Infection of a French Population of Aedes albopictus and of Aedes aegypti (Paea Strain) with Zika Virus Reveals Low Transmission Rates to These Vectors’ Saliva


Abstract
:1. Introduction
2. Results
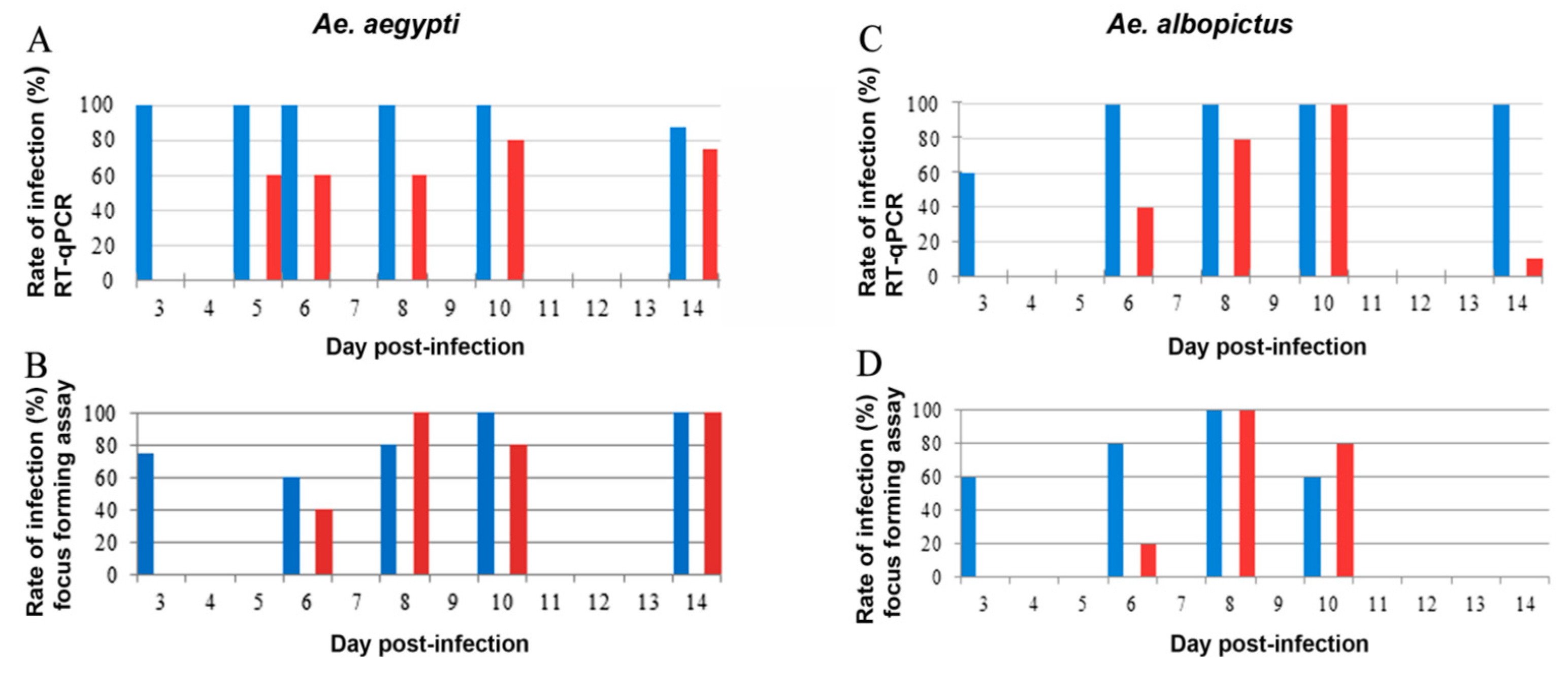
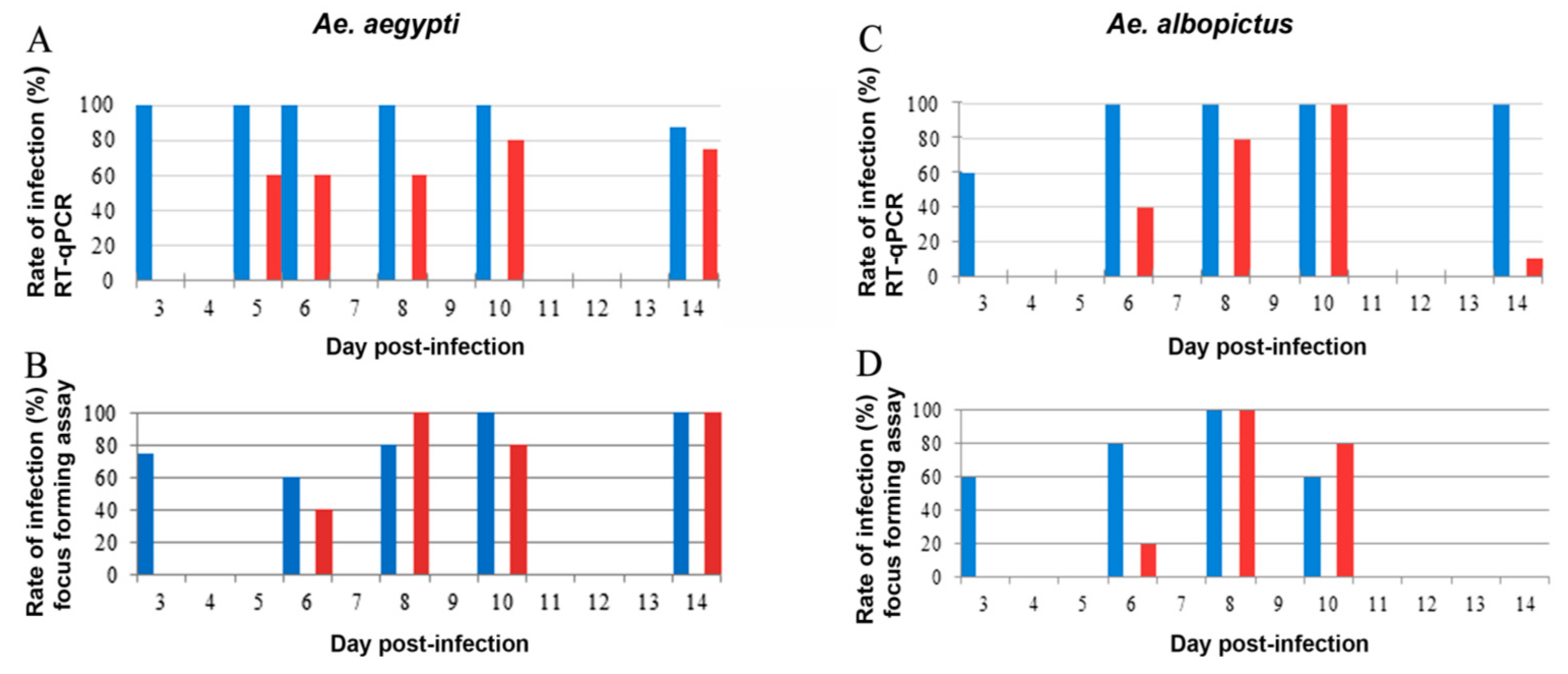
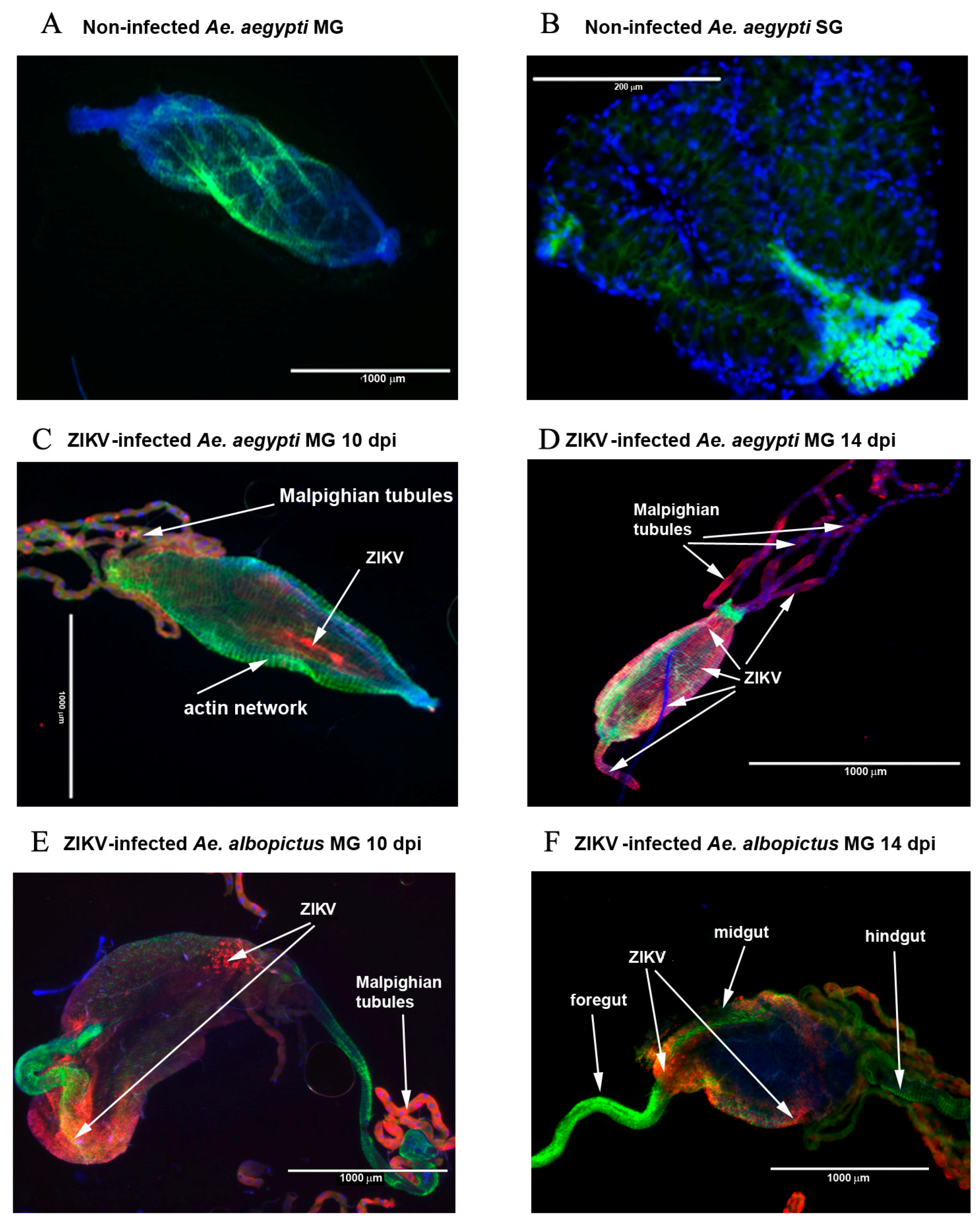
2.1. Rate of Infection of Ae. aegypti and Ae. albopictus Midguts and Salivary Glands
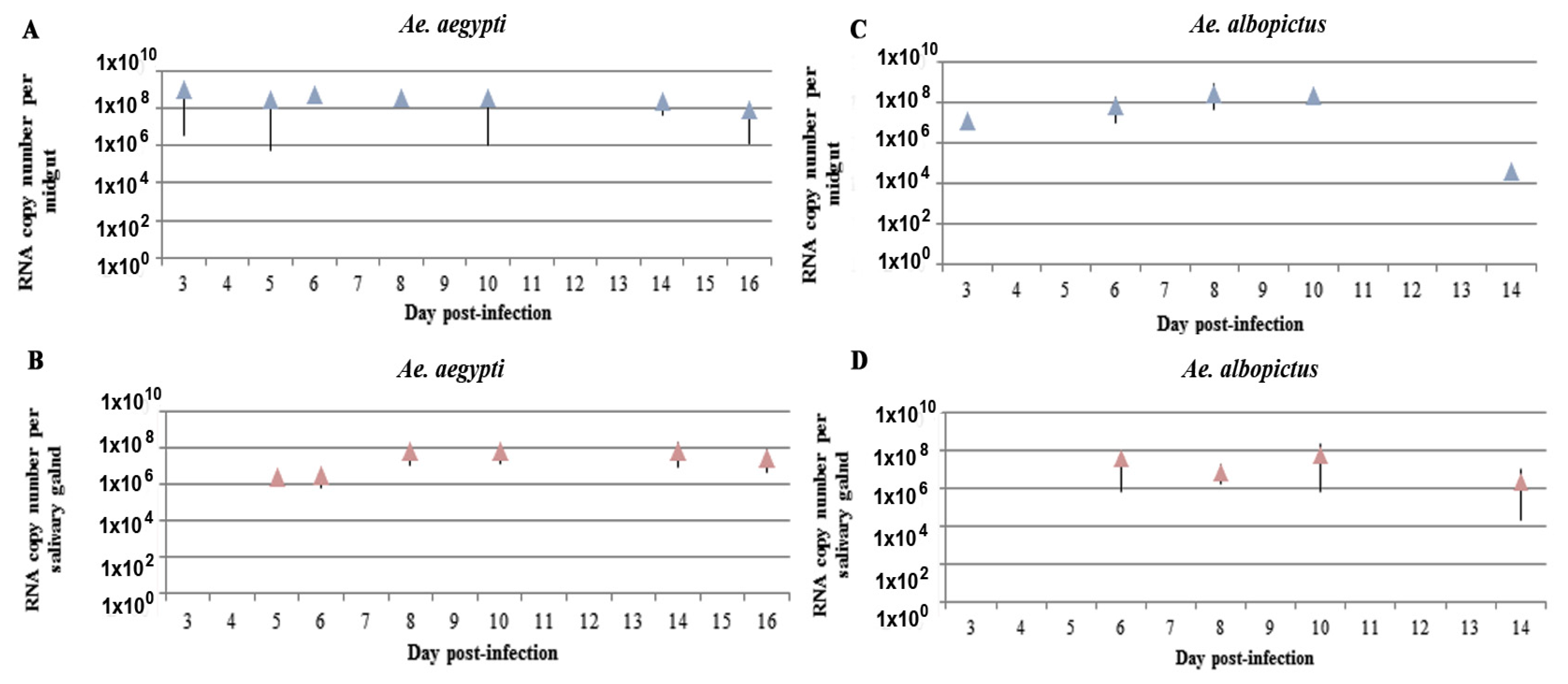
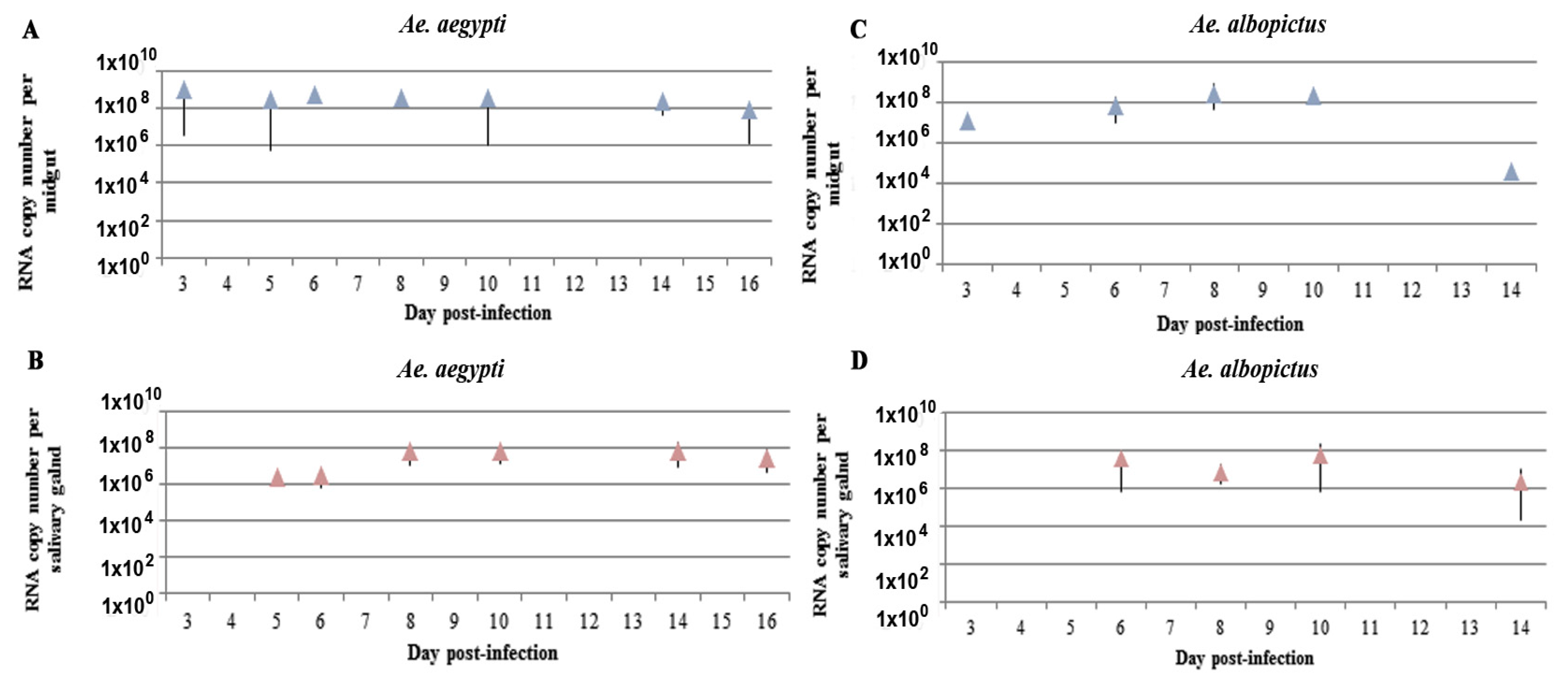
2.2. Kinetics of ZIKV RNA Levels in MG and SG of Aedes Mosquitoes
2.3. Transmission Rate of ZIKV in Aedes Saliva
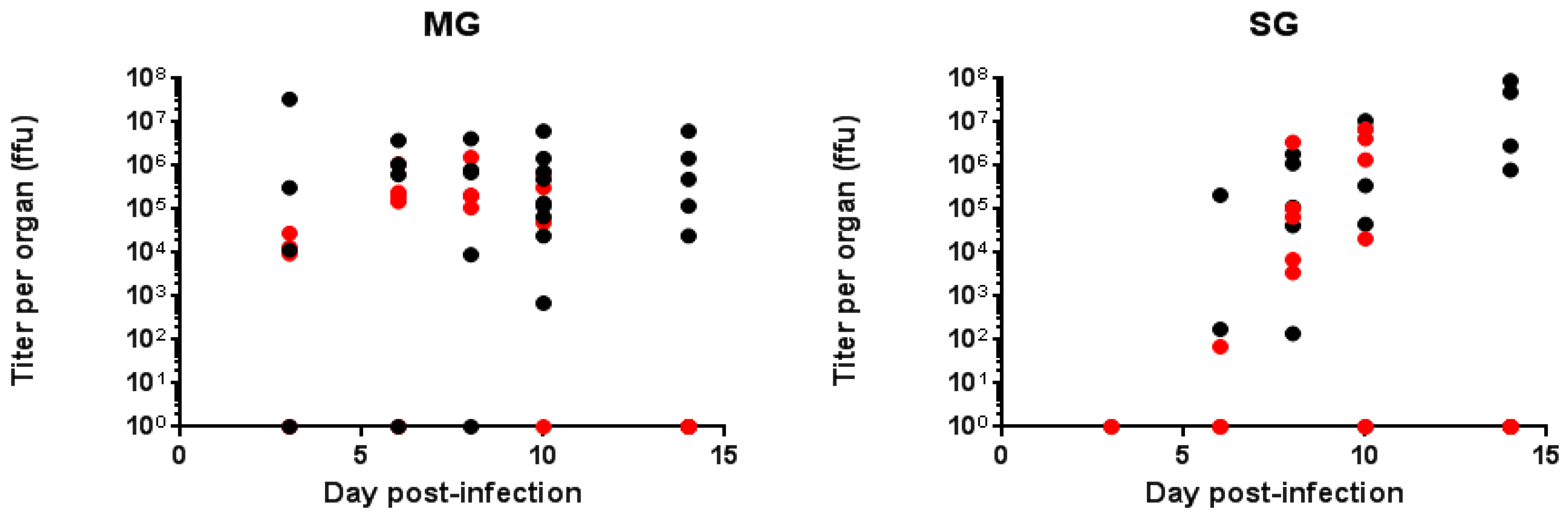
2.4. Analysis of Viral Particles in Both Mosquito Species
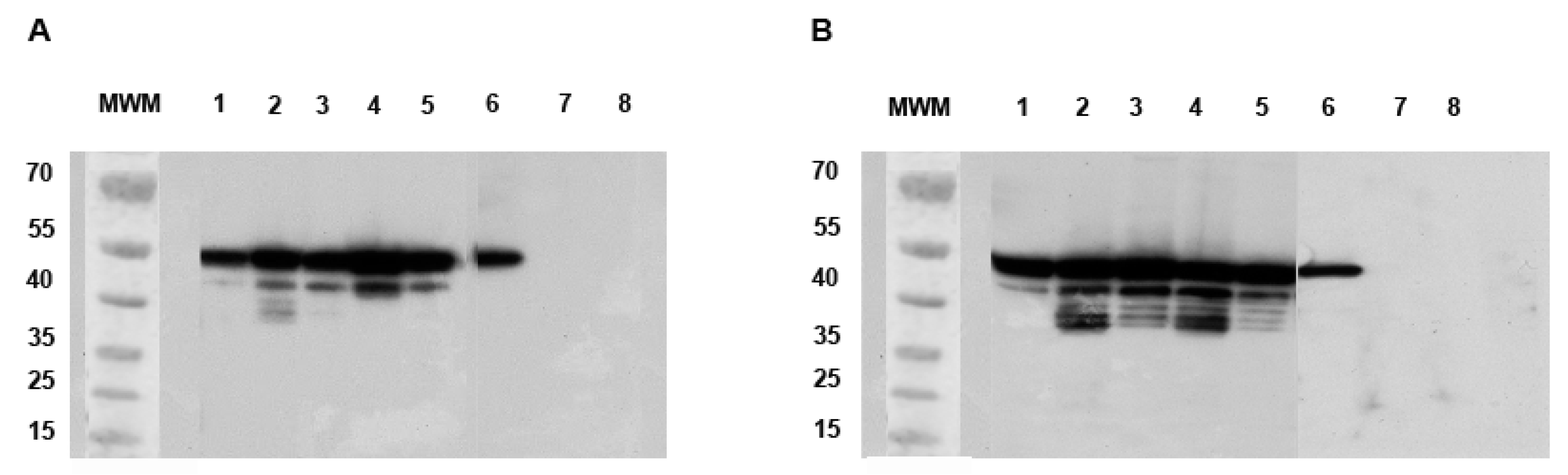
2.4.1. Quantification of Defective ZIKV Particles in SG and MG of Ae. aegypti and Ae. albopictus at Different Time Points
