Role of Placental VDR Expression and Function in Common Late Pregnancy Disorders


Abstract
:1. Introduction
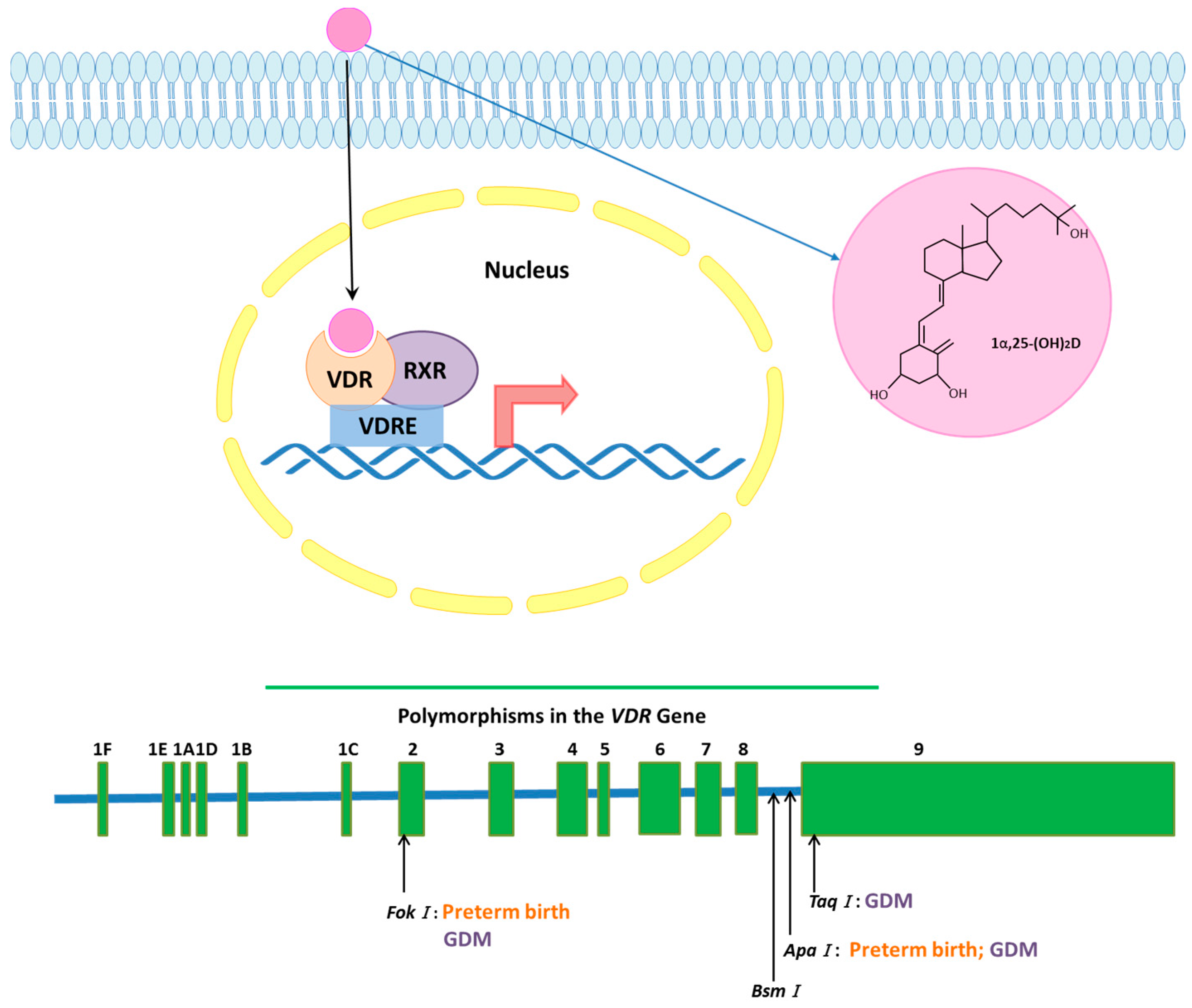
1.1. Vitamin D Receptor (VDR) Signaling
1.2. VDR Gene and Polymorphisms
2. VDR Expression in Physiologic Pregnancy
3. VDR and Preeclampsia
3.1. VDR Expression Changes in Preeclampsia
3.2. Role of VDR Polymorphisms in Preeclampsia
4. VDR and Fetal Growth Restriction
4.1. VDR Expression and Signaling in Fetal Growth Restriction
4.2. VDR Polymorphisms in Fetal Growth Restriction
5. VDR and Diabetes in Pregnancy
5.1. VDR Expression in Gestational Diabetes Mellitus
5.2. VDR Polymorphisms and Gestational Diabetes Mellitus
6. VDR and Preterm Birth
6.1. VDR Signaling in Preterm Birth
6.2. Role of VDR Polymorphisms in Preterm Birth
7. Existing Knowledge from VDR Knock-Out Models
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CAMP | Cathelicidin |
| EVT | Extra villous trophoblast |
| FGR | Fetal growth restriction |
| GDM | Gestational diabetes mellitus |
| LPS | Lipopolysaccharide |
| PTB | Preterm birth |
| SGA | Small for gestational age |
| T1DM | Type 1 diabetes mellitus |
| T2DM | Type 2 diabetes mellitus |
| TL4 | Toll-like receptor 4 |
| UtSM | Myometrial smooth muscle |
| VDR | Vitamin D receptor |
| VSMC | Smooth muscle cells of the placental vessels |
References
- Holick, M.F. Vitamin D: Importance in the prevention of cancers, type 1 diabetes, heart disease, and osteoporosis. Am. J. Clin. Nutr. 2004, 79, 362–371. [Google Scholar] [PubMed]
- Barrera, D.; Avila, E.; Hernandez, G.; Mendez, I.; Gonzalez, L.; Halhali, A.; Larrea, F.; Morales, A.; Diaz, L. Calcitriol affects hcg gene transcription in cultured human syncytiotrophoblasts. Reprod. Biol. Endocrinol. 2008, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Jones, G. Promise of vitamin D analogues in the treatment of hyperproliferative conditions. Mol. Cancer Ther. 2006, 5, 797–808. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin d. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [PubMed]
- Kovacs, C.S.; Kronenberg, H.M. Maternal-fetal calcium and bone metabolism during pregnancy, puerperium, and lactation. Endocr. Rev. 1997, 18, 832–872. [Google Scholar] [CrossRef] [PubMed]
- El Baba, K.; Zantout, M.S.; Akel, R.; Azar, S.T. Seasonal variation of vitamin D and hba(1c) levels in patients with type 1 diabetes mellitus in the middle east. Int. J. Gen. Med. 2011, 4, 635–638. [Google Scholar] [PubMed]
- Baz-Hecht, M.; Goldfine, A.B. The impact of vitamin D deficiency on diabetes and cardiovascular risk. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qiu, C.; Hu, F.B.; David, R.M.; van Dam, R.M.; Bralley, A.; Williams, M.A. Maternal plasma 25-hydroxyvitamin d concentrations and the risk for gestational diabetes mellitus. PLoS ONE 2008, 3, e3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haussler, M.R.; Haussler, C.A.; Bartik, L.; Whitfield, G.K.; Hsieh, J.C.; Slater, S.; Jurutka, P.W. Vitamin D receptor: Molecular signaling and actions of nutritional ligands in disease prevention. Nutr. Rev. 2008, 66, S98–S112. [Google Scholar] [CrossRef] [PubMed]
- Palomer, X.; González-Clemente, J.M.; Blanco-Vaca, F.; Mauricio, D. Role of vitamin D in the pathogenesis of type 2 diabetes mellitus. Diabetes Obes. Metab. 2008, 10, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W.; Bishop, J.E.; Bula, C.M.; Olivera, C.J.; Mizwicki, M.T.; Zanello, L.P.; Ishida, H.; Okamura, W.H. Molecular tools for study of genomic and rapid signal transduction responses initiated by 1 alpha,25(oh)(2)-vitamin d(3). Steroids 2002, 67, 457–466. [Google Scholar] [CrossRef]
- Menegaz, D.; Mizwicki, M.T.; Barrientos-Duran, A.; Chen, N.; Henry, H.L.; Norman, A.W. Vitamin d receptor (vdr) regulation of voltage-gated chloride channels by ligands preferring a vdr-alternative pocket (vdr-ap). Mol. Endocrinol. 2011, 25, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Choi, M.Y.; Longtine, M.S.; Nelson, D.M. Vitamin d effects on pregnancy and the placenta. Placenta 1027, 31, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Fujiwara, T.M.; Ross, M.V.; Morgan, K.; Garcia-Heras, J.; Ledbetter, D.H.; Hughes, M.R.; Glorieux, F.H. Two hereditary defects related to vitamin D metabolism map to the same region of human chromosome 12q13–14. J. Bone Miner. Res. 1992, 7, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Uitterlinden, A.G.; Fang, Y.; van Meurs, J.B.J.; van Leeuwen, H.; Pols, H.A.P. Vitamin D receptor gene polymorphisms in relation to vitamin D related disease states. J. Steroid Biochem. Mol. Biol. 2004, 89, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Daiger, S.P.; Schanfield, M.S.; Cavalli-Sforza, L.L. Group-specific component (gc) proteins bind vitamin D and 25-hydroxyvitamin d. Proc. Natl. Acad. Sci. USA 1975, 72, 2076–2080. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; van Meurs, J.B.; Berry, D.; Kiel, D.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef]
