Inhibition of Starvation-Triggered Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in ARPE-19 Cells by Taurine through Modulating the Expression of Calpain-1 and Calpain-2


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
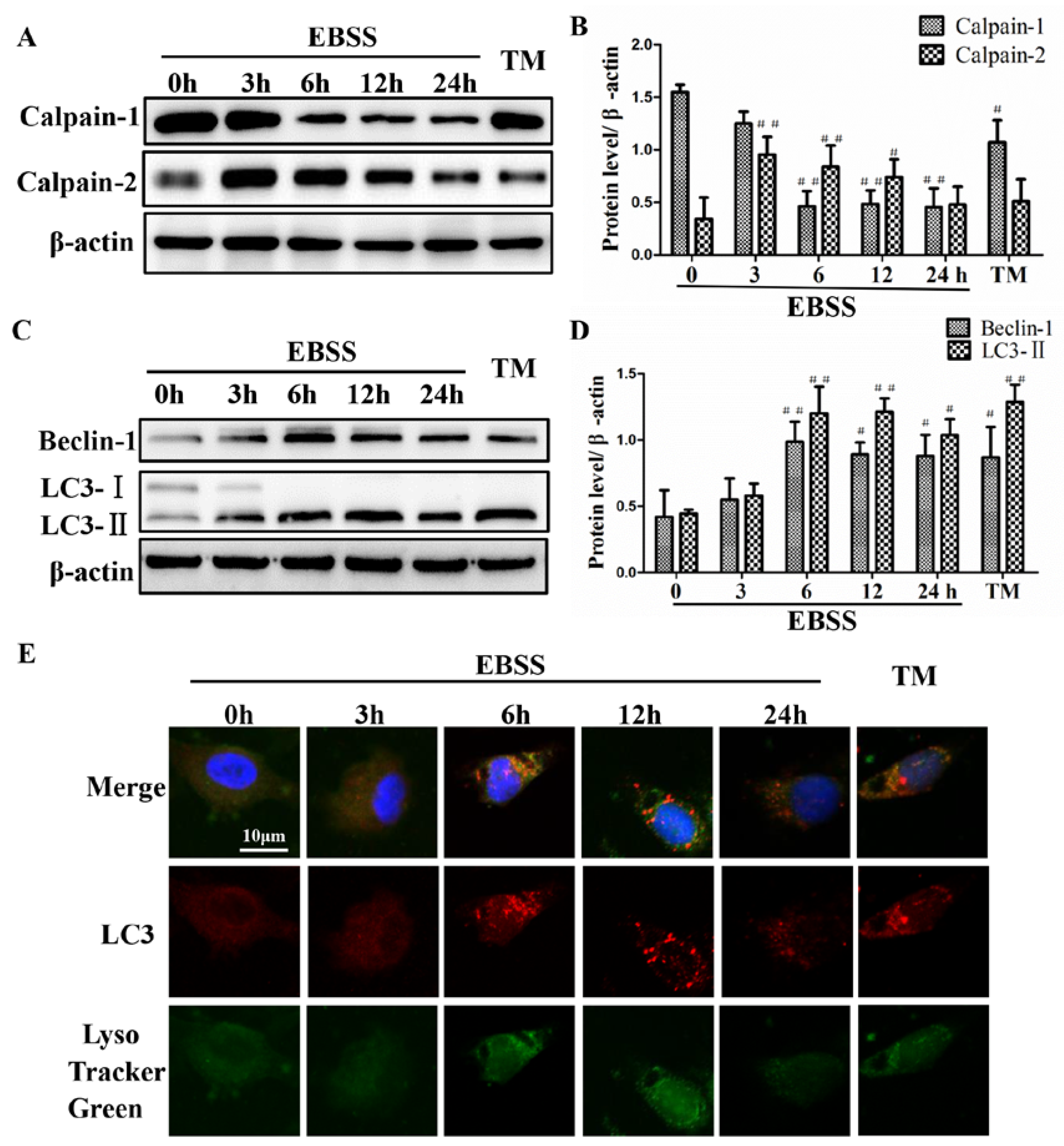
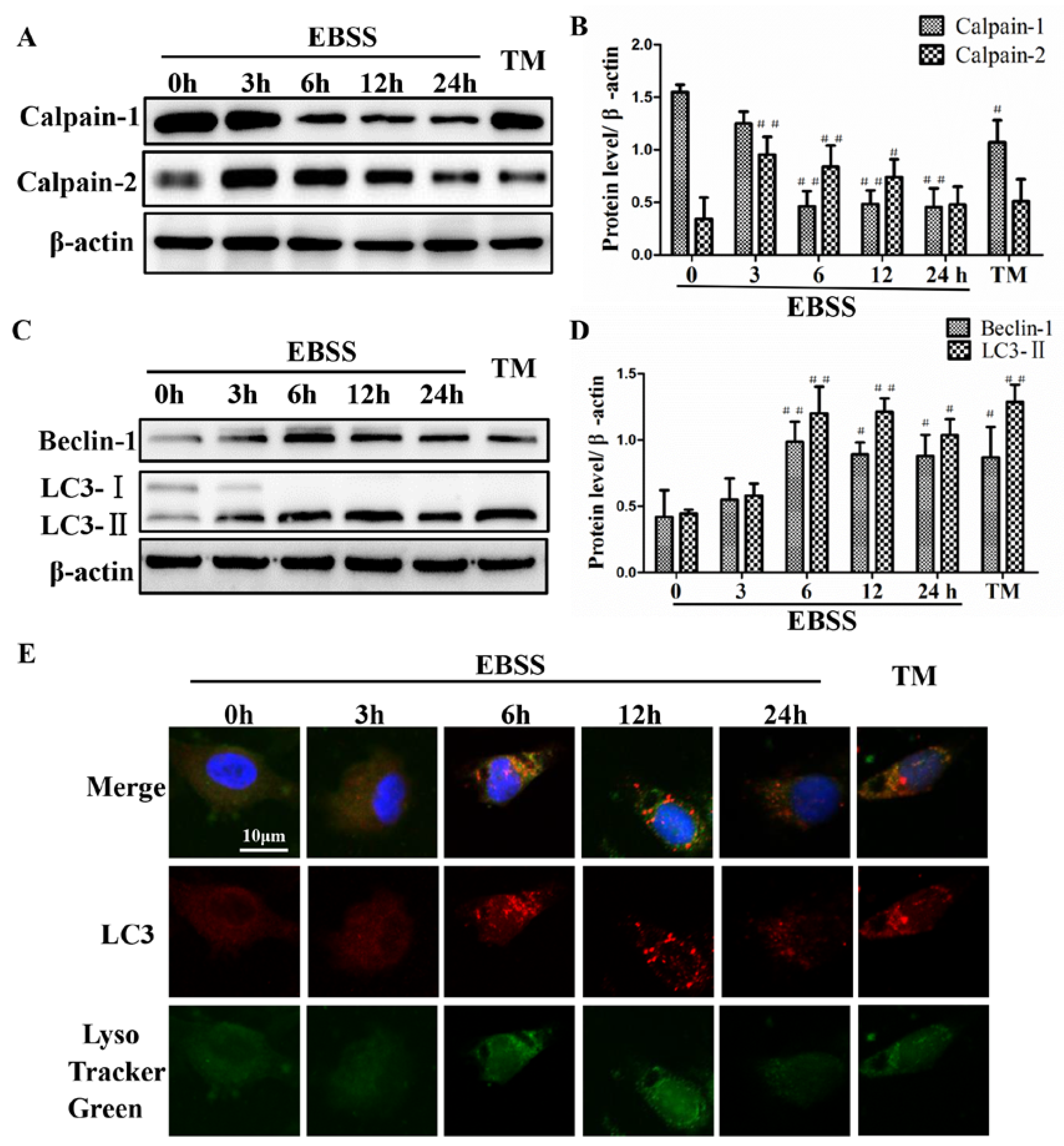
2.1. Effects of EBSS Treatment for Different Durations on Expression of Calpains and Autophagy-Relative Proteins in ARPE-19 Cells
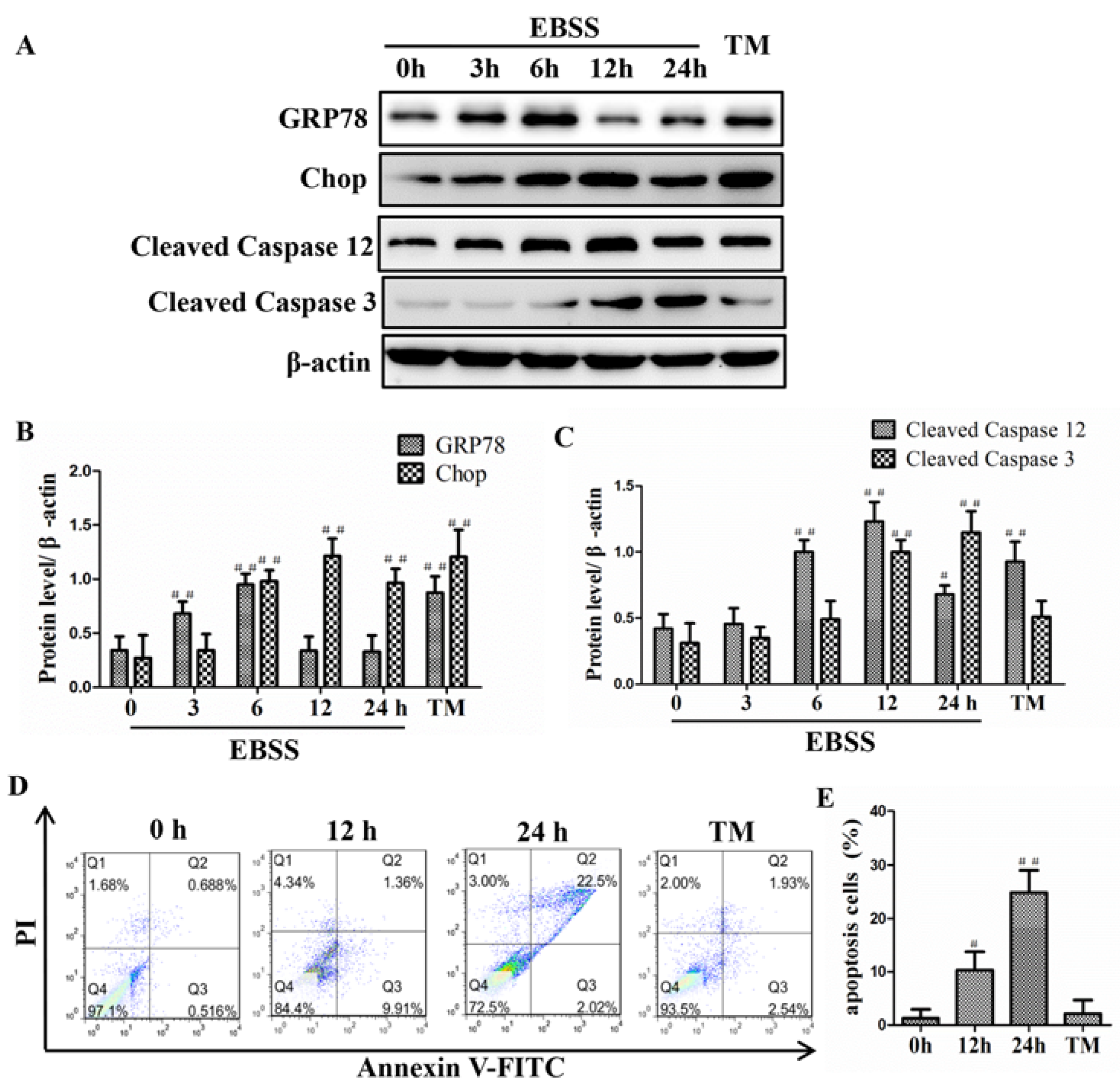
2.2. Effects of EBSS Treatment for Different Durations on Expression of ER Stress-Relative and Apoptosis-Relative Proteins in ARPE-19 Cells
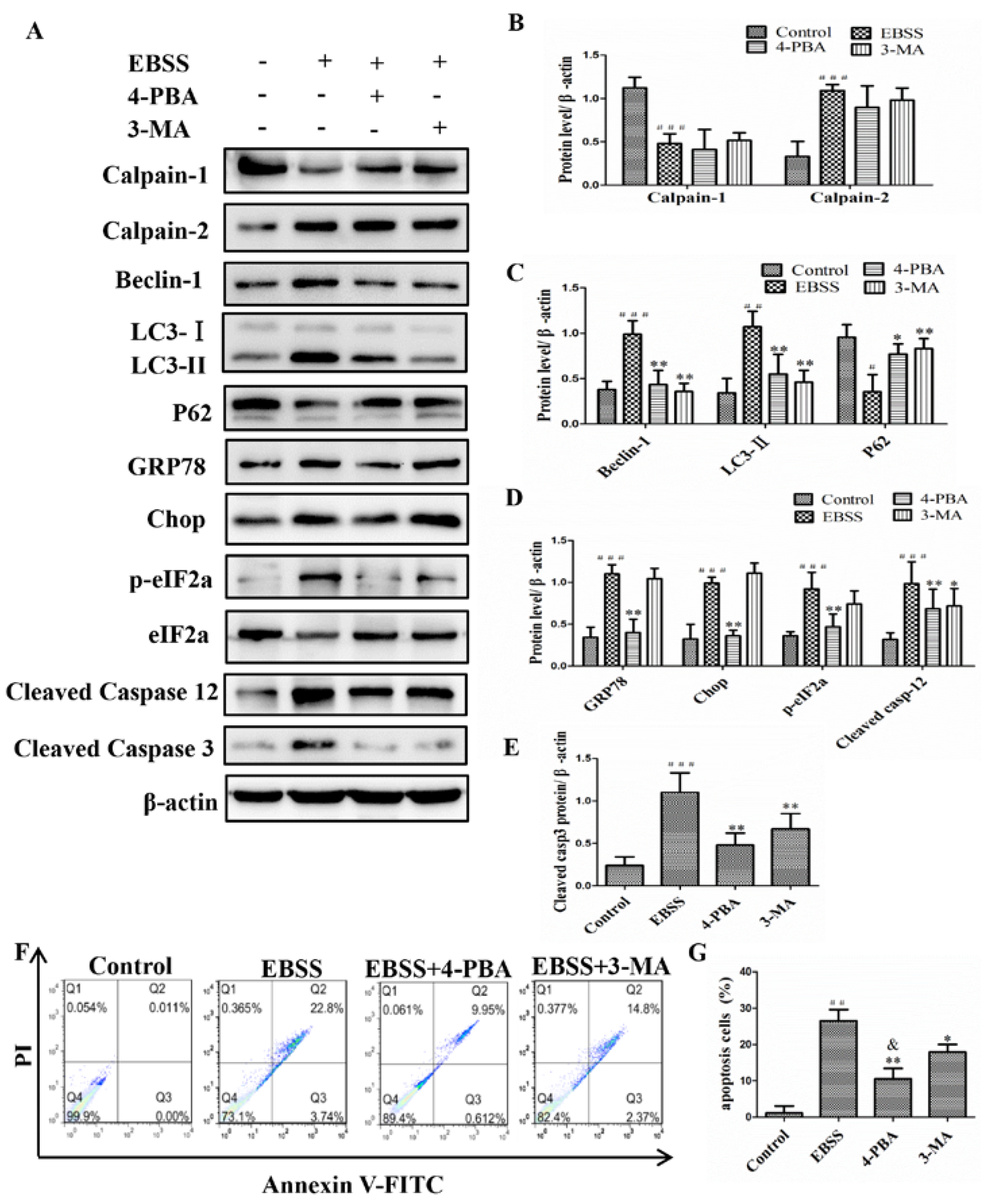
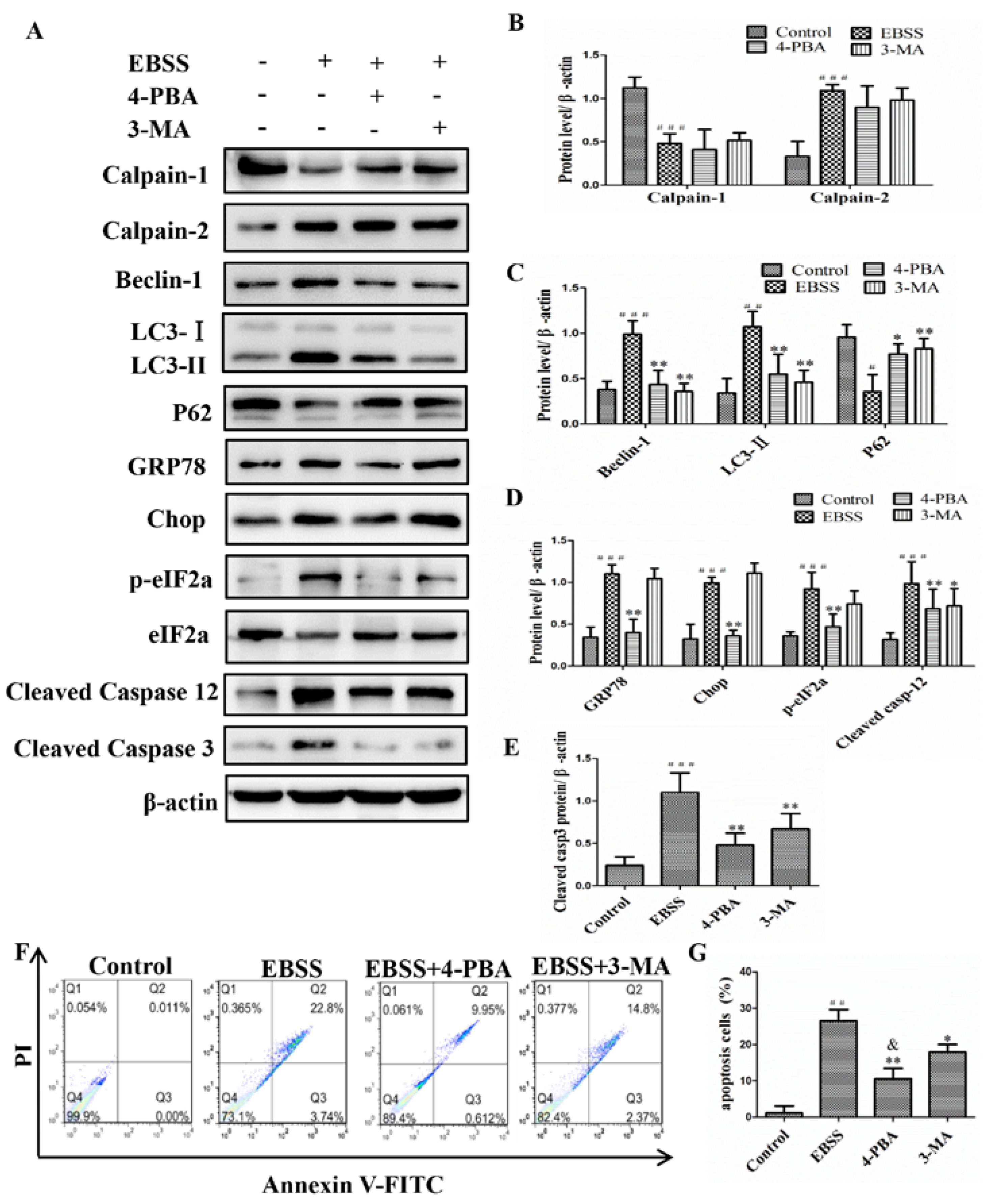
2.3. Inhibiting ER Stress and Autophagy Attenuated EBSS-Induced Apoptosis in ARPE-19 Cells
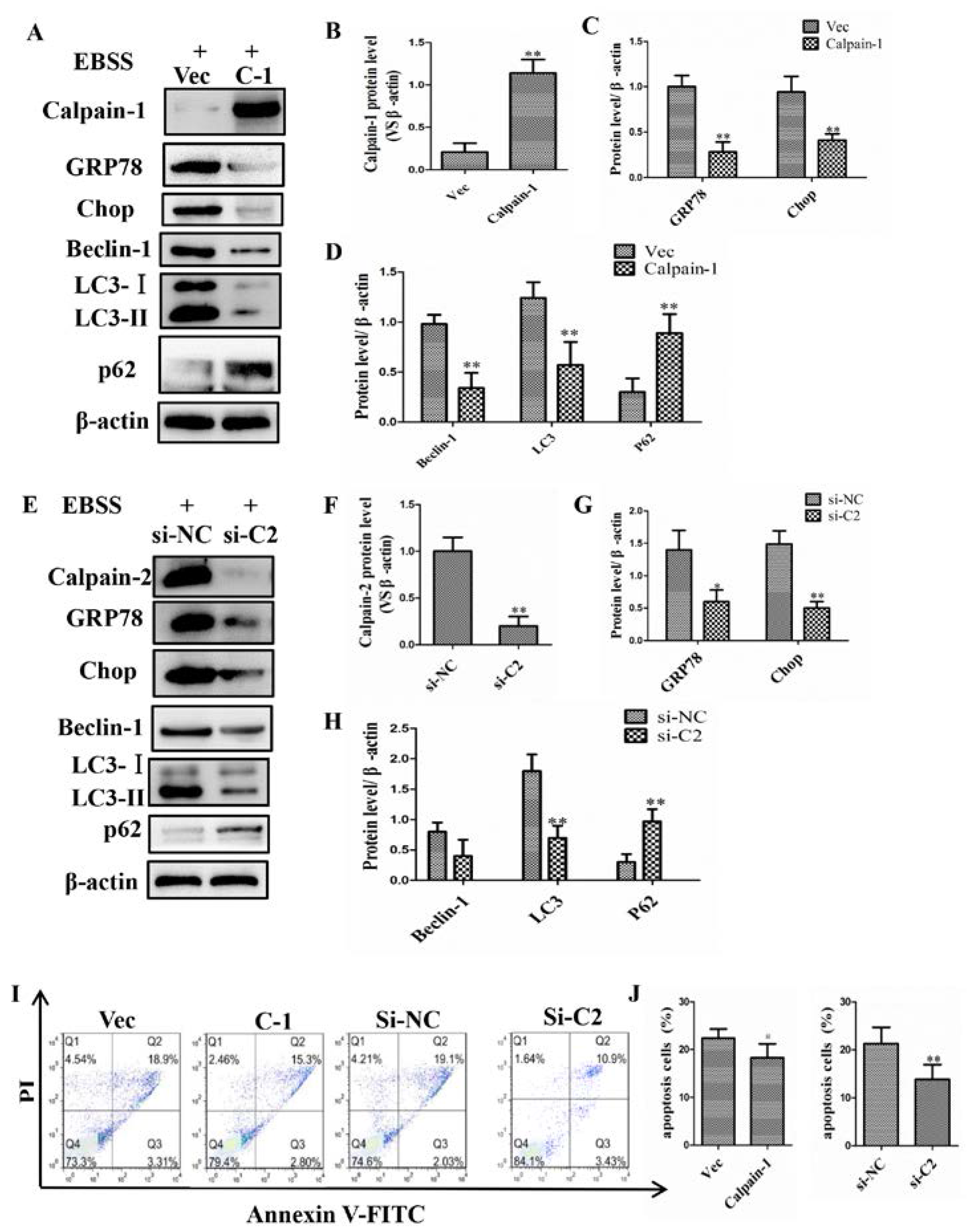
2.4. Calpain-1 and Calpain-2 Played Key Roles in EBSS-Induced Cell Injuries
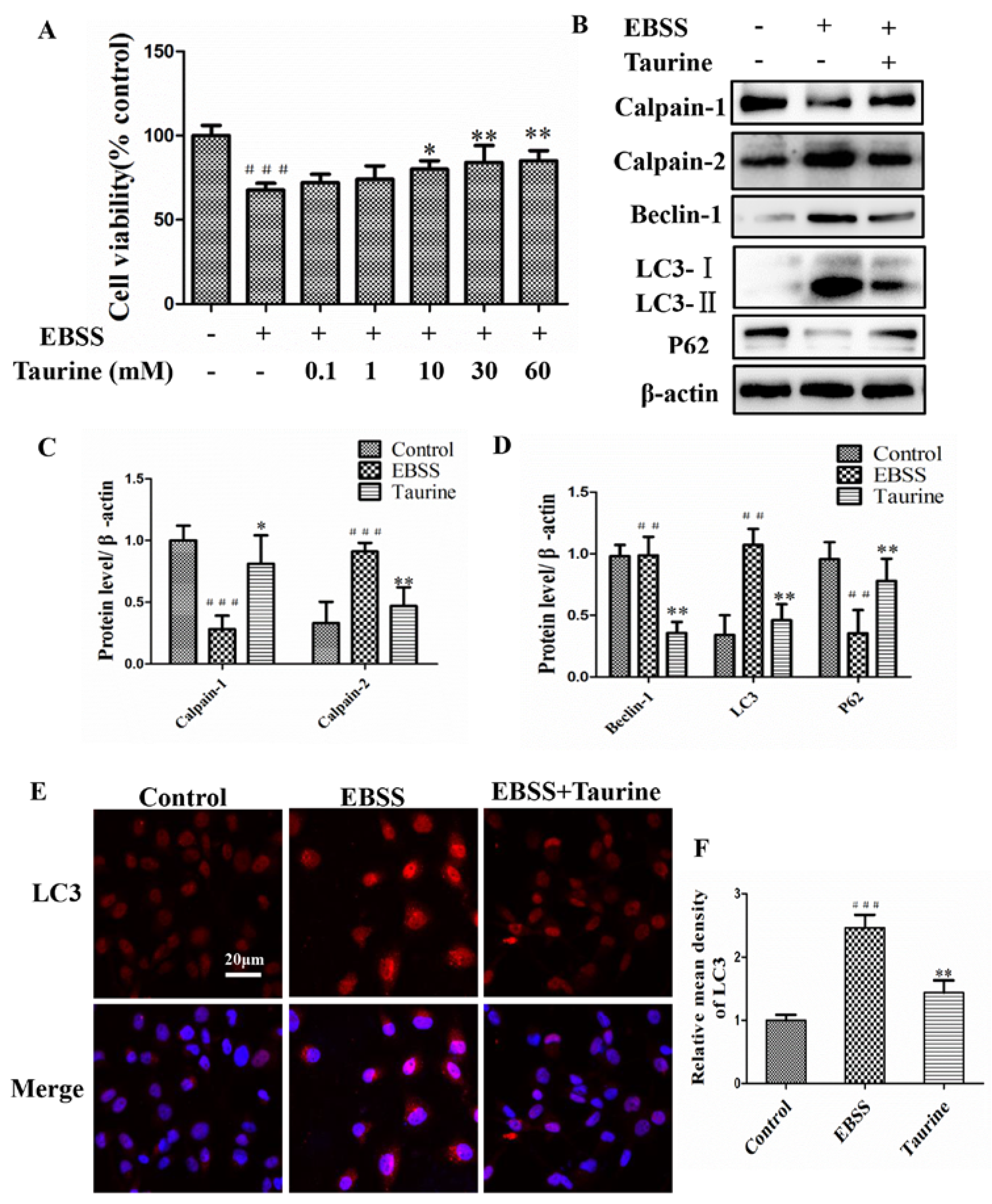
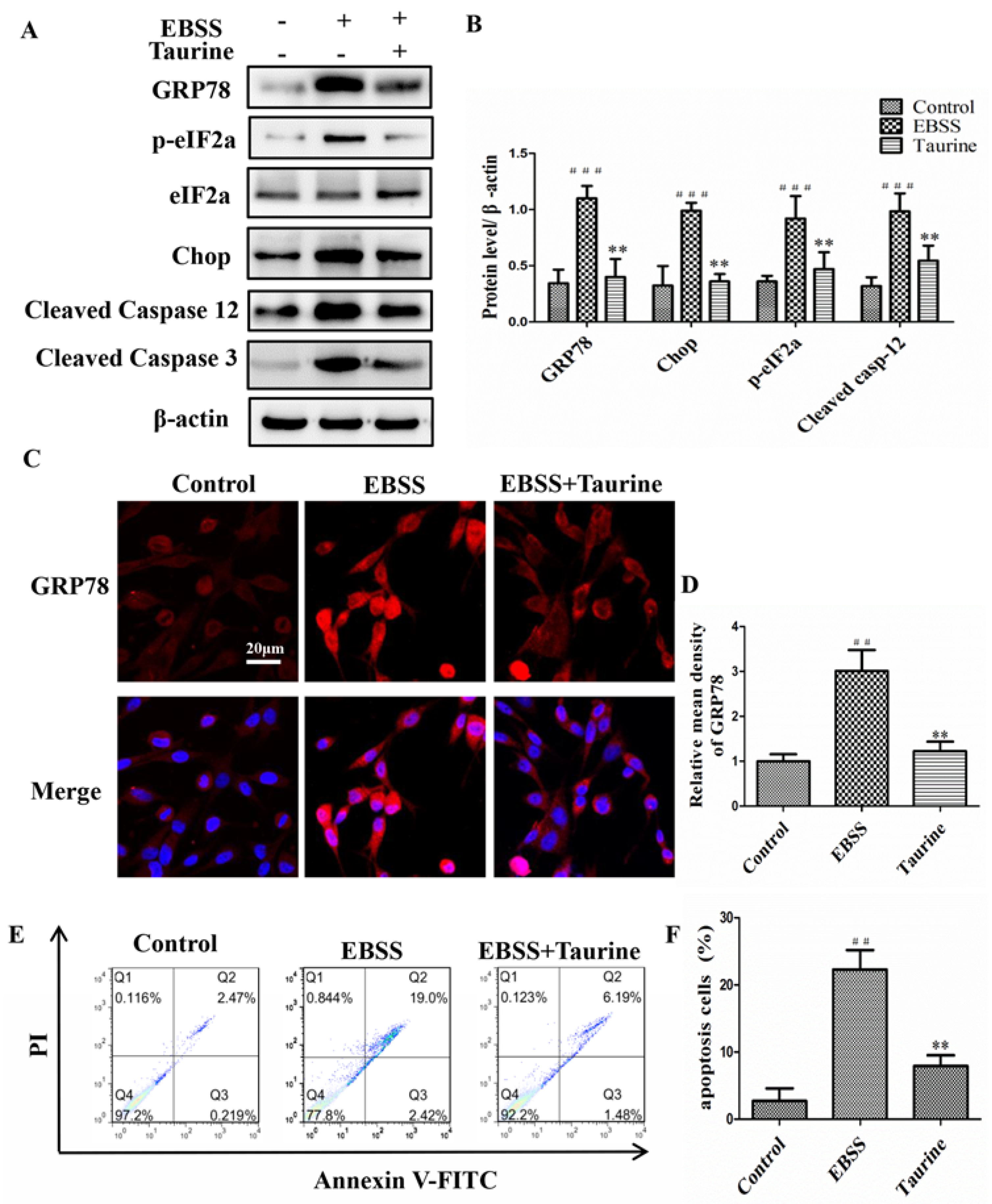
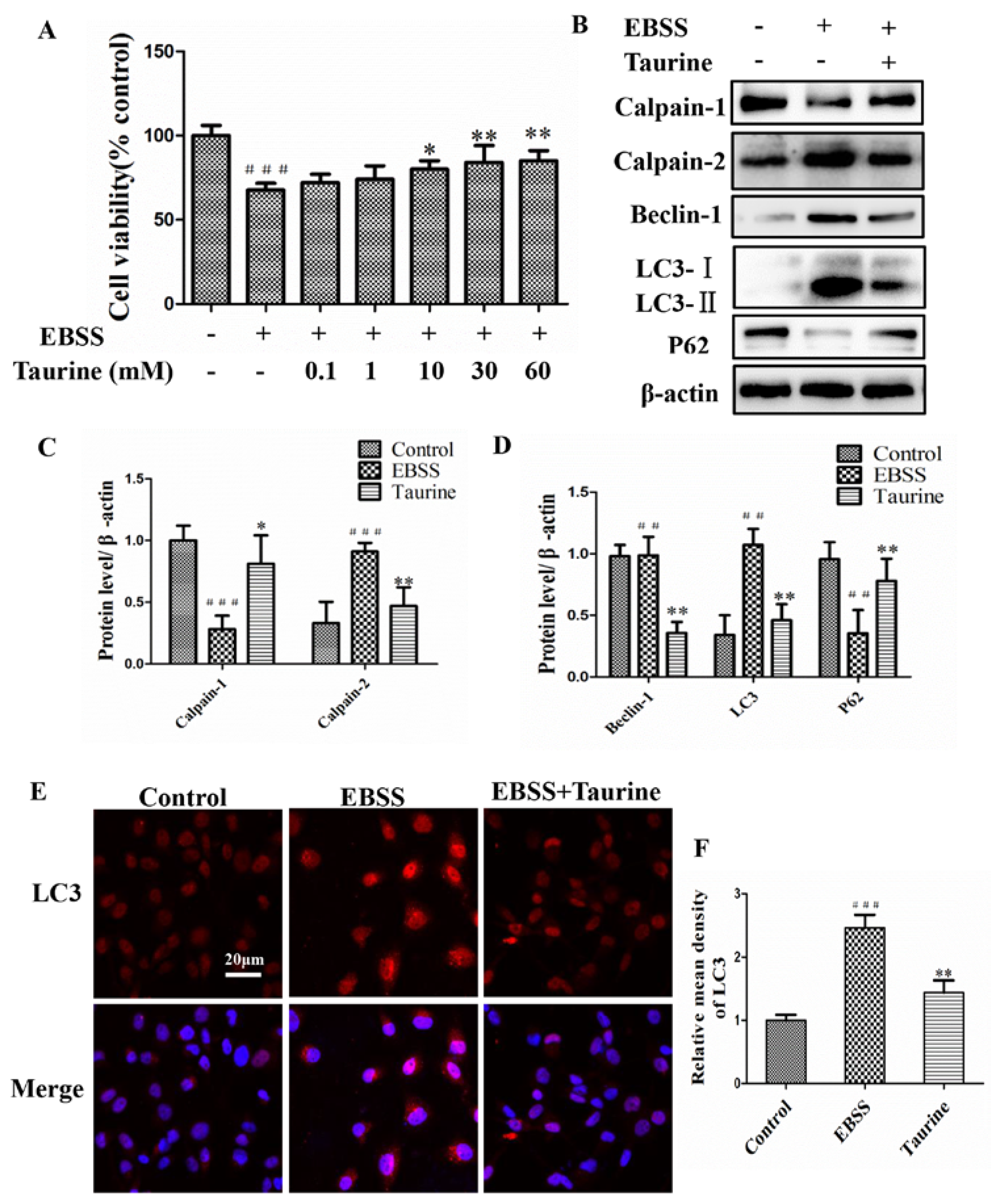
2.5. Effects of Taurine Treatment on Expression of Calpains and Autophagy-Relative Proteins Induced by EBSS in ARPE-19 Cells
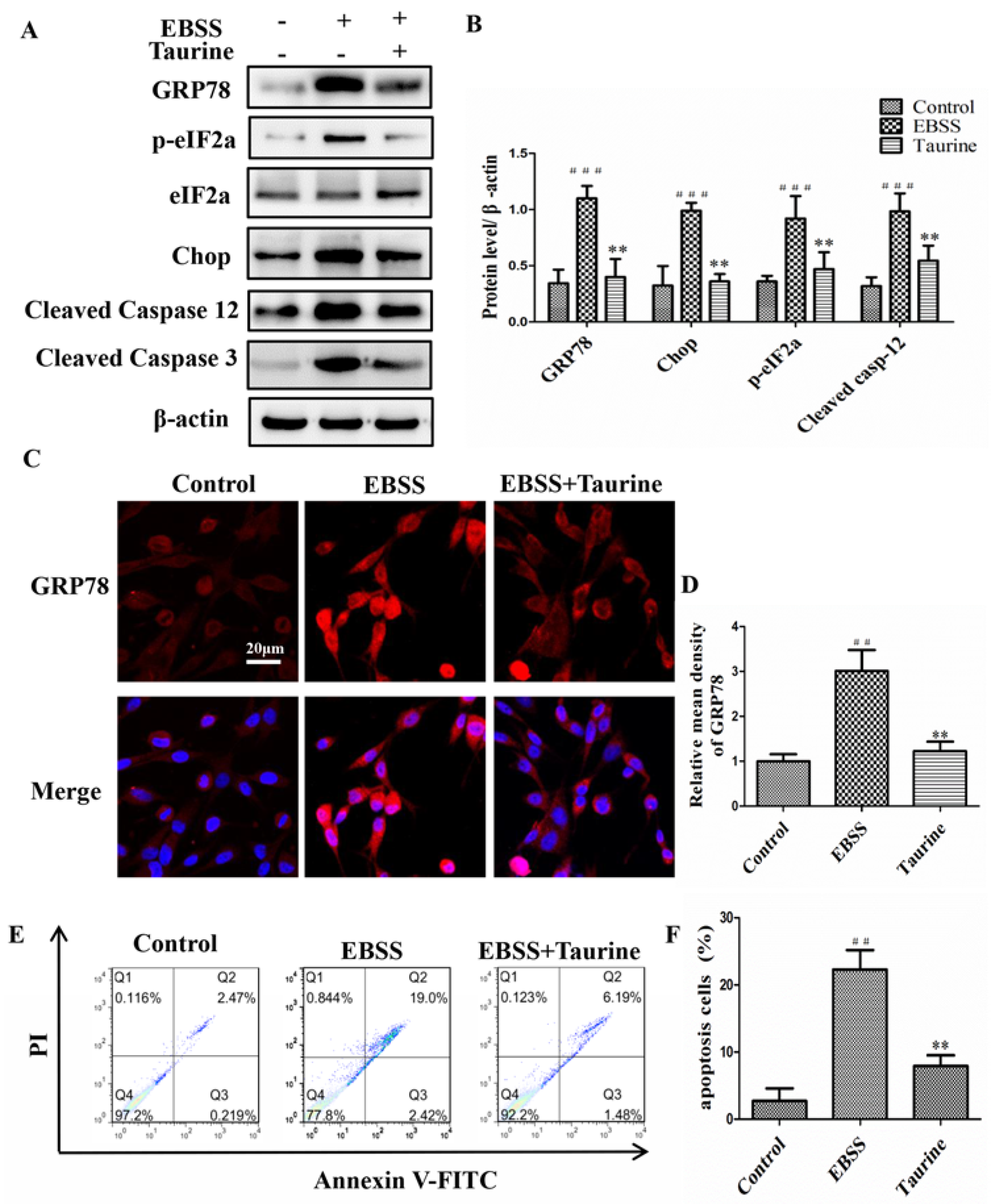
2.6. Effects of Taurine Treatment on the Expression of ER Stress-Relative and Apoptosis-Relative Proteins Induced by EBSS in ARPE-19 Cells
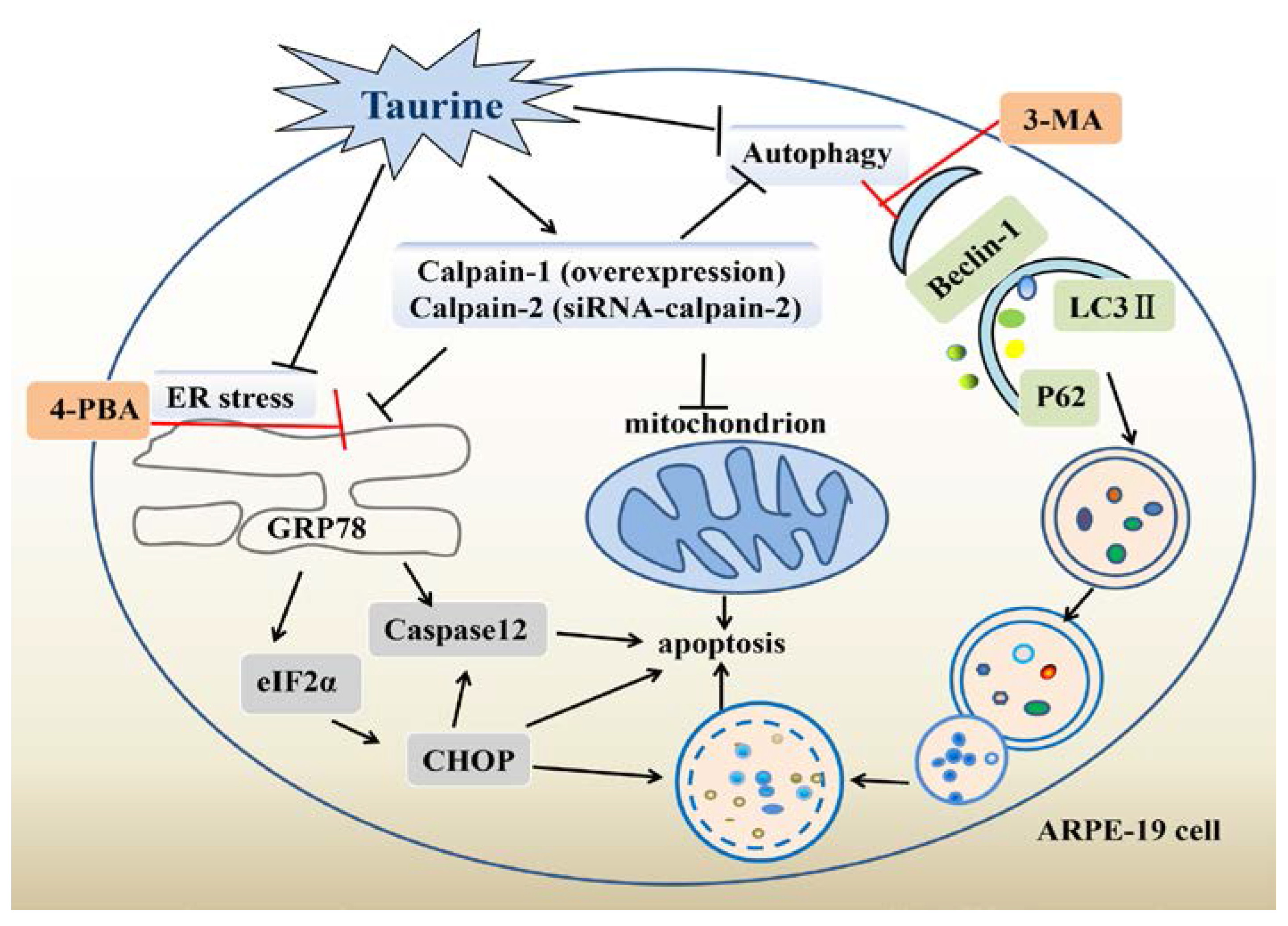
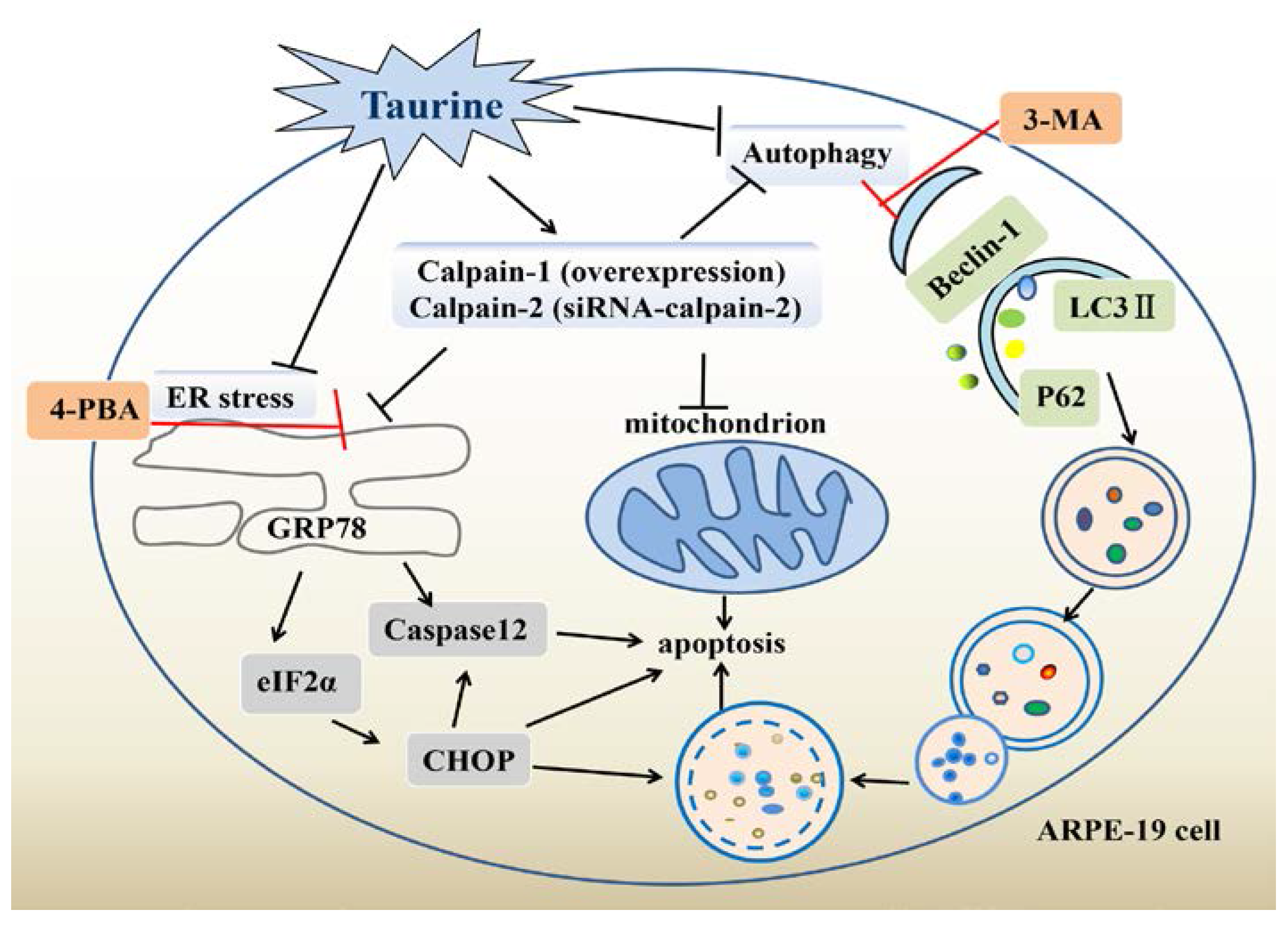
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Transfection Experiments
4.3. Cell Viability Assay
4.4. Double Immunofluorescence Staining
4.5. Annexin V/PI Double Staining
4.6. Western Blotting
4.7. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bardak, H.; Uguz, A.C.; Bardak, Y. Protective effects of melatonin and memantine in human retinal pigment epithelium (ARPE-19) cells against 2-ethylpyridine-induced oxidative stress: Implications for age-related macular degeneration. Cutan. Ocul. Toxicol. 2017, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cootjans, K.; Vanhaecke, J.; Dezillie, M.; Barth, J.; Pottel, H.; Stockmans, F. Joint Survival Analysis and Clinical Outcome of Total Joint Arthroplasties With the ARPE Implant in the Treatment of Trapeziometacarpal Osteoarthritis with a Minimal Follow-Up of 5 Years. J. Hand Surg. 2017, 42, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zhang, Z.; Wang, X.; Li, R.; Hou, W.; Bi, W.; Zhang, X. Inhibition of autophagy induces IL-1beta release from ARPE-19 cells via ROS mediated NLRP3 inflammasome activation under high glucose stress. Biochem. Biophys. Res. Commun. 