Alteration of Vascular Responsiveness to Uridine Adenosine Tetraphosphate in Aortas Isolated from Male Diabetic Otsuka Long-Evans Tokushima Fatty Rats: The Involvement of Prostanoids

 ,
,
Abstract
:1. Introduction
2. Results
2.1. General Parameters
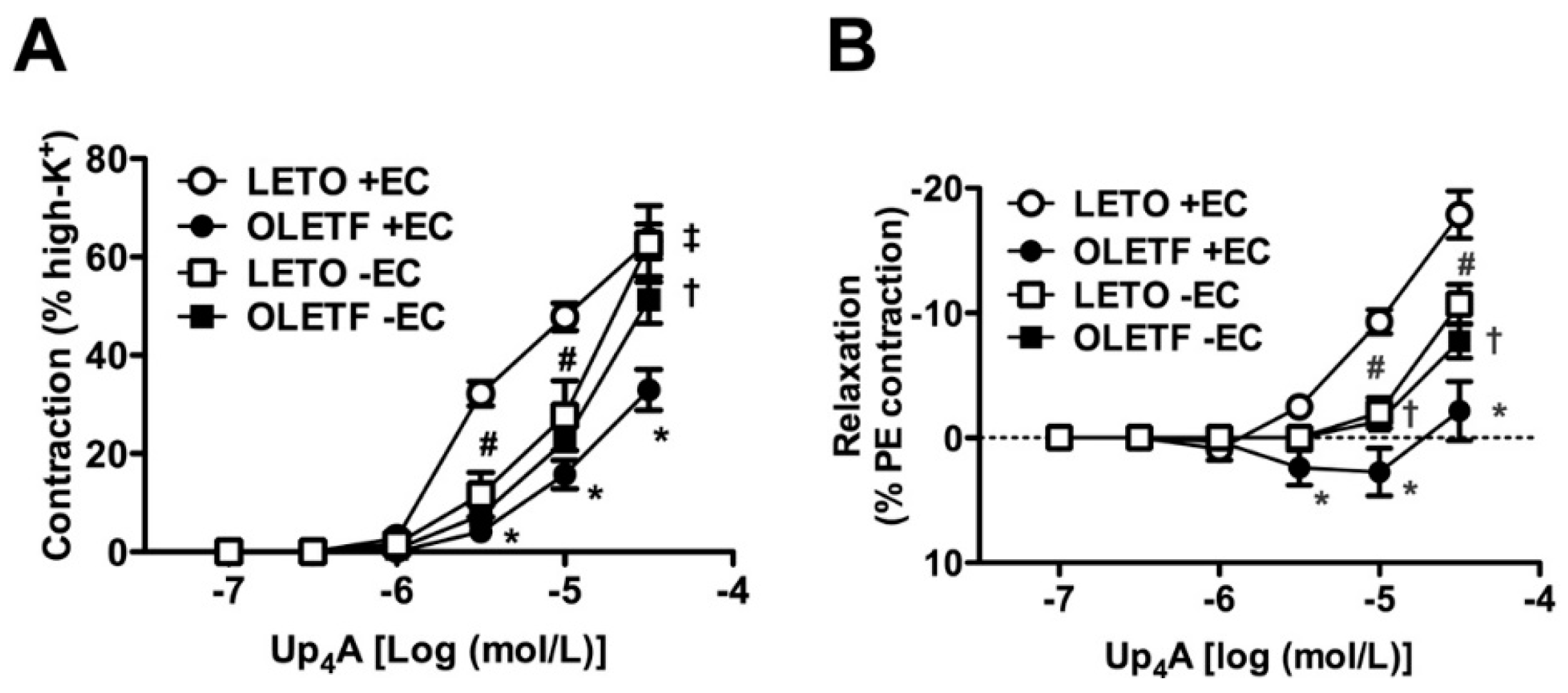
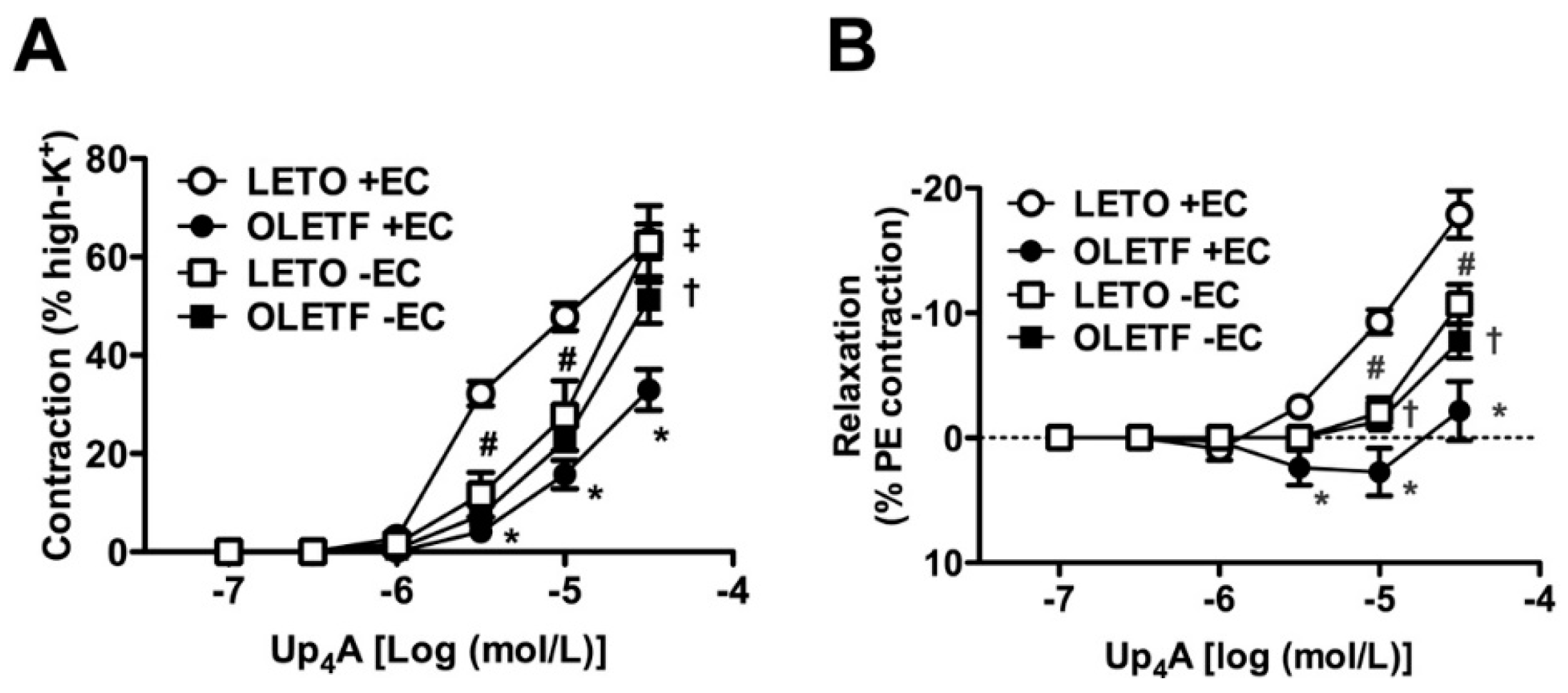
2.2. Role of Endothelium in Up4A-Mediated Responses in the Aorta
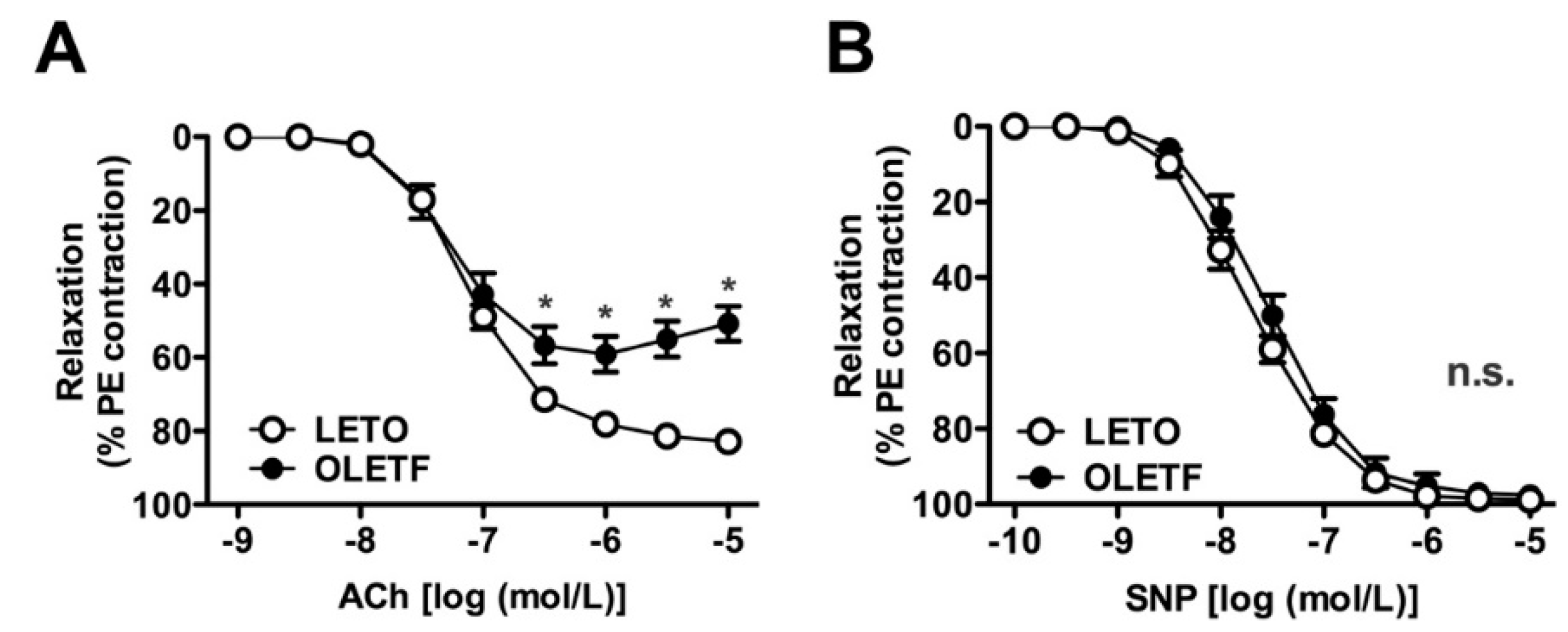
2.3. Relaxation Induced by Acetylcholine and Sodium Nitroprusside in Endothelium-Intact Aortas
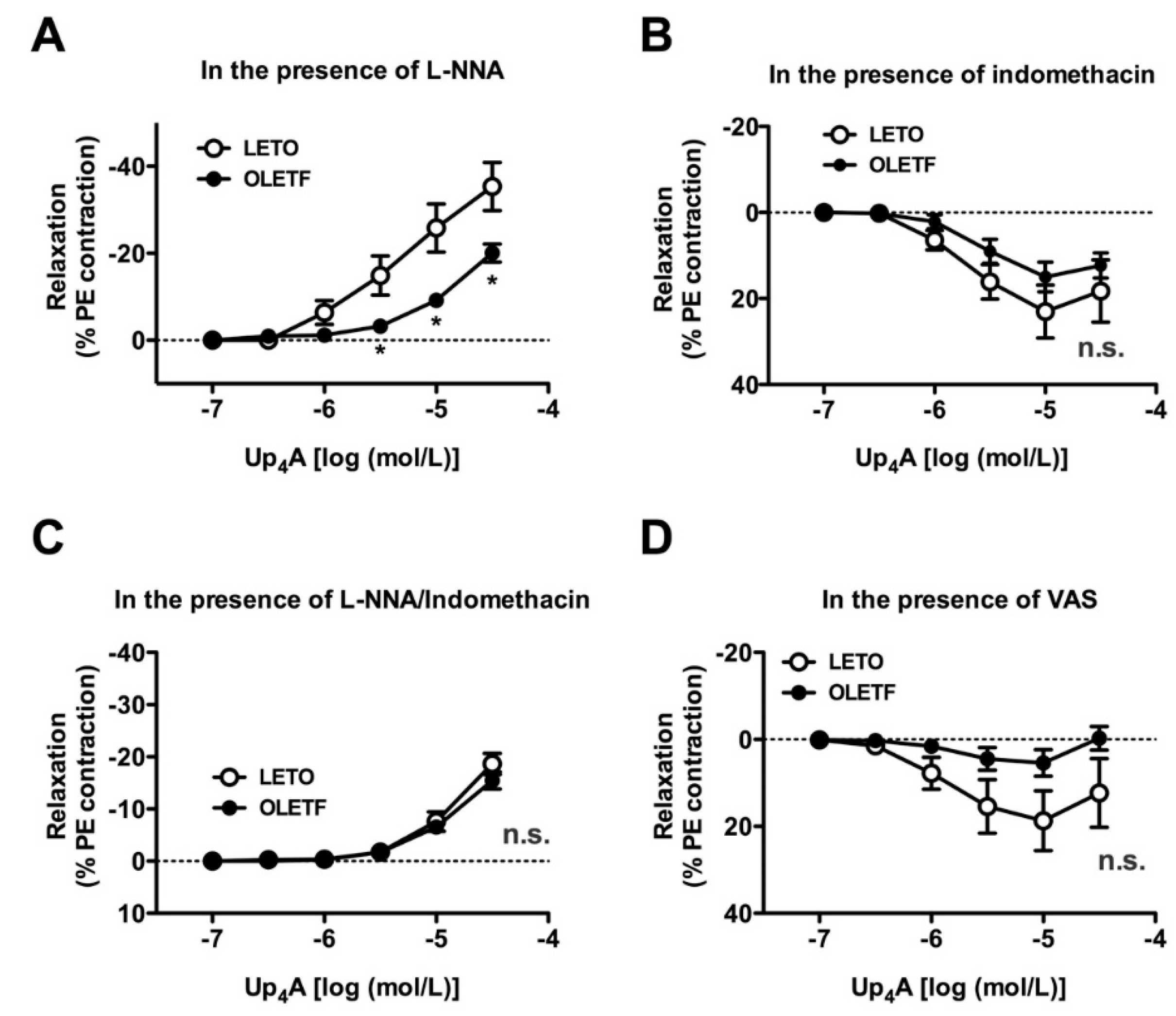
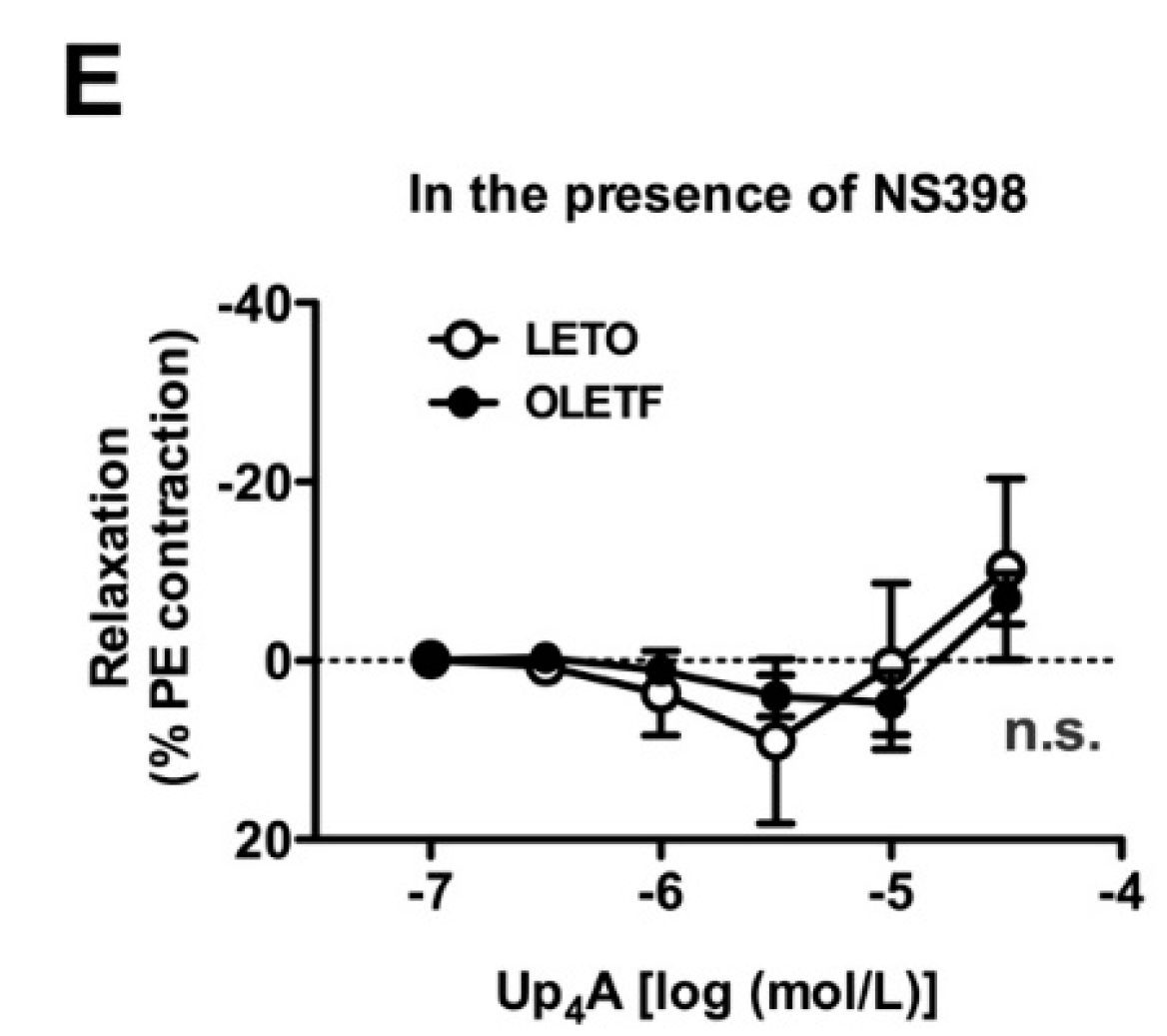
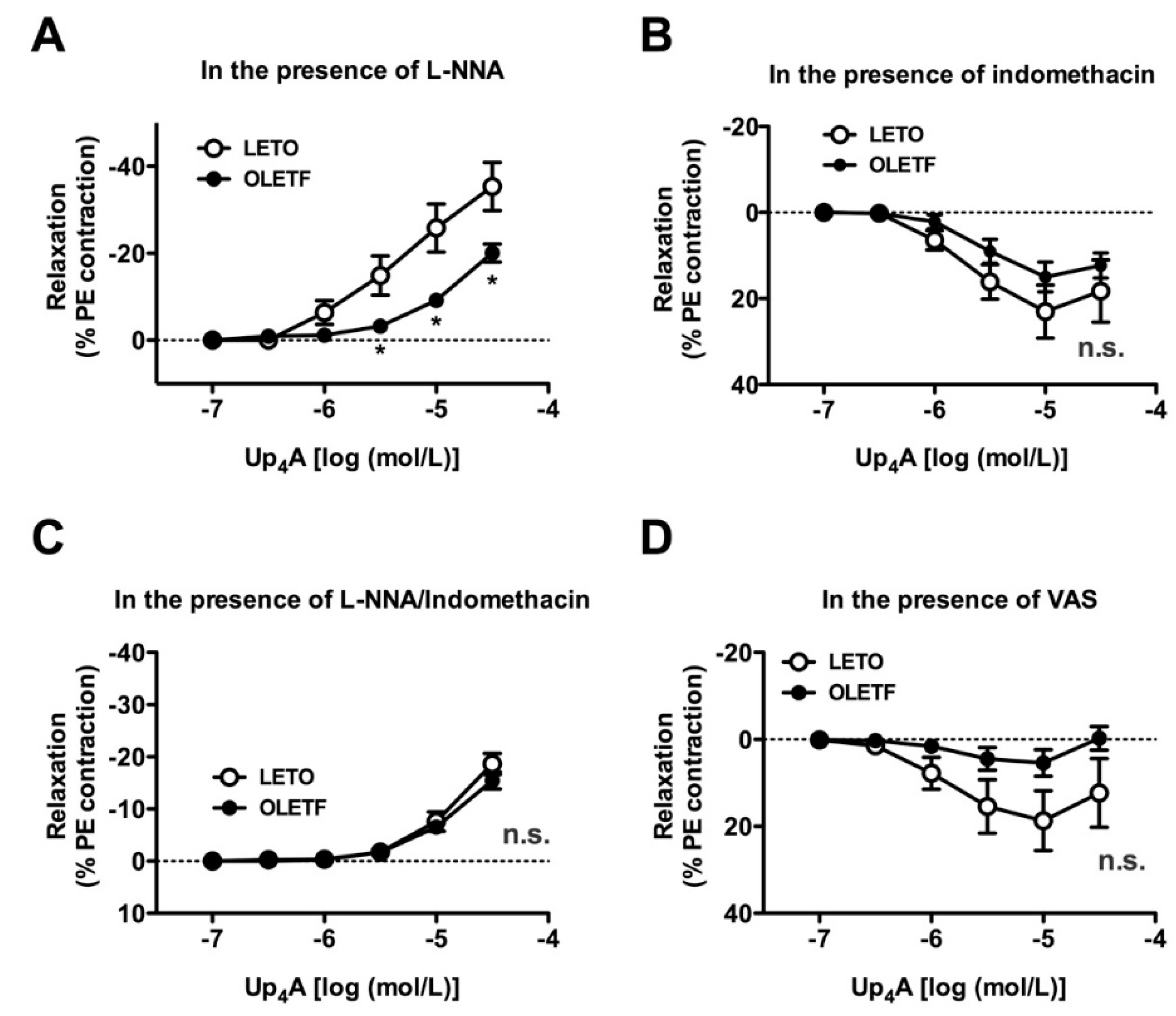
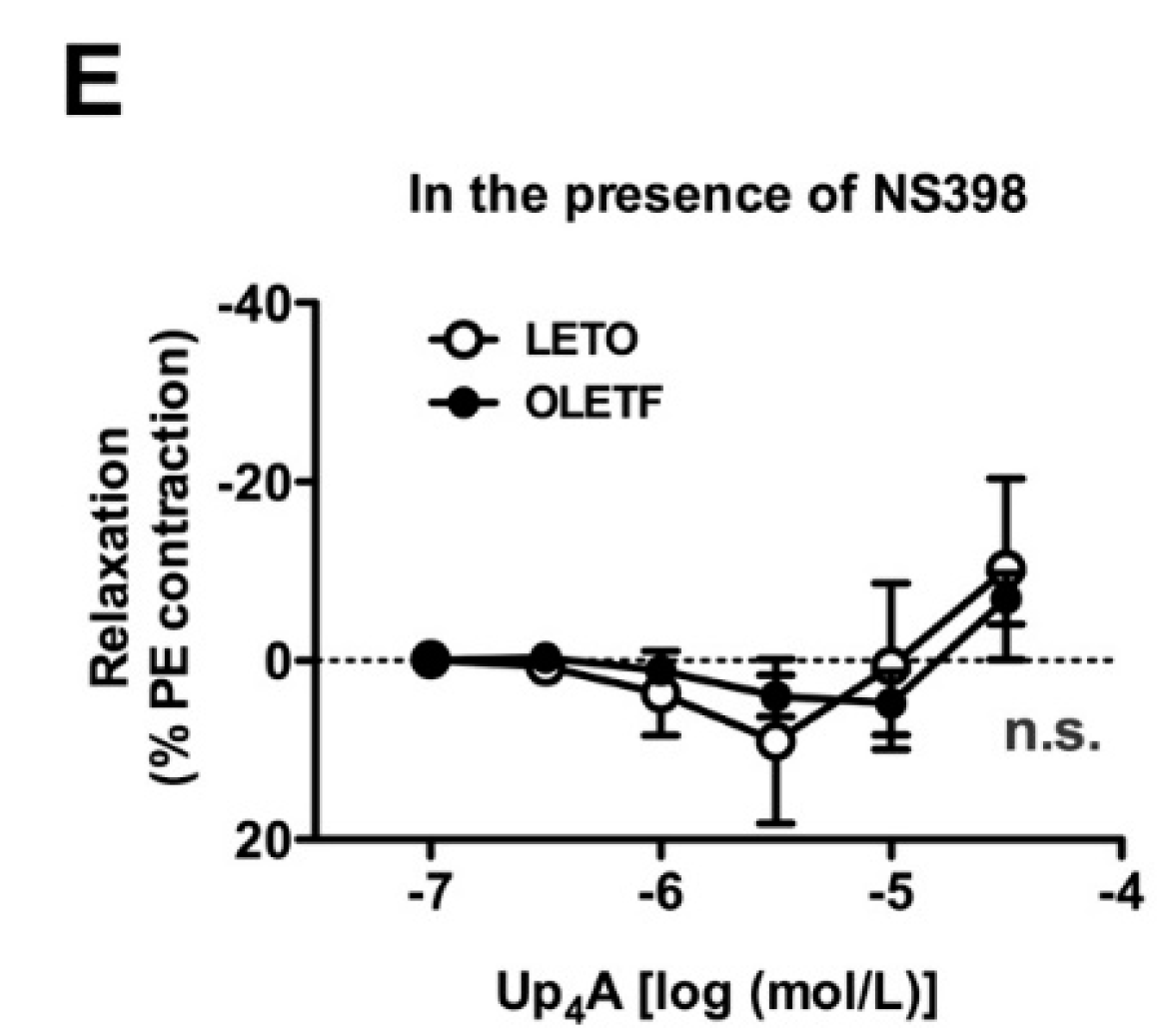
2.4. Effects of Nitric Oxide Synthase (NOS) and COX Inhibitors on Up4A-Induced Aortic Relaxation
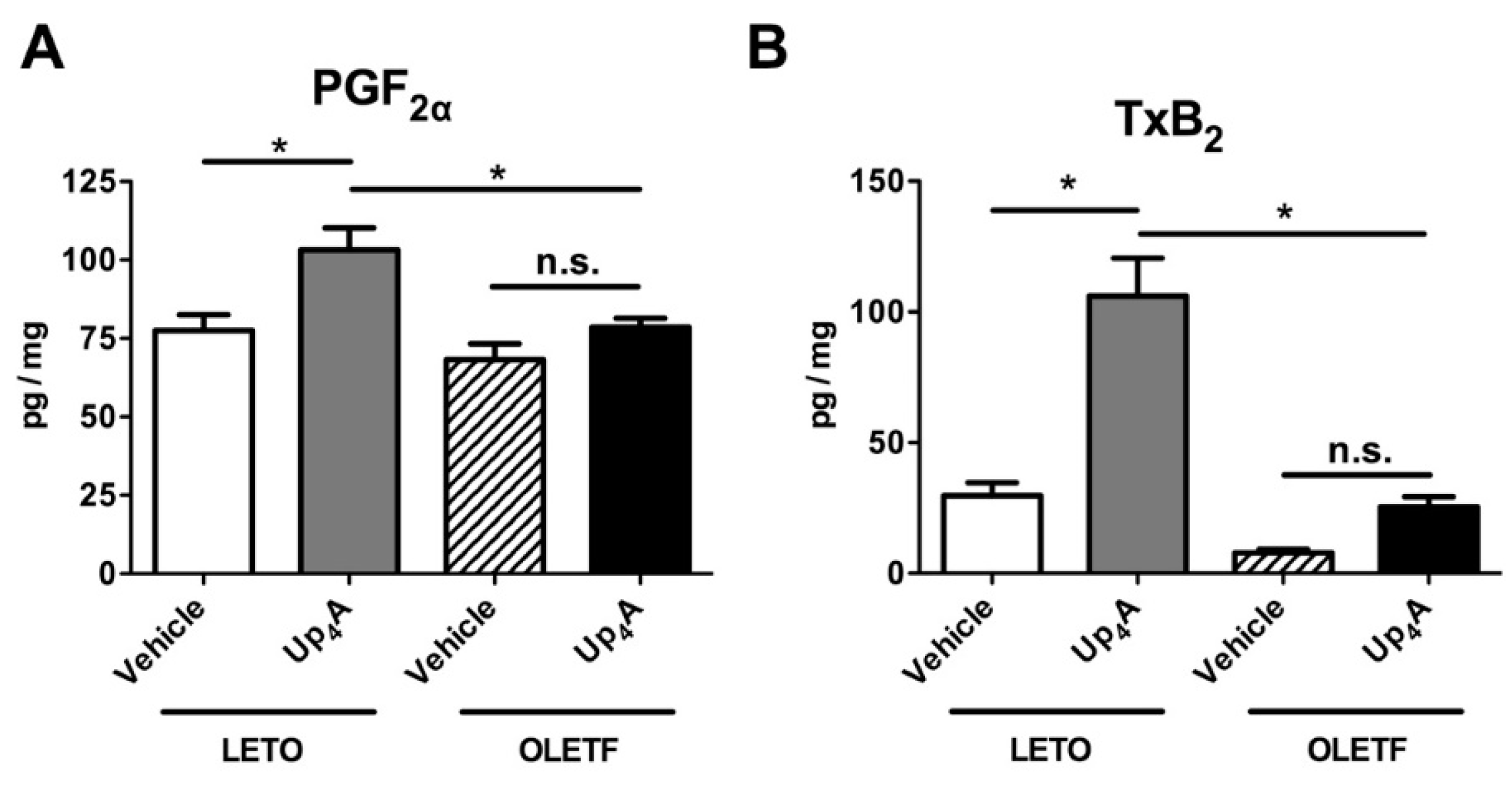
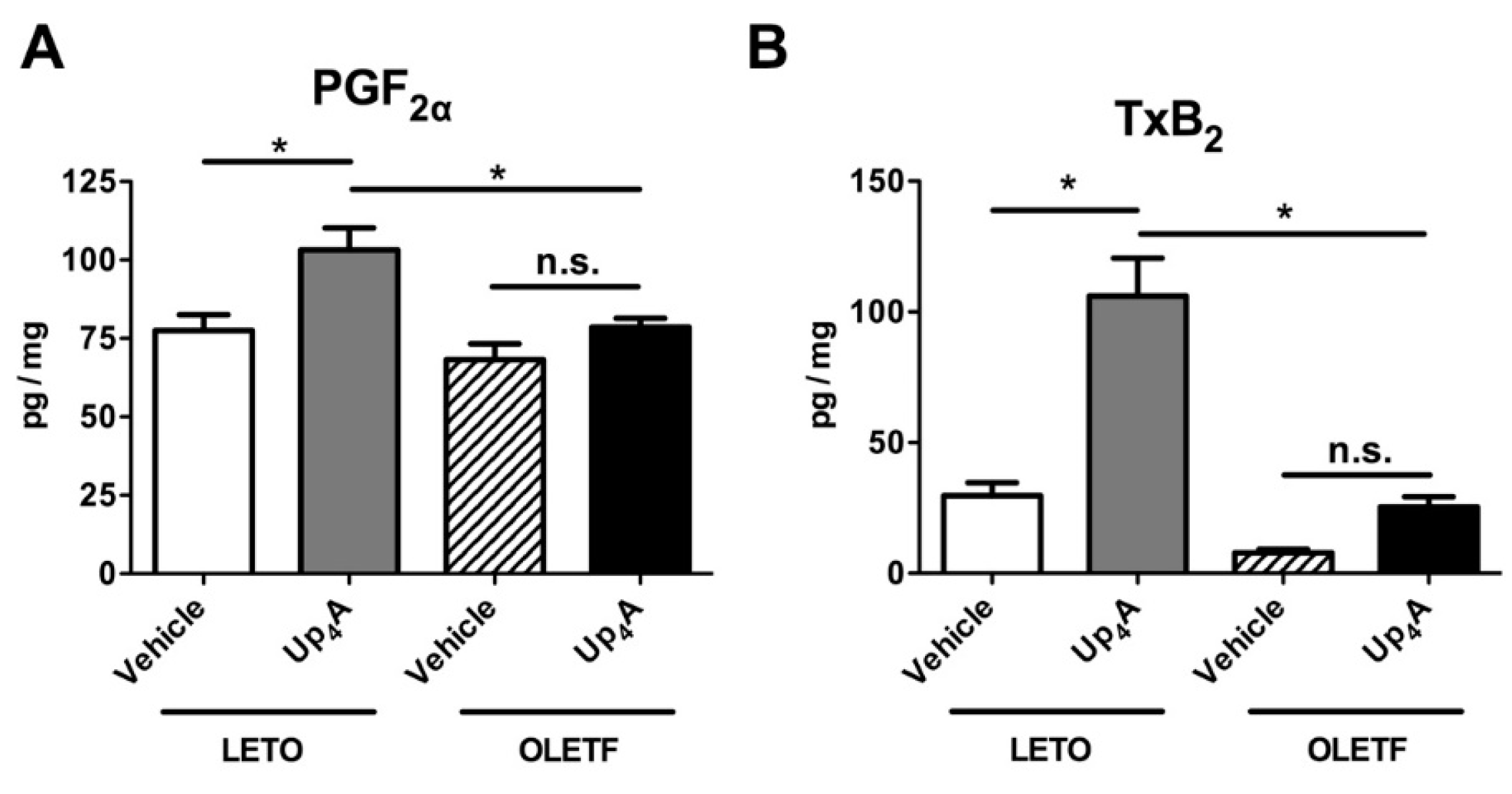
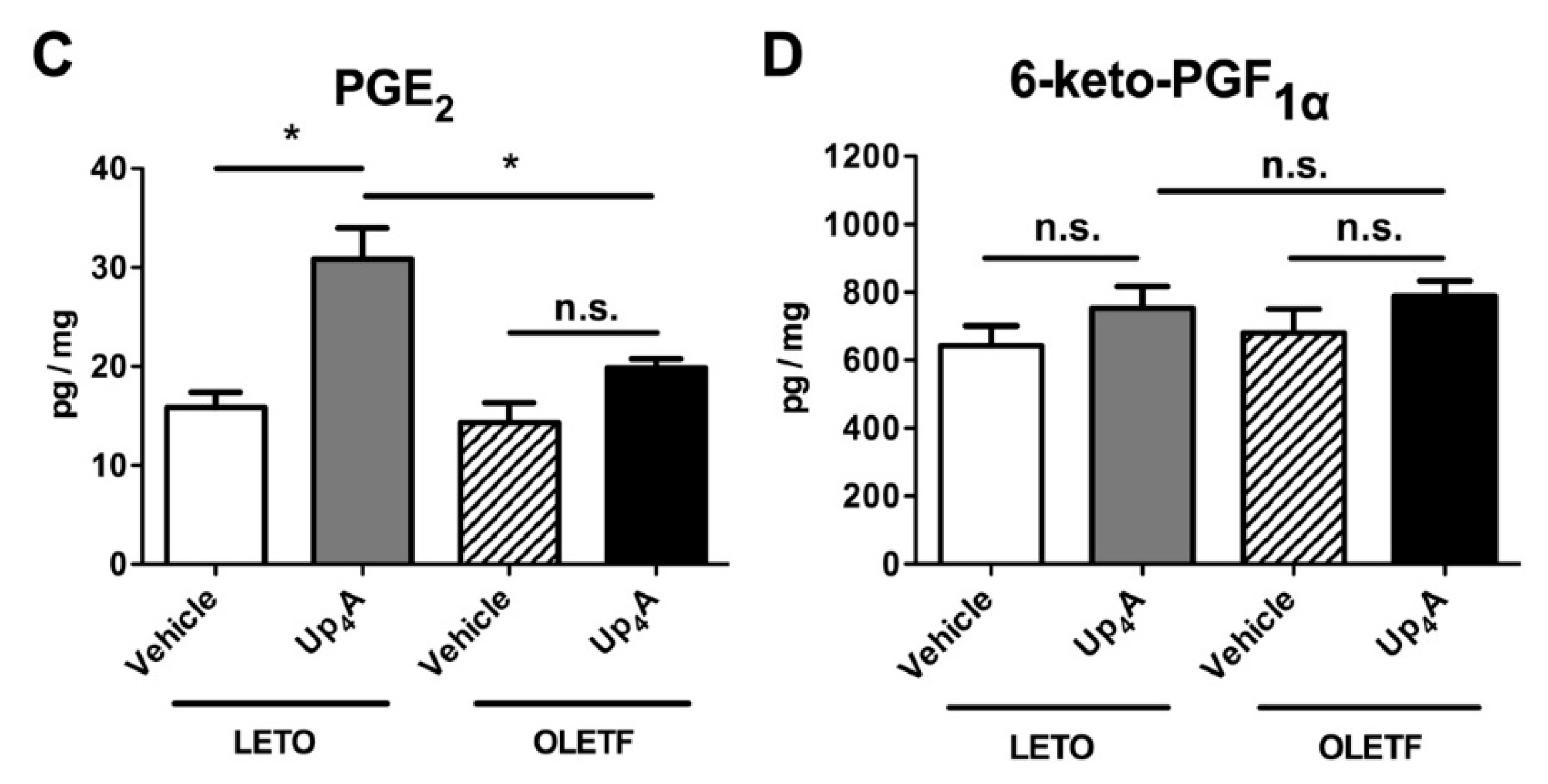
2.5. Effect of Up4A on Prostanoid Release in the Aorta
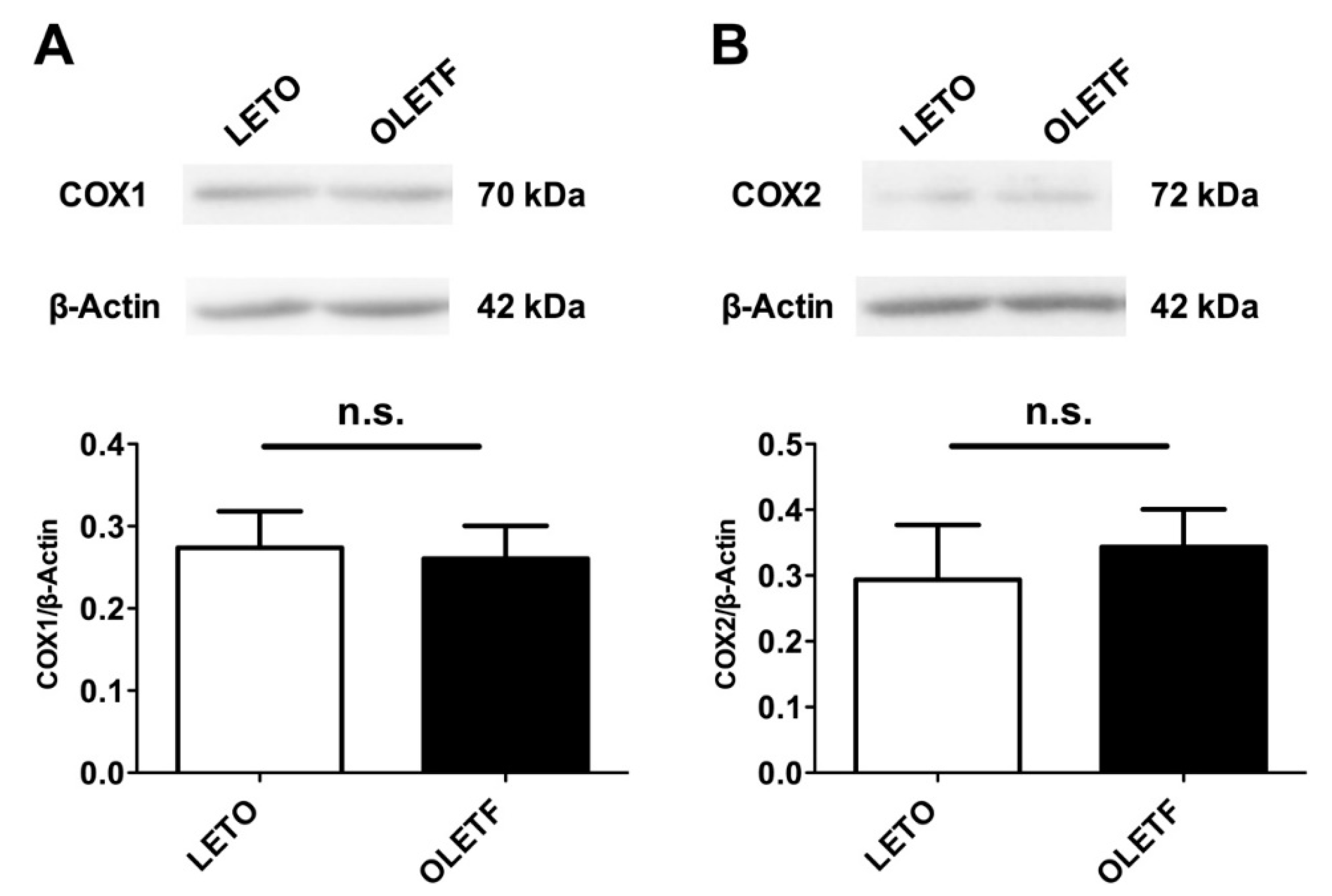
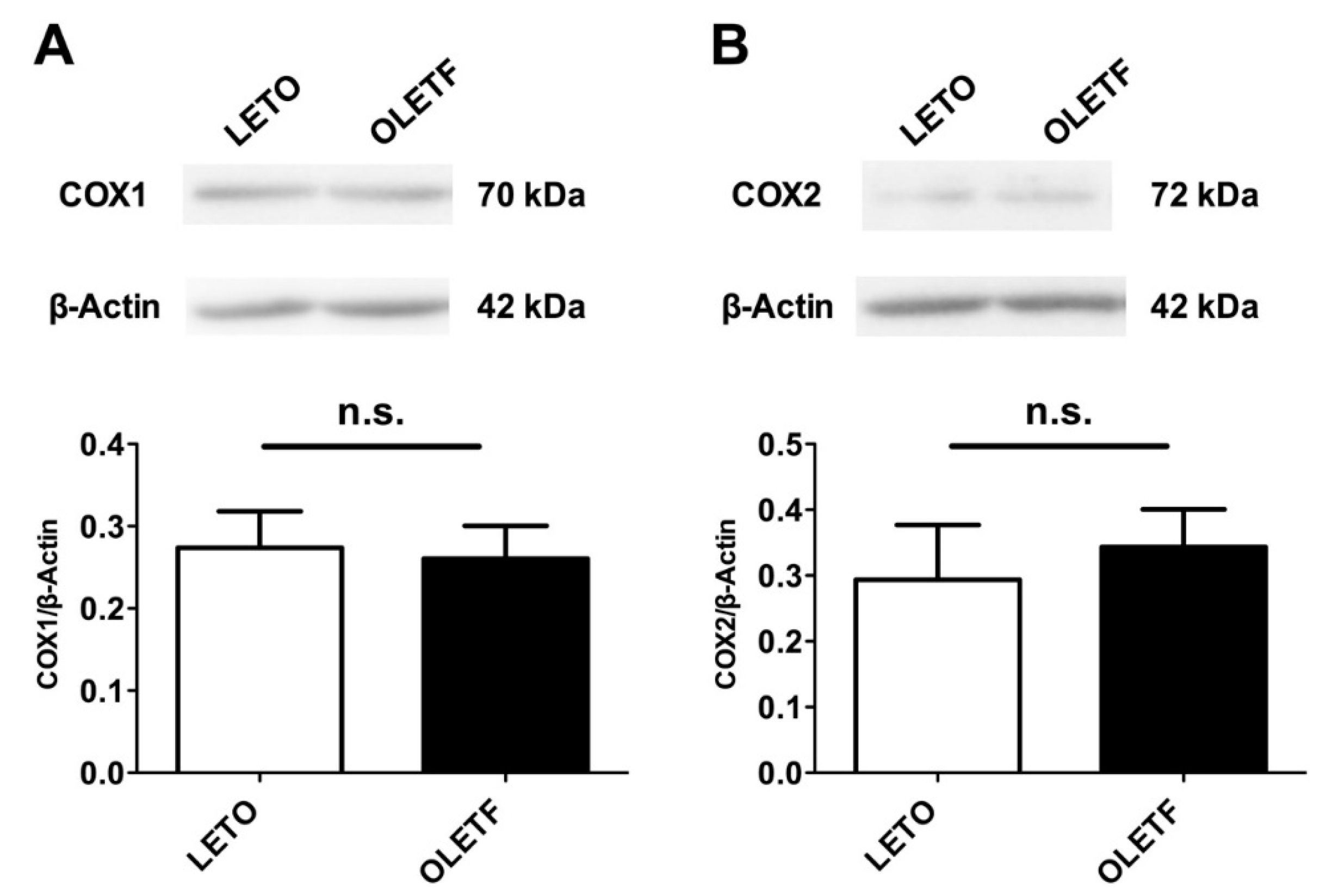
2.6. Expressions of COX Proteins in the Aorta
3. Discussion
4. Materials and Methods
4.1. Animals and Procedures
4.2. Preparation of Rat Aortic Rings and Evaluation of Vascular Function
4.3. Measurement of Prostanoid Release
4.4. Western Blotting
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACh | Acetylcholine |
| ANOVA | Analysis of variance |
| COX | Cyclooxygenase |
| DOCA | Deoxycorticosterone acetate |
| EDCF | Endothelium-derived contracting factor |
| EDRF | Endothelium-derived relaxing factor |
| GK | Goto-Kakizaki |
| KHS | Krebs-Henseleit solution |
| LETO | Long-Evans Tokushima Otsuka |
| L-NNA | NG-nitro-l-arginine |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| OLETF | Otsuka Long-Evans Tokushima Fatty |
| PE | Phenylephrine |
| PG | Prostaglandin |
| SBP | Systolic blood pressure |
| SNP | Sodium nitroprusside |
| TP | Thromboxane-prostanoid receptor |
| Tx | Thromboxane |
| Up4A | Uridine adenosine tetraphosphate |
| VAS | Valeroyl salicylate |
References
- Ferrannini, E.; DeFronzo, R.A. Impact of glucose-lowering drugs on cardiovascular disease in type 2 diabetes. Eur. Heart J. 2015, 36, 2288–2296. [Google Scholar] [CrossRef] [PubMed]
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Munzel, T. Targeting vascular (endothelial) dysfunction. Br. J. Pharmacol. 2017, 174, 1591–1619. [Google Scholar] [CrossRef] [PubMed]
