Vitamin D Axis in Inflammatory Bowel Diseases: Role, Current Uses and Future Perspectives


1
Department of Life, Health and Environmental Sciences, Division of Internal Medicine and Nephrology, University of L’Aquila, 67100 L’Aquila, Italy
2
Division of Gastrointestinal and Liver Disease, Case Western Reserve University, Cleveland, OH 44106, USA
3
Digestive Health Institute, University Hospitals Cleveland Medical Center, Cleveland, OH 44106, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(11), 2360; https://doi.org/10.3390/ijms18112360
Submission received: 1 October 2017
/
Revised: 27 October 2017
/
Accepted: 29 October 2017
/
Published: 7 November 2017
(This article belongs to the Special Issue Vitamin D and Its Analogues: New Insights on Biological Effects and Therapeutic Uses)
Abstract
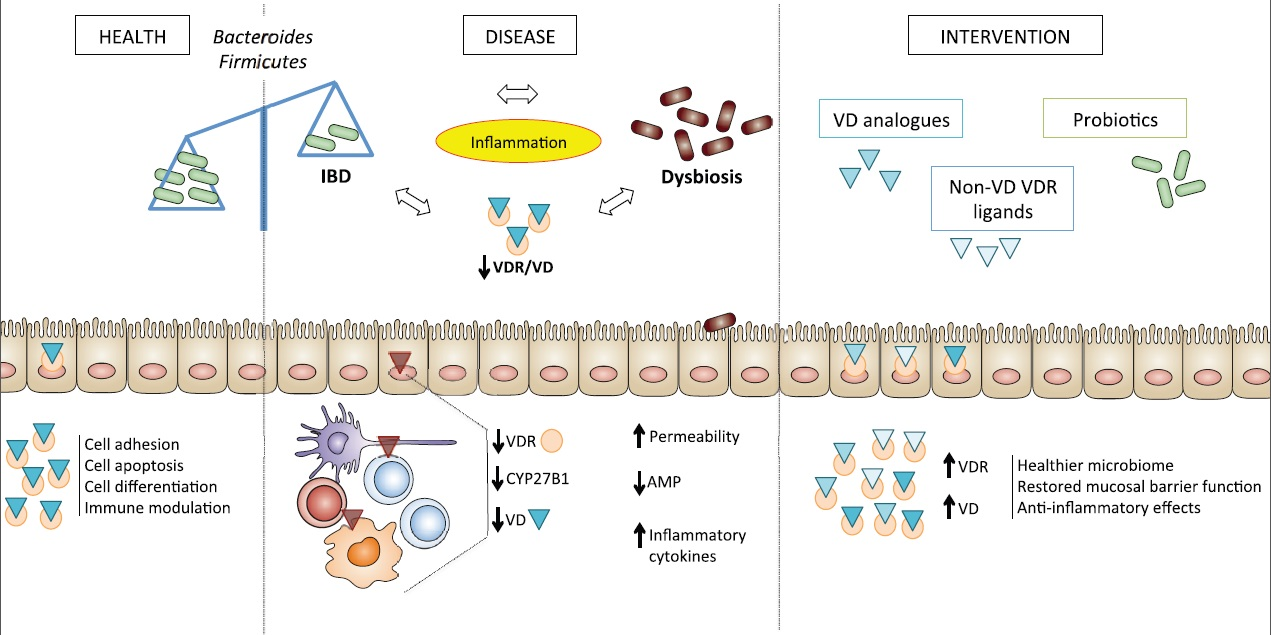
:Increasing evidence supports the concept that the vitamin D axis possesses immunoregulatory functions, with vitamin D receptor (VDR) status representing the major determinant of vitamin D’s pleiotropic effects. Vitamin D promotes the production of anti-microbial peptides, including β-defensins and cathelicidins, the shift towards Th2 immune responses, and regulates autophagy and epithelial barrier integrity. Impairment of vitamin D-mediated pathways are associated with chronic inflammatory conditions, including inflammatory bowel diseases (IBD). Interestingly, inhibition of vitamin D pathways results in dysbiosis of the gut microbiome, which has mechanistically been implicated in the development of IBD. Herein, we explore the role of the vitamin D axis in immune-mediated diseases, with particular emphasis on its interplay with the gut microbiome in the pathogenesis of IBD. The potential clinical implications and therapeutic relevance of this interaction will also be discussed, including optimizing VDR function, both with vitamin D analogues and probiotics, which may represent a complementary approach to current IBD treatments.

{kind=link}
1. Introduction
Vitamin D is a pleiotropic hormone of the steroid/thyroid superfamily, classically known for calcium homeostasis, but with several, additional non-calcemic effects, ranging from immune modulation to cell differentiation and intercellular adhesion. The active form of the vitamin is obtained after a two-step hydroxylation of the inactive precursor, cholecalciferol. Cholecalciferol undergoes hydroxylation at position 25, and the resultant inactive intermediate calcifediol [25(OH)D3] is further hydroxylated at position 1 to obtain the active vitamin, calcitriol [1,25(OH)2D3]. Both the inactive, as well as the active, forms of vitamin D circulate in the bloodstream bound to vitamin D-binding protein (VDBP). The active form exerts its effects by binding to a specific transcription-regulating molecule, the vitamin D receptor (VDR). Low levels of vitamin D have been associated with a variety of immune-mediated diseases, as well as with altered immune responses to pathogens and increased susceptibility to infection and cancer [1,2,3,4]. In addition, its binding protein has the ability to directly mediate some immunoregulatory functions [5], while its receptor, also expressed on immune cells, participates in modulation of inflammatory pathways [6].
Inflammatory bowel diseases (IBD) are chronic, inflammatory disorders that can affect the entire gastrointestinal tract, and are thought to result from inappropriate and ongoing immune activation in response to gut luminal agents in genetically predisposed individuals [7,8]. Vitamin D deficiency is common in the setting of the two major forms of IBD, Crohn’s disease (CD) and ulcerative colitis (UC) [9,10]. Interestingly, growing evidence supports the concept that intestinal dysbiosis and vitamin D metabolism are related in several ways, which may be of interest for uncovering novel pathogenic mechanisms and for future therapeutic directions in IBD [9,10].
In this context, we reviewed the current literature to summarize emerging evidence regarding the critical role of the vitamin D axis in the setting of IBD, with particular emphasis on the cross-talk between the gut microbiome and Vitamin D/VDR-mediated genetic and immune responses. Our results depict an interesting concept, wherein a balanced intervention on VDR function, both with vitamin D analogues and probiotics, may represent a complementary approach to IBD treatment.
2. Why Target the Vitamin D Axis in IBD?
Vitamin D, its binding protein and its receptor, constitute the so-called vitamin D axis, for which many interesting properties at the level of gut physiology have been emerging. The cellular actions of vitamin D are specifically mediated by the VDR, a ligand-dependent transcriptional regulator of the nuclear receptor superfamily, which is expressed in a variety of cell types, including mucosal immune cells and the intestinal epithelium. In addition, the enzyme, Cyp27B1, which converts circulating, inactive vitamin D [25(OH)D3] into its active, VDR-binding form [1,25(OH)2D3], is also expressed in different immune cell populations, as well as the intestinal epithelium. Co-localization of such key players in different cells of the gastrointestinal tract suggests the role of active vitamin D as a paracrine molecule, whose levels are modulated according to local needs [11,12,13]. Interestingly, intestinal bacteria have been shown to regulate the vitamin D axis within the gut, acting on the intestinal epithelium, as well as on local mucosal immune cells. In particular, the expression of Cyp27B1 has been reported to be reduced in intestinal epithelial cells of germ-free and antibiotic-treated mice, as is the expression of several genes involved in innate immunity (e.g., antibacterial peptides, tight junction proteins, cytokines and their receptors), suggesting that the synthesis of active vitamin D by the ‘‘microbiota-dependent’’ Cyp27B1 enzyme may be a requirement for the proper development of local innate immunity [14].
