1. Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterized by systemic inflammation, presence of autoantibodies, and targeted synovitis, affecting approximately 0.5–1% of population [
1]. Articular manifestation of inflammatory arthritis is the hallmark of RA and a major determinant of the disease activity [
2]. Numerous cell types are involved in the pathophysiology of RA, including immune cells like T cell, B cells and macrophages, synoviocytes, and chondrocytes [
3,
4]. Synoviocytes and chondrocytes are cell types within the joint dominantly affected by RA. Activated synovial fibroblasts within the inflamed synovium have altered morphology and behavior, and attach directly to cartilage and release matrix degradation enzymes, leading to the destruction of cartilage tissue [
1,
5,
6]. They also interact indirectly with adjacent macrophages through the release of receptor activator of nuclear factor κB ligand (RANKL) and mediate the differentiation of macrophage precursors into osteoclasts [
6,
7].
Bone erosion is a characteristic feature of affected joints in RA, known to be triggered mainly by synovitis, producing pro-inflammatory cytokines and RANKL [
8]. Within the inflamed joint structure, the destructive process by the pannus, a structure formed by proliferative synovium containing infiltrates of immune cells, proliferative vessels, and increased osteoclasts, leads to bone erosion particularly at the synovium–bone interface where the pannus invades [
9]. Through the direct contact of the invading pannus to the bone and increased angiogenesis within the pannus structure, the capacity of the synovial fibroblasts to damage structures within the joint has been widely proposed [
8,
9,
10,
