
The HIV-1 Vpr Protein: A Multifaceted Target for Therapeutic Intervention


Abstract
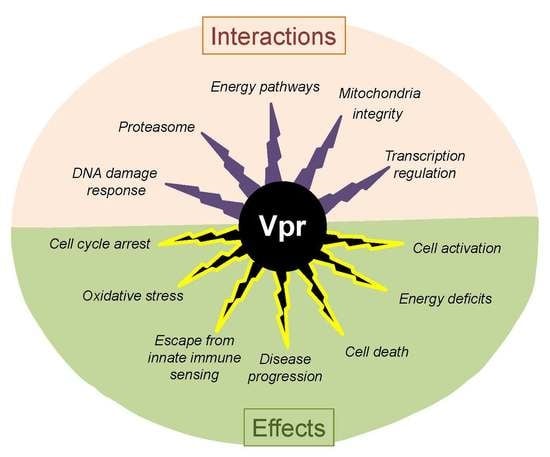
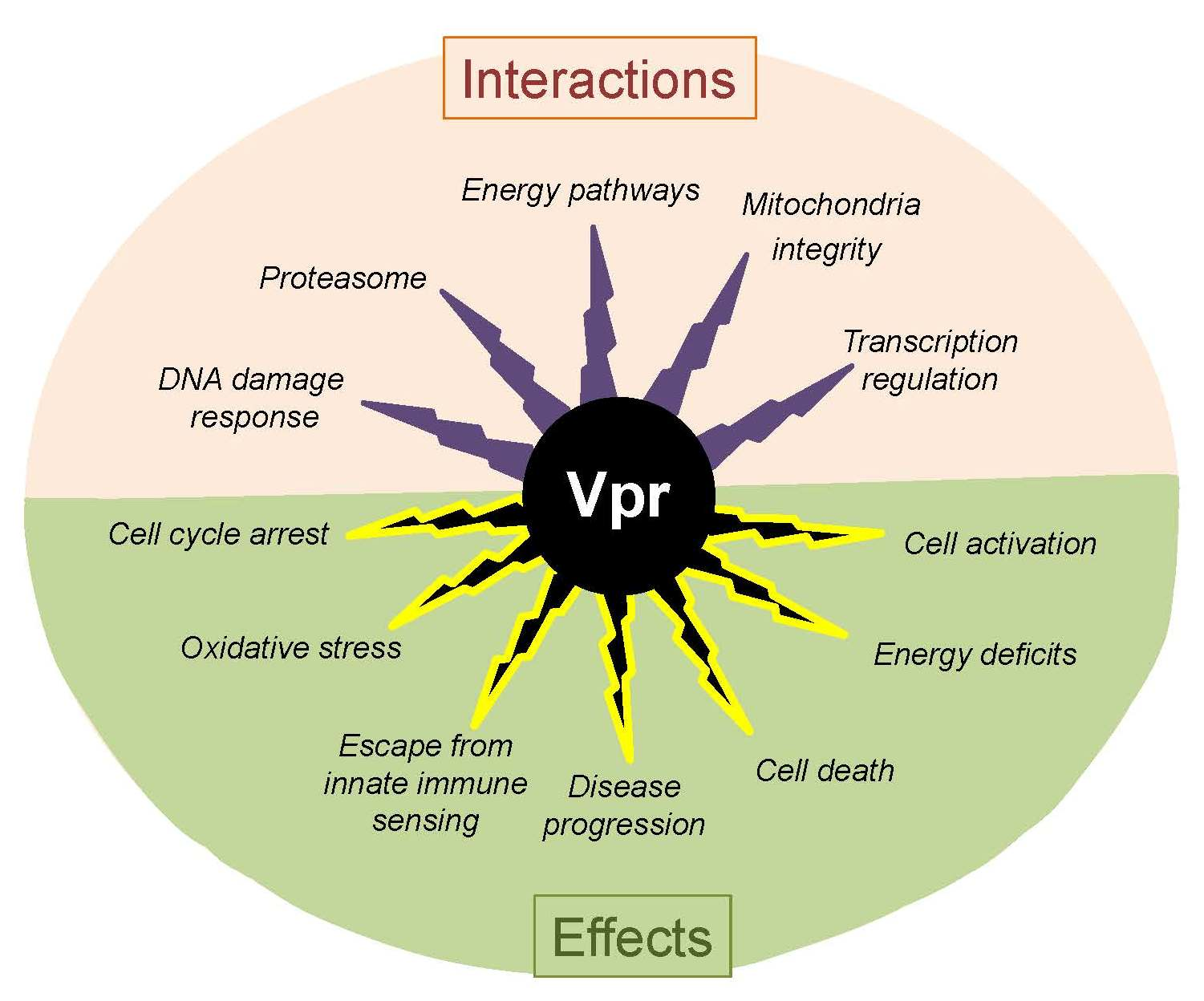
:
1. Introduction
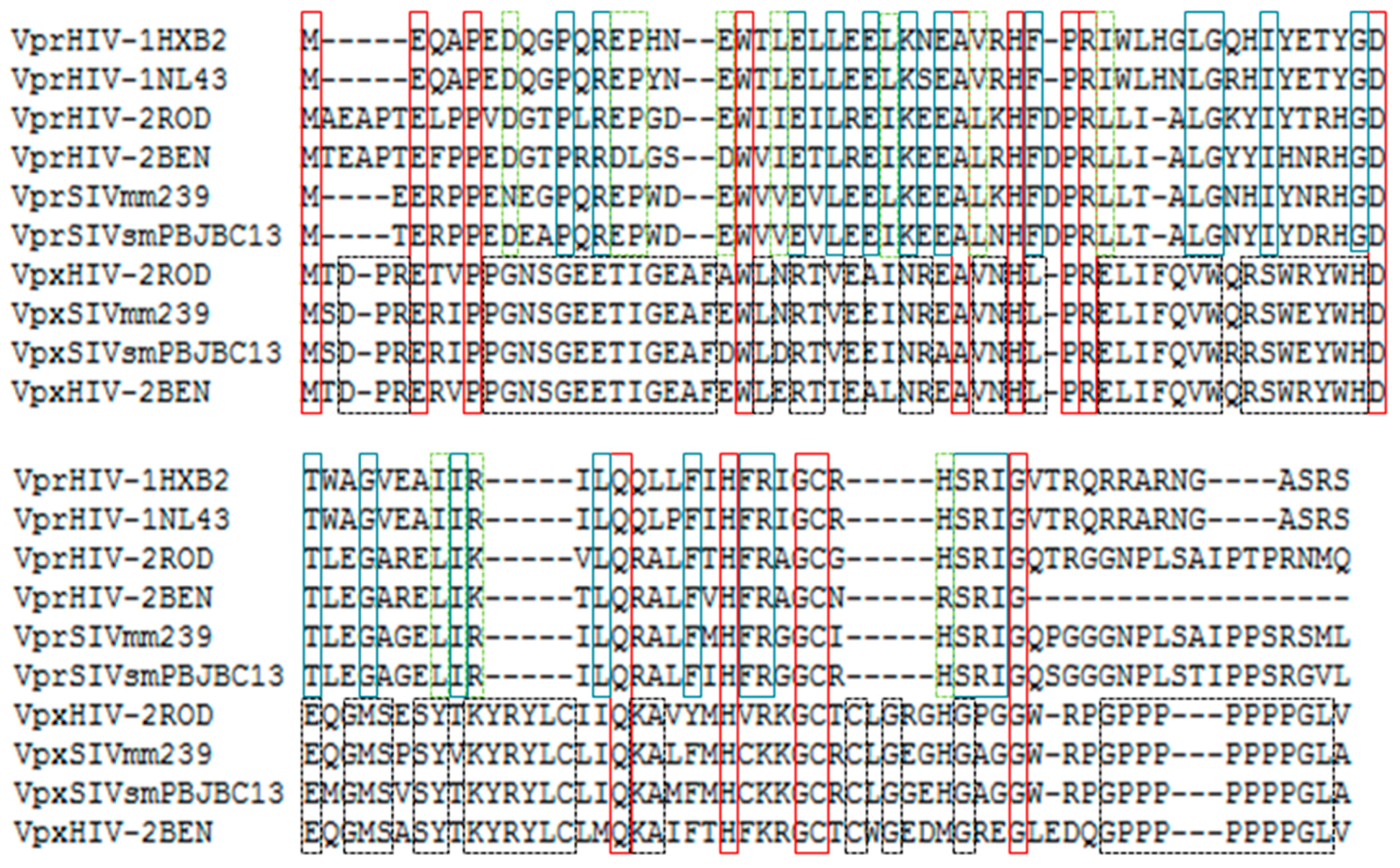
2. Origin and Conservation of Vpr Protein from Human Lentiviruses
3. The Role of Vpr Protein in HIV-1 Infection and Disease Progression
3.1. Concurrence and Independence of Cytopathogenic Activities of Vpr
3.2. The Influence of Vpr on Disease Progression
4. Functional Perturbation of Cells by the HIV-1 Vpr Protein
5. Molecular Mechanisms That Are Affected by Vpr Interaction with Cellular Factors
5.1. Energy Pathways, Redox Homeostasis, and Cell Cycle
5.2. Proteasomal Activity and Cell Death
5.3. Transcriptional Regulation
5.4. DNA Repair Mechanisms
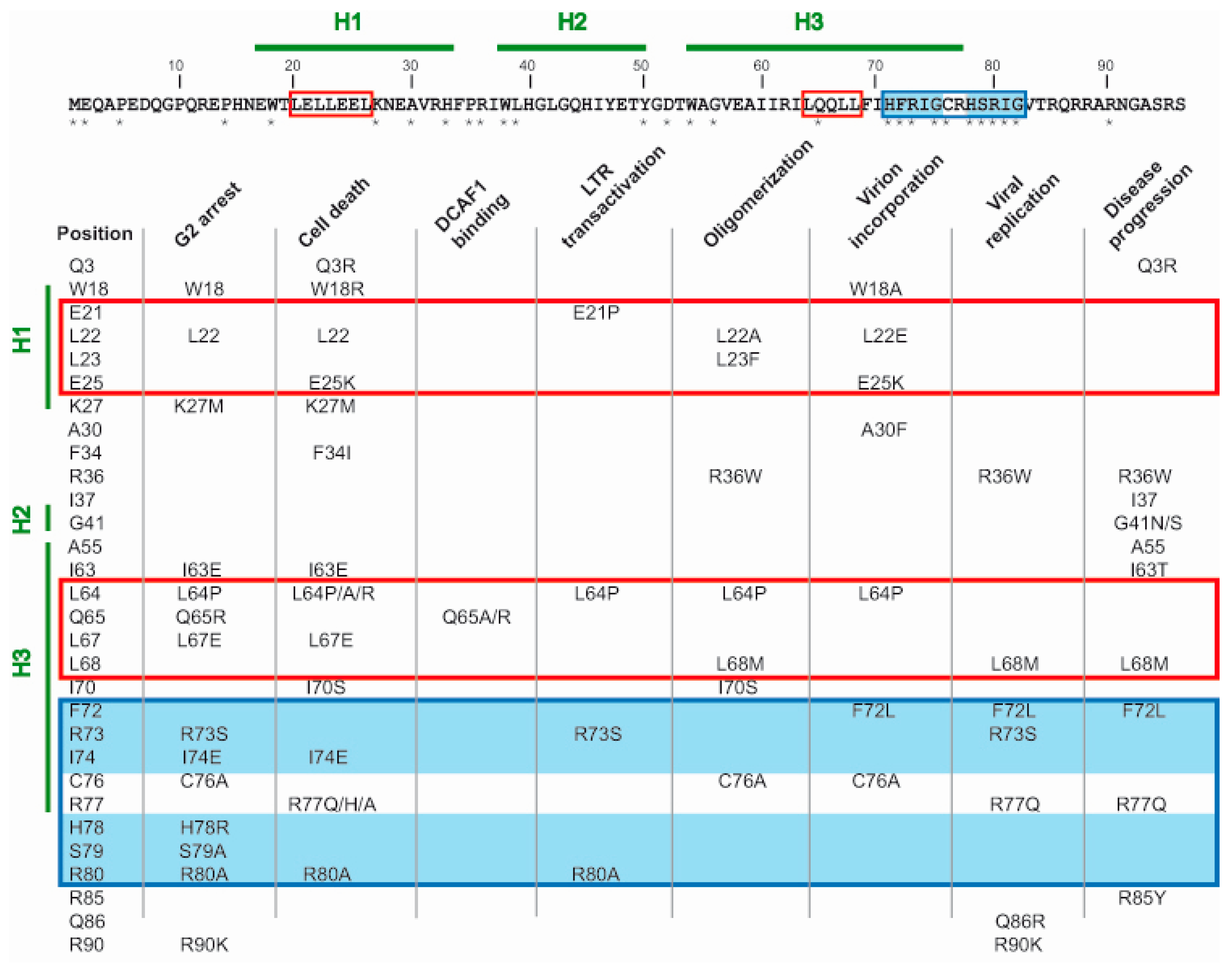
6. Amino Acid Residues Contributing to Vpr Functions
7. Progress in Searching for Vpr-Targeted Drugs
8. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Ogawa, K.; Shibata, R.; Kiyomasu, T.; Higuchi, I.; Kishida, Y.; Ishimoto, A.; Adachi, A. Mutational analysis of the human immunodeficiency virus Vpr open reading frame. J. Virol. 1989, 63, 4110–4114. [Google Scholar] [PubMed]
- Wong-Staal, F.; Chanda, P.K.; Ghrayeb, J. Human immunodeficiency virus: The eighth gene. AIDS Res. Hum. Retrovir. 1987, 3, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Dedera, D.; Hu, W.; Vander, H.N.; Ratner, L. Viral protein R of human immunodeficiency virus types 1 and 2 is dispensable for replication and cytopathogenicity in lymphoid cells. J. Virol. 1989, 63, 3205–3208. [Google Scholar] [PubMed]
- Levy, D.N.; Refaeli, Y.; MacGregor, R.R.; Weiner, D.B. Serum Vpr regulates productive infection and latency of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 1994, 91, 10873–10877. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.A.; Terwilliger, E.F.; Jalinoos, Y.; Proulx, J.; Sodroski, J.G.; Haseltine, W.A. Identification of HIV-1 Vpr product and function. J. Acquir. Immune Defic. Syndr. 1990, 3, 11–18. [Google Scholar] [PubMed]