Similarly, probiotics and pathogenic bacteria have shown to modulate VDR expression in opposite directions, with the former increasing [15], and the latter decreasing [16], its expression. In particular, VDR is subject to the actions of antagonist molecules, in an attempt of pathogens to escape immune surveillance and manipulate host genes to increase their own survival [6,17]. The VDR gene (VDR, 12q12—14) is among the candidate genes that have been extensively studied for associations with IBD. Results from two recent meta-analyses showed that the risk of CD is increased in the presence of the VDR ApaI polymorphism and the TaqI tt genotype, whereas the risk of UC may decrease in the presence of the VDR TaqI polymorphism, especially in Caucasians [18,19]. For Asians, the VDR FokI polymorphism has been associated to susceptibility to UC [19]. In experimental animal models, VDR knockout (VDR KO) mice showed greater susceptibility to experimental colitis, manifested as worse histology scores, increased expression of genes encoding proinflammatory cytokines, and the development of intestinal dysbiosis [9,20,21]. The latter, in turn, was shown to dramatically alter the composition of bile acids in feces, and this may profoundly affect further molecular signaling, with particular focus on the cellular responses involved in immune regulation [22,23].
Vitamin D binding protein (VDBP), or Gc globulin (human group-specific component (Gc)), is a 55 kDa serum protein secreted by the liver and belonging to the albumin superfamily that is responsible for transporting active and inactive vitamin D in the plasma [24]. Single nucleotide polymorphisms (SNPs) in the gene encoding VDBP have been shown to affect circulating levels of this protein, as well as of circulating 25(OH)D3 [25]. VDBP is essential for the proper functioning of the endocytic pathway required for the renal uptake of 25(OH)D3 into renal tubular cells and consequent activation of the vitamin [26]. An association has been reported between specific SNPs in VDBP (VDBP 420 variant Lys; 416 Asp 420 Lys) and IBD, although their exact meaning in the pathogenesis of the disease remains to be determined [27]. VDBP has shown additional properties aside from a vitamin D carrier, particularly serving as a chemotactic and scavenger agent, as well as a macrophage activator. In fact, plasma VDBP effectively scavenges G-actin released at sites of necrotic cells and prevents polymerization of actin in the circulation [24]. In addition, it functions as a co-chemotactic factor for C5a, which is a very potent chemotactic factor for all leukocytes, as well as several other cell types, and is generated by limited proteolytic cleavage of C5 during complement activation [28]. After stepwise modification of its sugar moiety, VDBP is also converted into macrophage-derived macrophage activating factor (GcMAF), which not only produces a fully active ingestion function and cytotoxic capacity in 3 hours [29], but also has additional functions, such as antitumor [30,31,32] and antiangiogenic [33,34,35] activities. As a result, cloned GcMAF constructs [36] and GcMAF-mimicking peptides [37] have been developed for the purpose of studying their potential clinical use as immunopotentiators.
3. The Vitamin D Axis, Gut Microbiome, and the Gut Mucosal Immune System: Interplay at the Intestinal Level
Intestinal homeostasis is determined by the interplay among multiple factors, linked through complex molecular signaling, including the intestinal epithelial barrier, the gut microbiome, and components of the innate and adaptive immune systems. Interesting effects of the vitamin D axis on each of these components have been described.
3.1. Intestinal Epithelial Barrier
The differentiated intestinal epithelium constitutes a barrier for the free exchange of molecules between the intestinal lumen and the gut mucosa. In fact, the presence of adhesion structures between adjacent epithelial cells, namely tight junctions (occludin, proteins of the zonula occludens, and claudins), adherens junctions (E-cadherin, catenins, nectin [38]), desmosomes and gap junctions, guarantees the sealing of the paracellular space and regulates the permeability of the mucosal barrier. The integrity of the gut mucosa is also crucial for protection against microorganisms. Disruption of barrier function, in fact, facilitates infection with enteropathogenic bacteria and the development of intestinal inflammation [39] and IBD [40,41,42,43,44]. Conversely, probiotics have been shown to decrease paracellular permeability, evaluated by transepithelial electrical resistance (TEER), as well as to decrease epithelial apoptosis, in different models of intestinal inflammation [45,46,47]. Impaired mucosal barrier function with hyperpermeability is also common in the setting of several infectious and immune-mediated diseases of the lung (cystic fibrosis [48], interstitial lung disease [49], asthma [50,51], tuberculosis [48], chronic obstructive pulmonary disease [52]), skin (atopic dermatitis [53]), oral mucosa [54,55] and eyes [56], where impairment of the vitamin D axis has been described. In addition, intestinal epithelial cells cooperate with the hematopoietic compartments for the management of enteric infections and play an essential role in the initiation of type 2 immune responses [57,58]. Examples of epithelial-derived immunocompetent cells, include Paneth cells, goblet cells and the specialized phagocytic, antigen-presenting M cells located in the follicle-associated epithelium overlying organized lymphoid structures.
Vitamin D and its receptor have a protective effect on epithelial barriers in various tissues, including the gut mucosa [59,60,61]. In fact, it is well documented that active vitamin D increases the expression of several tight junction and adherent junction proteins [62]. In particular, active vitamin D induces the expression and/or membrane translocation of occludin, the zonula occludens proteins, ZO-1 and ZO-2, claudins 2, -7 and -12, and vinculin at several anatomic sites, including corneal epithelium, podocytes, and enterocytes [63,64]. In vitro studies demonstrated that pretreatment with 1,25(OH)2D3 protects intestinal epithelial cells from increased permeability induced by dextran sulfate sodium (DSS), and in vivo studies using VDR KO mice showed increased susceptibility to DSS-induced colitis when compared to their wild-type littermates [65]. Moreover, it has been demonstrated that intestinal epithelial VDR signaling plays a key role in maintaining mucosal barrier integrity by suppressing intestinal epithelial cell apoptosis, thus regulating gut mucosal inflammation [59,66]. In addition to their sealing properties, adherent proteins are actively involved in signal transduction, and VDR can regulate such pathways acting on VDR-regulated promoters. For instance, active vitamin D attenuates growth and promotes differentiation in colon cancer cells by the VDR-mediated induction of E-cadherin and inhibition of β-catenin signaling [67,68,69]. Taken together, these data confirm the role of the vitamin D axis in mucosal barrier development, integrity and healing capacity.
3.2. Intestinal Microbiome
In recent years, the Human Microbiome Project has provided unprecedented information regarding the diversity and function of microbial communities and their genes, referred to as the human microbiome [70]. Sequencing of microbial ribosomal RNA obtained from different body sites showed that the number and relative distribution of distinct microbial species characterized health and disease states in humans; for instance, decreased diversity in the gut was observed in IBD [71]. The intestinal microbiome has a role in several functions, including metabolism, mucosal barrier physiology, immunity, and inflammatory signaling, and its disruption, or dysbiosis, is associated with the development, maintenance, and perpetuation of various clinical conditions, both intestinal and extraintestinal. By regulating the expression of antimicrobial peptides [72,73], mucosal barrier function, and innate immunity [61], vitamin D and its receptor have been shown to influence the composition and functions of bacterial communities in the gut, protect from dysbiosis, as well as experimental IBD and its symptoms [9,74].
