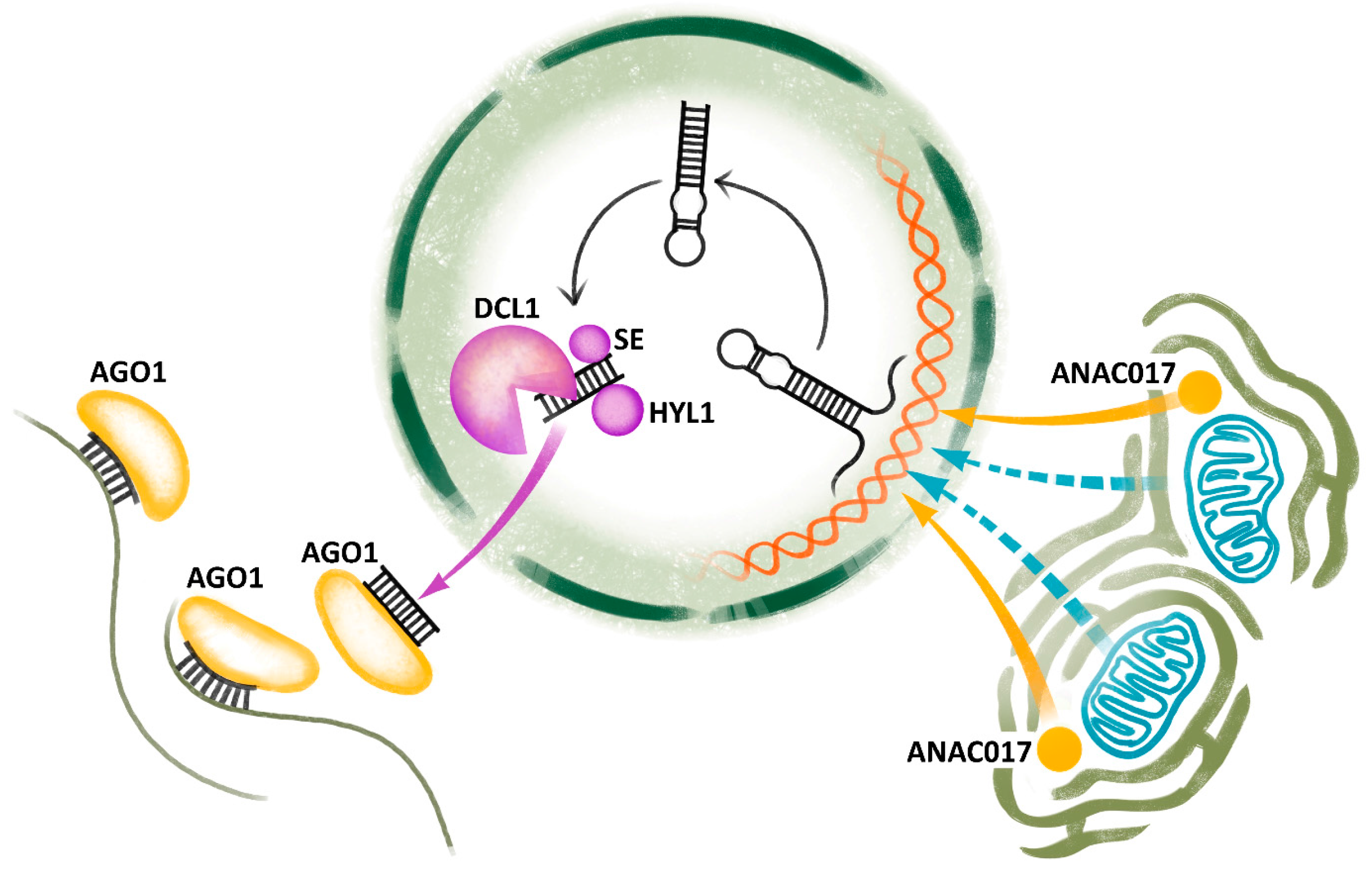
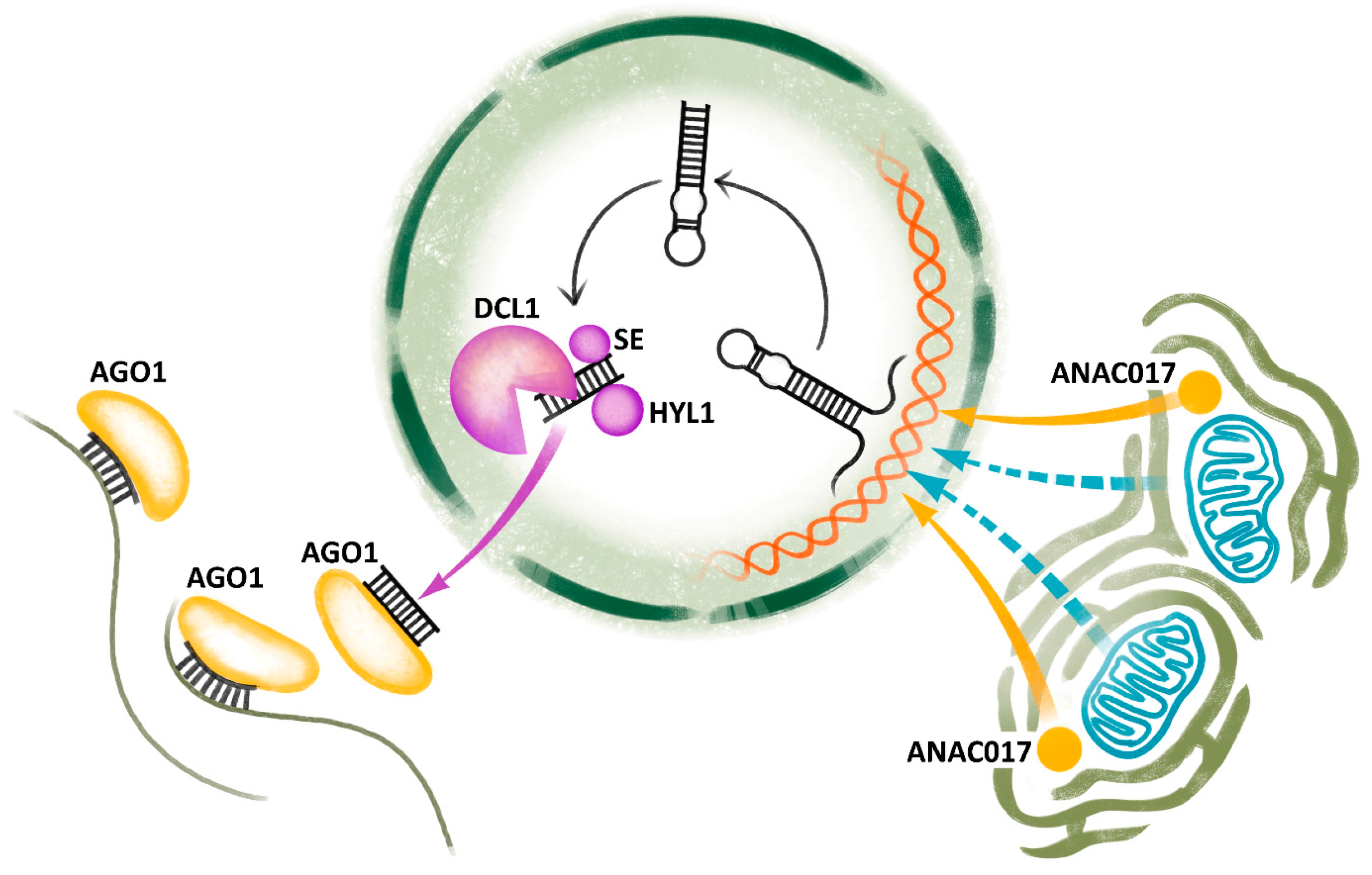
Figure 1.
Induction and biogenesis of miRNA during CMS. Mitochondrial biogenesis is altered by the action of cytoplasmic male sterility (CMS)-associated genes sending retrograde signals to the nucleus by means of the NAC transcription factor ANAC017 localized close to endoplasmatic reticulum and/or by other unknown factors. They trigger miRNA gene expression and the production of pri-miRNA, which is subsequently trimmed by the complex containing DICER LIKE1 (DCL1), HYPONASTIC LEAVES1 (HYL), SERRATE (SE) and other proteins in the nucleus. Afterwards, miRNA duplexes are transported to the cytoplasm, where they join ARGONAUTE1 (AGO1), find target mRNAs and initiate its cleavage or translation inhibition by RNA-induced silencing complex (RISC). The figure is based on well-supported model except for blue dashed arrows representing an unknown signal.
Figure 1.
Induction and biogenesis of miRNA during CMS. Mitochondrial biogenesis is altered by the action of cytoplasmic male sterility (CMS)-associated genes sending retrograde signals to the nucleus by means of the NAC transcription factor ANAC017 localized close to endoplasmatic reticulum and/or by other unknown factors. They trigger miRNA gene expression and the production of pri-miRNA, which is subsequently trimmed by the complex containing DICER LIKE1 (DCL1), HYPONASTIC LEAVES1 (HYL), SERRATE (SE) and other proteins in the nucleus. Afterwards, miRNA duplexes are transported to the cytoplasm, where they join ARGONAUTE1 (AGO1), find target mRNAs and initiate its cleavage or translation inhibition by RNA-induced silencing complex (RISC). The figure is based on well-supported model except for blue dashed arrows representing an unknown signal.