11]. Together with synovial fibroblasts and immune cells, the critical role of osteoclasts in the disrupted bone homeostasis under pathological condition such as RA is being studied. The imbalance of bone homeostasis in RA leads to peri-articular bone loss, which usually precedes bone erosion and further progression of joint destruction [
12].
While osteoclasts are the major cells responsible for bone loss and bone erosion in RA, the impaired differentiation and function of osteoblasts have been proposed in recent studies, and inflammatory tissue in RA may impair osteoblast activity, which respond differently from osteoblasts of osteoarthritic condition [
12,
13,
14,
15]. The precursors of immune cells are formed and maintained in the bone marrow, where osteoclasts and osteoblasts reside and act to maintain a balanced bone remodeling; therefore, research on the interplay between the immune system and the skeletal system has emerged to gain more knowledge on the novel field termed osteoimmunology [
8,
16,
17].
MicroRNAs (miRNAs) are non-coding single strand RNAs consisting of 20–22 nucleotides, acting primarily on the 3’UTR of mRNAs to regulate gene expressions through a post-transcriptional manner [
18]. These small non-coding RNAs participate in the regulation of numerous cellular processes and dysregulation of miRNAs are associated with various diseases [
19]. The role of miRNA regulation in bone diseases has been reported, including osteoporosis and arthritis that are associated with altered bone homeostasis [
20,
21]. The study of gene associations using the next generation sequencing (NGS) technique and bioinformatics approaches has evolved, providing high-throughput genomic profiling and further understanding and analysis of functional annotations of identified genes and/or miRNAs [
22,