Although there appears to be no unique and optimal composition of the microbiome to promote gut health, Bacteroides and Firmicutes species are the most highly represented under normal conditions [70]. During IBD, a reduction in the number of species within the phylum Firmicutes—specifically the Clostridium clusters XIVa and IV- and Bacteroidetes-namely Bifidobacterium, Lactobacillus, and Ruminococcaceae (particularly the butyrate-producing genus Faecalibacterum)—and an increase in Bacillus spp and Enterobacteriacae, is observed [75,76,77]. Studies using VDR KO mice and wild-type littermates showed defective autophagy and consequent gut dysbiosis in the former, with depletion of fecal Lactobacillus and Bacteroides [78]. In a colitis model, administration of butyrate, a fermentation product of gut bacteria, increased intestinal VDR expression and suppressed inflammation [78]. Interaction between the vitamin D axis and the gut microbiome was further demonstrated in a model of experimental colitis on CYP27b1 KO and VDR KO mice compared to littermates [9]. Results of this study showed greater susceptibility of KO mice to DSS colitis, which was associated with bacterial imbalance, with more Proteobacteria and less Firmicutes, similarly to that observed in patients with IBD. Vitamin D deficiency itself was also shown to be a co-factor for dysbiosis in the setting of a high-fat dietary regimen, and this effect was mediated by the downregulation of specific α-defensins from ileal Paneth cells, as well as of tight junction genes in the absence of vitamin D, with consequent endotoxemia and systemic inflammation [79]. Pathogens may also regulate the monocyte/macrophage vitamin D axis in their own favor through DNA methylation on specific sequences, namely micro-RNAs (miRs) [13]. As an example, miR-21 can interact with CYP27B1 mRNA and suppress its activity, thus decreasing localized synthesis of active vitamin D in monocytes [80].
Probiotics, consisting of ingestible non-pathogenic living microorganisms with the ability to confer some beneficial effects to the host when consumed in adequate amounts as food components [81], have been widely used in clinical trials for the treatment of IBD with variable results [82,83]. It has been recently shown that a properly functioning VDR pathway is required for probiotic protection against colitis [84], a finding that is of importance since VDR expression can be significantly decreased in IBD patients as a consequence of chronic inflammation [85] or dysbiosis [6,17]. VDR KO mice, in fact, did not respond to probiotics such as Lactobacillus rhamnosus strain GG (LGG) and Lactobacillus plantarum (LP) and had worse severity of Salmonella-induced colitis compared to littermates [84]. The same probiotics in wild-type mice, indeed, were able to increase VDR expression and its transcriptional activity, with increased expression of antimicrobial peptides, and had the ability to confer physiological and histologic protection from Salmonella-induced colitis [84]. Taken together, the interplay of the vitamin D axis with the intestinal microbiome is an intriguing, yet undiscovered field of research, with potential clinical implications.
3.3. Immune System
Besides its role as a site for nutrient absorption, the gut also hosts a unique immune system, composed of coordinated immunocompetent cells that cooperate in the difficult, unparalleled task of discriminating between harmful and beneficial antigens, among the plethora of diverse antigenic components of its intraluminal content [86]. Such ability is crucial for the induction of tolerance towards nutrients and commensal bacteria, as well as for first-line protection against pathogens. Apart from epithelial-derived immunocompetent cells, the intestinal immune system consists of organized lymphoid structures, such as Peyer’s patches (PPs), cryptopatches and isolated lymphoid follicles (ILFs), which are located immediately below the epithelial layer within the lamina propria. Within the gut mucosa reside several immune populations, including intraepithelial effector lymphocytes interspersed within the epithelial lining, polarized CD4+ T cells, such as T regulatory cells (Treg), T helper 1, 2 and 17 (Th1, Th2, Th17) cells, and IgA-producing plasma cells, as well as innate immune cells with antigen-presenting cell function, such as dendritic cells (DCs) and monocytes/macrophages, and the recently identified heterogeneous group of innate lymphoid cells (ILCs) [87].
The vitamin D axis is an important regulator of the innate and adaptive immune systems. Its effects include decreasing Th1/Th17 T cells and pro-inflammatory cytokines, such as IL-1, IL-6, IL-8, IFNγ, and TNFα, in favor of Th2 response, increasing Tregs, downregulating T cell-driven IgG production, inhibiting DC differentiation, and helping maintain self-tolerance, while enhancing protective innate immune responses [88]. In particular, active vitamin D [89] and glycosylated VDBP [29] boost autophagy in human monocytes/macrophages, with inhibitory effects on intracellular pathogens, including Mycobacterium tuberculosis and human immunodeficiency virus type 1 [90]. By boosting C5a activity, VDBP enhances cell recruitment during inflammation, with particular reference to neutrophils, monocytes and fibroblasts [28,91]. VDBP also modulates the availability of 25(OH)D3 to DCs, indirectly regulating the amount of active vitamin derived from DCs that can become available to T cells [92]. Of importance in IBD, VDR mediates enhanced production of antimicrobial peptides, such as β-defensin 2 (DEFB4/HBD2) and cathelicidin (CAMP) that are traditionally boosted by the activation of nucleotide-binding oligomerization domain-containing protein 2 (NOD2) after stimulation by microbial muramyl dipeptides [73]. Interesting, a vitamin D deficient diet can repress the expression of defensins and their activating enzyme, matrix metalloproteinase 7 (MMP7), with consequent dysbiosis [79]. Notably, the region encoding DEFB2/HBD2 has been identified as a CD susceptibility locus. The NOD2 gene itself has a VDRE [93,94], and mutations or dysregulation in NOD2, with consequent decreased expression of antimicrobial peptides, impaired autophagy, and dysbiosis, are also associated with IBD [93,95,96]. In addition, NOD2 KO-associated altered microbial composition, with greater susceptibility to DSS colitis, was transmissible to co-housed wild-type mice [96]. Another mechanism for vitamin D to activate innate immunity occurs by enhancing the function of another pattern recognition receptor, the Toll-like receptor 4 (TLR4), which is traditionally activated after recognition of lipopolysaccharide (LPS), a cell wall component of gram-negative bacteria [97,98]. Interestingly, VDR levels were found to be reduced by more than 50%, and the pro-inflammatory cytokines, TNFα and IL-1β, were elevated, in colonic biopsies from patients with CD and UC, indicating that VDR can be repressed by inflammatory mediators [59]. Studies on wild-type, specific pathogen-free IL-10 KO mice, and VDR KO mice showed impaired T cell homing to the gut in the absence of vitamin D signaling, with less CD8+ intraepithelial lymphocytes, low levels of IL-10 and consequent increased inflammatory response to the normally harmless commensal flora [99]. Stimulation of VDR by microbial-derived bile acids is another pathway of immune modulation mediated by the vitamin D axis, of which changes in bile acid profiles after dysbiosis can impact such signaling [23].
4. Role of the Vitamin D Axis and Dysbiosis in Immune-Mediated Diseases: Focus on Experimental Studies Targeting the Vitamin D Axis in IBD
Autoimmune diseases are self-directed pathologies resulting from an aberrant activation of the immune system against harmless self-antigens, with consequent inflammation and tissue damage compromising the affected target organs or systems. An association has been consistently described between autoimmunity and vitamin D epidemiology and genetics, thus promoting great interest in the potential clinical applications of targeting the vitamin D axis in such conditions, including IBD. Considering the interplay between the vitamin D axis, the gut microbiome and the mucosal immune system, the reciprocal effects of targeted interventions on each of these components represents an intriguing therapeutic opportunity.