2015, 463, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Vainio, I.; Abu Khamidakh, A.; Paci, M.; Skottman, H.; Juuti-Uusitalo, K.; Hyttinen, J.; Nymark, S. Computational Model of Ca2+ Wave Propagation in Human Retinal Pigment Epithelial ARPE-19 Cells. PLoS ONE 2015, 10, e0128434. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xing, Y.; Chen, C.; Chen, Z.; Qian, Z. Advanced glycation end-product (AGE) induces apoptosis in human retinal ARPE-19 cells via promoting mitochondrial dysfunction and activating the Fas-FasL signaling. Biosci. Biotechnol. Biochem. 2016, 80, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.G.; Zhang, L.; Chen, G.; Zhang, T.; Liu, J.; Jin, M.; Ma, X.; Ma, D.; Yuan, J. Control of basal autophagy by calpain1 mediated cleavage of ATG5. Autophagy 2010, 6, 61–66. [Google Scholar] [CrossRef] [PubMed]
- McKernan, D.P.; Guerin, M.B.; O’Brien, C.J.; Cotter, T.G. A key role for calpains in retinal ganglion cell death. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5420–5430. [Google Scholar] [CrossRef] [PubMed]
- Komarova, Y.; Malik, A.B. Regulation of endothelial permeability via paracellular and transcellular transport pathways. Annu. Rev. Physiol. 2010, 72, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhang, J.; Sun, P.; Yao, C.; Su, C.; Sui, T.; Huang, H.; Cao, X.; Ge, Y. EIF2alpha and caspase-12 activation are involved in oxygen-glucose-serum deprivation/restoration-induced apoptosis of spinal cord astrocytes. Neurosci. Lett. 2010, 478, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; DeRoo, E.P.; Stecyk, C.; Wolsey, M.; Szuchnicki, M.; Hagos, E.G. Impaired autophagy in mouse embryonic fibroblasts null for Kruppel-like Factor 4 promotes DNA damage and increases apoptosis upon serum starvation. Mol. Cancer 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Parra, M.A.; Walia, M.; Sankar, M.; Gronemeyer, H. Dissecting the retinoid-induced differentiation of F9 embryonal stem cells by integrative genomics. Mol. Syst. Biol. 2011, 7, 538. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.S.; Du, J.; Fan, Y.J.; Liu, F.J.; Cao, L.L.; Liang, N.; Xu, D.G.; Zhang, J.D. Activation of endoplasmic reticulum stress promotes autophagy and apoptosis and reverses chemoresistance of human small cell lung cancer cells by inhibiting the PI3K/AKT/mTOR signaling pathway. Oncotarget 2016, 7, 76827–76839. [Google Scholar] [CrossRef] [PubMed]
- Luan, Q.; Jin, L.; Jiang, C.C.; Tay, K.H.; Lai, F.; Liu, X.Y.; Liu, Y.L.; Guo, S.T.; Li, C.Y.; Yan, X.G.; et al. RIPK1 regulates survival of human melanoma cells upon endoplasmic reticulum stress through autophagy. Autophagy 2015, 11, 975–994. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.P.; Chien, J.Y.; Tsai, R.K. Ethambutol induces impaired autophagic flux and apoptosis in the rat retina. Dis. Models Mech. 2015, 8, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, S.K.; Dash, R.; Das, S.K.; Azab, B.; Su, Z.Z.; Lee, S.G.; Grant, S.; Yacoub, A.; Dent, P.; Curiel, D.T.; et al. Mechanism of autophagy to apoptosis switch triggered in prostate cancer cells by antitumor cytokine melanoma differentiation-associated gene 7/interleukin-24. Cancer Res. 2010, 70, 3667–3676. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Park, K.I.; Kim, S.H.; Yu, S.N.; Park, S.G.; Kim, Y.W.; Seo, Y.K.; Ma, J.Y.; Ahn, S.C. Inhibition of Autophagy Promotes Salinomycin-Induced Apoptosis via Reactive Oxygen Species-Mediated PI3K/AKT/mTOR and ERK/p38 MAPK-Dependent Signaling in Human Prostate Cancer Cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Tzekov, R.; Li, H.; McDowell, J.H.; Gao, G.; Smith, W.C.; Tang, S.; Kaushal, S. Inhibition or Stimulation of Autophagy Affects Early Formation of Lipofuscin-Like Autofluorescence in the Retinal Pigment Epithelium Cell. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Ryu, E.S.; Oh, E.S.; Kang, D.H. Endoplasmic reticulum stress as a novel target to ameliorate epithelial-to-mesenchymal transition and apoptosis of human peritoneal mesothelial cells. Lab. Investig. 2015, 95, 1157–1173. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Shan, S.; Hu, T.; Chen, H.; Tian, J.; Ren, A.; Zhou, X.; Yuan, W.; Lin, L. Salusins protect myocardium against ischemic injury by alleviating endoplasmic reticulum stress. Sci. China Life Sci. 2012, 55, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Du, L. PERK pathway is involved in oxygen-glucose-serum deprivation-induced NF-kB activation via ROS generation in spinal cord astrocytes. Biochem. Biophys. Res. Commun. 