23]. In this study, we used various bioinformatics tools and databases to assist in identifying potential miRNA–mRNA interactions in osteoblasts of RA population, including miRmap [
24], Gene Expression Omnibus (GEO) [
25], Ingenuity
® Pathway Analysis (IPA) [
26], and Database for Annotation, Visualization and Integrated Discovery (DAVID) [
27].
The role of osteoblasts in the development of peri-articular bone loss and limited repair of bone erosion in RA has received much attention, and miRNAs are hypothesized to play critical roles in the regulation of osteoblast function in RA. The aim of our current study is to explore novel miRNAs differentially expressed in osteoblasts of RA bone and to identify genes potentially involved in the dysregulated bone homeostasis in RA. Using the NGS for genomic profiling and various bioinformatics approaches, we expect the findings will provide novel insights into potential therapeutic targets that contribute to better control of RA disease activity.
3. Discussion
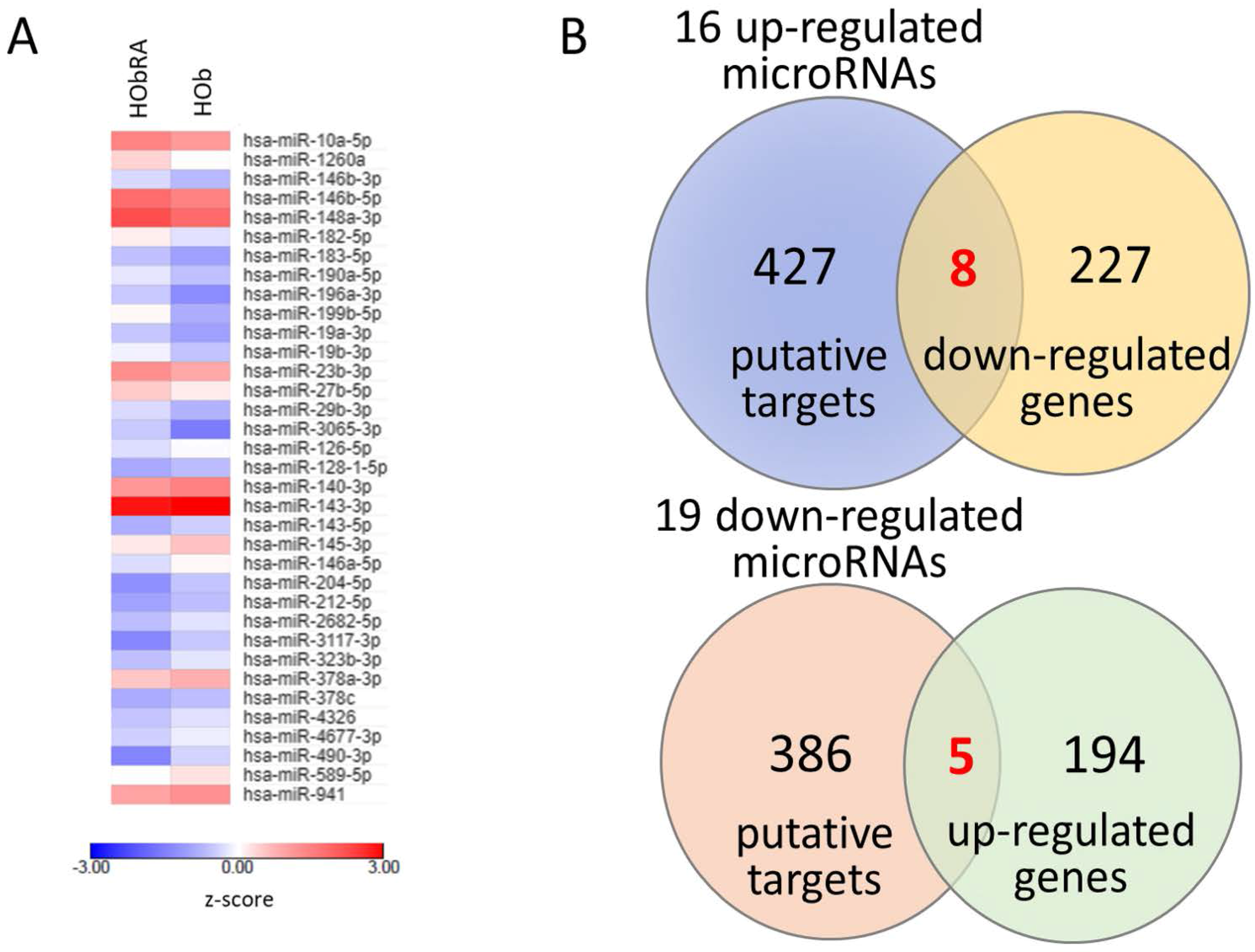
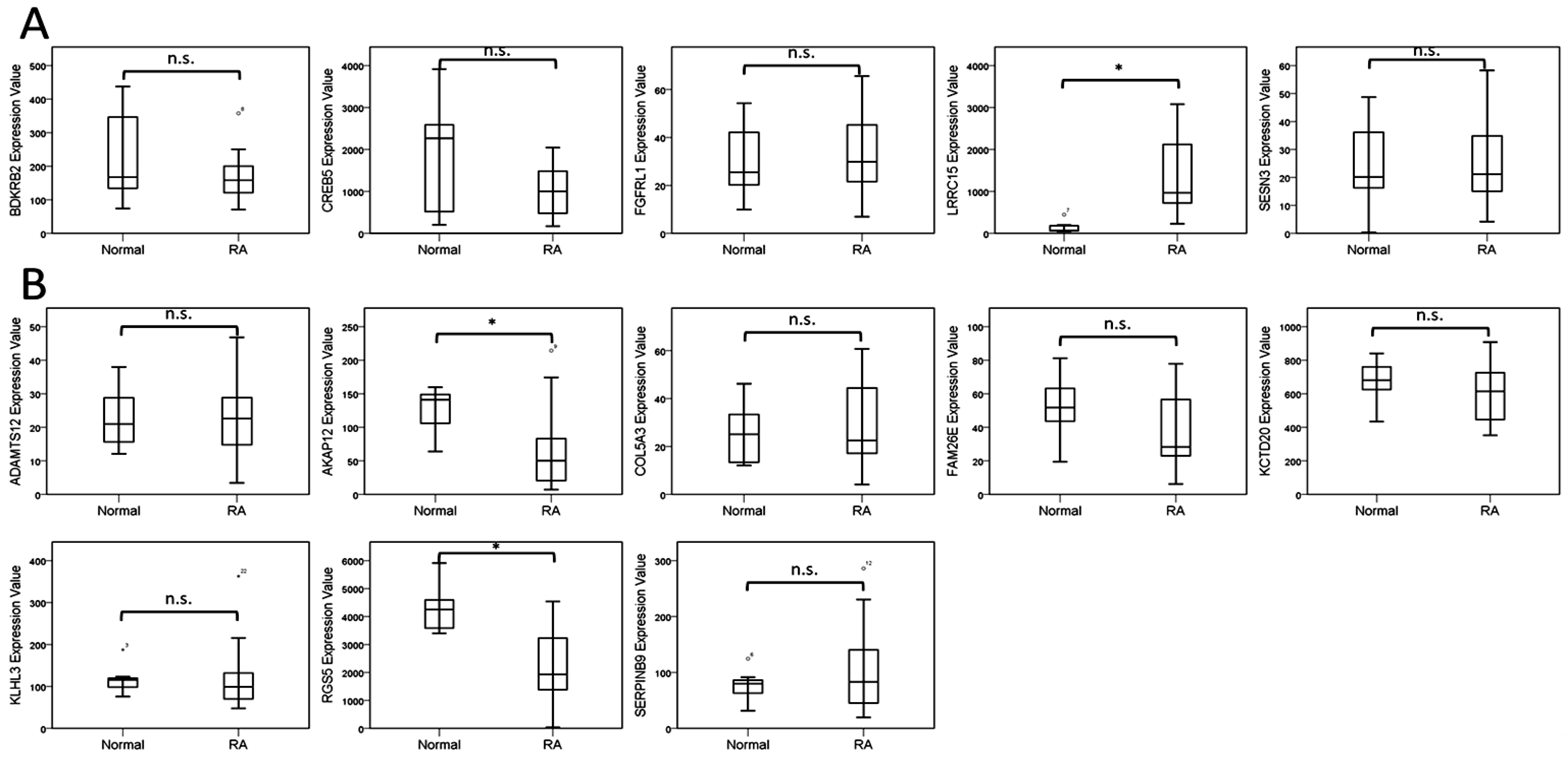
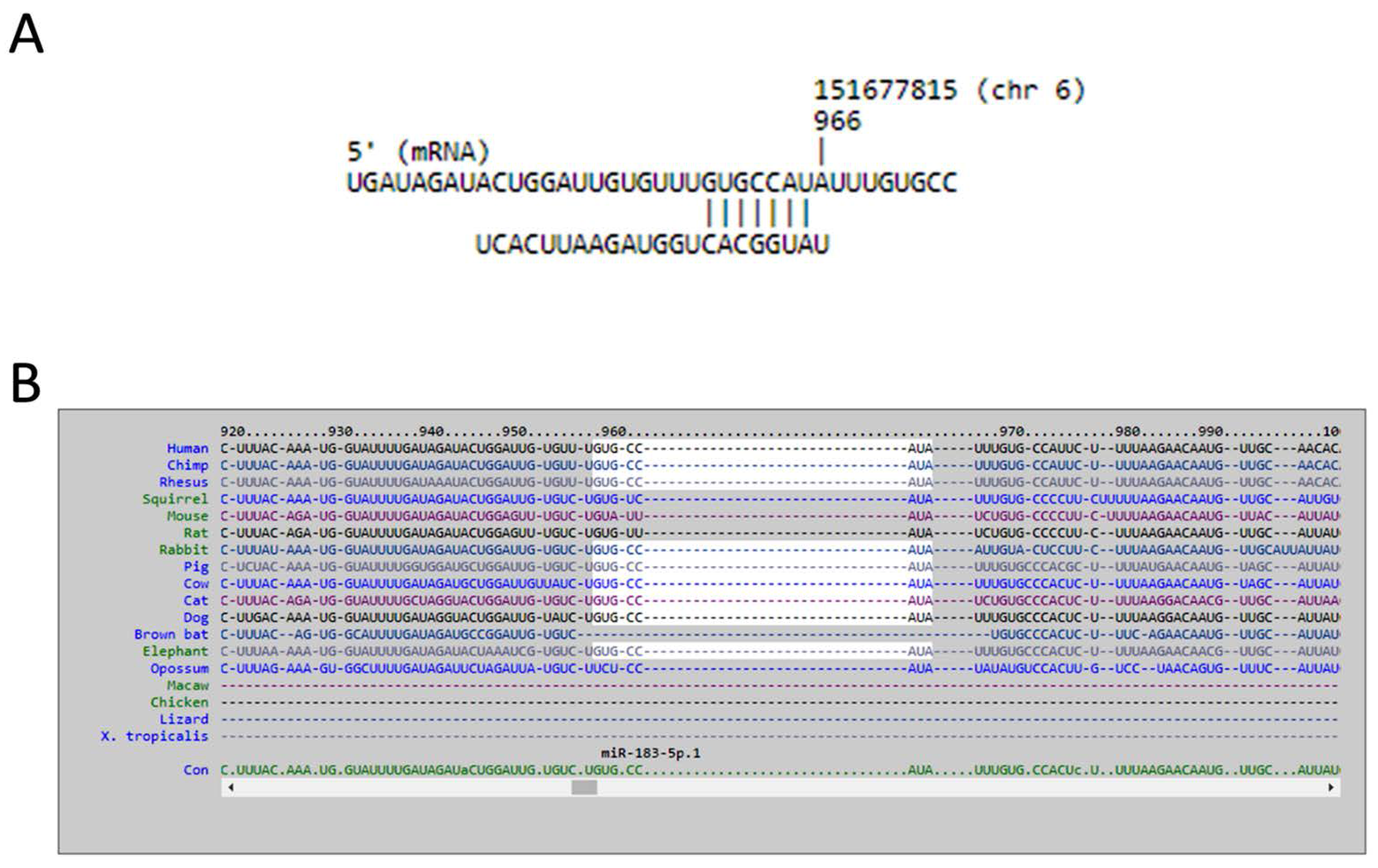
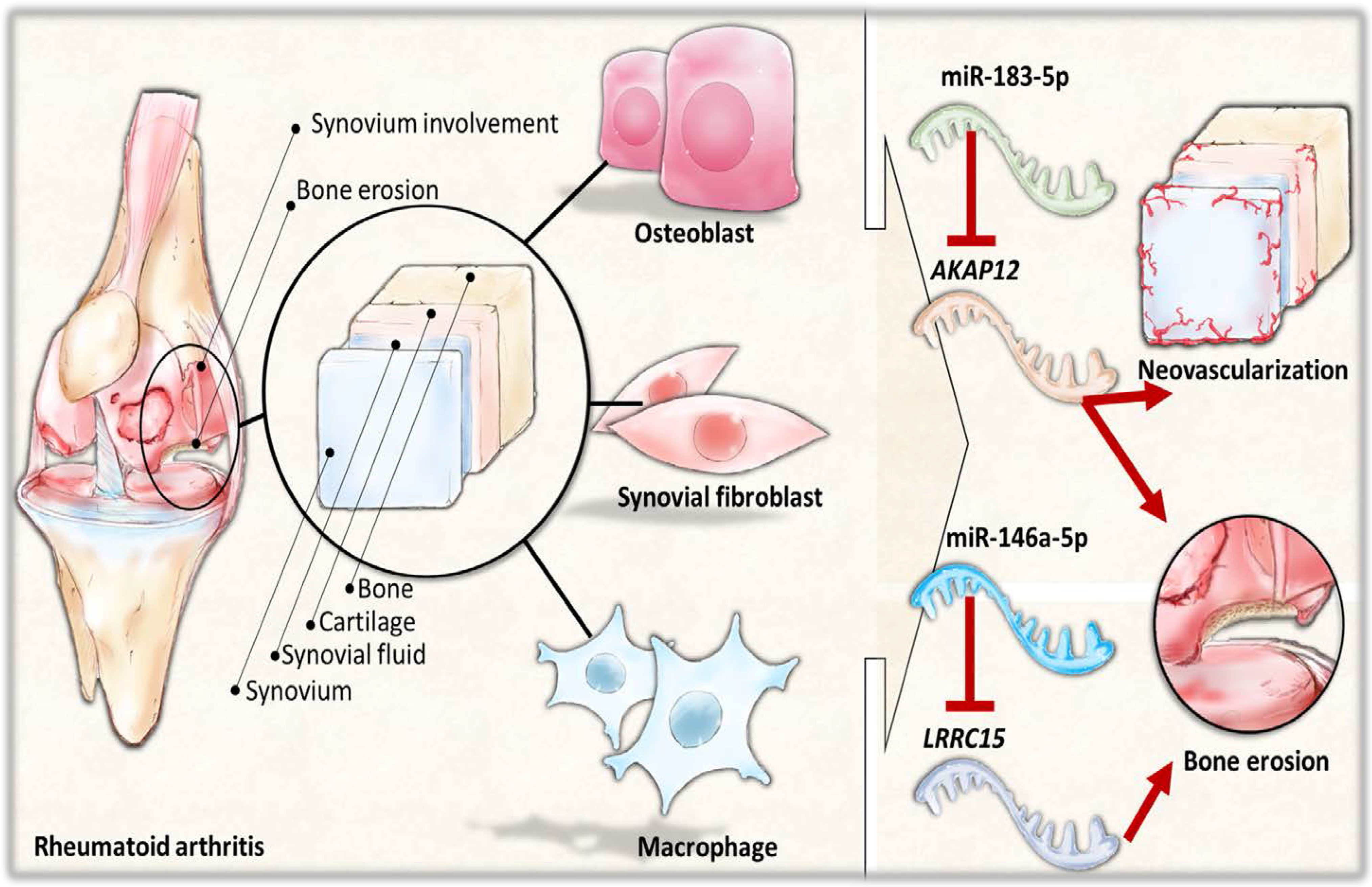
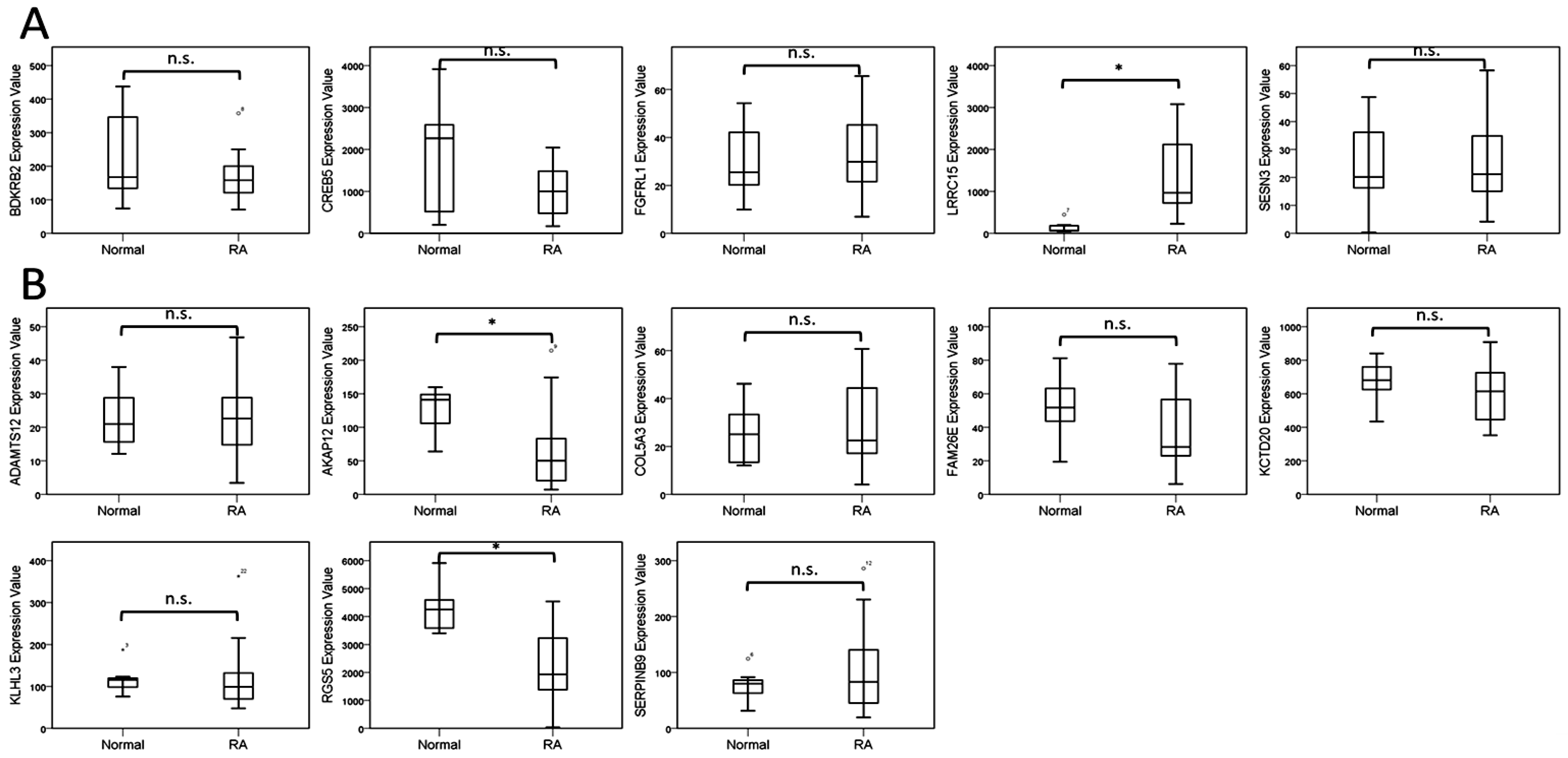
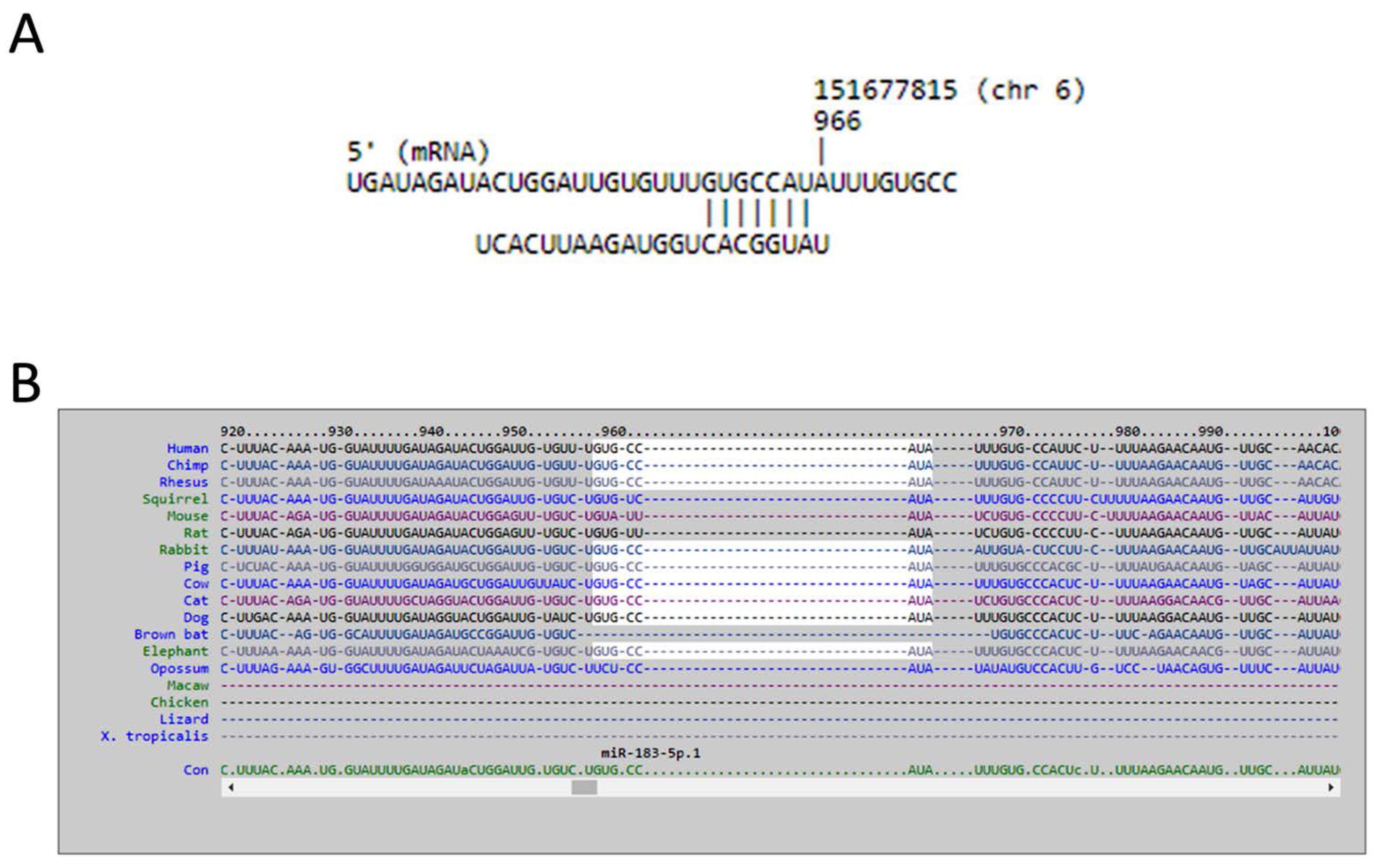
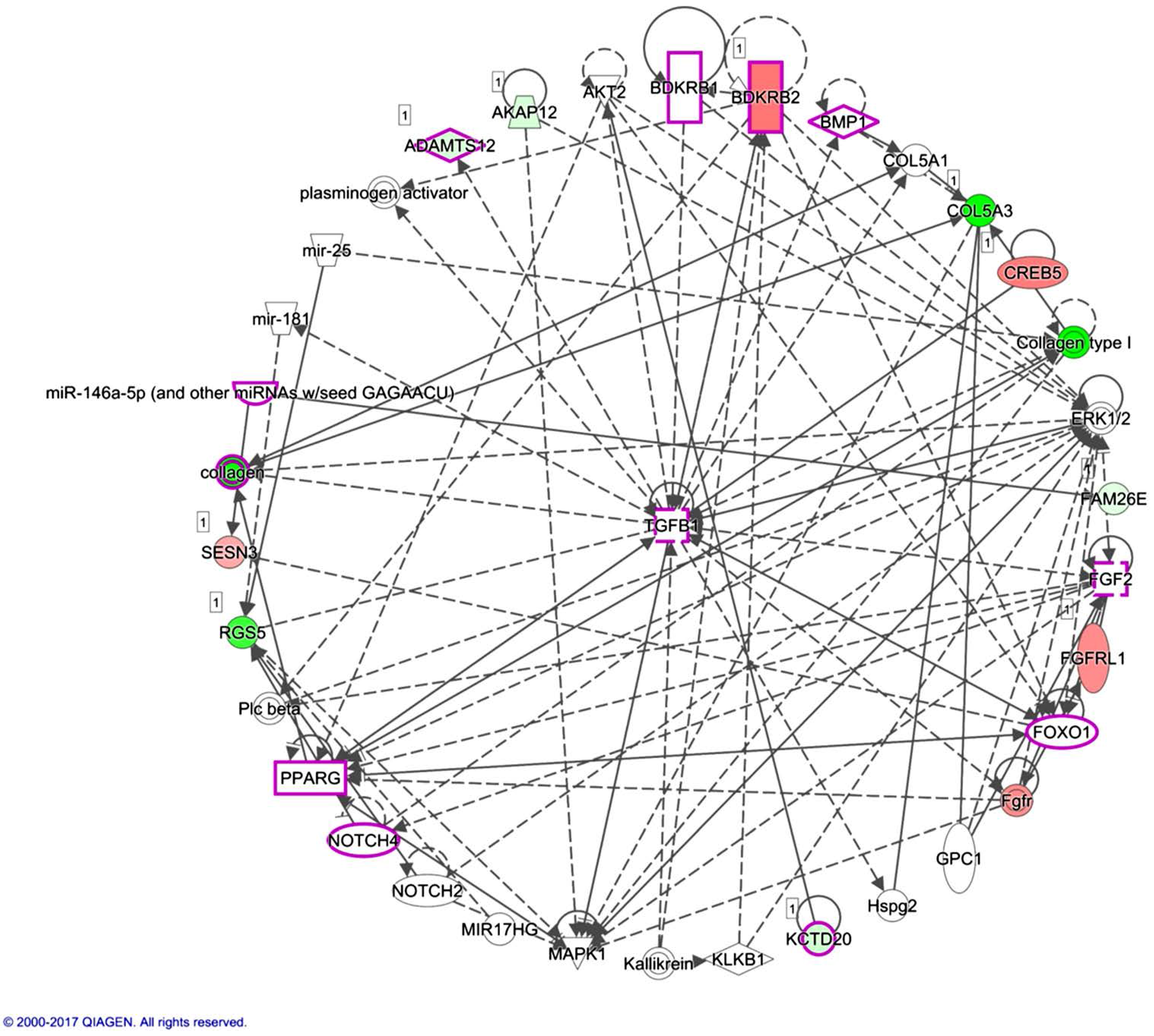
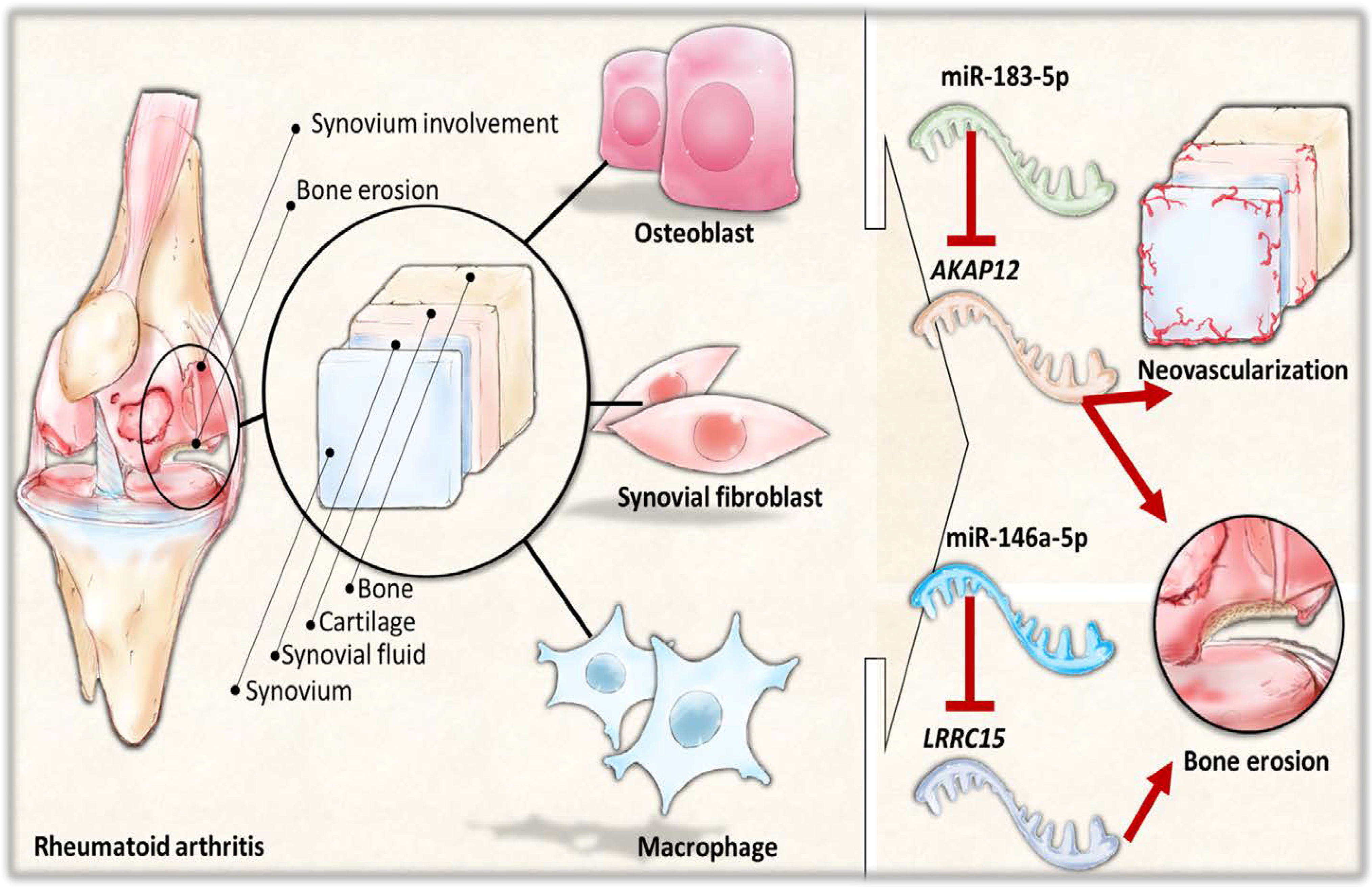
The current study identified 35 differentially expressed miRNAs and 13 candidate genes potentially involved in osteoblasts of RA, using NGS and bioinformatics analysis. Two of the 13 candidate genes differentially expressed in osteoblasts of RA, LRRC15 and AKAP12, were found to have consistent direction of expression in the synovial tissue of RA patients identified in four of the five RA array datasets (GSE7307, GSE55475, GSE77298, GSE55235, and GSE1919). The potential miRNA regulation on LRRC15 