Human clinical trials targeting the vitamin D axis in IBD are often heterogeneous in their design and methods, and this prevents the exact comparability of results. However, despite differences in disease history and treatment, as well as additional environmental factors that can influence vitamin D status, most studies are in agreement regarding the beneficial effects of vitamin D supplementation on disease activity and/or quality of life [100,101,102,103,104,105,106]. As an example, a randomized, double-blind placebo-controlled study on 94 CD patients with inactive disease, assigned to either 1200 IU vitamin D3 daily or placebo for 12 months, showed that the relapse rate had a trend towards being lower in the treatment versus placebo group (p = 0.06) [100]. Similar results were described after a trial of high-dose vitamin D3 at 10,000 IU daily (n = 18) compared to 1000 IU daily (n = 16) for 12 months in patients with CD in remission, with less clinical relapse of disease in patients receiving a higher dose of vitamin D (0% vs. 37.5%, p = 0.049) [105]. Similarly, a recent prospective randomized controlled trial on 18 patients with UC and hypovitaminosis D showed that vitamin D3 supplementation improved quality of life and reduced UC disease activity, especially at higher doses (4000 IU daily versus 2000 IU daily) [103]. Other studies have reported a short-term beneficial effect on disease activity in CD patients treated with vitamin D, particularly in its active form [101]. A recent pilot study on CD and UC patients assigned to oral vitamin D supplementation targeting a serum concentration of 100–125 nmol/L showed successful and safe improvement of symptom-based activity scores, but did not show significant changes in objective measures of intestinal or systemic inflammation after 12 weeks [104]. In addition, a randomized placebo-controlled clinical trial on 108 IBD patients with vitamin D deficiency (serum 26(OH)D < 30 ng/mL) showed that oral supplementation with cholecalciferol 50,000 UI/week was not significantly efficacious in reducing serum TNFα levels after 12 weeks (p = 0.07) [106]. In patients with multiple sclerosis, daily vitamin D supplementation (5000 IU for 3 months) increased the abundance of Akkermansia, which promotes immune tolerance, as well as Faecalibacterium and Coprococcus, which produce the VDR-activating, anti-inflammatory fermentation product butyrate [107]. In a pilot study, the gut microbiome was modified and intraepithelial CD8+ T-cells in the terminal ileum increased, even in healthy volunteers after 2-months of vitamin D supplementation [108].
Given the growing evidence supporting the intimate relationship between the gut microbiome and the vitamin D axis in autoimmunity, potential contributing factors may help to explain the observed results. For instance, bacterial-induced epigenetic modifications in cytochromes involved in vitamin D metabolism, as well as changes in bile acid profiles after dysbiosis, can influence VDR-mediated signaling. Use of probiotics, indeed, promotes VDR expression and its antimicrobial effects, which is beneficial to dampening colonic inflammation. In turn, vitamin D may restore a healthier gut microbiome and attenuate inflammation. Therefore, a correct nutraceutical approach to immune-mediated diseases, including IBD, should contemporarily exert beneficial effects on both VDR expression and signaling, and the gut microbiome.
5. Future Directions
The role of vitamin D in immune-mediated diseases appears to be intimately associated with bacteria metabolism, with chronic dysbiosis causing VDR dysfunction and triggering a vicious cycle, wherein a compromised immune system perpetuates disease. Restoring VDR function at different cellular levels should therefore be considered as a therapeutic option. Probiotics and olmesartan proved to be effective in this sense in experimental settings, but require additional testing in human studies in specific clinical settings [15,16,84,109]. To date, in fact, sparse research has been performed on the reciprocal effects of probiotics and vitamin D in humans. An example is a double-blind, placebo-controlled, randomized trial on 127 otherwise healthy hypercholesterolemic adults randomized to consume L. reuteri NCIMB 30242 or placebo capsules over a 9 week intervention period. The study showed a significant increase in circulating vitamin D in response to oral probiotic supplementation compared to placebo (p = 0.003) [110]. Olmesartan is an angiotensin receptor blocker with VDR-binding properties. According to some evidence, it acts as a VDR agonist and is able to restore proper VDR function by displacing bacterial products bound to the receptor with inhibitory effects [16]. Olmesartan was proposed in combination with pulsed, low dose, broad-spectrum, bacteriostatic antibiotics as an approach to reverse the disease process in autoimmune diseases [16]. Raising epithelial VDR levels by vitamin D analogues or by anti-TNF therapy may represent an additional mechanism to ameliorate IBD by reducing IEC apoptosis [59]. In addition, several VDR ligands with low calcemic effects, but high therapeutic potential have drawn attention as possible alternatives to active vitamin D [111]. In fact, despite major side effects of vitamin D supplementation, like hypercalcemia, have been rarely reported and are usually only observed after exposure to high doses of the active hormone, the risk of vascular calcifications, hypercalciuria and renal complications following long-term exposure to vitamin D remains uncertain [112]. Non-vitamin D VDR ligands demonstrated in vivo efficacy in protecting against, or reducing the severity of, experimental colitis [113,114,115,116]. Similarly, in vitro studies on human immune cells have proven the ability of these analogues to modulate the immune system through the switch of Th1 into Th2 immune response and the down-regulation of pro-inflammatory cytokines in peripheral mononuclear cells [117,118,119,120]. Glucuronide conjugates of vitamin D represent an additional option for targeted delivery of active vitamin D at specific sites of the gastrointestinal tract, namely the ileum and colon, a mechanism that exploits bacteria metabolism. In fact, bacteria residing in the lower gastrointestinal tract produce β-glucuronidase enzymes that can cleave glucuronide and liberate 1,25(OH)2D3 for local actions [121,122]. Duodenal bacteria did not appear to produce the same enzyme [122]. Therefore, the oral administration of such water-soluble vitamin D compounds allows their selective action at the lower intestinal tract, which is typically affected by IBD, without hypercalcemic effects, and theoretically improves the efficacy of rectally-administered vitamin D, whose diffusion would exclude the ileum. In accordance with this, βGluc-1,25(OH)2D3 proved to ameliorate the severity of experimental IBD in mice, without rising blood concentrations of calcium [122]. Finally, in the era of personalized medicine, a better knowledge of targeting gene expression, or anticipating the potential response to treatment based on genetic variants of specific genes [123], may further help improving quality of life in chronic diseases.
In conclusion, the horizon opened by current advances in knowledge in the field of microbiomics and nutraceuticals depicts interesting implications in the treatment of immune-mediated diseases. Despite persistent gaps preventing the possibility of recommendations to incorporate manipulation of the vitamin D axis and microbiome into clinical practice guidelines, results of recent research encourage the pursuit of this goal for better, targeted therapy for the treatment of patients with IBD.
Acknowledgments
The Authors wish to thank Theresa Pizarro for critical revision of the manuscript. No specific source of funding was used for this study.
Author Contributions
Rita Del Pinto and Fabio Cominelli conceived the manuscript; Rita Del Pinto performed the literature search; Rita Del Pinto and Claudio Ferri wrote the first draft and Fabio Cominelli supervised the final version of the paper. All Authors have read and approved the final version of the manuscript.
Conflicts of Interest
The Authors declare no conflict of interest.