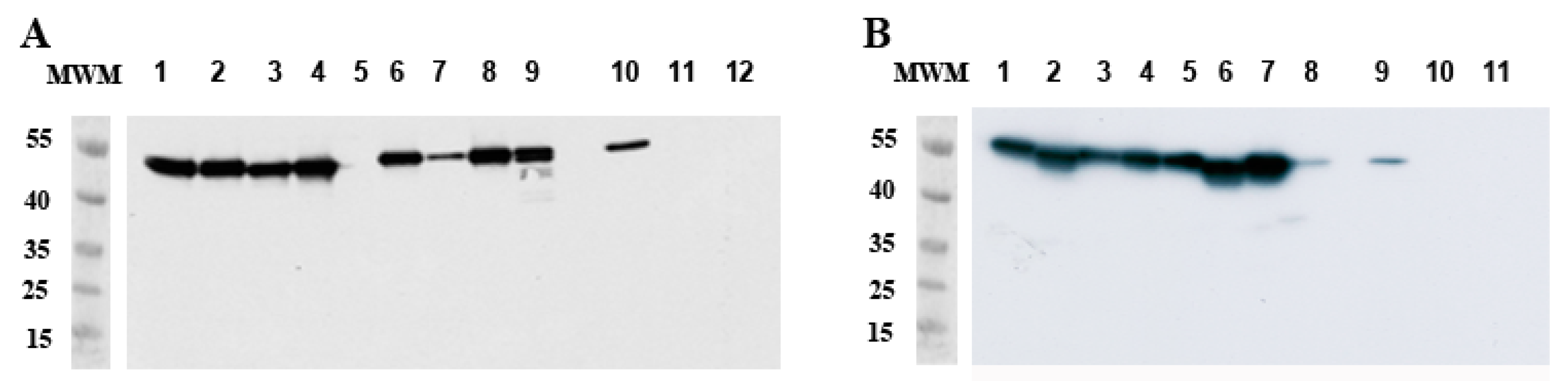
2.4.2. Degradation of Viral Proteins
3. Discussion
4. Materials and Methods
4.1. Viral Strain
4.2. Mosquito Population
4.3. Oral Infection of Mosquitoes and Dissections
4.4. Mosquito Salivation
4.5. Focus Forming Assay
4.6. RT-qPCR
4.7. Immunofluorescence
4.8. Western Blot
4.9. Dot-Blot
4.10. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MG | Midgut |
| SG | Salivary gland |
| ffa | Focus forming assay |
| ffu | Focus forming unit |
| dpi | Day post-infection |
References
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap island, federated states of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lormeau, V.M.; Blake, A.; Mons, S.; Lastere, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barre syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Musso, D.; Rouault, E.; Teissier, A.; Lanteri, M.C.; Zisou, K.; Broult, J.; Grange, E.; Nhan, T.X.; Aubry, M. Molecular detection of Zika virus in blood and RNA load determination during the French Polynesian outbreak. J. Med. Virol. 2017, 89, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, M.; Fischer, M.; Staples, J.E. Zika virus spreads to new areas—Region of the Americas, May 2015–January 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastere, S.; Valour, F.; Baudouin, L.; Mallet, H.; Musso, D.; Ghawche, F. Zika virus infection complicated by Guillain-Barre syndrome—Case report, French Polynesia, December 2013. Eurosurveillance 2014, 19, 20720. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonca, M.C.L.; de Oliveira, L.; et al. Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: A case study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Culjat, M.; Darling, S.E.; Nerurkar, V.R.; Ching, N.; Kumar, M.; Min, S.K.; Wong, R.; Grant, L.; Melish, M.E. Clinical and imaging findings in an infant with Zika embryopathy. Clin. Infect. Dis. 2016, 63, 805–811. [Google Scholar] [PubMed]
- Oliveira Melo, A.S.; Malinger, G.; Ximenes, R.; Szejnfeld, P.O.; Alves Sampaio, S.; Bispo de Filippis, A.M. Zika virus intrauterine infection causes fetal brain abnormality and microcephaly: Tip of the iceberg? Ultrasound Obstet. Gynecol. 2016, 47, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Faccini, L.; Ribeiro, E.M.; Feitosa, I.M.; Horovitz, D.D.; Cavalcanti, D.P.; Pessoa, A.; Doriqui, M.J.; Neri, J.I.; Neto, J.M.; Wanderley, H.Y.; et al. Possible association between Zika virus infection and microcephaly—Brazil, 