was miR-146a-5p, whereas the potential miRNA regulation on AKAP12 was miR-183-5p, predicted by miRmap, TargetScan and miRDB database. We proposed the novel findings of miR-146a-5p–LRRC15 and miR-183-5p–AKAP12 regulations in the altered function of osteoblasts in RA microenvironment.
The role of osteoblasts in the development of bone loss and limited capacity of repair of bone erosion in RA has received more attention recently, and miRNAs are hypothesized to play critical roles in the regulation of the function of osteoblasts in RA [
13,
15,
28]. The results of IPA analysis identified miR-29b-3p to be a potential upstream regulator of the differentially expressed genes in RA. The regulation of the miR-29 family is proposed to participate in osteoarthritis and cartilage homeostasis [
29]. In addition, the miR-29 family has been shown to promote osteoblast differentiation by targeting inhibitors of the Wnt signaling pathway, and to possess numerous distinct activities at different stages of osteoblast differentiation. In the mature osteoblasts, miR-29 targets collagen type I, reducing the rate of collagen synthesis and facilitating structural stability of the bone [
30]. In a review article by Miao et al., the Wnt signaling pathway participates in the pathogenesis of RA and bone remodeling. They suggested that inhibition of the Wnt signaling pathway may contribute to impaired osteoblast function in RA, and increased expression of several inhibitors of the Wnt signaling pathway may contribute to bone resorption in RA [
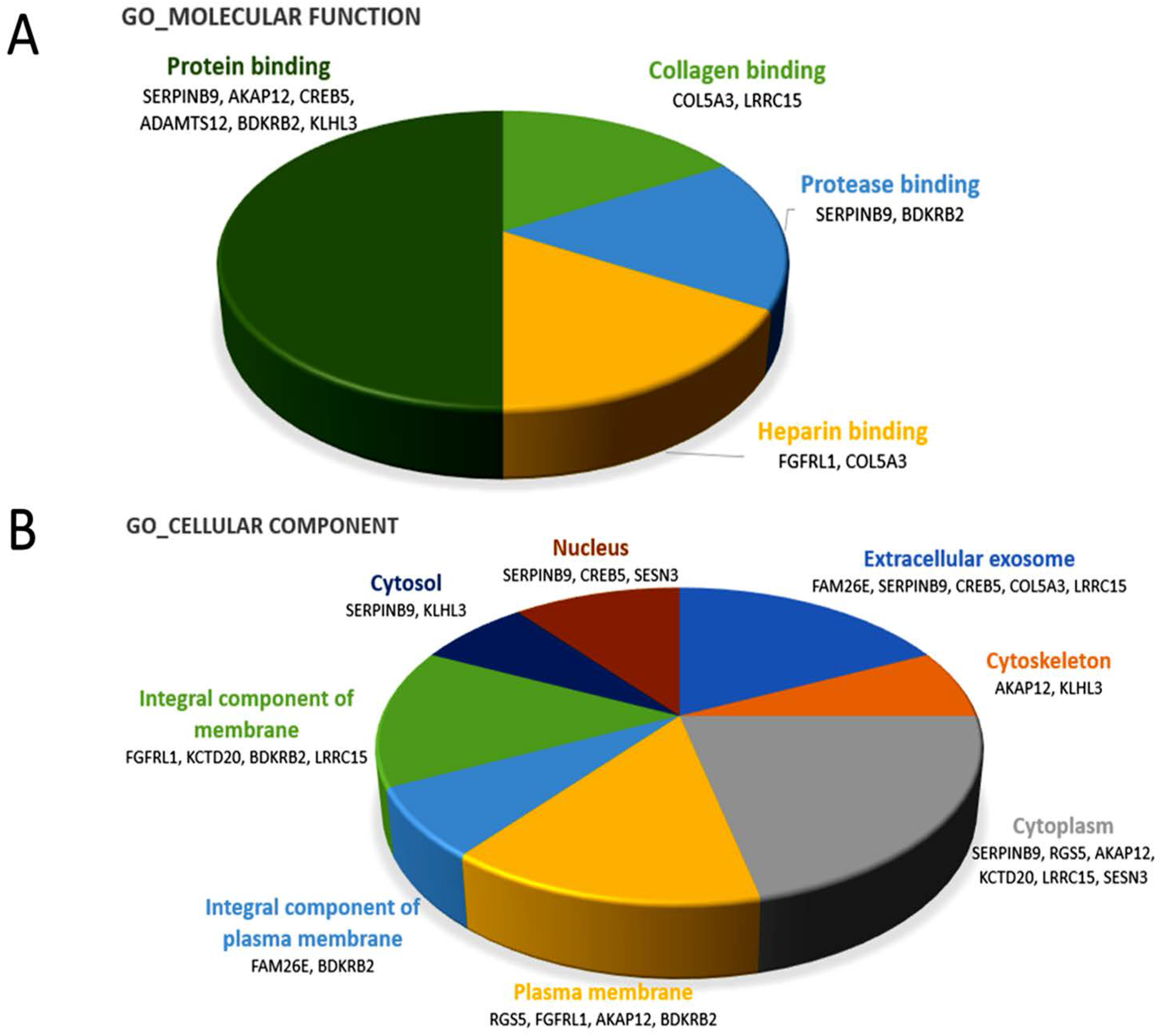
31]. Two of the effectors of miR-29b-3p in our candidate genes,
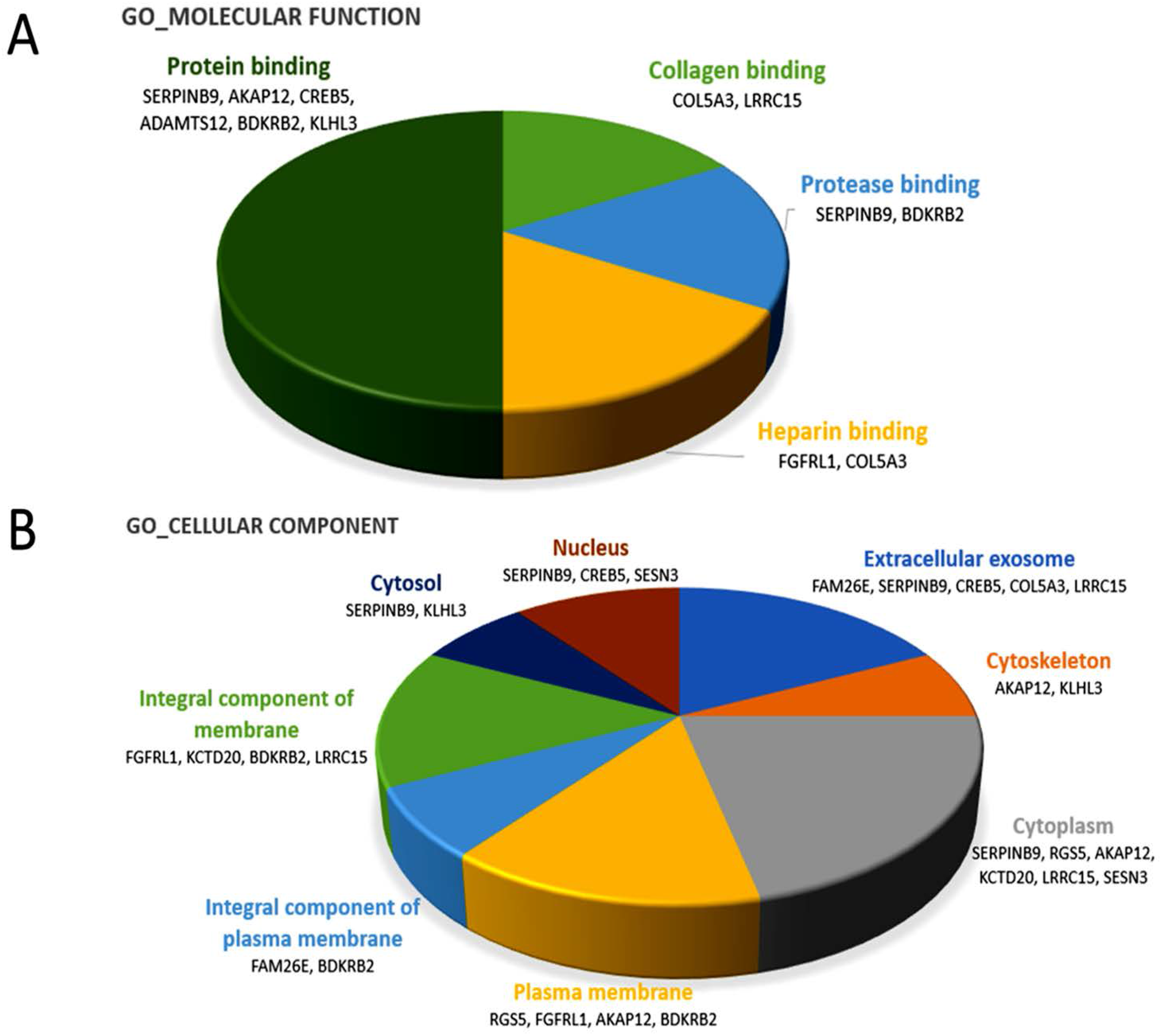
BDKRB2 and
SERPINB9, were categorized into the molecular function of protease binding by gene ontology, as shown in