- Uitterlinden, A.G.; Fang, Y.; Van Meurs, J.B.; Pols, H.A.; Van Leeuwen, J.P. Genetics and biology of vitamin D receptor polymorphisms. Gene 2004, 338, 143–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzon, L.; Altarescu, G.; Tevet, A.; Schimmel, M.S.; Elstein, D.; Samueloff, A.; Grisaru-Granovsky, S. Vitamin d receptor polymorphism foki is associated with spontaneous idiopathic preterm birth in an israeli population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 177, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Schirmer, H.; Wilsgaard, T.; Joakimsen, R.M.; Mathiesen, E.B.; Njølstad, I.; Løchen, M.-L.; Figenschau, Y.; Berg, J.P.; Svartberg, J.; et al. Polymorphisms related to the serum 25-hydroxyvitamin d level and risk of myocardial infarction, diabetes, cancer and mortality. The tromsø study. PLoS ONE 2012, 7, e37295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdivielso, J.M.; Fernandez, E. Vitamin d receptor polymorphisms and diseases. Clin. Chim. Acta 2006, 371, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, G.K.; Remus, L.S.; Jurutka, P.W.; Zitzer, H.; Oza, A.K.; Dang, H.T.; Haussler, C.A.; Galligan, M.A.; Thatcher, M.L.; Encinas Dominguez, C.; et al. Functionally relevant polymorphisms in the human nuclear vitamin D receptor gene. Mol. Cell. Endocrinol. 2001, 177, 145–159. [Google Scholar] [CrossRef]
- Murthi, P.; Yong, H.E.J.; Ngyuen, T.P.H.; Ellery, S.; Singh, H.; Rahman, R.; Dickinson, H.; Walker, D.W.; Davies-Tuck, M.; Wallace, E.M.; et al. Role of the placental vitamin D receptor in modulating feto-placental growth in fetal growth restriction and preeclampsia-affected pregnancies. Front. Physiol. 2016, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Avila, E.; Hernandez, G.; Halhali, A.; Biruete, B.; Larrea, F.; Diaz, L. Estradiol and progesterone synthesis in human placenta is stimulated by calcitriol. J. Steroid Biochem. Mol. Biol. 2007, 103, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.N.; Bulmer, J.N.; Kilby, M.D.; Hewison, M. Vitamin D and placental-decidual function. J. Soc. Gynecol. Investig. 2004, 11, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Acker, G.M.; Garabedian, M. 1,25-dihydroxyvitamin d3 induces in vivo the decidualization of rat endometrial cells. J. Reprod. Fertil. 1991, 91, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Stephanou, A.; Ross, R.; Handwerger, S. Regulation of human placental lactogen expression by 1,25-dihydroxyvitamin d3. Endocrinology 1994, 135, 2651–2656. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, D.; Evans, K.N.; Kilby, M.D.; Bulmer, J.N.; Innes, B.A.; Stewart, P.M.; Hewison, M. The ontogeny of 25-hydroxyvitamin d(3) 1α-hydroxylase expression in human placenta and decidua. Am. J. Pathol. 2002, 161, 105–114. [Google Scholar] [CrossRef]
- Avila, E.; Diaz, L.; Barrera, D.; Halhali, A.; Mendez, I.; Gonzalez, L.; Zuegel, U.; Steinmeyer, A.; Larrea, F. Regulation of vitamin D hydroxylases gene expression by 1,25-dihydroxyvitamin d3 and cyclic amp in cultured human syncytiotrophoblasts. J. Steroid Biochem. Mol. Biol. 2007, 103, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Kaplan, A.T.; Low, J.; Nguyen, L.; Liu, G.Y.; Equils, O.; Hewison, M. Vitamin D induces innate antibacterial responses in human trophoblasts via an intracrine pathway. Biol. Reprod. 2009, 80, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.; Jeddi-Tehrani, M.; Zareie, M.; Salek-Moghaddam, A.; Akhondi, M.M.; Bahmanpoor, M.; Sadeghi, M.R.; Zarnani, A.H. Expression profiling of vitamin D receptor in placenta, decidua and ovary of pregnant mice. Placenta 2011, 32, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Pospechova, K.; Rozehnal, V.; Stejskalova, L.; Vrzal, R.; Pospisilova, N.; Jamborova, G.; May, K.; Siegmund, W.; Dvorak, Z.; Nachtigal, P.; et al. Expression and activity of vitamin D receptor in the human placenta and in choriocarcinoma bewo and jeg-3 cell lines. Mol. Cell. Endocrinol. 2009, 299, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.N.; Nguyen, L.; Chan, J.; Innes, B.A.; Bulmer, J.N.; Kilby, M.D.; Hewison, M. Effects of 25-hydroxyvitamin d3 and 1,25-dihydroxyvitamin d3 on cytokine production by human decidual cells. Biol. Reprod. 2006, 75, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; Kaplan, A.T.; Lagishetty, V.; Ouyang, Y.B.; Ouyang, Y.; Simmons, C.F.; Equils, O.; Hewison, M. Vitamin d and the regulation of placental inflammation. J. Immunol. 2011, 186, 5968–5974. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, molecular mechanism of action, and pleiotropic effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-P.; Bajoria, R.; Aplin, J.D. Decreased vascularization and cell proliferation in placentas of intrauterine growth–restricted fetuses with abnormal umbilical artery flow velocity waveforms. Am. J. Obstet. Gynecol. 2002, 187, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Catov, J.M.; Simhan, H.N.; Holick, M.F.; Powers, R.W.; Roberts, J.M. Maternal vitamin D deficiency increases the risk of preeclampsia. J. Clin. Endocrinol. Metab. 2007, 92, 3517–3522. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Schroer, A.; Ludders, D.; Cordes, T.; Bucker, B.; Reichrath, J.; Friedrich, M. Metabolism of vitamin d3 in the placental tissue of normal and preeclampsia complicated pregnancies and premature births. Clin. Exp. Obstet. Gynecol. 2007, 34, 80–84. [Google Scholar] [PubMed]
- Baker, A.M.; Haeri, S.; Camargo, C.A., Jr.; Espinola, J.A.; Stuebe, A.M. A nested case-control study of midgestation vitamin D deficiency and risk of severe preeclampsia. J. Clin. Endocrinol. Metab. 2010, 95, 5105–5109. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Alanis, M.C.; Wagner, C.L.; Hollis, B.W.; Johnson, D.D. Plasma 25-hydroxyvitamin d levels in early-onset severe preeclampsia. Am. J. Obstet. Gynecol. 2010, 203. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Simhan, H.N.; Catov, J.M.; Roberts, J.M.; Platt, R.W.; Diesel, J.C.; Klebanoff, M.A. Maternal vitamin D status and the risk of mild and severe preeclampsia. Epidemiology 2014, 25, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Tamblyn, J.A.; Susarla, R.; Jenkinson, C.; Jeffery, L.E.; Ohizua, O.; Chun, R.F.; Chan, S.Y.; Kilby, M.D.; Hewison, M. Dysregulation of maternal and placental vitamin D metabolism in preeclampsia. Placenta 2017, 50, 70–77. [Google Scholar] [CrossRef] [PubMed]