- Balotta, C.; Lusso, P.; Crowley, R.; Gallo, R.C.; Franchini, G. Antisense phosphorothioate oligodeoxynucleotides targeted to the vpr gene inhibit human immunodeficiency virus type 1 replication in primary human macrophages. J. Virol. 1993, 67, 4409–4414. [Google Scholar] [PubMed]
- Hattori, N.; Michaels, F.; Fargnoli, K.; Marcon, L.; Gallo, R.C.; Franchini, G. The human immunodeficiency virus type 2 vpr gene is essential for productive infection of human macrophages. Proc. Natl. Acad. Sci. USA 1990, 87, 8080–8084. [Google Scholar] [CrossRef] [PubMed]
- Connor, R.I.; Chen, B.K.; Choe, S.; Landau, N.R. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology 1995, 206, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Heinzinger, N.K.; Bukinsky, M.I.; Haggerty, S.A.; Ragland, A.M.; Kewalramani, V.; Lee, M.A.; Gendelman, H.E.; Ratner, L.; Stevenson, M.; Emerman, M. The Vpr protein of human immunodeficiency virus type 1 influences nuclear localization of viral nucleic acids in nondividing host cells. Proc. Natl. Acad. Sci. USA 1994, 91, 7311–7315. [Google Scholar] [CrossRef] [PubMed]
- Hohne, K.; Businger, R.; van Nuffel, A.; Bolduan, S.; Koppensteiner, H.; Baeyens, A.; Vermeire, J.; Malatinkova, E.; Verhasselt, B.; Schindler, M. Virion encapsidated HIV-1 Vpr induces NFAT to prime non-activated T cells for productive infection. Open Biol. 2016, 6, 160046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, C.A.; Mukhtar, M.; Pomerantz, R.J. Human immunodeficiency virus type 1 Vpr induces apoptosis in human neuronal cells. J. Virol. 2000, 74, 9717–9726. [Google Scholar] [CrossRef] [PubMed]