Figure 2A and
Table 6, which may support the involvement of miR-29b-3p in the regulation of bone matrix in RA.
A-kinase anchoring protein 12 (AKAP12), one of the A-kinase anchoring proteins, is a scaffold protein for protein kinase A (PKA) and protein kinase C (PKC) that control cytoskeleton dynamics, cell migration, and cell adhesion [
32,
33]. Studies suggested that the down-regulation of AKAP12 induces the formation of stress fibers and proliferation of adhesion complexes; increased cellular senescence was also observed in AKAP12-null mice [
34]. The findings potentially link
AKAP12 to the disrupted bone homeostasis in RA, where the study by Yudoh and colleagues revealed the higher rate of cellular senescence and greater decline in the replicative capacity of peri-articular osteoblasts in RA patients, compared to osteoarthritic patients [
15]. The expressions of AKAP12 in inflammatory responses were studied. The increased protein expression of AKAP12 in the fibrotic scar may restrict excessive inflammation during central nervous system repair [
35], and decreased protein levels of AKAP12 were observed in the lung tissue of patients with chronic obstructive pulmonary disease [
36], suggesting the participation of AKAP12 in the regulation of inflammatory response. There is not much literature discussing the role of AKAP12 in the bone homeostasis or the inflamed joint microenvironment. One study identified
Akap12 to be one of the genes possibly involved in the alternative splicing in bone following mechanical loading in rat model [
37]. The altered joint structures along with inflamed peri-articular soft tissue in RA predispose the affected joint to increased mechanical loading, which may disrupt the PKA-mediated mechanotransduction.