Abbreviations
| VDBP | Vitamin D binding protein |
| VDR | Vitamin D receptor |
| IBD | Inflammatory bowel diseases |
| CD | Crohn’s disease |
| UC | Ulcerative colitis |
| KO | Knockout |
| SNPs | Single nucleotide polymorphisms |
| GcMAF | Group-specific component macrophage activating factor |
| TEER | Transepithelial electrical resistance |
| IECs | Intestinal epithelial cells |
| DSS | Dextran sodium sulfate |
| RNA | Ribonucleic acid |
| DNA | Deoxyribonucleic acid |
| PPs | Peyer’s patches |
| ILFs | Isolated lymphoid follicles |
| Treg | Regulatory T cells |
| Th | T helper |
| DCs | Dendritic cells |
| ILCs | Isolated lymphatic cells |
| IL | Interleukin |
| IFN | Interferon |
| TNF | Tumor necrosis factor |
| HBD | Human beta defensin |
| CAMP | Cathelicidin antimicrobial peptide |
| NOD2 | Nucleotide-binding oligomerization domain-containing protein 2 |
| MMP | Matrix metalloproteinase |
| TLR | Toll like receptor |
| LPS | Lipopolysaccharide |
| IU | International unit |
| OR | Odds ratio |
| CI | Confidence interval |
References
- Holick, M.F. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Dankers, W.; Colin, E.M.; van Hamburg, J.P.; Lubberts, E. Vitamin D in Autoimmunity: Molecular Mechanisms and Therapeutic Potential. Front. Immunol. 2016, 7, 697. [Google Scholar] [CrossRef] [PubMed]
- Gois, P.H.F.; Ferreira, D.; Olenski, S.; Seguro, A.C. Vitamin D and infectious diseases: Simple bystander or contributing factor? Nutrients 2017, 9, 651. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Homma, S. Vitamin D3 binding protein (group-specific component) is a precursor for the macrophage-activating signal factor from lysophosphatidylcholine-treated lymphocytes. Proc. Natl. Acad. Sci. USA 1991, 88, 8539–8543. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Cantorna, M.T. Vitamin D and the immune system. In Diet, Immunity and Inflammation; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 244–263. [Google Scholar]
- Del Pinto, R.; Pietropaoli, D.; Chandar, A.K.; Ferri, C.; Cominelli, F. Association between inflammatory bowel disease and vitamin D deficiency: A systematic review and meta-analysis. Inflamm. Bowel Dis. 2015, 21, 2708–2717. [Google Scholar] [CrossRef] [PubMed]
- Basson, A.; Trotter, A.; Rodriguez-Palacios, A.; Cominelli, F. Mucosal Interactions between Genetics, Diet, and Microbiome in Inflammatory Bowel Disease. Front. Immunol. 2016, 7, 290. [Google Scholar] [CrossRef] [PubMed]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef] [PubMed]
- Proal, A.D.; Albert, P.J.; Marshall, T. Autoimmune disease in the era of the metagenome. Autoimmun. Rev. 2009, 8, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T. Why do T cells express the vitamin D receptor? Ann. N. Y. Acad. Sci. 2011, 1217, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Kongsbak, M.; Levring, T.B.; Geisler, C.; von Essen, M.R. The vitamin d receptor and T cell function. Front. Immunol. 2013, 4, 148. [Google Scholar] [CrossRef] [PubMed]
- Chun, R.F.; Liu, P.T.; Modlin, R.L.; Adams, J.S.; Hewison, M. Impact of vitamin D on immune function: Lessons learned from genome-wide analysis. Front. Physiol. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, A.; Kobiita, A.; Ye, T.; Chambon, P. Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by TLRs. Cell 2013, 153, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Appleyard, C.B.; Cruz, M.L.; Isidro, A.A.; Arthur, J.C.; Jobin, C.; de Simone, C. Pretreatment with the probiotic VSL#3 delays transition from inflammation to dysplasia in a rat model of colitis-associated cancer. AJP Gastrointest. Liver Physiol. 2011, 301, G1004–G1013. [Google Scholar]
- Waterhouse, J.C.; Perez, T.H.; Albert, P.J. Reversing bacteria-induced vitamin D receptor dysfunction is key to autoimmune disease. Ann. N. Y. Acad. Sci. 2009, 1173, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Proal, A.D.; Albert, P.J.; Marshall, T.G. The human microbiome and autoimmunity. Curr. Opin. Rheumatol. 2013, 25, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.T.; Hu, J.J.; Fan, R.; Zhou, J.; Zhong, J. Polymorphisms of the vitamin D receptor gene and the risk of inflammatory bowel disease: A meta-analysis. Genet. Mol. Res. 2014, 13, 2598–2610. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.-N.; Xu, K.-Q.; Zhang, W.; Wang, Q.; Wu, J.; Wang, X.-Y. Associations between vitamin D receptor polymorphisms and susceptibility to ulcerative colitis and Crohn’s disease: A meta-analysis. Inflamm. Bowel Dis. 2013, 19, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Yamaori, S.; Tanabe, T.; Johnson, C.H.; Krausz, K.W.; Kato, S.; Gonzalez, F.J. Implication of intestinal VDR deficiency in inflammatory bowel disease. Biochim. Biophys. Acta 2013, 1830, 2118–2128. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Waddell, A.; Lin, Y.-D.; Cantorna, M.T. Dysbiosis caused by vitamin D receptor deficiency confers colonization resistance to Citrobacter rodentium through modulation of innate lymphoid cells. Mucosal Immunol. 2015, 8, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Ko, J.I.; Kim, W.B.; Kang, H.K.; Kim, B.K. Taurine can ameliorate inflammatory bowel disease in rats. Adv. Exp. Med. Biol. 1998, 442, 291–298. [Google Scholar] [PubMed]
- Joyce, S.A.; Gahan, C.G.M. Disease-Associated Changes in Bile Acid Profiles and Links to Altered Gut Microbiota. Dig. Dis. 2017, 35, 169–177. [Google Scholar] [CrossRef] [PubMed]
- White, P.; Cooke, N. The multifunctional properties and characteristics of vitamin D-binding protein. Trends Endocrinol. Metab. 2000, 11, 320–327. [Google Scholar] [CrossRef]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Streeten, C.; Koller, D.L. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef]
- Nykjaer, A.; Dragun, D.; Walther, D.; Vorum, H.; Jacobsen, C.; Herz, J.; Melsen, F.; Christensen, E.I.; Willnow, T.E. An endocytic pathway essential for renal uptake and activation of the steroid 25-(OH) vitamin D3. Cell 1999, 96, 507–515. [Google Scholar] [CrossRef]
- Eloranta, J.J.; Wenger, C.; Mwinyi, J.; Hiller, C.; Gubler, C.; Vavricka, S.R.; Fried, M.; Kullak-Ublick, G.A. Association of a common vitamin D-binding protein polymorphism with inflammatory bowel disease. Pharmacogenet. Genom. 2011, 21, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kew, R.R. Identification of a region in the vitamin D-binding protein that mediates its C5a chemotactic cofactor function. J. Biol. Chem. 2004, 279, 53282–53287. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, H.; Uto, Y.; Sasaki, H.; Okamura, N.; Murakami, A.; Kubo, S.; Kirk, K.L.; Hori, H. Gc protein (vitamin D-binding protein): Gc genotyping and GcMAF precursor activity. Anticancer Res. 2005, 25, 3689–3695. [Google Scholar] [PubMed]
- Yamamoto, N.; Naraparaju, V.R. Immunotherapy of BALB/c mice bearing Ehrlich ascites tumor with vitamin D-binding protein-derived macrophage activating factor. Cancer Res. 1997, 57, 2187–2192. [Google Scholar] [PubMed]