- Piller, S.C.; Jans, P.; Gage, P.W.; Jans, D.A. Extracellular HIV-1 virus protein R causes a large inward current and cell death in cultured hippocampal neurons: Implications for AIDS pathology. Proc. Natl. Acad. Sci. USA 1998, 95, 4595–4600. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.A.; Simon, M.N.; Gross, I.; Krausslich, H.G.; Fuller, S.D.; Vogt, V.M.; Johnson, M.C. The stoichiometry of Gag protein in HIV-1. Nat. Struct. Mol. Biol. 2004, 11, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.A.; Dehni, G.; Sodroski, J.G.; Haseltine, W.A. Human immunodeficiency virus Vpr product is a virion-associated regulatory protein. J. Virol. 1990, 64, 3097–3099. [Google Scholar] [PubMed]
- Muller, B.; Tessmer, U.; Schubert, U.; Krausslich, H.G. Human immunodeficiency virus type 1 Vpr protein is incorporated into the virion in significantly smaller amounts than gag and is phosphorylated in infected cells. J. Virol. 2000, 74, 9727–9731. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.L.; Spearman, P.; Ratner, L. Human immunodeficiency virus type 1 viral protein R localization in infected cells and virions. J. Virol. 1993, 67, 6542–6550. [Google Scholar] [PubMed]
- Desai, T.M.; Marin, M.; Sood, C.; Shi, J.; Nawaz, F.; Aiken, C.; Melikyan, G.B. Fluorescent protein-tagged Vpr dissociates from HIV-1 core after viral fusion and rapidly enters the cell nucleus. Retrovirology 2015, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.N.; Refaeli, Y.; Weiner, D.B. Extracellular Vpr protein increases cellular permissiveness to human immunodeficiency virus replication and reactivates virus from latency. J. Virol. 1995, 69, 1243–1252. [Google Scholar] [PubMed]
- Wu, X.; Liu, H.; Xiao, H.; Kim, J.; Seshaiah, P.; Natsoulis, G.; Boeke, J.D.; Hahn, B.H.; Kappes, J.C. Targeting foreign proteins to human immunodeficiency virus particles via fusion with Vpr and Vpx. J. Virol. 1995, 69, 3389–3398. [Google Scholar] [PubMed]
- Stivahtis, G.L.; Soares, M.A.; Vodicka, M.A.; Hahn, B.H.; Emerman, M. Conservation and host specificity of Vpr-mediated cell cycle arrest suggest a fundamental role in primate lentivirus evolution and biology. J. Virol. 1997, 71, 4331–4338. [Google Scholar] [PubMed]