Scaffold proteins serve as connecting hubs that modulate both upstream signaling molecules and the downstream effectors within cells. The expression and activity of AKAP12 is proposed to be affected by the hypoxic tumor microenvironment [
38]. Studies also suggested
AKAP12 to be a tumor suppressor and angiogenesis suppressor gene that down-regulates vascular endothelial growth factor, potentially through epigenetic regulation [
39]. The role of miRNA regulation in altered bone homeostasis has been reported [
20,
21]. Few studies also reported that miR-183 increased osteoclastogenesis through the binding on the 3′UTR of heme oxygenase-1 [
40], and oxidative stress within the bone marrow microenvironment may alter miRNA cargo of extracellular vesicles, expressing high abundance of miR-183 cluster and miR-183-5p transfection inhibits the osteogenic differentiation of bone marrow stromal cells [
41]. Altogether, with these literature reviews, the up-regulated miR-183-5p with its putative target of down-regulated
AKAP12 in our NGS result suggests the novel finding of potential miR-183-5p–
AKAP12 regulation in the changed bone homeostasis in RA joint microenvironment.
Leucine rich repeat containing 15 (
LRRC15), also named
LIB, is a gene encoding leucine-rich transmembrane protein that participates in the cell–matrix adhesion and cell migration, and is induced and highly expressed in various cancer types [
42,
43,
44]. The role of
LRRC15 in inflammation is also proposed, as it is induced by beta-amyloid and pro-inflammatory cytokines in astrocytes of Alzheimer's disease brain [
45,
46] and up-regulated by pro-inflammatory stimuli during the process of dental caries [
47]. There is still lack of related literature on the role of
LRRC15 in the arthritic joint microenvironment or other bone diseases.