- August, P.; Marcaccio, B.; Gertner, J.M.; Druzin, M.L.; Resnick, L.M.; Laragh, J.H. Abnormal 1,25-dihydroxyvitamin d metabolism in preeclampsia. Am. J. Obstet. Gynecol. 1992, 166, 1295–1299. [Google Scholar] [CrossRef]
- Halhali, A.; Tovar, A.R.; Torres, N.; Bourges, H.; Garabedian, M.; Larrea, F. Preeclampsia is associated with low circulating levels of insulin-like growth factor i and 1,25-dihydroxyvitamin d in maternal and umbilical cord compartments. J. Clin. Endocrinol. Metab. 2000, 85, 1828–1833. [Google Scholar] [PubMed]
- Seely, E.W.; Wood, R.J.; Brown, E.M.; Graves, S.W. Lower serum ionized calcium and abnormal calciotropic hormone levels in preeclampsia. J. Clin. Endocrinol. Metab. 1992, 74, 1436–1440. [Google Scholar] [PubMed]
- Hypponen, E. Vitamin d for the prevention of preeclampsia? A hypothesis. Nutr. Rev. 2005, 63, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Susarla, R.; Canovas, D.; Vasilopoulou, E.; Ohizua, O.; McCabe, C.J.; Hewison, M.; Kilby, M.D. Vitamin D promotes human extravillous trophoblast invasion in vitro. Placenta 2015, 36, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Tarcin, O.; Yavuz, D.G.; Ozben, B.; Telli, A.; Ogunc, A.V.; Yuksel, M.; Toprak, A.; Yazici, D.; Sancak, S.; Deyneli, O.; et al. Effect of vitamin D deficiency and replacement on endothelial function in asymptomatic subjects. J. Clin. Endocrinol. Metab. 2009, 94, 4023–4030. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Sibson, M.; Ng, H.K.; Manuelpillai, U.; Rakyan, V.; Down, T.; Beck, S.; Fournier, T.; Evain-Brion, D.; Dimitriadis, E.; et al. Placenta-specific methylation of the vitamin D 24-hydroxylase gene: Implications for feedback autoregulation of active vitamin D levels at the fetomaternal interface. J. Biol. Chem. 2009, 284, 14838–14848. [Google Scholar] [CrossRef] [PubMed]
- Saffery, R.; Ellis, J.; Morley, R. A convergent model for placental dysfunction encompassing combined sub-optimal one-carbon donor and vitamin D bioavailability. Med. Hypotheses 2009, 73, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, N.C.; Said, E.S.; Stathopoulou, M.G.; Siest, G.; Tsai, M.Y.; Visvikis-Siest, S. Epistatic study reveals two genetic interactions in blood pressure regulation. BMC Med. Genet. 2013, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, J.; Manson, J.E.; Buring, J.E.; Gaziano, J.M.; Sesso, H.D. A prospective study of plasma vitamin D metabolites, vitamin D receptor gene polymorphisms, and risk of hypertension in men. Eur. J. Nutr. 2013, 52, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Swapna, N.; Vamsi, U.M.; Usha, G.; Padma, T. Risk conferred by foki polymorphism of vitamin D receptor (vdr) gene for essential hypertension. Indian J. Hum. Genet. 2011, 17, 201–206. [Google Scholar] [PubMed]
- Li, Y.C. Vitamin d regulation of the renin–angiotensin system. J. Cell. Biochem. 2003, 88, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.; Sun, B.; Forman, J.P.; Hopkins, P.N.; Brown, N.J.; Kolatkar, N.S.; Williams, G.H.; Williams, J.S. The fok1 vitamin D receptor gene polymorphism is associated with plasma renin activity in caucasians. Clin. Endocrinol. 2011, 74, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Sandrim, V.C.; Palei, A.C.; Cavalli, R.C.; Araujo, F.M.; Ramos, E.S.; Duarte, G.; Tanus-Santos, J.E. Enos haplotypes associated with gestational hypertension or preeclampsia. Pharmacogenomics 2008, 9, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Sandrim, V.C.; Palei, A.C.; Luizon, M.R.; Izidoro-Toledo, T.C.; Cavalli, R.C.; Tanus-Santos, J.E. Enos haplotypes affect the responsiveness to antihypertensive therapy in preeclampsia but not in gestational hypertension. Pharmacogenom. J. 2010, 10, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Palei, A.C.; Sandrim, V.C.; Duarte, G.; Cavalli, R.C.; Gerlach, R.F.; Tanus-Santos, J.E. Matrix metalloproteinase (mmp)-9 genotypes and haplotypes in preeclampsia and gestational hypertension. Clin. Chim. Acta 2010, 411, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Rezende, V.B.; Sandrim, V.C.; Palei, A.C.; Machado, L.; Cavalli, R.C.; Duarte, G.; Tanus-Santos, J.E. Vitamin D receptor polymorphisms in hypertensive disorders of pregnancy. Mol. Biol. Rep. 2012, 39, 10903–10906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntire, D.D.; Bloom, S.L.; Casey, B.M.; Leveno, K.J. Birth weight in relation to morbidity and mortality among newborn infants. N. Engl. J. Med. 1999, 340, 1234–1238. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, M.; Gardosi, J. Fetal growth. Curr. Opin. Obstet. Gynecol. 2000, 12, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Scifres, C.M.; Nelson, D.M. Intrauterine growth restriction, human placental development and trophoblast cell death. J. Physiol. 2009, 587, 3453–3458. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Barker, D.J. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Eriksson, J.G.; Forsen, T.; Osmond, C. Fetal origins of adult disease: Strength of effects and biological basis. Int. J. Epidemiol. 2002, 31, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, F.H.; Sorensen, H.T.; Gillman, M.W.; Rothman, K.J.; Sabroe, S.; Fischer, P.; Olsen, J. Low birth weight and preterm delivery as risk factors for asthma and atopic dermatitis in young adult males. Epidemiology 2000, 11, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Frisk, V.; Amsel, R.; Whyte, H.E. The importance of head growth patterns in predicting the cognitive abilities and literacy skills of small-for-gestational-age children. Dev. Neuropsychol. 2002, 22, 565–593. [Google Scholar] [CrossRef] [PubMed]