- McNicholas, E.; Abdelhafiz, A.H. Diabetes management in older people: A focus on cardiovascular risk reduction. Postgrad. Med. 2017, 129, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Urbina, E.M. Vascular and endothelial function in youth with type 2 diabetes mellitus. Curr. Diabetes Rep. 2017, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Barton, M. Obesity and aging: Determinants of endothelial cell dysfunction and atherosclerosis. Pflug. Arch. 2010, 460, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Goligorski, M.S. Vascular endothelium in diabetes. Am. J. Physiol. Renal Physiol. 2017, 312, F266–F275. [Google Scholar] [CrossRef] [PubMed]
- Michel, T.; Vanhoutte, P.M. Cellular signaling and NO production. Pflug. Arch. 2010, 459, 807–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nava, E.; Llorens, S. The paracrine control of vascular motion. A historical perspective. Pharmacol. Res. 2016, 113, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Shimokawa, H.; Feletou, M.; Tang, E.H. Endothelial dysfunction and vascular disease—A 30th anniversary update. Acta Physiol. 2017, 219, 22–96. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Zhao, Y.; Leung, S.W. Thirty years saying NO: Sources, fate, actions, and misfortunes of the endothelium-derived vasodilator mediator. Circ. Res. 2016, 119, 375–396. [Google Scholar] [CrossRef] [PubMed]
- Feletou, M.; Huang, Y.; Vanhoutte, P.M. Vasoconstrictor prostanoids. Pflug. Arch. 2010, 459, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feletou, M.; Huang, Y.; Vanhoutte, P.M. Endothelium-mediated control of vascular tone: COX-1 and COX-2 products. Br. J. Pharmacol. 2011, 164, 894–912. [Google Scholar] [CrossRef] [PubMed]
- Feletou, M.; Kohler, R.; Vanhoutte, P.M. Nitric oxide: Orchestrator of endothelium-dependent responses. Ann. Med. 2012, 44, 694–716. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Goulopoulou, S.; Taguchi, K.; Tostes, R.C.; Kobayashi, T. Constrictor prostanoids and uridine adenosine tetraphosphate: Vascular mediators and therapeutic targets in hypertension and diabetes. Br. J. Pharmacol. 2015, 172, 3980–4001. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Hida, M.; Narimatsu, H.; Matsumoto, T.; Kobayashi, T. Glucose and angiotensin II-derived endothelial extracellular vesicles regulate endothelial dysfunction via ERK1/2 activation. Pflug. Arch. 2017, 469, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Triggle, C.R. Endothelial dysfunction in diabetes: Multiple targets for treatment. Pflug. Arch. 2010, 459, 977–994. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Tolle, M.; Vanholder, R.; Schonfelder, G.; van der Giet, M.; Henning, L.; Schluter, H.; Paul, M.; Zidek, W.; Jankowski, J. Uridine adenosine tetraphosphate: A novel endothelium-derived vasoconstrictive factor. Nat. Med. 2005, 11, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; He, G.; Walsh, M.P.; Zheng, X.L. Signaling mechanisms mediating uridine adenosine tetraphosphate-induced proliferation of human vascular smooth muscle cells. J. Cardiovasc. Pharmacol. 2011, 58, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Walsh, M.P.; Jankowski, V.; Jankowski, J.; Zheng, X.L. Up4A stimulates endothelium-independent contraction of isolated rat pulmonary artery. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, L733–L738. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.B.; Hristovska, A.; Wolff, H.; Vanhoutte, P.; Jensen, B.L.; Bie, P. Uridine adenosine tetraphosphate affects contractility of mouse aorta and decreases blood pressure in conscious rats and mice. Acta Physiol. 2010, 200, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Meyer, A.A.; Schlattmann, P.; Gui, Y.; Zheng, X.L.; Stamcou, I.; Radtke, K.; Tran, T.N.; van der Giet, M.; Tolle, M.; et al. Increased uridine adenosine tetraphosphate concentrations in plasma of juvenile hypertensives. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1776–1781. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, V.; Patzak, A.; Herget-Rosenthal, S.; Tran, T.N.; Lai, E.Y.; Gunthner, T.; Buschmann, I.; Zidek, W.; Jankowski, J. Uridine adenosine tetraphosphate acts as an autocrine hormone affecting glomerular filtration rate. J. Mol. Med. 2008, 86, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Linder, A.E.; Tumbri, M.; Linder, F.F.; Webb, R.C.; Leite, R. Uridine adenosine tetraphosphate induces contraction and relaxation in rat aorta. Vasc. Pharmacol. 2008, 48, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Tostes, R.C.; Webb, R.C. The role of uridine adenosine tetraphosphate in the vascular system. Adv. Pharmacol. Sci. 2011, 2011, 435132. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Tostes, R.C.; Webb, R.C. Alterations in vasoconstrictor responses to the endothelium-derived contracting factor uridine adenosine tetraphosphate are region specific in DOCA-salt hypertensive rats. Pharmacol. Res. 2012, 65, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Tostes, R.C.; Webb, R.C. Uridine adenosine tetraphosphate-induced contraction is increased in renal but not pulmonary arteries from DOCA-salt hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H409–H417. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Watanabe, S.; Ando, M.; Yamada, K.; Iguchi, M.; Taguchi, K.; Kobayashi, T. Diabetes and age-related differences in vascular function of renal artery: Possible involvement of endoplasmic reticulum stress. Rejuvenation Res. 2016, 19, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Watanabe, S.; Kawamura, R.; Taguchi, K.; Kobayashi, T. Enhanced uridine adenosine tetraphosphate-induced contraction in renal artery from type 2 diabetic Goto-Kakizaki rats due to activated cyclooxygenase/thromboxane receptor axis. Pflug. Arch. 2014, 466, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, M.; Prufer, J.; Prufer, N.; Wiedon, A.; Huang, T.; Chebli, M.; Jankowski, V.; Jankowski, J.; Schafer-Korting, M.; Zidek, W.; et al. The endothelium-derived contracting factor uridine adenosine tetraphosphate induces P2Y(2)-mediated pro-inflammatory signaling by monocyte chemoattractant protein-1 formation. J. Mol. Med. 2011, 89, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, M.; Tolle, M.; Prufer, J.; Prufer, N.; Huang, T.; Jankowski, V.; Jankowski, J.; Zidek, W.; van der Giet, M. Uridine adenosine tetraphosphate activation of the purinergic receptor P2Y enhances in vitro vascular calcification. Kidney Int. 2012, 81, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Tolle, M.; Schuchardt, M.; Wiedon, A.; Huang, T.; Klockel, L.; Jankowski, J.; Jankowski, V.; Zidek, W.; van der Giet, M. Differential effects of uridine adenosine tetraphosphate on purinoceptors in the rat isolated perfused kidney. Br. J. Pharmacol. 2010, 161, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Wiedon, A.; Tolle, M.; Bastine, J.; Schuchardt, M.; Huang, T.; Jankowski, V.; Jankowski, J.; Zidek, W.; van der Giet, M. Uridine adenosine tetraphosphate (Up4A) is a strong inducer of smooth muscle cell migration via activation of the P2Y2 receptor and cross-communication to the PDGF receptor. Biochem. Biophys. Res. Commun. 2012, 417, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Chrifi, I.; Xu, Y.; Pernow, J.; Duncker, D.J.; Merkus, D.; Cheng, C. Uridine adenosine tetraphosphate acts as a proangiogenic factor in vitro through purinergic P2Y receptors. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H299–H309. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; de Wijs-Meijler, D.; Lankhuizen, I.; Jankowski, J.; Jankowski, V.; Jan Danser, A.H.; Duncker, D.J.; Merkus, D. Blunted coronary vasodilator response to uridine adenosine tetraphosphate in post-infarct remodeled myocardium is due to reduced P1 receptor activation. Pharmacol. Res. 2013, 77, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Merkus, D.; Cheng, C.; Duckers, H.J.; Jan Danser, A.H.; Duncker, D.J. Uridine adenosine tetraphosphate is a novel vasodilator in the coronary microcirculation which acts through purinergic P1 but not P2 receptors. Pharmacol. Res. 2013, 67, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sorop, O.; de Beer, V.J.; Heinonen, I.; Cheng, C.; Jan Danser, A.H.; Duncker, D.J.; Merkus, D. Altered purinergic signaling in uridine adenosine tetraphosphate-induced coronary relaxation in swine with metabolic derangement. Purinergic Signal. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sun, C.; Tilley, S.L.; Mustafa, S.J. Mechanisms underlying uridine adenosine tetraphosphate-induced vascular contraction in mouse aorta: Role of thromboxane and purinergic receptors. Vasc. Pharmacol. 2015, 73, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yadav, V.R.; Sun, C.; Teng, B.; Mustafa, J.S. Impaired aortic contractility to uridine adenosine tetraphosphate in angiotensin II-induced hypertensive mice: Receptor desensitization? Am. J. Hypertens. 2017, 30, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Kawano, K.; Hirashima, T.; Mori, S.; Natori, T. OLETF (Otsuka Long-Evans Tokushima Fatty) rat: A new NIDDM rat strain. Diabetes Res. Clin. Pract. 1994, 24, S317–S320. [Google Scholar] [CrossRef]
- Kawano, K.; Hirashima, T.; Mori, S.; Saitoh, Y.; Kurosumi, M.; Natori, T. Spontaneous long-term hyperglycemic rat with diabetic complications Otsuka Long-Evans Tokushima Fatty (OLETF) strain. Diabetes 1992, 41, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Moran, T.H. Obesity in the Otsuka Long Evans Tokushima Fatty Rat: Mechanisms and Discoveries. Front. Nutr. 2016, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Kagota, S.; Yamaguchi, Y.; Nakamura, K.; Kunitomo, M. Altered endothelium-dependent responsiveness in the aortas and renal arteries of Otsuka Long-Evans Tokushima Fatty (OLETF) rats, a model of non-insulin-dependent diabetes mellitus. Gen. Pharmacol. 2000, 34, 201–209. [Google Scholar] [CrossRef]
- Liu, P.; Xie, Q.; Wei, T.; Chen, Y.; Chen, H.; Shen, W. Activation of the NLRP3 inflammasome induces vascular dysfunction in obese OLETF rats. Biochem. Biophys. Res. Commun. 