- Korbelik, M.; Naraparaju, V.R.; Yamamoto, N. Macrophage-directed immunotherapy as adjuvant to photodynamic therapy of cancer. Br. J. Cancer 1997, 75, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Koga, Y.; Naraparaju, V.R.; Yamamoto, N. Antitumor effect of vitamin D-binding protein-derived macrophage activating factor on Ehrlich ascites tumor-bearing mice. Proc. Soc. Exp. Biol. Med. 1999, 220, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Kanda, S.; Mochizuki, Y.; Miyata, Y.; Kanetake, H.; Yamamoto, N. Effects of vitamin D3-binding protein-derived macrophage activating factor (GcMAF) on angiogenesis. J. Natl. Cancer Inst. 2002, 94, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Kisker, O.; Onizuka, S.; Becker, C.M.; Fannon, M.; Flynn, E.; D’Amato, R.; Zetter, B.; Folkman, J.; Ray, R.; Swamy, N. Vitamin D binding protein-macrophage activating factor (DBP-maf) inhibits angiogenesis and tumor growth in mice. Neoplasia 2003, 5, 32–40. [Google Scholar] [CrossRef]
- Onizuka, S.; Kawakami, S.; Taniguchi, K.; Fujioka, H.; Miyashita, K. Pancreatic Carcinogenesis. Pancreas 2004, 28, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Naraparaju, V.R.; Urade, M. Prognostic utility of serum alpha-N-acetylgalactosaminidase and immunosuppression resulted from deglycosylation of serum Gc protein in oral cancer patients. Cancer Res. 1997, 57, 295–299. [Google Scholar] [PubMed]
- Schneider, G.B.; Grecco, K.J.; Safadi, F.F.; Popoff, S.N. The Anabolic Effects of Vitamin D-Binding Protein-Macrophage Activating Factor (DBP-MAF) and a Novel Small Peptide on Bone. Crit. Rev. Eukaryot. Gene Expr. 2003, 13, 8. [Google Scholar] [CrossRef]
- Zhang, Y.-G.; Wu, S.; Sun, J. Vitamin D, Vitamin D Receptor, and Tissue Barriers. Tissue Barriers 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.R.; Dahlhoff, M.; Horst, D.; Hirschi, B.; Trülzsch, K.; Müller-Höcker, J.; Vogelmann, R.; Allgauer, M.; Gerhard, M.; Steininger, S.; et al. A key role for E-cadherin in intestinal homeostasis and Paneth cell maturation. PLoS ONE 2010, 5, e14325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollander, D. Crohn’s disease—A permeability disorder of the tight junction? Gut 1988, 29, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Soderholm, J.D. Augmented increase in tight junction permeability by luminal stimuli in the non-inflamed ileum of Crohn’s disease. Gut 2002, 50, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.R. Increased gut permeability in Crohn’s disease: Is TNF the link? Gut 2004, 53, 1724–1725. [Google Scholar] [CrossRef] [PubMed]
- Schulzke, J.D.; Ploeger, S.; Amasheh, M.; Fromm, A.; Zeissig, S.; Troeger, H.; Richter, J.; Bojarski, C.; Schumann, M.; Fromm, M. Epithelial tight junctions in intestinal inflammation. Ann. N. Y. Acad. Sci. 2009, 1165, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, J.; Vogelsang, H.; Hübl, W.; Waldhöer, T.; Lochs, H. Intestinal permeability and the prediction of relapse in Crohn’s disease. Lancet 1993, 341, 1437–1439. [Google Scholar] [CrossRef]
- Mennigen, R.; Nolte, K.; Rijcken, E.; Utech, M.; Loeffler, B.; Senninger, N.; Bruewer, M. Probiotic mixture VSL#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1140–G1149. [Google Scholar] [PubMed]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; van Langer, M.L.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Corridoni, D.; Pastorelli, L.; Mattioli, B.; Locovei, S.; Ishikawa, D.; Arseneau, K.O.; Chieppa, M.; Cominelli, F.; Pizarro, T.T. Probiotic bacteria regulate intestinal epithelial permeability in experimental ileitis by a TNF-dependent mechanism. PLoS ONE 2012, 7, e42067. [Google Scholar] [CrossRef] [PubMed]
- Finklea, J.D.; Grossmann, R.E.; Tangpricha, V. Vitamin D and chronic lung disease: A review of molecular mechanisms and clinical studies. Adv. Nutr. 2011, 2, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Zosky, G.R.; Berry, L.J.; Elliot, J.G.; James, A.L.; Gorman, S.; Hart, P.H. Vitamin D deficiency causes deficits in lung function and alters lung structure. Am. J. Respir. Crit. Care Med. 2011, 183, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, T.; Gupta, G.K.; Agrawal, D.K. Vitamin D deficiency decreases the expression of VDR and prohibitin in the lungs of mice with allergic airway inflammation. Exp. Mol. Pathol. 2012, 93, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Mak, G.; Hanania, N.A. Vitamin D and asthma. Curr. Opin. Pulm. Med. 2011, 17, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sundar, I.K.; Hwang, J.-W.; Wu, S.; Sun, J.; Rahman, I. Deletion of vitamin D receptor leads to premature emphysema/COPD by increased matrix metalloproteinases and lymphoid aggregates formation. Biochem. Biophys. Res. Commun. 2011, 406, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, B.; Riedel, R.; Jörss, K.; Loddenkemper, C.; Steinmeyer, A.; Zügel, U.; Babina, M.; Radbruch, A.; Worm, M. Vitamin D receptor activation improves allergen-triggered eczema in mice. J. Invest. Dermatol. 2012, 132, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, S.N.; Hagner, M.; Nogueira, A.V.B.; Franke, A.; Jäger, A.; Deschner, J. Inflammatory bowel disease and oral health: Systematic review and a meta-analysis. J. Clin. Periodontol. 2017, 44, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Pietropaoli, D.; del Pinto, R.; Corridoni, D.; Rodriguez-Palacios, A.; di Stefano, G.; Monaco, A.; Weinberg, A.; Cominelli, F. Occurrence of spontaneous periodontal disease in the SAMP1/YitFc murine model of Crohn disease. J. Periodontol. 2014, 85, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Pintea, V.; Lin, Y.; Hammock, B.D.; Watsky, M.A. Vitamin D enhances corneal epithelial barrier function. Invest. Ophthalmol. Vis. Sci. 2011, 52, 7359–7364. [Google Scholar] [CrossRef] [PubMed]
- Gerbe, F.; Jay, P. Intestinal tuft cells: Epithelial sentinels linking luminal cues to the immune system. Mucosal Immunol. 2016, 9, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Ivanov, I.I. Intestinal epithelial cells as mediators of the commensal–host immune crosstalk. Immunol. Cell Biol. 2013, 91, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Chen, Y.; Du, J. Critical roles of intestinal epithelial vitamin D receptor signaling in controlling gut mucosal inflammation. J. Steroid Biochem. Mol. Biol. 2015, 148, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.-Y.; Liu, T.-J.; Fu, J.-H.; Xu, W.; Wu, L.-L.; Hou, A.-N.; Xue, X.-D. Vitamin D/VDR signaling attenuates lipopolysaccharide‑induced acute lung injury by maintaining the integrity of the pulmonary epithelial barrier. Mol. Med. Rep. 2016, 13, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Barbáchano, A.; Fernández-Barral, A.; Ferrer-Mayorga, G.; Costales-Carrera, A.; Larriba, M.J.; Muñoz, A. The endocrine vitamin D system in the gut. Mol. Cell. Endocrinol. 2017, 453, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Stio, M.; Retico, L.; Annese, V.; Bonanomi, A.G. Vitamin D regulates the tight-junction protein expression in active ulcerative colitis. Scand. J. Gastroenterol. 2016, 51, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Migliori, M.; Giovannini, L.; Panichi, V.; Filippi, C.; Taccola, D.; Origlia, N.; Mannari, C.; Camussi, G. Treatment with 1,25-dihydroxyvitamin D3 preserves glomerular slit diaphragm-associated protein expression in experimental glomerulonephritis. Int. J. Immunopathol. Pharmacol. 2005, 18, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, Y.; Golan, M.A.; Annunziata, M.L.; Du, J.; Dougherty, U.; Kong, J.; Musch, M.; Huang, Y.; Pekow, J.; et al. Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis. J. Clin. Investig. 2013, 123, 3983–3996. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Islam, M.N.; Dakshanamurthy, S.; Rizvi, I.; Rao, M.; Herrell, R.; Zinser, G.; Valrance, M.; Aranda, A.; Moras, D.; et al. The molecular basis of vitamin D receptor and β-catenin crossregulation. Mol. Cell 2006, 21, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. WNT signaling pathway and stem cell signaling network. Clin. Cancer Res. 2007, 13, 4042–4045. [Google Scholar] [CrossRef] [PubMed]