- Rosso, I.M.; Cannon, T.D.; Huttunen, T.; Huttunen, M.O.; Lonnqvist, J.; Gasperoni, T.L. Obstetric risk factors for early-onset schizophrenia in a finnish birth cohort. Am. J. Psychiatry 2000, 157, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.R.; Martyn, C.N. Birth weight and later risk of depression in a national birth cohort. Br. J. Psychiatry J. Ment. Sci. 2004, 184, 28–33. [Google Scholar] [CrossRef]
- Sankaran, S.; Kyle, P.M. Aetiology and pathogenesis of iugr. Best Pract. Res. Clin. Obstet. Gynaecol. 2009, 23, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.S.; Moore, V.M.; Owens, J.A.; McMillen, I.C. Origins of fetal growth restriction. Eur J. Obstet. Gynecol. Reprod. Biol. 2000, 92, 13–19. [Google Scholar] [CrossRef]
- Wei, S.Q.; Qi, H.P.; Luo, Z.C.; Fraser, W.D. Maternal vitamin D status and adverse pregnancy outcomes: A systematic review and meta-analysis. J. Matern.-Fetal Neonatal Med. 2013, 26, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Billaudel, B.; Labriji-Mestaghanmi, H.; Sutter, B.C.; Malaisse, W.J. Vitamin D and pancreatic islet function. II. Dynamics of insulin release and cationic fluxes. J. Endocrinol. Investig. 1988, 11, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Galthen-Sorensen, M.; Andersen, L.B.; Sperling, L.; Christesen, H.T. Maternal 25-hydroxyvitamin d level and fetal bone growth assessed by ultrasound: A systematic review. Ultrasound Obstet. Gynecol. 2014, 44, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Schneuer, F.J.; Roberts, C.L.; Guilbert, C.; Simpson, J.M.; Algert, C.S.; Khambalia, A.Z.; Tasevski, V.; Ashton, A.W.; Morris, J.M.; Nassar, N. Effects of maternal serum 25-hydroxyvitamin d concentrations in the first trimester on subsequent pregnancy outcomes in an australian population. Am. J. Clin. Nutr. 2014, 99, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, C.L.; Gernand, A.D.; Roth, D.E.; Bodnar, L.M. Maternal vitamin D status and infant anthropometry in a us multi-centre cohort study. Ann. Hum. Biol. 2015, 42, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Gernand, A.D.; Simhan, H.N.; Klebanoff, M.A.; Bodnar, L.M. Maternal serum 25-hydroxyvitamin d and measures of newborn and placental weight in a U.S. Multicenter cohort study. J. Clin. Endocrinol. Metab. 2013, 98, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Eggemoen, A.R.; Jenum, A.K.; Mdala, I.; Knutsen, K.V.; Lagerlov, P.; Sletner, L. Vitamin d levels during pregnancy and associations with birth weight and body composition of the newborn: A longitudinal multiethnic population-based study. Br. J. Nutr. 2017, 117, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Crocker, I.P.; Cooper, S.; Ong, S.C.; Baker, P.N. Differences in apoptotic susceptibility of cytotrophoblasts and syncytiotrophoblasts in normal pregnancy to those complicated with preeclampsia and intrauterine growth restriction. Am. J. Pathol. 2003, 162, 637–643. [Google Scholar] [CrossRef]
- Newhouse, S.M.; Davidge, S.T.; Winkler-Lowen, B.; Demianczuk, N.; Guilbert, L.J. In vitro differentiation of villous trophoblasts from pregnancies complicated by intrauterine growth restriction with and without pre-eclampsia. Placenta 2007, 28, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Samuel, S.; Sitrin, M.D. Vitamin d's role in cell proliferation and differentiation. Nutr. Rev. 2008, 66, S116–S124. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.P.; Yong, H.E.; Chollangi, T.; Borg, A.J.; Brennecke, S.P.; Murthi, P. Placental vitamin D receptor expression is decreased in human idiopathic fetal growth restriction. J. Mol. Med. (Berl.) 2015, 93, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B.; Kingdom, J.C. Apoptosis in the trophoblast--role of apoptosis in placental morphogenesis. J. Soc. Gynecol. Investig. 2004, 11, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Toaldo, C.; Pizzimenti, S.; Cerbone, A.; Pettazzoni, P.; Dianzani, M.U.; Ferretti, C. The role of ppar ligands in controlling growth-related gene expression and their interaction with lipoperoxidation products. PPAR Res. 2008, 2008, 524671. [Google Scholar] [CrossRef] [PubMed]
- Tuan, R.S.; Moore, C.J.; Brittingham, J.W.; Kirwin, J.J.; Akins, R.E.; Wong, M. In vitro study of placental trophoblast calcium uptake using JEG-3 human choriocarcinoma cells. J. Cell Sci. 1991, 98 Pt 3, 333–342. [Google Scholar] [PubMed]
- Avila, E.; Diaz, L.; Halhali, A.; Larrea, F. Regulation of 25-hydroxyvitamin d3 1alpha-hydroxylase, 1,25-dihydroxyvitamin d3 24-hydroxylase and vitamin D receptor gene expression by 8-bromo cyclic AMP in cultured human syncytiotrophoblast cells. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sandovici, I.; Hoelle, K.; Angiolini, E.; Constancia, M. Placental adaptations to the maternal-fetal environment: Implications for fetal growth and developmental programming. Reprod. Biomed. Online 2012, 25, 68–89. [Google Scholar] [CrossRef] [PubMed]