2015, 468, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Ishida, K.; Nakayama, N.; Taguchi, K.; Kobayashi, T.; Kamata, K. Mechanisms underlying the losartan treatment-induced improvement in the endothelial dysfunction seen in mesenteric arteries from type 2 diabetic rats. Pharmacol. Res. 2010, 62, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Ishida, K.; Kobayashi, T.; Kamata, K. Pyrrolidine dithocarbamate reduces vascular prostanoid-induced responses in aged type 2 diabetic rat model. J. Pharmacol. Sci. 2009, 110, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kakami, M.; Noguchi, E.; Kobayashi, T.; Kamata, K. Imbalance between endothelium-derived relaxing and contracting factors in mesenteric arteries from aged OLETF rats, a model or Type 2 diabetes. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1480–H1490. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Nakayama, N.; Ishida, K.; Kobayashi, T.; Kamata, K. Eicosapentaenoic acid improves imbalance between vasodilator and vasoconstrictor actions of endothelium-derived factors in mesenteric arteries from rats at chronic stage of type 2 diabetes. J. Pharmacol. Exp. Ther. 2009, 329, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Noguchi, E.; Ishida, K.; Kobayashi, T.; Yamada, N.; Kamata, K. Metformin normalizes endothelial function by suppressing vasoconstrictor prostanoids in mesenteric arteries from OLETF rats, a model of type 2 diabetes. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1165–H1176. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, S.; Taguchi, K.; Matsumoto, T.; Kamata, K.; Kobayashi, T. Pravastatin normalizes ET-1-induced contraction in the aorta of type 2 diabetic OLETF rats by suppressing the KSR1/ERK complex. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H893–H902. [Google Scholar] [CrossRef] [PubMed]
- Saito, F.; Kawaguchi, M.; Izumida, J.; Asakura, T.; Maehara, K.; Maruyama, Y. Alteration in haemodynamics and pathological changes in the cardiovascular system during the development of Type 2 diabetes mellitus in OLETF rats. Diabetologia 2003, 46, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Nemoto, S.; Ishida, K.; Taguchi, K.; Matsumoto, T.; Kamata, K. Involvement of CaM kinase II in the impairment of endothelial function and eNOS activity in aortas of Type 2 diabetic rats. Clin. Sci. 2012, 123, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Roviezzo, F.; Bucci, M.; Brancaleone, V.; Di Lorenzo, A.; Geppetti, P.; Farneti, S.; Parente, L.; Lungarella, G.; Fiorucci, S.; Cirino, G. Proteinase-activated receptor-2 mediated arterial vasodilation in diabetes. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2349–2354. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Morishige, A.; Matsumoto, T.; Kamata, K.; Kobayashi, T. Enhanced estradiol-induced vasorelaxation in aortas from type 2 diabetic mice may reflect a compensatory role of p38 MAPK-mediated eNOS activation. Pflug. Arch. 2012, 464, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Matsumoto, T.; Taguchi, K.; Kamata, K.; Kobayashi, T. Mechanisms underlying altered extracellular nucleotide-induced contractions in mesenteric arteries from rats in later-stage type 2 diabetes: Effect of ANG II type 1 receptor antagonism. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1850–H1861. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Matsumoto, T.; Taguchi, K.; Kamata, K.; Kobayashi, T. Protein kinase C delta contributes to increase in EP3 agonist-induced contraction in mesenteric arteries from type 2 diabetic Goto-Kakizaki rats. Pflug. Arch. 2012, 463, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Gluais, P.; Vanhoutte, P.M.; Feletou, M. Mechanisms underlying ATP-induced endothelium-dependent contractions in the SHR aorta. Eur. J. Pharmacol. 2007, 556, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, F.; Eto, M.; De Paolis, P.; van der Loo, B.; Bachschmid, M.; Ullrich, V.; Kouroedov, A.; Delli Gatti, C.; Joch, H.; Volpe, M.; et al. High glucose causes upregulation of cyclooxygenase-2 and alters prostanoid profile in human endothelial cells: Role of protein kinase C and reactive oxygen species. Circulation 2003, 107, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Wu, J.L.; Zhang, M.; Xu, J.; Zou, M.H. Endothelial nitric oxide synthase-dependent tyrosine nitration of prostacyclin synthase in diabetes in vivo. Diabetes 2006, 55, 3133–3141. [Google Scholar] [CrossRef] [PubMed]
- Tang, E.H.; Vanhoutte, P.M. Gene expression changes of prostanoid synthases in endothelial cells and prostanoid receptors in vascular smooth muscle cells caused by aging and hypertension. Physiol. Genom. 2008, 32, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Abiru, T.; Machida, H.; Kasuya, Y.; Kamata, K. Endothelium-derived hyperpolarizing factor does not contribute to the decrease in endothelium-dependent relaxation in the aorta of streptozotocin-induced diabetic rats. Gen. Pharmacol. 1995, 26, 149–153. [Google Scholar] [CrossRef]