- Pendás-Franco, N.; Aguilera, O.; Pereira, F.; González-Sancho, J.M.; Muñoz, A. Vitamin D and Wnt/β-catenin pathway in colon cancer: Role and regulation of DICKKOPF genes. Anticancer Res. 2008, 28, 2613–2623. [Google Scholar] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-T.; Dabbas, B.; Laperriere, D.; Bitton, A.J.; Soualhine, H.; Tavera-Mendoza, L.E.; Dionne, S.; Servant, M.J.; Bitton, A.; Seidman, E.G.; et al. Direct and indirect induction by 1,25-dihydroxyvitamin D3 of the NOD2/CARD15-defensin β2 innate immune pathway defective in Crohn disease. J. Biol. Chem. 2010, 285, 2227–2231. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Wu, S.; Zhang, Y.-G.; Lu, R.; Xia, Y.; Dong, H.; Sun, J. Lack of vitamin D receptor causes dysbiosis and changes the functions of the murine intestinal microbiome. Clin. Ther. 2015, 37, 996–1009.e7. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Seksik, P.; Rigottier-Gois, L.; Lay, C.; Lepage, P.; Podglajen, I.; Marteau, P.; Dore, J. Specificities of the fecal microbiota in inflammatory bowel disease. Inflamm. Bowel Dis. 2006, 12, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.N.; St. Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; Leleiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Zhang, Y.-G.; Lu, R.; Xia, Y.; Zhou, D.; Petrof, E.O.; Claud, E.C.; Chen, D.; Chang, E.B.; Carmeliet, G.; et al. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut 2015, 64, 1082–1094. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Nie, Y.; Zhu, A.; Chen, Z.; Wu, P.; Zhang, L.; Luo, M.; Sun, Q.; Cai, L.; Lai, Y.; et al. Vitamin D signaling through induction of Paneth cell defensins maintains gut microbiota and improves metabolic disorders and hepatic steatosis in animal models. Front. Physiol. 2016, 7, 498. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Wheelwright, M.; Teles, R.; Komisopoulou, E.; Edfeldt, K.; Ferguson, B.; Mehta, M.D.; Vazirnia, A.; Rea, T.H.; Sarno, E.N.; et al. MicroRNA-21 targets the vitamin D-dependent antimicrobial pathway in leprosy. Nat. Med. 2012, 18, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria (2001, Córdoba, Río Primero). In Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria: Report of a Joint FAO WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria, American Córdoba Park Hotel, Córdoba, Argentina, 1–4 October 2001; FAO: Rome, Italy, 2001.
- Derwa, Y.; Gracie, D.J.; Hamlin, P.J.; Ford, A.C. Systematic review with meta-analysis: The efficacy of probiotics in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2017, 46, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yoon, S.; Zhang, Y.-G.; Lu, R.; Xia, Y.; Wan, J.; Petrof, E.O.; Claud, E.C.; Chen, D.; Sun, J. Vitamin D receptor pathway is required for probiotic protection in colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G341–G349. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Tanaka, H.; Maeda, K.; Inoue, T.; Noda, E.; Amano, R.; Kubo, N.; Muguruma, K.; Yamada, N.; Yashiro, M.; et al. Vitamin D receptor expression is associated with colon cancer in ulcerative colitis. Oncol. Rep. 2009, 22, 1021–1025. [Google Scholar] [PubMed]
- Ahluwalia, B.; Magnusson, M.K.; Öhman, L. Mucosal immune system of the gastrointestinal tract: Maintaining balance between the good and the bad. Scand. J. Gastroenterol. 2017, 52, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, E.; Bedoui, S. Intestinal innate immune cells in gut homeostasis and immunosurveillance. Immunol. Cell Biol. 2013, 91, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Kamen, D.L.; Tangpricha, V. Vitamin D and molecular actions on the immune system: Modulation of innate and autoimmunity. J. Mol. Med. 2010, 88, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Yuk, J.-M.; Shin, D.-M.; Lee, H.-M.; Yang, C.-S.; Jin, H.S.; Kim, K.-K.; Lee, Z.-W.; Lee, S.-H.; Kim, J.-M.; Jo, E.-K.; et al. Vitamin D3 induces autophagy in human monocytes/macrophages via cathelicidin. Cell Host Microbe 2009, 6, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.R.; Spector, S.A. Autophagy induction by vitamin D inhibits both Mycobacterium tuberculosis and human immunodeficiency virus type 1. Autophagy 2012, 8, 1523–1525. [Google Scholar] [CrossRef] [PubMed]
- McVoy, L.A.; Kew, R.R. CD44 and annexin A2 mediate the C5a chemotactic cofactor function of the vitamin D binding protein. J. Immunol. 2005, 175, 4754–4760. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, L.E.; Wood, A.M.; Qureshi, O.S.; Hou, T.Z.; Gardner, D.; Briggs, Z.; Kaur, S.; Raza, K.; Sansom, D.M. Availability of 25-hydroxyvitamin D3 to APCs controls the balance between regulatory and inflammatory T cell responses. J. Immunol. 2012, 189, 5155–5164. [Google Scholar] [CrossRef] [PubMed]
- Verway, M.; Behr, M.A.; White, J.H. Vitamin D, NOD2, autophagy and Crohn’s disease. Expert Rev. Clin. Immunol. 2010, 6, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Hruz, P.; Eckmann, L. Innate immune defence: NOD2 and autophagy in the pathogenesis of Crohn’s disease. Swiss Med. Wkly. 2010. [Google Scholar] [CrossRef] [PubMed]
- Corridoni, D.; Kodani, T.; Rodriguez-Palacios, A.; Pizarro, T.T.; Xin, W.; Nickerson, K.P.; McDonald, C.; Ley, K.F.; Abbott, D.W.; et al. Dysregulated NOD2 predisposes SAMP1/YitFc mice to chronic intestinal inflammation. Proc. Natl. Acad. Sci. USA 2013, 110, 16999–17004. [Google Scholar] [CrossRef] [PubMed]
- Couturier-Maillard, A.; Secher, T.; Rehman, A.; Normand, S.; de Arcangelis, A.; Haesler, R.; Huot, L.; Grandiean, T.; Bressenot, A.; Delanoye-Crespin, A.; et al. NOD2-mediated dysbiosis predisposes mice to transmissible colitis and colorectal cancer. J. Clin. Investig. 2013, 123, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, V.; White, J.H. Vitamin D signaling in intestinal innate immunity and homeostasis. Mol. Cell. Endocrinol. 2017, 453, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Bruce, D.; Froicu, M.; Weaver, V.; Cantorna, M.T. Failure of T cell homing, reduced CD4/CD8 intraepithelial lymphocytes, and inflammation in the gut of vitamin D receptor KO mice. Proc. Natl. Acad. Sci. USA 2008, 105, 20834–20839. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.P.; Agnholt, J.; Glerup, H.; Lyhne, S.; Villadsen, G.E.; Hvas, C.L.; Bartels, L.E.; Kelsen, J.; Christensen, L.A.; Dahlerup, J.F. Clinical trial: Vitamin D3 treatment in Crohn’s disease—A randomized double-blind placebo-controlled study. Aliment. Pharmacol. Ther. 2010, 32, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Miheller, P.; Müzes, G.; Hritz, I.; Lakatos, G.; Pregun, I.; Lakatos, P.L.; Herszenyi, L. Comparison of the effects of 1,25 dihydroxyvitamin D and 25 hydroxyvitamin D on bone pathology and disease activity in Crohn’s disease patients. Inflamm. Bowel Dis. 2009, 15, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.; Hosseinzadeh-Attar, M.J.; Vahedi, H.; Nedjat, S. A randomized controlled trial on the effect of vitamin D3 on inflammation and cathelicidin gene expression in ulcerative colitis patients. Saudi J. Gastroenterol. 2016, 22, 316–323. [Google Scholar] [PubMed]