- Jurutka, P.W.; Whitfield, G.K.; Hsieh, J.C.; Thompson, P.D.; Haussler, C.A.; Haussler, M.R. Molecular nature of the vitamin D receptor and its role in regulation of gene expression. Rev. Endocr. Metab. Disord. 2001, 2, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Morley, J.E. Vitamin D redux. J. Am. Med. Dir. Assoc. 2009, 10, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Swamy, G.K.; Garrett, M.E.; Miranda, M.L.; Ashley-Koch, A.E. Maternal vitamin D receptor genetic variation contributes to infant birthweight among black mothers. Am. J. Med. Genet. Part A 2011, 155, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Metzger, B.E.; Coustan, D.R. Summary and recommendations of the fourth international workshop-conference on gestational diabetes mellitus. The organizing committee. Diabetes Care 1998, 21 (Suppl. 2), B161–B167. [Google Scholar] [PubMed]
- Knabl, J.; Huttenbrenner, R.; Hutter, S.; Gunthner-Biller, M.; Riedel, C.; Hiden, U.; Kainer, F.; Desoye, G.; Jeschke, U. Gestational diabetes mellitus upregulates vitamin D receptor in extravillous trophoblasts and fetoplacental endothelial cells. Reprod. Sci. 2015, 22, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Butte, N.F. Carbohydrate and lipid metabolism in pregnancy: Normal compared with gestational diabetes mellitus. Am. J. Clin. Nutr. 2000, 71, 1256s–1261s. [Google Scholar] [PubMed]
- Hartling, L.; Dryden, D.M.; Guthrie, A.; Muise, M.; Vandermeer, B.; Aktary, W.M.; Pasichnyk, D.; Seida, J.C.; Donovan, L. Screening and diagnosing gestational diabetes mellitus. Evid. Rep./Technol. Assess. 2012, 210, 1–327. [Google Scholar]
- Nar, G.; Inci, S.; Aksan, G.; Unal, O.K.; Nar, R.; Soylu, K. The relationship between epicardial fat thickness and gestational diabetes mellitus. Diabetol. Metab. Syndr. 2014, 6, 120. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D. Type 2 diabetes mellitus after gestational diabetes: A systematic review and meta-analysis. Lancet 2009, 373, 1773–1779. [Google Scholar] [CrossRef]
- Soheilykhah, S.; Mojibian, M.; Rashidi, M.; Rahimi-Saghand, S.; Jafari, F. Maternal vitamin D status in gestational diabetes mellitus. Nutr. Clin. Pract. 2010, 25, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Maghbooli, Z.; Hossein-Nezhad, A.; Karimi, F.; Shafaei, A.R.; Larijani, B. Correlation between vitamin d3 deficiency and insulin resistance in pregnancy. Diabetes Metab. Res. Rev. 2008, 24, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Clifton-Bligh, R.J.; McElduff, P.; McElduff, A. Maternal vitamin D deficiency, ethnicity and gestational diabetes. Diabet. Med. 2008, 25, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Haeri, S.; Camargo, C.A., Jr.; Stuebe, A.M.; Boggess, K.A. First-trimester maternal vitamin D status and risk for gestational diabetes (gdm) a nested case-control study. Diabetes Metab. Res. Rev. 2012, 28, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Makgoba, M.; Nelson, S.M.; Savvidou, M.; Messow, C.M.; Nicolaides, K.; Sattar, N. First-trimester circulating 25-hydroxyvitamin d levels and development of gestational diabetes mellitus. Diabetes Care 2011, 34, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Farrant, H.J.; Krishnaveni, G.V.; Hill, J.C.; Boucher, B.J.; Fisher, D.J.; Noonan, K.; Osmond, C.; Veena, S.R.; Fall, C.H. Vitamin D insufficiency is common in indian mothers but is not associated with gestational diabetes or variation in newborn size. Eur. J. Clin. Nutr. 2009, 63, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Burris, H.H.; Rifas-Shiman, S.L.; Kleinman, K.; Litonjua, A.A.; Huh, S.Y.; Rich-Edwards, J.W.; Camargo, C.A., Jr.; Gillman, M.W. Vitamin D deficiency in pregnancy and gestational diabetes mellitus. Am. J. Obstet. Gynecol. 2012, 207. [Google Scholar] [CrossRef] [PubMed]
- Aghajafari, F.; Nagulesapillai, T.; Ronksley, P.E.; Tough, S.C.; O'Beirne, M.; Rabi, D.M. Association between maternal serum 25-hydroxyvitamin d level and pregnancy and neonatal outcomes: Systematic review and meta-analysis of observational studies. BMJ 2013, 346, f1169. [Google Scholar] [CrossRef] [PubMed]
- Triunfo, S.; Lanzone, A.; Lindqvist, P.G. Low maternal circulating levels of vitamin D as potential determinant in the development of gestational diabetes mellitus. J. Endocrinol. Investig. 2017, 40, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Darwish, H.; DeLuca, H.F. Vitamin D-regulated gene expression. Crit. Rev. Eukaryot. Gene Expr. 1993, 3, 89–116. [Google Scholar] [PubMed]
- Potera, C. Diet and nutrition: Vitamin D regulates ms gene. Environ. Health Perspect. 2009, 117, A196. [Google Scholar] [CrossRef] [PubMed]
- Senti, J.; Thiele, D.K.; Anderson, C.M. Maternal vitamin D status as a critical determinant in gestational diabetes. J. Obstet. Gynecol. Neonatal Nurs. JOGNN 2012, 41, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Bourlon, P.M.; Billaudel, B.; Faure-Dussert, A. Influence of vitamin d3 deficiency and 1,25 dihydroxyvitamin d3 on de novo insulin biosynthesis in the islets of the rat endocrine pancreas. J. Endocrinol. 1999, 160, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Cade, C.; Norman, A.W. Vitamin d3 improves impaired glucose tolerance and insulin secretion in the vitamin d-deficient rat in vivo. Endocrinology 1986, 119, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.; Bonner-Weir, S.; Norman, A.W.; Orci, L. Immunocytochemistry of vitamin d-dependent calcium binding protein in chick pancreas: Exclusive localization. Endocrinology 1982, 110, 2216–2218. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, S.; Norman, A.W. Dietary vitamin D is essential for normal insulin secretion from the perfused rat pancreas. J. Clin. Investig. 1984, 73, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W.; Frankel, J.B.; Heldt, A.M.; Grodsky, G.M. Vitamin D deficiency inhibits pancreatic secretion of insulin. Science 1980, 209, 823–825. [Google Scholar] [CrossRef] [PubMed]