- Tristem, M.; Marshall, C.; Karpas, A.; Hill, F. Evolution of the primate lentiviruses: Evidence from Vpx and Vpr. EMBO J. 1992, 11, 3405–3412. [Google Scholar] [PubMed]
- Lang, S.M.; Weeger, M.; Stahl-Hennig, C.; Coulibaly, C.; Hunsmann, G.; Muller, J.; Muller-Hermelink, H.; Fuchs, D.; Wachter, H.; Daniel, M.M. Importance of Vpr for infection of rhesus monkeys with simian immunodeficiency virus. J. Virol. 1993, 67, 902–912. [Google Scholar] [PubMed]
- Hartman, T.L.; Buckheit, R.W. The continuing evolution of HIV-1 therapy: Identification and development of novel antiretroviral agents targeting viral and cellular targets. Mol. Biol. Int. 2012, 2012, 401965. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.C.; Rogel, M.E.; Kinsey, C.M.; Michael, S.F.; Fultz, P.N.; Nowak, M.A.; Hahn, B.H.; Emerman, M. HIV-1 Vpr increases viral expression by manipulation of the cell cycle: A mechanism for selection of Vpr in vivo. Nat. Med. 1998, 4, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Rogel, M.E.; Wu, L.I.; Emerman, M. The human immunodeficiency virus type 1 vpr gene prevents cell proliferation during chronic infection. J. Virol. 1995, 69, 882–888. [Google Scholar] [PubMed]
- Sharp, P.M.; Bailes, E.; Stevenson, M.; Emerman, M.; Hahn, B.H. Gene acquisition in HIV and SIV. Nature 1996, 383, 586–587. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, T.M.; Brichacek, B.; Sharova, N.; Newman, M.A.; Stivahtis, G.; Sharp, P.M.; Emerman, M.; Hahn, B.H.; Stevenson, M. Nuclear import and cell cycle arrest functions of the HIV-1 Vpr protein are encoded by two separate genes in HIV-2/SIV(SM). EMBO J. 1996, 15, 6155–6165. [Google Scholar] [PubMed]