- Burnstock, G. Control of vascular tone by purines and pyrimidines. Br. J. Pharmacol. 2010, 161, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic Signaling and Endothelium. Curr. Vasc. Pharmacol. 2016, 14, 130–145. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic Signaling in the Cardiovascular System. Circ. Res. 2017, 120, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Delicado, E.G.; Miras-Portugal, M.T.; Carrasquero, L.M.; Leon, D.; Perez-Sen, R.; Gualix, J. Dinucleotide polyphosphates and their interaction with other nucleotide signaling pathways. Pflug. Arch. 2006, 452, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Hernandez, M.; Pintor, J.; Miras-Portugal, M.T. Modulation of the dinucleotide receptor present in rat midbrain synaptosomes by adenosine and ATP. Br. J. Pharmacol. 2000, 130, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Sunggip, C.; Oda, S.; Numaga-Tomita, T.; Tsuda, M.; Nishida, M. Purinergic P2Y receptors: Molecular diversity and implications for treatment of cardiovascular diseases. Pharmacol. Ther. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tagashira, H.; Matsumoto, T.; Taguchi, K.; Zhang, C.; Han, F.; Ishida, K.; Nemoto, S.; Kobayashi, T.; Fukunaga, K. Vascular endothelial σ1-receptor stimulation with SA4503 rescues aortic relaxation via Akt/eNOS signaling in ovariectomized rats with aortic banding. Circ. J. 2013, 77, 2831–2840. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (g) | Blood Glucose (mg/dL) | Systolic Blood Pressure (mmHg) |
|---|---|---|---|
| LETO | 574.7 ± 10.7 (21) | 108.5 ± 3.6 (21) | 123 ± 4 (9) |
| OLETF | 521.0 ± 14.7 (20) * | 380.9 ± 28.8 (20) * | 146 ± 6 (8) * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, T.; Kobayashi, S.; Ando, M.; Iguchi, M.; Takayanagi, K.; Kojima, M.; Taguchi, K.; Kobayashi, T. Alteration of Vascular Responsiveness to Uridine Adenosine Tetraphosphate in Aortas Isolated from Male Diabetic Otsuka Long-Evans Tokushima Fatty Rats: The Involvement of Prostanoids. Int. J. Mol. Sci. 2017, 18, 2378. https://doi.org/10.3390/ijms18112378
Matsumoto T, Kobayashi S, Ando M, Iguchi M, Takayanagi K, Kojima M, Taguchi K, Kobayashi T. Alteration of Vascular Responsiveness to Uridine Adenosine Tetraphosphate in Aortas Isolated from Male Diabetic Otsuka Long-Evans Tokushima Fatty Rats: The Involvement of Prostanoids. International Journal of Molecular Sciences. 2017; 18(11):2378. https://doi.org/10.3390/ijms18112378
Chicago/Turabian StyleMatsumoto, Takayuki, Shota Kobayashi, Makoto Ando, Maika Iguchi, Keisuke Takayanagi, Mihoka Kojima, Kumiko Taguchi, and Tsuneo Kobayashi. 2017. "Alteration of Vascular Responsiveness to Uridine Adenosine Tetraphosphate in Aortas Isolated from Male Diabetic Otsuka Long-Evans Tokushima Fatty Rats: The Involvement of Prostanoids" International Journal of Molecular Sciences 18, no. 11: 2378. https://doi.org/10.3390/ijms18112378