{kind=link}
Table 1.
The examples of miRNA and their putative target genes differentially expressed between the CMS lines and their maintainers in various crops.
Table 1.
The examples of miRNA and their putative target genes differentially expressed between the CMS lines and their maintainers in various crops.
| miRNA | Putative Target Genes | Target Gene Functions | References |
|---|---|---|---|
| Maize CMS C48-2 | |||
| Zma-miR397c | Laccase | Oxidation of phenolic substrates | |
| Zma-miR601 | Flavin-containing monooxygenase (FMO) | Auxin biosynthesis | [61] |
| Enoyl-CoA hydratase | Catabolism of fatty acids | ||
| Zma-miR604 | Monosaccharide transport protein 2 (STP2) | Uptake of glucose from callose degradation | |
| Brassica juncea hybrid | |||
| miR156a | SPL transcription factors | Floral transition, tapetum development | [57] |
| miR167a | Auxin response factor (ARF6/ARF8) | Anther dehiscence | |
| miR319a | TCP transcription factors | Floral induction | |
| miR395a | ATP sulphurylase (APS) | Sulphur metabolism | |
| Rice MeixiangA | |||
| osa-miR528-3p | F-box containing protein | Proteolytic turnover through proteasome | [42] |
| osa-miR1432-5p | Metal cation transporter | Cation homeostasis | |
| osa-miR2118c | NBS-LRR | Disease-resistance related proteins | |
| Brassica oleracea Bo01-12A | |||
| bol-miR157a | SPL transcription factors | Floral transition, tapetum development | [40] |
| bol-miR171a | SCARECROW-like (SCL) transcription factor | GA mediated action | |
| bol-miR172 | APETALA2 (AP2) transcription factor | Floral transition | |
| bol-miR824 | MADS-box transcription factor-like | Plant development | |
| Brassica rapa CMS-Ogura | |||
| bra-miR157a | SPL transcription factors | Floral transition, tapetum development | [59] |
| bra-miR158-3p | PPR-RFL | RNA metabolism in organelles | |
| bra-miR159a | MYB81 transcription factor | Flowering | |
| bra-miR164a | CUP SHAPED COTYLEDON 1 | Meristem development | |
| bra-miR172a | APETALA2 (AP2) transcription factor | Floral transition | |
| bra-miR5712 | VACUOLAR ATP SYNTHASE SUBUNIT A | Male gametophyte development | |
| bra-miR5716 | Zinc finger transcription factor | Drought stress response | |
| bra-miR6030 | CC-NBS-LRR | Disease-resistance related proteins | |
| Glycine max NJCMS1A | |||
| gma-miR166a-3p | HD-ZIPIII transcription factor | Vascular nad cell wall development | [60] |
| gma-miR169b | Nuclear factor Y (NF-YA) transcription factor | Flowering | |
| gma-miR171a | SCARECROW-like (SCL) transcription factor | GA mediated action | |
| gma-miR394b-5p | F-box protein | Proteolytic turnover through proteasome | |
| gma-miR395c | Sulphate transporter 2.1-like | Sulphur metabolism | |
| gma-miR396k-5p | bHLH79 transcription factor | Floral development | |
| gma-miR397a | Laccase | Oxidation of phenolic substrates | |
| gma-miR408c-3p | Plastocyanin-like | Copper metabolism | |
| Raphanus sativus CMS-WA | |||
| miR-158b-3p | PPR-RFL | RNA metabolism in organelles | [43] |
| miR161 | Mechanosensitive channel of small conductance-like 10 (MSL10) | Mechanosensitive ion channel, cell death induction | |
| miR395a | putative F-box/kelch-repeat (KFB) | Proteolytic turnover through proteasome | |
| Pummelo cybrid line | |||
| cga-miR156a.1 | SPL transcription factors | Floral transition, tapetum development | [58] |
| cga-miR399a.1 | UBC (ubiquitin-conjugating E2 enzyme) | Phosphate (Pi) homeostasis | |
| cga-miR827 | Basic leucine zipper (bZIP) | Pollen and flower development | |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Štorchová, H. The Role of Non-Coding RNAs in Cytoplasmic Male Sterility in Flowering Plants. Int. J. Mol. Sci. 2017, 18, 2429. https://doi.org/10.3390/ijms18112429
AMA Style
Štorchová H. The Role of Non-Coding RNAs in Cytoplasmic Male Sterility in Flowering Plants. International Journal of Molecular Sciences. 2017; 18(11):2429. https://doi.org/10.3390/ijms18112429
Chicago/Turabian StyleŠtorchová, Helena. 2017. "The Role of Non-Coding RNAs in Cytoplasmic Male Sterility in Flowering Plants" International Journal of Molecular Sciences 18, no. 11: 2429. https://doi.org/10.3390/ijms18112429
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.