2015, 467, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Zhu, H.Z.; Li, S.W.; Yang, J.M.; Xiao, Y.; Kang, Q.R.; Li, C.Y.; Zhao, Y.S.; Zeng, Y.; Li, Y.; et al. Crude Saponins of Panax notoginseng Have Neuroprotective Effects To Inhibit Palmitate-Triggered Endoplasmic Reticulum Stress-Associated Apoptosis and Loss of Postsynaptic Proteins in Staurosporine Differentiated RGC-5 Retinal Ganglion Cells. J. Agric. Food Chem. 2016, 64, 1528–1539. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.F.; Liu, X.; Gao, M.; Zhang, Y.N.; Liu, J. Endoplasmic reticulum stress induces autophagy and apoptosis while inhibiting proliferation and drug resistance in multiple myeloma through the PI3K/Akt/mTOR signaling pathway. Oncotarget 2017. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Berliocchi, L.; Adornetto, A.; Varano, G.P.; Cavaliere, F.; Nucci, C.; Rotiroti, D.; Morrone, L.A.; Bagetta, G.; Corasaniti, M.T. Calpain-mediated cleavage of Beclin-1 and autophagy deregulation following retinal ischemic injury in vivo. Cell Death Dis. 2011, 2, e144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Briz, V.; Chishti, A.; Bi, X.; Baudry, M. Distinct roles for mu-calpain and m-calpain in synaptic NMDAR-mediated neuroprotection and extrasynaptic NMDAR-mediated neurodegeneration. J. Neurosci. 2013, 33, 18880–18892. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cui, W.; He, S.M.; Duan, Y.H.; Heng, L.J.; Wang, L.; Gao, G.D. Human U87 astrocytoma cell invasion induced by interaction of betaig-h3 with integrin alpha5beta1 involves calpain-2. PLoS ONE 2012, 7, e37297. [Google Scholar] [CrossRef]
- Tang, L.; Pei, H.; Yang, Y.; Wang, X.; Wang, T.; Gao, E.; Li, D.; Yang, Y.; Yang, D. The inhibition of calpains ameliorates vascular restenosis through MMP2/TGF-beta1 pathway. Sci. Rep. 2016, 6, 29975. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lopez, D.; Davey, P.G.; Cameron, D.J.; Nguyen, K.; Tran, J.; Marquez, E.; Liu, Y.; Bi, X.; Baudry, M. Calpain-1 and calpain-2 play opposite roles in retinal ganglion cell degeneration induced by retinal ischemia/reperfusion injury. Neurobiol. Dis. 2016, 93, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Bei, L.; Hjort, E.E.; Eklund, E.A. Decreased Calpain activity in chronic myeloid leukemia impairs apoptosis by increasing Survivin in myeloid progenitors and xiap1 in differentiating granulocytes. Oncotarget 2017. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Ma, J.; Wang, X.; Yang, W.; Zhang, J.; Ji, Q. Free fatty acid induces endoplasmic reticulum stress and apoptosis of beta-cells by Ca2+/calpain-2 pathways. PLoS ONE 2013, 8, e59921. [Google Scholar] [CrossRef]
- Bucolo, C.; Fidilio, A.; Platania, C.B.M.; Geraci, F.; Lazzara, F.; Drago, F. Antioxidant and osmoprotecting activity of taurine in dry eye models. J. Ocul. Pharmacol. Ther. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, L.; Zhou, Y.; Lu, X.; Wang, Z.; Wang, J.; Li, W. Taurine ameliorated homocysteine-induced H9C2 cardiomyocyte apoptosis by modulating endoplasmic reticulum stress. Apoptosis 2017, 22, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, Q.; Wang, J.; Liu, F.; Mi, M.; Xu, H.; Chen, F.; Zeng, K. Taurine protects transformed rat retinal ganglion cells from hypoxia-induced apoptosis by preventing mitochondrial dysfunction. Brain Res. 2009, 1279, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, Z.; Chen, B.; Bu, Q.; Lu, W.; Deng, Y.; Zhu, R.; Shao, X.; Hou, J.; Zhao, J.; et al. Taurine attenuates methamphetamine-induced autophagy and apoptosis in PC12 cells through mTOR signaling pathway. Toxicol. Lett. 2012, 215, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Froger, N.; Moutsimilli, L.; Cadetti, L.; Jammoul, F.; Wang, Q.P.; Fan, Y.; Gaucher, D.; Rosolen, S.G.; Neveux, N.; Cynober, L.; et al. Taurine: The comeback of a neutraceutical in the prevention of retinal degenerations. Prog. Retin. Eye Res. 2014, 41, 44–63. [Google Scholar] [CrossRef] [PubMed]
- Messer, J.S. The cellular autophagy/apoptosis checkpoint during inflammation. Cell. Mol. Life Sci. 2017, 74, 1281–1296. [Google Scholar] [CrossRef] [PubMed]