- Wu, Y.; Zhou, X.; Barnes, C.O.; DeLucia, M.; Cohen, A.E.; Gronenborn, A.M.; Ahn, J.; Calero, G. The DDB1-DCAF1-Vpr-UNG2 crystal structure reveals how HIV-1 Vpr steers human UNG2 toward destruction. Nat. Struct. Mol. Biol. 2016, 23, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Morellet, N.; Bouaziz, S.; Petitjean, P.; Roques, B.P. NMR structure of the HIV-1 regulatory protein Vpr. J. Mol. Biol. 2003, 327, 215–227. [Google Scholar] [CrossRef]
- Khamsri, B.; Murao, F.; Yoshida, A.; Sakurai, A.; Uchiyama, T.; Shirai, H.; Matsuo, Y.; Fujita, M.; Adachi, A. Comparative study on the structure and cytopathogenic activity of HIV Vpr/Vpx proteins. Microbes Infect. 2006, 8, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, Z.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Etienne, L.; Hahn, B.H.; Sharp, P.M.; Matsen, F.A.; Emerman, M. Gene loss and adaptation to hominids underlie the ancient origin of HIV-1. Cell Host Microbe 2013, 14, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Sakai, Y.; Doi, N.; Miyazaki, Y.; Adachi, A.; Nomaguchi, M. Phylogenetic insights into the functional relationship between primate lentiviral reverse transcriptase and accessory proteins Vpx/Vpr. Front. Microbiol. 2016, 7, 1655. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Choe, S.; Walker, R.; di, M.P.; Morgan, D.O.; Landau, N.R. Human immunodeficiency virus type 1 viral protein R (Vpr) arrests cells in the G2 phase of the cell cycle by inhibiting p34cdc2 activity. J. Virol. 1995, 69, 6705–6711. [Google Scholar] [PubMed]
- Jowett, J.B.; Planelles, V.; Poon, B.; Shah, N.P.; Chen, M.L.; Chen, I.S. The human immunodeficiency virus type 1 vpr gene arrests infected T cells in the G2 + M phase of the cell cycle. J. Virol. 1995, 69, 6304–6313. [Google Scholar] [PubMed]
- Re, F.; Braaten, D.; Franke, E.K.; Luban, J. Human immunodeficiency virus type 1 Vpr arrests the cell cycle in G2 by inhibiting the activation of p34cdc2-cyclin B. J. Virol. 1995, 69, 6859–6864. [Google Scholar] [PubMed]
- Popov, S.; Rexach, M.; Ratner, L.; Blobel, G.; Bukrinsky, M. Viral protein R regulates docking of the HIV-1 preintegration complex to the nuclear pore complex. J. Biol. Chem. 1998, 273, 13347–13352. [Google Scholar] [CrossRef] [PubMed]