2015. Morb. Mortal. Wkly. Rep. 2016, 65, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Schuler-Faccini, L.; Sanseverino, M.; Vianna, F.; da Silva, A.A.; Larrandaburu, M.; Marcolongo-Pereira, C.; Abeche, A.M. Zika virus: A new human teratogen? Implications for women of reproductive age. Clin. Pharmacol. Ther. 2016, 100, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.V.; Maia, M.; Bravo-Filho, V.; Gois, A.L.; Belfort, R., Jr. Zika virus in Brazil and macular atrophy in a child with microcephaly. Lancet 2016, 387, 228. [Google Scholar] [CrossRef]
- Ayres, C.F. Identification of Zika virus vectors and implications for control. Lancet Infect. Dis. 2016, 16, 278–279. [Google Scholar] [CrossRef]
- Saiz, J.C.; Blazquez, A.B.; De Oya, N.J.; Merino-Ramos, T.; Martin-Acebes, M.A.; Escribano-Romero, E.; Vazquez-Calvo, A. Response: Commentary: Zika virus: The latest newcomer. Front. Microbiol. 2016, 7, 1398. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.N.; Otaguiri, K.K.; Kashima, S.; Covas, D.T. Overview of Zika virus (ZIKV) infection in regards to the brazilian epidemic. Braz. J. Med. Biol. Res. 2016, 49, e5420. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, J.; Recker, M. The 2012 Madeira dengue outbreak: Epidemiological determinants and future epidemic potential. PLoS Negl. Trop. Dis. 2014, 8, e3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control (ECDC). Mosquito Maps. Aedes Aegypti—Current Known Distribution—September 2017 & Aedes albopictus—Current Known Distribution—September 2017 (ECDC, Stockholm, 2017). Available online: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps (accessed on 10 October 2017).