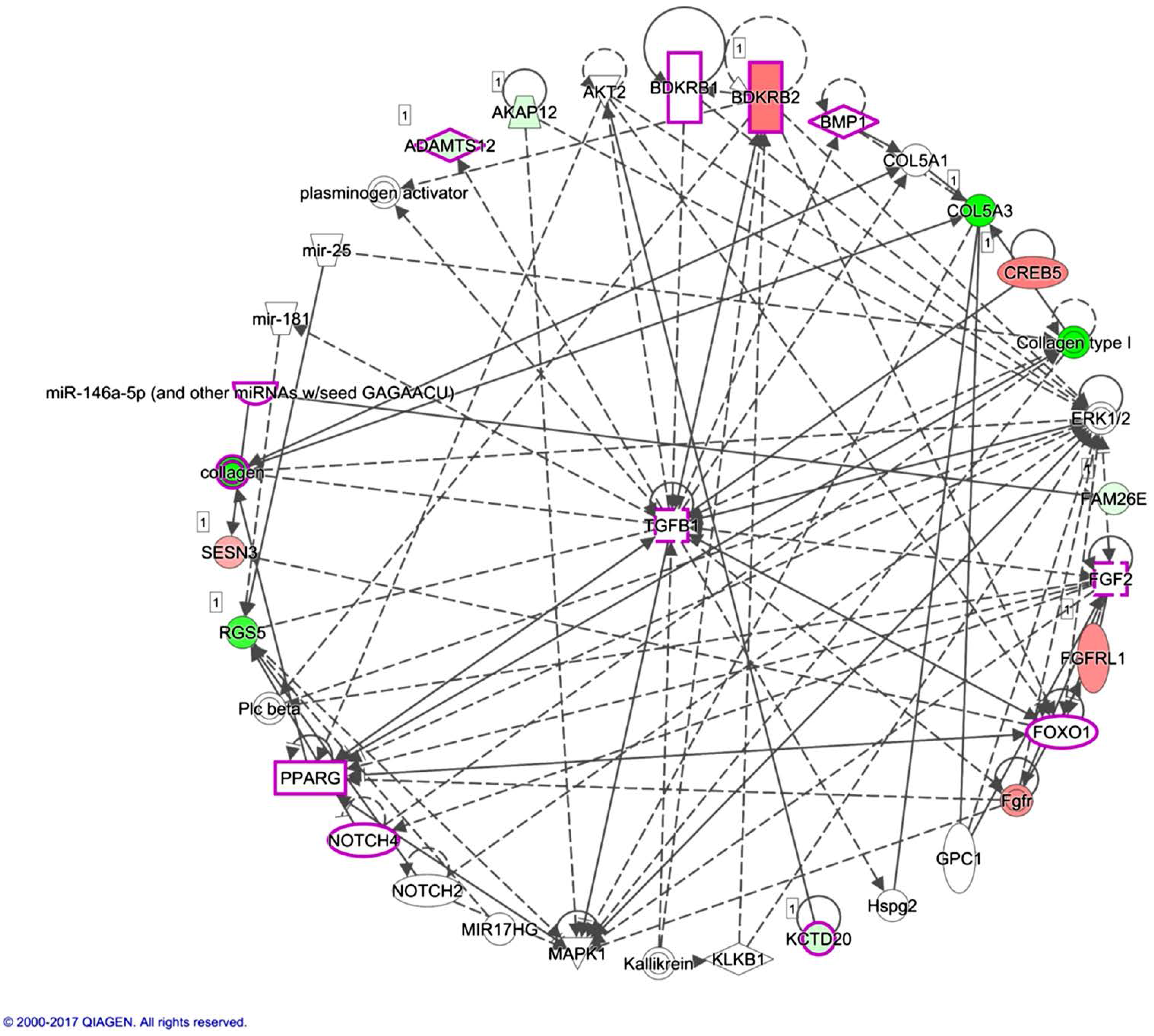
miR-146a-5p was identified to be one of the molecules associated with the inflammation of joint in the IPA analysis, as indicated in
Figure 6. miR-146 is one of the miRNAs strongly implicated in RA, regulating a group of target genes related to inflammation. The level of miR-146a is increased in synovial tissue, synovial fluid, whole blood and many other cells types such as T cells, B cells and macrophages. However, the association between the expression level of miR-146a and disease activity is still inconclusive [
48]. miR-146a is also proposed to prevent joint destruction in arthritic mice by inhibiting osteoclastogenesis [
49]. Our results indicated the potential regulation of miR-146a-5p on
LRRC15. Whether the regulation of miR-146a-5p in bone is mediated by inflammatory joint microenvironment or other stimuli merits further clarification.
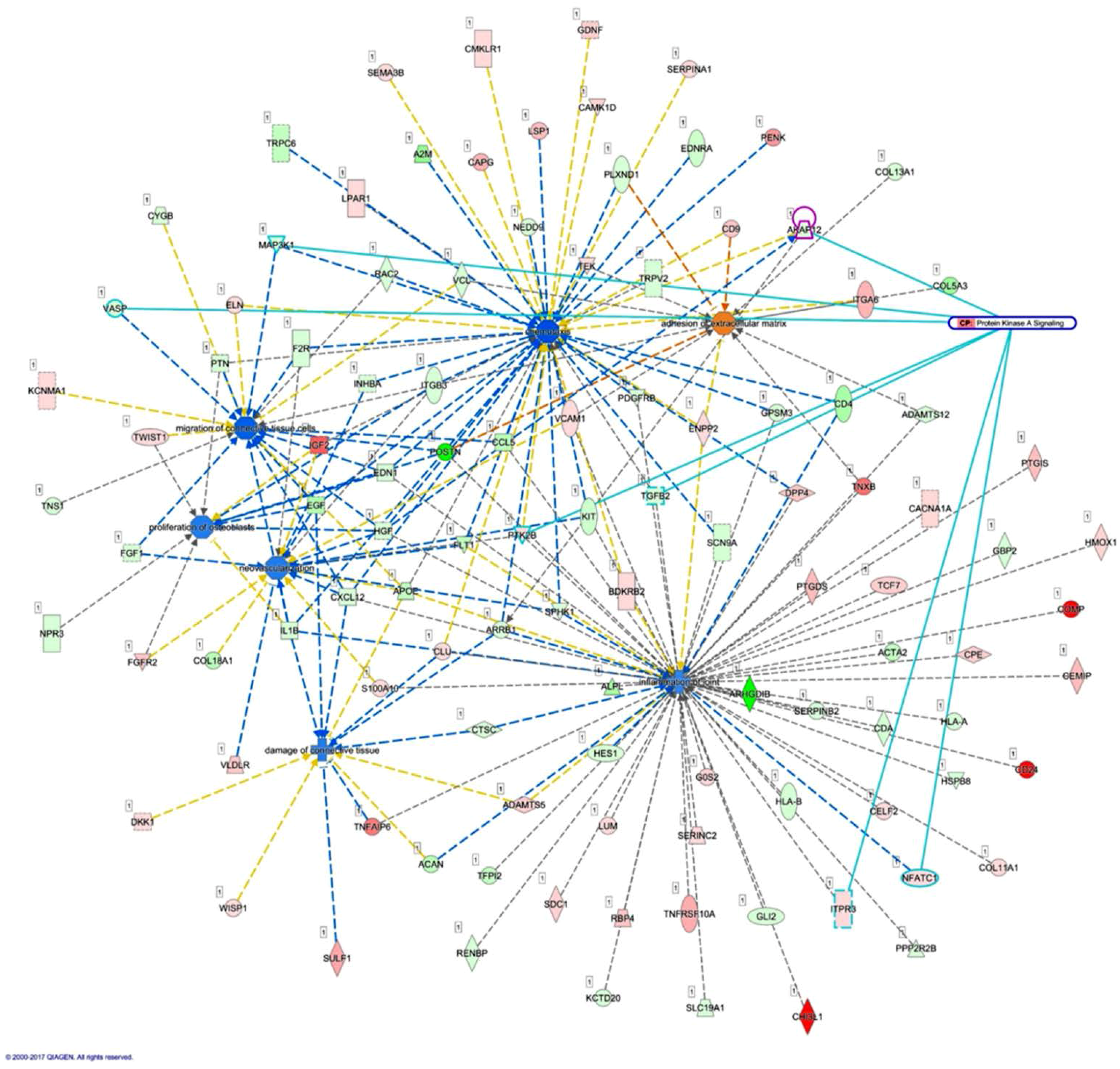
The interconnection between different cell types and the arthritic microenvironment is complex yet important in the understanding of the RA disease entity. The area of pannus formation is infiltrated by synoviocytes, immune cells, osteoclasts and proliferative vessels, and comes into direct contact with adjacent bone surface, where osteoblasts reside [
9]. Along with immune cells and synovial fibroblasts, the role of osteoblasts in the pathogenesis of articular destruction in RA have gained much attention, with the function of osteoblasts being compromised at sites of focal erosion and reduced mineralization of the newly formed bone in the arthritic joints [
50]. The activity and function of osteoblasts is also inhibited by the hypoxia within the arthritic joint microenvironment [
51]. In the current study, we explored novel miRNAs and the putative targets expressed in the osteoblasts of RA origin. Several of the target genes identified in our study were also found to have consistent directions of expression in the synovial tissues of RA patients from related GEO array datasets. The schematic potential molecular mechanisms are summarized in
Figure 9. These identified miRNA–mRNA regulations may provide novel perspectives into deeper understanding of the cell–cell communication between different cell types within the joint structure and the role of arthritic joint microenvironment in the altered bone homeostasis.