- Arunagiri, C.; Macreadie, I.; Hewish, D.; Azad, A. A C-terminal domain of HIV-1 accessory protein Vpr is involved in penetration, mitochondrial dysfunction and apoptosis of human CD4+ lymphocytes. Apoptosis 1997, 2, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ayyavoo, V.; Mahboubi, A.; Mahalingam, S.; Ramalingam, R.; Kudchodkar, S.; Williams, W.V.; Green, D.R.; Weiner, D.B. HIV-1 Vpr suppresses immune activation and apoptosis through regulation of nuclear factor κB. Nat. Med. 1997, 3, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Majumder, B.; Venkatachari, N.J.; Srinivasan, A.; Ayyavoo, V. HIV-1 mediated immune pathogenesis: Spotlight on the role of viral protein R (Vpr). Curr. HIV Res. 2009, 7, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Vermeire, J.; Roesch, F.; Sauter, D.; Rua, R.; Hotter, D.; van Nuffel, A.; Vanderstraeten, H.; Naessens, E.; Iannucci, V.; Landi, A.; et al. HIV triggers a cGAS-dependent, Vpu- and Vpr-regulated type I interferon response in CD4+ T cells. Cell Rep. 2016, 17, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.J.; Wang, L.; Mukherjee, S.; Narayan, O. Biochemical mechanism of HIV-1 Vpr function. Oligomerization mediated by the N-terminal domain. J. Biol. Chem. 1994, 269, 32131–32137. [Google Scholar] [PubMed]
- Bolton, D.L.; Lenardo, M.J. Vpr cytopathicity independent of G2/M cell cycle arrest in HIV-1-infected CD4+ T cells. J. Virol. 2007, 81, 8878–8890. [Google Scholar] [CrossRef] [PubMed]
- Venkatachari, N.J.; Walker, L.A.; Tastan, O.; Le, T.; Dempsey, T.M.; Li, Y.; Yanamala, N.; Srinivasan, A.; Klein-Seetharaman, J.; Montelaro, R.C.; et al. Human immunodeficiency virus type 1 Vpr: Oligomerization is an essential feature for its incorporation into virus particles. Virol. J. 2010, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Di, M.P.; Choe, S.; Ebright, M.; Knoblauch, R.; Landau, N.R. Mutational analysis of cell cycle arrest, nuclear localization and virion packaging of human immunodeficiency virus type 1 Vpr. J. Virol. 1995, 69, 7909–7916. [Google Scholar]