- Boorman, J.P.; Porterfield, J.S. A simple technique for infection of mosquitoes with viruses; transmission of Zika virus. Trans. R. Soc. Trop. Med. Hyg. 1956, 50, 238–242. [Google Scholar] [CrossRef]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Faye, O.; Ba, Y.; Hanley, K.A.; Buenemann, M.; Weaver, S.C.; Diallo, M. Zika virus emergence in mosquitoes in Southeastern Senegal, 2011. PLoS ONE 2014, 9, e109442. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Suhandiman, G.; Triwibowo, V. Zika virus, a cause of fever in central Java, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1981, 75, 389–393. [Google Scholar] [CrossRef]
- Marchette, N.J.; Garcia, R.; Rudnick, A. Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia. Am. J. Trop. Med. Hyg. 1969, 18, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Yakob, L.; Walker, T. Zika virus outbreak in the Americas: The need for novel mosquito control methods. Lancet Glob. Health 2016, 4, e148–e149. [Google Scholar] [CrossRef]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenco-de-Oliveira, R.; Failloux, A.B. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Jupille, H.; Seixas, G.; Mousson, L.; Sousa, C.A.; Failloux, A.B. Zika virus, a new threat for Europe? PLoS Negl. Trop. Dis. 2016, 10, e0004901. [Google Scholar] [CrossRef] [PubMed]
- Guzzetta, G.; Poletti, P.; Montarsi, F.; Baldacchino, F.; Capelli, G.; Rizzoli, A.; Rosa, R.; Merler, S. Assessing the potential risk of Zika virus epidemics in temperate areas with established Aedes albopictus populations. Eurosurveillance 2016, 21, 30199. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.M.; Moesker, B.; Rodenhuis-Zybert, I.; Wilschut, J. Flavivirus cell entry and membrane fusion. Viruses 2011, 3, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Fagbami, A.H. Zika virus infections in Nigeria: Virological and seroepidemiological investigations in Oyo State. J. Hyg. 1979, 83, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.L.; Causey, O.R.; Carey, D.E.; Reddy, S.; Cooke, A.R.; Akinkugbe, F.M.; David-West, T.S.; Kemp, G.E. Arthropod-borne viral infections of man in Nigeria, 1964–1970. Ann. Trop. Med. Parasitol. 1975, 69, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.I. Zika virus infection in man. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 335–338. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.M.; Roche, C.; Teissier, A.; Robin, E.; Berry, A.L.; Mallet, H.P.; Sall, A.A.; Musso, D. Zika virus, French Polynesia, South Pacific, 2013. Emerg. Infect. Dis. 2014, 20, 1085–1086. [Google Scholar] [CrossRef] [PubMed]
- Massad, E.; Burattini, M.N.; Khan, K.; Struchiner, C.J.; Coutinho, F.A.B.; Wilder-Smith, A. On the origin and timing of Zika virus introduction in Brazil. Epidemiol. Infect. 2017, 145, 2303–2312. [Google Scholar] [CrossRef] [PubMed]
- Venturi, G.; Zammarchi, L.; Fortuna, C.; Remoli, M.E.; Benedetti, E.; Fiorentini, C.; Trotta, M.; Rizzo, C.; Mantella, A.; Rezza, G.; et al. An autochthonous case of Zika due to possible sexual transmission, florence, Italy, 2014. Eurosurveillance 2016, 21, 30148. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Rouzeyrol, M.; Biron, A.; O’Connor, O.; Huguon, E.; Descloux, E. Infectious Zika viral particles in breastmilk. Lancet 2016, 387, 1051. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Akoua-Koffi, C.; Diarrassouba, S.; Benie, V.B.; Ngbichi, J.M.; Bozoua, T.; Bosson, A.; Akran, V.; Carnevale, P.; Ehouman, A. Investigation surrounding a fatal case of yellow fever in Côte d’Ivoire in 1999. Bull. Soc. Pathol. Exot. 2001, 94, 227–230. [Google Scholar] [PubMed]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Mboui Ondo, S.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika virus in Gabon (Central Africa)—2007: A new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8, e2681. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci. Data 2015, 2, 150035. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.S.; Li, M.Z.; Chong, C.S.; Ng, L.C.; Tan, C.H. Aedes (Stegomyia) albopictus (Skuse): A potential vector of Zika virus in singapore. PLoS Negl. Trop. Dis. 2013, 7, e2348. [Google Scholar] [CrossRef] [PubMed]