- Mathur, J.; Naing, S.; Mills, P.; Limsui, D. A randomized clinical trial of vitamin D3 (cholecalciferol) in ulcerative colitis patients with hypovitaminosis D3. PeerJ. 2017, 5, e3654. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Rosella, O.; Rosella, G.; Wu, Y.; Lubel, J.S.; Gibson, P.R. Evaluation of a 12-week targeted vitamin D supplementation regimen in patients with active inflammatory bowel disease. Clin. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Narula, N.; Cooray, M.; Anglin, R.; Muqtadir, Z.; Narula, A.; Marshall, J.K. Impact of High-Dose Vitamin D3 Supplementation in Patients with Crohn’s Disease in Remission: A Pilot Randomized Double-Blind Controlled Study. Dig. Dis. Sci. 2017, 62, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Dadaei, T.; Safapoor, M.H.; Asadzadeh Aghdaei, H.; Balaii, H.; Pourhoseingholi, M.A.; Naderi, N.; Zojaji, H.; Azimzadeh, P.; Mohammadi, P.; Zali, M.R. Effect of vitamin D3 supplementation on TNF-α serum level and disease activity index in Iranian IBD patients. Gastroenterol. Hepatol. Bed Bench 2015, 8, 49–55. [Google Scholar] [PubMed]
- Cantarel, B.L.; Waubant, E.; Chehoud, C.; Kuczynski, J.; DeSantis, T.Z.; Warrington, J.; Venkatesan, A.; Fraser, C.M.; Mowry, E.M. Gut microbiota in multiple sclerosis: Possible influence of immunomodulators. J. Investig. Med. 2015, 63, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.; Prietl, B.; Tauschmann, M.; Mautner, S.I.; Kump, P.K.; Treiber, G.; Wurm, P.; Gorkiewicz, G.; Hogenauer, C.; Pieber, T.R. Effects of high doses of vitamin D3 on mucosa-associated gut microbiome vary between regions of the human gastrointestinal tract. Eur. J. Nutr. 2016, 55, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Wu, S.; Zhang, Y.; Lu, R.; Petrof, E.O.; Yuan, L.; Claud, E.C.; Sun, J. Probiotic Regulation of Vitamin D Receptor in Intestinal Inflammation. Gastroenterology 2011. [Google Scholar] [CrossRef]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Oral supplementation with probiotic L. reuteri NCIMB 30242 increases mean circulating 25-hydroxyvitamin D: A post hoc analysis of a randomized controlled trial. J. Clin. Endocrinol. Metab. 2013, 98, 2944–2951. [Google Scholar] [CrossRef] [PubMed]
- Takada, I.; Makishima, M. Control of Inflammatory Bowel Disease and Colorectal Cancer by Synthetic Vitamin D Receptor Ligands. Curr. Med. Chem. 2017, 24, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Lubel, J.S.; Sparrow, M.P.; Holt, S.G.; Gibson, P.R. Review article: Vitamin D and inflammatory bowel disease—Established concepts and future directions. Aliment. Pharmacol. Ther. 2012, 36, 324–344. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Radeke, H.H.; Sartory, N.A.; Zahn, N.; Zuegel, U.; Steinmeyer, A.; Stein, J. The new low calcemic vitamin D analog 22-ene-25-oxa-vitamin D prominently ameliorates T helper cell type 1-mediated colitis in mice. J. Pharmacol. Exp. Ther. 2006, 319, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Strauch, U.G.; Obermeier, F.; Grunwald, N.; Dunger, N.; Rath, H.C.; Schölmerich, J.; Steinmeyer, A.; Zugel, U.; Herfarth, H.H. Calcitriol analog ZK191784 ameliorates acute and chronic dextran sodium sulfate-induced colitis by modulation of intestinal dendritic cell numbers and phenotype. World J. Gastroenterol. 2007, 13, 6529–6537. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, L.; Leyssens, C.; Beullens, I.; Marcelis, S.; Mathieu, C.; de Clercq, P.; Verstuyf, A. The vitamin D analog TX527 ameliorates disease symptoms in a chemically induced model of inflammatory bowel disease. J. Steroid Biochem. Mol. Biol. 2013, 136, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Liu, T.-J.; Shi, Y.-Y.; Zhao, Q. Vitamin D/VDR signaling pathway ameliorates 2,4,6-trinitrobenzene sulfonic acid-induced colitis by inhibiting intestinal epithelial apoptosis. Int. J. Mol. Med. 2015, 35, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Schlauch, T.; Zügel, U.; Steinmeyer, A.; Radeke, H.H.; Steinhilber, D.; Stein, J. 22-ene-25-oxa-vitamin D: A new vitamin D analogue with profound immunosuppressive capacities. Eur. J. Clin. Investig. 2005, 35, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Stio, M.; Martinesi, M.; Bruni, S.; Treves, C.; Mathieu, C.; Verstuyf, A.; d’Albasio, G.; Bagnoli, S.; Bonanomi, A.G. The vitamin D analogue TX 527 blocks NF-κB activation in peripheral blood mononuclear cells of patients with Crohn’s disease. J. Steroid Biochem. Mol. Biol. 2007, 103, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Van Belle, T.L.; Vanherwegen, A.-S.; Feyaerts, D.; de Clercq, P.; Verstuyf, A.; Korf, H.; Gysemans, C.; Mathieu, C. 1,25-Dihydroxyvitamin D3 and its analog TX527 promote a stable regulatory T cell phenotype in T cells from type 1 diabetes patients. PLoS ONE 2014, 9, e109194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laverny, G.; Penna, G.; Vetrano, S.; Correale, C.; Nebuloni, M.; Danese, S.; Adorini, L. Efficacy of a potent and safe vitamin D receptor agonist for the treatment of inflammatory bowel disease. Immunol. Lett. 2010, 131, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Koszewski, N.J.; Horst, R.L.; Goff, J.P. Importance of apical membrane delivery of 1,25-dihydroxyvitamin D3 to vitamin D-responsive gene expression in the colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G870–G878. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Koszewski, N.J.; Haynes, J.S.; Horst, R.L. Targeted delivery of vitamin D to the colon using β-glucuronides of vitamin D: Therapeutic effects in a murine model of inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G460–G469. [Google Scholar] [CrossRef] [PubMed]
- Barry, E.L.; Rees, J.R.; Peacock, J.L.; Mott, L.A.; Amos, C.I.; Bostick, R.M.; Figueiredo, J.C.; Ahnen, D.J.; Bresalier, R.S.; Burke, C.A.; et al. Genetic variants in CYP2R1, CYP24A1, and VDR modify the efficacy of vitamin D3 supplementation for increasing serum 25-hydroxyvitamin D levels in a randomized controlled trial. J. Clin. Endocrinol. Metab. 2014, 99, E2133–E2137. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Del Pinto, R.; Ferri, C.; Cominelli, F. Vitamin D Axis in Inflammatory Bowel Diseases: Role, Current Uses and Future Perspectives. Int. J. Mol. Sci. 2017, 18, 2360. https://doi.org/10.3390/ijms18112360
AMA Style
Del Pinto R, Ferri C, Cominelli F. Vitamin D Axis in Inflammatory Bowel Diseases: Role, Current Uses and Future Perspectives. International Journal of Molecular Sciences. 2017; 18(11):2360. https://doi.org/10.3390/ijms18112360
Chicago/Turabian StyleDel Pinto, Rita, Claudio Ferri, and Fabio Cominelli. 2017. "Vitamin D Axis in Inflammatory Bowel Diseases: Role, Current Uses and Future Perspectives" International Journal of Molecular Sciences 18, no. 11: 2360. https://doi.org/10.3390/ijms18112360
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.