- Chiu, K.C.; Chu, A.; Go, V.L.; Saad, M.F. Hypovitaminosis d is associated with insulin resistance and beta cell dysfunction. Am. J. Clin. Nutr. 2004, 79, 820–825. [Google Scholar] [PubMed]
- Scragg, R.; Holdaway, I.; Singh, V.; Metcalf, P.; Baker, J.; Dryson, E. Serum 25-hydroxyvitamin d3 levels decreased in impaired glucose tolerance and diabetes mellitus. Diabetes Res. Clin. Pract. 1995, 27, 181–188. [Google Scholar] [CrossRef]
- Ojuka, E.O. Role of calcium and amp kinase in the regulation of mitochondrial biogenesis and glut4 levels in muscle. Proc. Nutr. Soc. 2004, 63, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.C.; Hucker, K.A.; Holloszy, J.O.; Han, D.H. Ca2+ and ampk both mediate stimulation of glucose transport by muscle contractions. Diabetes 2004, 53, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.F.; Caterson, I.D.; Cooney, G.J.; Zilkens, R.R.; Turtle, J.R. High affinity insulin binding and insulin receptor-effector coupling: Modulation by Ca2+. Cell Calcium 1990, 11, 547–556. [Google Scholar] [CrossRef]
- Draznin, B.; Sussman, K.; Kao, M.; Lewis, D.; Sherman, N. The existence of an optimal range of cytosolic free calcium for insulin-stimulated glucose transport in rat adipocytes. J. Biol. Chem. 1987, 262, 14385–14388. [Google Scholar] [PubMed]
- Plehwe, W.E.; Williams, P.F.; Caterson, I.D.; Harrison, L.C.; Turtle, J.R. Calcium-dependence of insulin receptor phosphorylation. Biochem. J. 1983, 214, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Zemel, M.B. Nutritional and endocrine modulation of intracellular calcium: Implications in obesity, insulin resistance and hypertension. Mol. Cell. Biochem. 1998, 188, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Reusch, J.E.; Begum, N.; Sussman, K.E.; Draznin, B. Regulation of glut-4 phosphorylation by intracellular calcium in adipocytes. Endocrinology 1991, 129, 3269–3273. [Google Scholar] [CrossRef] [PubMed]
- Maestro, B.; Davila, N.; Carranza, M.C.; Calle, C. Identification of a vitamin D response element in the human insulin receptor gene promoter. J. Steroid Biochem. Mol. Biol. 2003, 84, 223–230. [Google Scholar] [CrossRef]
- Maestro, B.; Molero, S.; Bajo, S.; Davila, N.; Calle, C. Transcriptional activation of the human insulin receptor gene by 1,25-dihydroxyvitamin d(3). Cell Biochem. Funct. 2002, 20, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D. Nonclassic actions of vitamin D. J. Clin. Endocrinol. Metab. 2009, 94, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Riachy, R.; Vandewalle, B.; Kerr Conte, J.; Moerman, E.; Sacchetti, P.; Lukowiak, B.; Gmyr, V.; Bouckenooghe, T.; Dubois, M.; Pattou, F. 1,25-dihydroxyvitamin d3 protects rinm5f and human islet cells against cytokine-induced apoptosis: Implication of the antiapoptotic protein a20. Endocrinology 2002, 143, 4809–4819. [Google Scholar] [CrossRef] [PubMed]
- Gysemans, C.A.; Cardozo, A.K.; Callewaert, H.; Giulietti, A.; Hulshagen, L.; Bouillon, R.; Eizirik, D.L.; Mathieu, C. 1,25-dihydroxyvitamin d3 modulates expression of chemokines and cytokines in pancreatic islets: Implications for prevention of diabetes in nonobese diabetic mice. Endocrinology 2005, 146, 1956–1964. [Google Scholar] [CrossRef] [PubMed]
- van Etten, E.; Mathieu, C. Immunoregulation by 1,25-dihydroxyvitamin d3: Basic concepts. J. Steroid Biochem. Mol. Biol. 2005, 97, 93–101. [Google Scholar] [CrossRef] [PubMed]
- D'Ambrosio, D.; Cippitelli, M.; Cocciolo, M.G.; Mazzeo, D.; Di Lucia, P.; Lang, R.; Sinigaglia, F.; Panina-Bordignon, P. Inhibition of il-12 production by 1,25-dihydroxyvitamin d3. Involvement of nf-kappab downregulation in transcriptional repression of the p40 gene. J. Clin. Investig. 1998, 101, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Pittas, A.G.; Joseph, N.A.; Greenberg, A.S. Adipocytokines and insulin resistance. J. Clin Endocrinol. Metab. 2004, 89, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, A.; Suarez-Pinzon, W.L.; Sooy, K.; Strynadka, K.; Christakos, S. Expression of calbindin-d(28k) in a pancreatic islet beta-cell line protects against cytokine-induced apoptosis and necrosis. Endocrinology 2001, 142, 3649–3655. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, S.; Norman, A.W. Pancreatic vitamin D-dependent calcium binding protein: Biochemical properties and response to vitamin D. Arch. Biochem. Biophys. 1984, 233, 228–236. [Google Scholar] [CrossRef]
- Radaelli, T.; Varastehpour, A.; Catalano, P.; Hauguel-de Mouzon, S. Gestational diabetes induces placental genes for chronic stress and inflammatory pathways. Diabetes 2003, 52, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Herrero, L.; Naaz, A. Obesity, inflammation, and insulin resistance. Gastroenterology 2007, 132, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wong, S.L.; Lau, C.W.; Lee, H.K.; Ng, C.F.; Zhang, L.; Yao, X.; Chen, Z.Y.; Vanhoutte, P.M.; Huang, Y. Calcitriol protects renovascular function in hypertension by down-regulating angiotensin ii type 1 receptors and reducing oxidative stress. Eur. Heart J. 2012, 33, 2980–2990. [Google Scholar] [CrossRef] [PubMed]