- Rocklov, J.; Quam, M.B.; Sudre, B.; German, M.; Kraemer, M.U.; Brady, O.; Bogoch, I.I.; Liu-Helmersson, J.; Wilder-Smith, A.; Semenza, J.C.; et al. Assessing seasonal risks for the introduction and mosquito-borne spread of Zika virus in Europe. EBioMedicine 2016, 9, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sanchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Nalim, S.; Tan, R.; Saipan, H.; Sulianti Saroso, J. Variation in susceptibility to oral infection with dengue viruses among geographic strains of Aedes aegypti. Am. J. Trop. Med. Hyg. 1979, 28, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Rosen, L. Variation among geographic strains of Aedes albopictus in susceptibility to infection with dengue viruses. Am. J. Trop. Med. Hyg. 1976, 25, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Li, M.I.; Wong, P.S.; Ng, L.C.; Tan, C.H. Oral susceptibility of Singapore Aedes (Stegomyia) aegypti (linnaeus) to Zika virus. PLoS Negl. Trop. Dis. 2012, 6, e1792. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rua, A.; Zouache, K.; Caro, V.; Diancourt, L.; Delaunay, P.; Grandadam, M.; Failloux, A.B. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the southeast of France. PLoS ONE 2013, 8, e59716. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika virus infection in human skin cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [PubMed]
- Smartt, C.T.; Stenn, T.M.S.; Chen, T.Y.; Teixeira, M.G.; Queiroz, E.P.; Souza Dos Santos, L.; Queiroz, G.A.N.; Ribeiro Souza, K.; Kalabric Silva, L.; Shin, D.; et al. Evidence of Zika virus RNA fragments in Aedes albopictus (Diptera: Culicidae) field-collected eggs from Camacari, Bahia, Brazil. J. Med. Entomol. 2017, 23, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Bialosuknia, S.M.; Ehrbar, D.J.; Kramer, L.D. Vertical transmission of Zika virus by Aedes aegypti and Ae. albopictus mosquitoes. Emerg. Infect. Dis. 2017, 23, 880–882. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day Post-Infection | 8 | 10 | 14 | 17 |
|---|---|---|---|---|
| Ae. aegypti | ||||
| % infection | 11 | 33 | 16 | 6.7 |
| RNA copy number mean | 7.4 × 104 | 3.2 × 105 | 1.1 × 105 | 1.6 × 105 |
| Standard error | - | 2.1 × 105 | 6.8 × 104 | - |
| Number of saliva tested | 9 | 9 | 25 | 15 |
| Ae. albopictus | ||||
| % infection | 0 | 28.6 | 25 | Not collected |
| RNA copy number mean | 4.5 × 104 | 5 × 104 | ||
| Standard error | 2.6 × 104 | 3 × 104 | ||
| Number of saliva tested | 9 | 9 | 16 | |
| Day Post-Infection | 8 | 8 | 10 | 10 | 14 | 14 |
|---|---|---|---|---|---|---|
| ZIKV − Saliva | ZIKV + Saliva | ZIKV − Saliva | ZIKV + Saliva | ZIKV − Saliva | ZIKV + Saliva | |
| RNA copy number per total Ae. aegypti mosquito | ||||||
| Mean | 3.4 × 107 | 2.5 × 107 | 8.0 × 107 | 5.2 × 107 | 1.3 × 108 | 2.5 × 108 |
| SD | 7.6 × 107 | - | 6.1 × 107 | 4.5 × 107 | 1.9 × 108 | 1.5 × 108 |
| N | 8 | 1 | 7 | 3 | 2 | 4 |
| RNA copy number per total Ae. albopictus mosquito | ||||||
| Mean | 6.1 × 107 | - | 8.8 × 107 | 2.1 × 108 | 2.1 × 108 * | 9.4 × 108 * |
| SD | 7.6 × 107 | - | 6.1 × 107 | 2.5 × 108 | 1.9 × 108 | 8.32 × 108 |
| N | 8 | 4 | 2 | 5 | 6 | |
| Day Post-Infection | Ae. aegypti | Ae. albopictus | ||||||
|---|---|---|---|---|---|---|---|---|
| Midguts | Salivary Glands | Midguts | Salivary Glands | |||||
| Viral Copy/ffu | R | Viral Copy/ffu | R | Viral Copy:ffu | R | Viral Copy/ffu | R | |
| 3 | 1.16 × 102 | 3 | - | 4.26 × 102 | 10.9 | - | ||
| 6 | 2.46 × 102 | 6.3 | 1.48 × 101 | 0.4 | 1.63 × 102 | 4.2 | 1.5 × 102 | 3.8 |
| 8 | 2.31 × 102 | 5.9 | 1.07 × 102 | 2.7 | 3.98 × 102 | 10.2 | 8.80 | 0.2 |
| 10 | 1.31 × 102 | 3.3 | 1.63 × 101 | 0.4 | 6.47 × 102 | 16.6 | 1.81 × 101 | 0.5 |
| 14 | 1.49 × 102 | 3.8 | 1.67 | 0.04 | - | - | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryckebusch, F.; Berthet, M.; Missé, D.; Choumet, V. Infection of a French Population of Aedes albopictus and of Aedes aegypti (Paea Strain) with Zika Virus Reveals Low Transmission Rates to These Vectors’ Saliva. Int. J. Mol. Sci. 2017, 18, 2384. https://doi.org/10.3390/ijms18112384
Ryckebusch F, Berthet M, Missé D, Choumet V. Infection of a French Population of Aedes albopictus and of Aedes aegypti (Paea Strain) with Zika Virus Reveals Low Transmission Rates to These Vectors’ Saliva. International Journal of Molecular Sciences. 2017; 18(11):2384. https://doi.org/10.3390/ijms18112384
Chicago/Turabian StyleRyckebusch, Faustine, Michèle Berthet, Dorothée Missé, and Valérie Choumet. 2017. "Infection of a French Population of Aedes albopictus and of Aedes aegypti (Paea Strain) with Zika Virus Reveals Low Transmission Rates to These Vectors’ Saliva" International Journal of Molecular Sciences 18, no. 11: 2384. https://doi.org/10.3390/ijms18112384