4. Materials and Methods
4.1. Primary Cell Culture
Osteoblasts isolated from normal human bones (HOb, Catalog No. 406-05a, Lot No. 3145) and bones of patients with RA (HObRA, Catalog No. 406RA-05a, Lot No. 1796) were purchased from Cell Applications, Inc. (San Diego, CA, USA). In detail, osteoblasts of normal bone were obtained from a 66-year-old female, and osteoblasts of RA bone were obtained from a 72-year-old female with RA. Osteoblasts were grown in human osteoblast growth medium (Cell Applications, Inc.) and maintained in 37 °C incubator containing 5% CO2 until confluence. The HOb and HObRA cells were then harvested for total RNA extraction and further mRNA and small RNA profiling.
4.2. RNA Sequencing
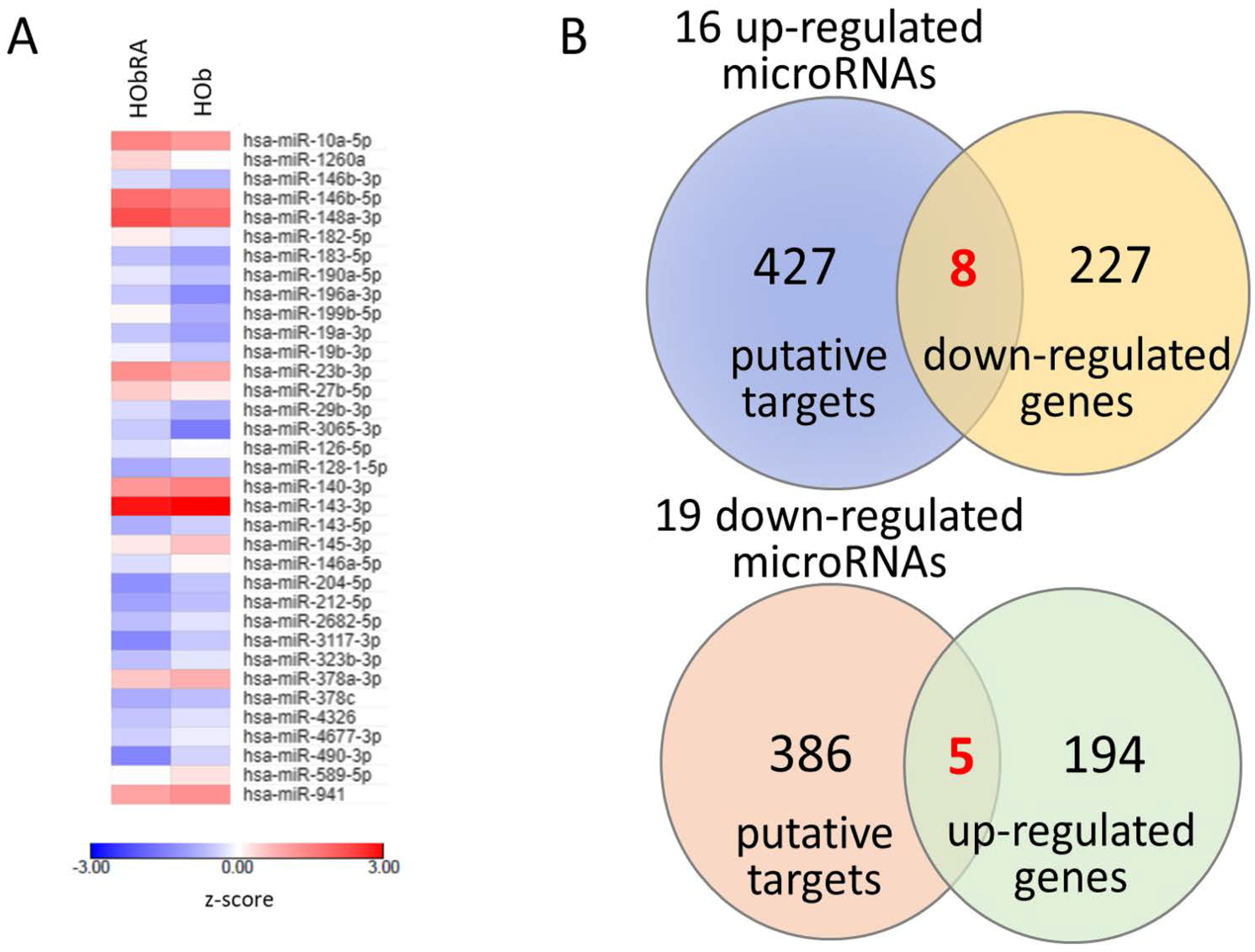
To prepare samples for mRNA and small RNA profiling, total RNAs from HOb and HObRA cells were extracted by Trizol® Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Before further sequencing, the quality of extracted RNAs were analyzed by OD260 detection using ND-1000 spectrophotometer (Nanodrop Technology, Wilmington, DE, USA) and validated by RNA integrity number (RIN) with Agilent Bioanalyzer (Agilent Technology, Santa Clara, CA, USA), where RINs for HOb and HObRA were 9.9 and 10, respectively. Total RNA sequencing analysis for RNA-seq and small RNA-seq were performed by Welgene Biotechnology Company (Welgene, Taipei, Taiwan). We set the criteria for differentially expressed mRNAs and miRNAs at fold change >2.0, FPKM >0.3 for mRNA and RPM >10 for miRNA.
4.3. miRmap Database
miRmap is an open-source database that provides miRNA target prediction using a comprehensive approach with various computational tools, including thermodynamic, evolutionary, probabilistic and sequence-based approaches. The repression strength of a miRNA–mRNA interaction of interest is indicated by the miRmap score. The higher miRmap score indicates higher repression strength. The 35 differentially expressed miRNAs were consecutively inputted to obtain putative target genes, and those with miRmap scores higher than 99.0 were selected for further analysis [
24].
4.4. Gene Expression Omnibus (GEO)
The GEO database provides public access to high-throughput array- and sequence-based data. The dataset of interest also provides link to web-based tool such as GEO2R and users can look for candidate genes and perform further analysis by obtaining raw data with expression values of specific genes in the array [
25]. The arrays related to joint tissues of RA patients (GSE1919, GSE55235, GSE55475, GSE7307, GSE77298, GSE29746, GSE10500 and GSE7779) were used in this study to identify genes that expressed in consistent directions with our NGS results.
4.5. Ingenuity® Pathway Analysis (IPA)
The Ingenuity
® Pathway Analysis (IPA) software (Ingenuity systems, Redwood City, CA, USA) contains large database with detailed and structured findings reviewed by experts which was derived from thousands of biological, chemical and medical researches, and provide researchers with quick searching. The IPA also enables analysis, integration, and recognition of data from gene and SNP arrays, RNA and small RNA sequencing, proteomics and many other biological experiments; in addition, deeper understanding and identification of related signaling pathways, upstream regulators, molecular interactions, disease process and candidate biomarkers are also available [
26].
4.6. DAVID Database
The DAVID database is a bioinformatics resource that assists in the analysis of a list of genes derived from high-throughput genomic sequencing experiments, using different tools such as functional annotation and gene functional classification. The analysis results help researchers gain overall understanding of the involved terms of gene ontology, signaling pathways and diseases within the genes of interest [
27].
4.7. Statistical Analysis
The expression values of target genes obtained from arrays of GEO database were analyzed using IBM SPSS Statistics for Windows, version 19 (IBM Corp., Armonk, NY, USA). To compare the differences of expression values between normal and RA groups, non-parametric method with Mann-Whitney U test was used. A statistically significant difference was determined by p-value < 0.05.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}