- Kumpel, B.M.; Manoussaka, M.S. Placental immunology and maternal alloimmune responses. Vox Sang. 2012, 102, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; Hewison, M. Vitamin D, the placenta and pregnancy. Arch. Biochem. Biophys. 2012, 523, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.J.; Hong, S.C.; Oh, M.J.; Kim, H.J. Vitamin D deficiency in gestational diabetes mellitus and the role of the placenta. Am. J. Obstet. Gynecol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Ben-Haroush, A.; Yogev, Y.; Hod, M. Epidemiology of gestational diabetes mellitus and its association with type 2 diabetes. Diabet. Med. 2004, 21, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Berger, D.K.; Chamany, S. Recurrence of gestational diabetes mellitus: A systematic review. Diabetes Care 2007, 30, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bao, W.; Rong, Y.; Yang, H.; Bowers, K.; Yeung, E.; Kiely, M. Genetic variants and the risk of gestational diabetes mellitus: A systematic review. Hum. Reprod. Update 2013, 19, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Aslani, S.; Hossein-Nezhad, A.; Mirzaei, K.; Maghbooli, Z.; Afshar, A.N.; Karimi, F. Vdr foki polymorphism and its potential role in the pathogenesis of gestational diabetes mellitus and its complications. Gynecol. Endocrinol. 2011, 27, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Rahmannezhad, G.; Mashayekhi, F.J.; Goodarzi, M.T.; Rezvanfar, M.R.; Sadeghi, A. Association between vitamin D receptor apai and taqi gene polymorphisms and gestational diabetes mellitus in an iranian pregnant women population. Gene 2016, 581, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, Y.; Yamada, S.; Yanagawa, T.; Maruyama, T.; Suzuki, R.; Niino, M.; Fukazawa, T.; Kasuga, A.; Hirose, H.; Matsubara, K.; et al. Vitamin D receptor gene polymorphism affects onset pattern of type 1 diabetes. J. Clin. Endocrinol. Metab. 2003, 88, 3137–3140. [Google Scholar] [CrossRef] [PubMed]
- Tizaoui, K.; Kaabachi, W.; Hamzaoui, A.; Hamzaoui, K. Contribution of vdr polymorphisms to type 1 diabetes susceptibility: Systematic review of case-control studies and meta-analysis. J. Steroid Biochem. Mol. Biol. 2014, 143, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Barrett-Connor, E. Association between vitamin D receptor polymorphism and type 2 diabetes or metabolic syndrome in community-dwelling older adults: The rancho bernardo study. Metabolism 2002, 51, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Chagoya, A.; Vazquez-Cardenas, P.; Moreno-Macias, H.; Tapia-Maruri, L.; Rodriguez-Guillen, R.; Lopez-Vite, E.; Garcia-Escalante, G.; Escobedo-Aguirre, F.; Parra-Covarrubias, A.; Cordero-Brieno, R.; et al. Genetic determinants for gestational diabetes mellitus and related metabolic traits in mexican women. PLoS ONE 2015, 10, e0126408. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Li, Q.; Gao, S. Meta-analysis of the relationship between common type 2 diabetes risk gene variants with gestational diabetes mellitus. PLoS ONE 2012, 7, e45882. [Google Scholar] [CrossRef] [PubMed]
- Bid, H.K.; Konwar, R.; Aggarwal, C.G.; Gautam, S.; Saxena, M.; Nayak, V.L.; Banerjee, M. Vitamin D receptor (foki, bsmi and taqi) gene polymorphisms and type 2 diabetes mellitus: A north indian study. Indian J. Med. Sci. 2009, 63, 187–194. [Google Scholar] [PubMed]
- Sahin, O.A.; Goksen, D.; Ozpinar, A.; Serdar, M.; Onay, H. Association of vitamin D receptor polymorphisms and type 1 diabetes susceptibility in children: A meta-analysis. Endocr. Connect. 2017, 6, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Cui, L.L.; Li, X.; Wang, C.J.; Ba, Y.; Wang, L.; Li, J.; Li, C.; Dai, L.P.; Li, W.J. The genetic polymorphisms in vitamin D receptor and the risk of type 2 diabetes mellitus: An updated meta-analysis. Asia Pac. J. Clin. Nutr. 2016, 25, 614–624. [Google Scholar] [PubMed]
- El-Beshbishy, H.A.; Tawfeek, M.A.; Taha, I.M.; FadulElahi, T.; Shaheen, A.Y.; Bardi, F.A.; Sultan, I.I. Association of vitamin D receptor gene bsmi (a > g) and foki (c > t) polymorphism in gestational diabetes among saudi women. Pak. J. Med. Sci. 2015, 31, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Wen, J.; Liu, G.; Liu, H.; Fu, Z.; Zhou, J.; Zhu, Y.; Liu, Y.; Guo, X.; Xu, J. Genetic variants in vitamin D signaling pathways and risk of gestational diabetes mellitus. Oncotarget 2016, 7, 67788–67795. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine infection and preterm delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG 2006, 113 (Suppl. 3), 17–42. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Espinoza, J.; Goncalves, L.F.; Kusanovic, J.P.; Friel, L.A.; Nien, J.K. Inflammation in preterm and term labour and delivery. Semin. Fetal Neonatal Med. 2006, 11, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Martineau, A.R.; Wilkinson, R.J.; Wilkinson, K.A.; Newton, S.M.; Kampmann, B.; Hall, B.M.; Packe, G.E.; Davidson, R.N.; Eldridge, S.M.; Maunsell, Z.J.; et al. A single dose of vitamin D enhances immunity to mycobacteria. Am. J. Respir. Crit. Care Med. 2007, 176, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.L.; McNeil, R.B.; Johnson, D.D.; Hulsey, T.C.; Ebeling, M.; Robinson, C.; Hamilton, S.A.; Hollis, B.W. Health characteristics and outcomes of two randomized vitamin D supplementation trials during pregnancy: A combined analysis. J. Steroid Biochem. Mol. Biol. 2013, 136, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Thorp, J.M.; Camargo, C.A.; McGee, P.L.; Harper, M.; Klebanoff, M.A.; Sorokin, Y.; Varner, M.W.; Wapner, R.J.; Caritis, S.N.; Iams, J.D.; et al. Vitamin D status and recurrent preterm birth: A nested case-control study in high-risk women. BJOG 2012, 119, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Klebanoff, M.A.; Gernand, A.D.; Platt, R.W.; Parks, W.T.; Catov, J.M.; Simhan, H.N. Maternal vitamin D status and spontaneous preterm birth by placental histology in the us collaborative perinatal project. Am. J. Epidemiol. 