- Au, A.K.; Aneja, R.K.; Bayir, H.; Bell, M.J.; Janesko-Feldman, K.; Kochanek, P.M.; Clark, R.S.B. Autophagy Biomarkers Beclin 1 and p62 are Increased in Cerebrospinal Fluid after Traumatic Brain Injury. Neurocrit. Care 2017, 26, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, J.; Du, Y.; Xu, Y.; Wang, Y.; Zhang, Z.; Xu, Z.; Zeng, Y.; Mao, X.; Cao, B. The Class I PI3K inhibitor S14161 induces autophagy in malignant blood cells by modulating the Beclin 1/Vps34 complex. J. Pharmacol. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Han, F.; Shi, Y.; Wen, L.; Han, D. Single-Prolonged-Stress-Induced Changes in Autophagy-Related Proteins Beclin-1, LC3, and p62 in the Medial Prefrontal Cortex of Rats with Post-traumatic Stress Disorder. J. Mol. Neurosci. 2017, 62, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, A.; Ferreira, E.; Branca, C.; Oddo, S. p62 improves AD-like pathology by increasing autophagy. Mol. Psychiatry 2017, 22, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.L.; Chen, C.H.; Hwang, C.S.; Chen, S.D.; Hwang, W.C.; Yang, D.I. Roles of p62 in BDNF-dependent autophagy suppression and neuroprotection against mitochondrial dysfunction in rat cortical neurons. J. Neurochem. 2017, 140, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, W.G.; Zhao, Y. Autophagy substrate SQSTM1/p62 regulates chromatin ubiquitination during the DNA damage response. Autophagy 2017, 13, 212–213. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, A.; Kaira, K.; Yasuda, M.; Asao, T.; Ishikawa, O. Clinical and Pathological Significance of ER Stress Marker (BiP/GRP78 and PERK) Expression in Malignant Melanoma. Pathol. Oncol. Res. 2017, 23, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Singh, N.; Sengupta, N.; Fatima, M.; Seth, P.; Mahadevan, A.; Shankar, S.K.; Bhattacharyya, A.; Basu, A. Japanese encephalitis virus induces human neural stem/progenitor cell death by elevating GRP78, PHB and hnRNPC through ER stress. Cell Death Dis. 2017, 8, e2556. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, M.; Oh, J.H. Effects of bevacizumab on endoplasmic reticulum stress in hypoxic retinal pigment epithelial cells. PLoS ONE 2017, 12, e0179048. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, C.; Lu, Y.; Liu, L.; Jiang, Y. ER stress related factor ATF6 and caspase-12 trigger apoptosis in neonatal hypoxic-ischemic encephalopathy. Int. J. Clin. Exp. Pathol. 2015, 8, 6960–6966. [Google Scholar] [PubMed]