2014, 179, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Rouse, D.J.; Momirova, V.; Peaceman, A.M.; Sciscione, A.; Spong, C.Y.; Varner, M.W.; Malone, F.D.; Iams, J.D.; Mercer, B.M.; et al. Maternal 25-hydroxyvitamin d and preterm birth in twin gestations. Obstet. Gynecol. 2013, 122, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Wetta, L.A.; Biggio, J.R.; Cliver, S.; Abramovici, A.; Barnes, S.; Tita, A.T. Is midtrimester vitamin D status associated with spontaneous preterm birth and preeclampsia? Am. J. Perinatol. 2014, 31, 541–546. [Google Scholar] [PubMed]
- Amegah, A.K.; Klevor, M.K.; Wagner, C.L. Maternal vitamin D insufficiency and risk of adverse pregnancy and birth outcomes: A systematic review and meta-analysis of longitudinal studies. PLoS ONE 2017, 12, e0173605. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Krohn, M.A.; Simhan, H.N. Maternal vitamin D deficiency is associated with bacterial vaginosis in the first trimester of pregnancy. J. Nutr. 2009, 139, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M. Antibacterial effects of vitamin D. Nat. Rev. Endocrinol. 2011, 7, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Sehringer, B.; Schafer, W.R.; Wetzka, B.; Deppert, W.R.; Brunner-Spahr, R.; Benedek, E.; Zahradnik, H.P. Formation of proinflammatory cytokines in human term myometrium is stimulated by lipopolysaccharide but not by corticotropin-releasing hormone. J. Clin. Endocrinol. Metab. 2000, 85, 4859–4865. [Google Scholar] [CrossRef] [PubMed]
- Helmer, H.; Tretzmuller, U.; Brunbauer, M.; Kaider, A.; Husslein, P.; Knofler, M. Production of oxytocin receptor and cytokines in primary uterine smooth muscle cells cultivated under inflammatory conditions. J. Soc. Gynecol. Investig. 2002, 9, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Mazor, M.; Tartakovsky, B. Systemic administration of interleukin-1 induces preterm parturition in mice. Am. J. Obstet. Gynecol. 1991, 165, 969–971. [Google Scholar] [CrossRef]
- Buhimschi, I.A.; Buhimschi, C.S.; Weiner, C.P. Protective effect of n-acetylcysteine against fetal death and preterm labor induced by maternal inflammation. Am. J. Obstet. Gynecol. 2003, 188, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Thota, C.; Farmer, T.; Garfield, R.E.; Menon, R.; Al-Hendy, A. Vitamin D elicits anti-inflammatory response, inhibits contractile-associated proteins, and modulates toll-like receptors in human myometrial cells. Reprod. Sci. 2013, 20, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, T.; Salem, H.; Altarescu, G.; Grisaru-Granovsky, S.; Tevet, A.; Birk, R. Maternal-fetal vitamin D receptor polymorphisms significantly associated with preterm birth. Arch. Gynecol. Obstet. 2017, 296, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Baczynska-Strzecha, M.; Kalinka, J. Influence of apa1 (rs7975232), taq1 (rs731236) and bsm1 (rs154410) polymorphisms of vitamin D receptor on preterm birth risk in the polish population. Ginekol. Pol. 2016, 87, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, C.S.; Woodland, M.L.; Fudge, N.J.; Friel, J.K. The vitamin D receptor is not required for fetal mineral homeostasis or for the regulation of placental calcium transfer in mice. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E133–E144. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.L.; Buckberry, S.; Spronk, F.; Laurence, J.A.; Leemaqz, S.; O'Leary, S.; Bianco-Miotto, T.; Du, J.; Anderson, P.H.; Roberts, C.T. Vitamin D receptor gene ablation in the conceptus has limited effects on placental morphology, function and pregnancy outcome. PLoS ONE 2015, 10, e0131287. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; Ouyang, Y.; Bulut, Y.; Lagishetty, V.; Chan, S.Y.; Hollis, B.W.; Wagner, C.; Equils, O.; Hewison, M. Dietary vitamin D restriction in pregnant female mice is associated with maternal hypertension and altered placental and fetal development. Endocrinology 2013, 154, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Placental oxidative stress: From miscarriage to preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Polymorphism | Pregnancy Complication |
|---|---|
| FokI | Preterm birth |
| Gestational diabetes | |
| Influence on offspring size | |
| ApaI | Preterm birth |
| Gestational diabetes | |
| Influence on Birth weight | |
| TaqI | Gestational diabetes |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knabl, J.; Vattai, A.; Ye, Y.; Jueckstock, J.; Hutter, S.; Kainer, F.; Mahner, S.; Jeschke, U. Role of Placental VDR Expression and Function in Common Late Pregnancy Disorders. Int. J. Mol. Sci. 2017, 18, 2340. https://doi.org/10.3390/ijms18112340
Knabl J, Vattai A, Ye Y, Jueckstock J, Hutter S, Kainer F, Mahner S, Jeschke U. Role of Placental VDR Expression and Function in Common Late Pregnancy Disorders. International Journal of Molecular Sciences. 2017; 18(11):2340. https://doi.org/10.3390/ijms18112340
Chicago/Turabian StyleKnabl, Julia, Aurelia Vattai, Yao Ye, Julia Jueckstock, Stefan Hutter, Franz Kainer, Sven Mahner, and Udo Jeschke. 2017. "Role of Placental VDR Expression and Function in Common Late Pregnancy Disorders" International Journal of Molecular Sciences 18, no. 11: 2340. https://doi.org/10.3390/ijms18112340