- Lee, W.; Kim, D.H.; Boo, J.H.; Kim, Y.H.; Park, I.S.; Mook-Jung, I. ER stress-induced caspase-12 activation is inhibited by PKC in neuronal cells. Apoptosis 2005, 10, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, H.; Okamoto, H.; Yoshimura, A.; Yoshida, H. ER stress-induced apoptosis and caspase-12 activation occurs downstream of mitochondrial apoptosis involving Apaf-1. J. Cell Sci. 2006, 119, 3958–3966. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Nam, S.M.; Kim, J.H.; Das, R.; Choi, S.K.; Nguyen, T.T.; Quan, X.; Choi, S.J.; Chung, C.H.; Lee, E.Y.; et al. Palmitate induces ER calcium depletion and apoptosis in mouse podocytes subsequent to mitochondrial oxidative stress. Cell Death Dis. 2015, 6, e1976. [Google Scholar] [CrossRef] [PubMed]
- Gorlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Barateiro, A.; Vaz, A.R.; Silva, S.L.; Fernandes, A.; Brites, D. ER stress, mitochondrial dysfunction and calpain/JNK activation are involved in oligodendrocyte precursor cell death by unconjugated bilirubin. Neuromol. Med. 2012, 14, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Qi, J.; Liu, L.M.; Li, J.; Xu, H.Y.; Liang, B.; Li, B. Valsartan reduces AT1-AA-induced apoptosis through suppression oxidative stress mediated ER stress in endothelial progenitor cells. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1159–1168. [Google Scholar] [PubMed]
- Zhou, X.; Wei, Y.; Qiu, S.; Xu, Y.; Zhang, T.; Zhang, S. Propofol Decreases Endoplasmic Reticulum Stress-Mediated Apoptosis in Retinal Pigment Epithelial Cells. PLoS ONE 2016, 11, e0157590. [Google Scholar] [CrossRef] [PubMed]
- Olchawa, M.M.; Pilat, A.K.; Szewczyk, G.M.; Sarna, T.J. Inhibition of phagocytic activity of ARPE-19 cells by free radical mediated oxidative stress. Free Radic. Res. 2016, 50, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Bootman, M.D.; Chehab, T.; Bultynck, G.; Parys, J.B.; Rietdorf, K. The regulation of autophagy by calcium signals: Do we have a consensus? Cell Calcium 2017. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhang, T.; Sun, L.; Luo, Y.; Liu, D.H.; Xie, S.T.; Song, X.Y.; Wang, G.F.; Chen, X.L.; Zhou, B.C.; et al. Calpain, Atg5 and Bak play important roles in the crosstalk between apoptosis and autophagy induced by influx of extracellular calcium. Apoptosis 2013, 18, 435–451. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Scull, C.; Ozcan, L.; Tabas, I. NADPH oxidase links endoplasmic reticulum stress, oxidative stress, and PKR activation to induce apoptosis. J. Cell Biol. 2010, 191, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Zhai, P.; Shao, D.; Zablocki, D.; Nagarajan, N.; Terada, L.S.; Volpe, M.; Sadoshima, J. Activation of NADPH oxidase 4 in the endoplasmic reticulum promotes cardiomyocyte autophagy and survival during energy stress through the protein kinase RNA-activated-like endoplasmic reticulum kinase/eukaryotic initiation factor 2alpha/activating transcription factor 4 pathway. Circ. Res. 2013, 113, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Froger, N.; Cadetti, L.; Lorach, H.; Martins, J.; Bemelmans, A.P.; Dubus, E.; Degardin, J.; Pain, D.; Forster, V.; Chicaud, L.; et al. Taurine provides neuroprotection against retinal ganglion cell degeneration. PLoS ONE 2012, 7, e42017. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.J.; Li, T.M.; Ko, C.H.; Cheng, C.F.; Ho, T.J.; Liu, X.; Tsang, H.; Lin, T.H.; Liao, C.C.; Li, J.P.; et al. Effects of Chinese herbal medicines on the occurrence of diabetic retinopathy in type 2 diabetes patients and protection of ARPE-19 retina cells by inhibiting oxidative stress. Oncotarget 2017. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ren, S.; Liu, Y.; Gao, K.; Liu, Z.; Zhang, Z. Inhibition of Starvation-Triggered Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in ARPE-19 Cells by Taurine through Modulating the Expression of Calpain-1 and Calpain-2. Int. J. Mol. Sci. 2017, 18, 2146. https://doi.org/10.3390/ijms18102146
Zhang Y, Ren S, Liu Y, Gao K, Liu Z, Zhang Z. Inhibition of Starvation-Triggered Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in ARPE-19 Cells by Taurine through Modulating the Expression of Calpain-1 and Calpain-2. International Journal of Molecular Sciences. 2017; 18(10):2146. https://doi.org/10.3390/ijms18102146
Chicago/Turabian StyleZhang, Yuanyuan, Shu Ren, Yuci Liu, Kun Gao, Zheng Liu, and Zhou Zhang. 2017. "Inhibition of Starvation-Triggered Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in ARPE-19 Cells by Taurine through Modulating the Expression of Calpain-1 and Calpain-2" International Journal of Molecular Sciences 18, no. 10: 2146. https://doi.org/10.3390/ijms18102146