- Ferrucci, A.; Nonnemacher, M.R.; Wigdahl, B. Human immunodeficiency virus viral protein R as an extracellular protein in neuropathogenesis. Adv. Virus Res. 2011, 81, 165–199. [Google Scholar] [PubMed]
- Huang, C.Y.; Chiang, S.F.; Lin, T.Y.; Chiou, S.H.; Chow, K.C. HIV-1 Vpr triggers mitochondrial destruction by impairing Mfn2-mediated ER-mitochondria interaction. PLoS ONE 2012, 7, e33657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, S.; Planelles, V.; Wu, L. Differential effects of Vpr on single-cycle and spreading HIV-1 infections in CD4+ T cells and dendritic cells. PLoS ONE 2012, 7, e35385. [Google Scholar] [CrossRef] [PubMed]
- Hrimech, M.; Yao, X.J.; Bachand, F.; Rougeau, N.; Cohen, E.A. Human immunodeficiency virus type 1 (HIV-1) Vpr functions as an immediate-early protein during HIV-1 infection. J. Virol. 1999, 73, 4101–4109. [Google Scholar] [PubMed]
- Conti, L.; Matarrese, P.; Varano, B.; Gauzzi, M.C.; Sato, A.; Malorni, W.; Belardelli, F.; Gessani, S. Dual role of the HIV-1 Vpr protein in the modulation of the apoptotic response of T cells. J. Immunol. 2000, 165, 3293–3300. [Google Scholar] [CrossRef] [PubMed]
- Trotard, M.; Tsopoulidis, N.; Tibroni, N.; Willemsen, J.; Binder, M.; Ruggieri, A.; Fackler, O.T. Sensing of HIV-1 infection in Tzm-bl cells with reconstituted expression of STING. J. Virol. 2015, 90, 2064–2076. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Riviere, L.; Jarrosson-Wuilleme, L.; Bernaud, J.; Rigal, D.; Darlix, J.L.; Cimarelli, A. SIVSM/HIV-2 Vpx proteins promote retroviral escape from a proteasome-dependent restriction pathway present in human dendritic cells. Retrovirology 2007, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Hrecka, K.; Hao, C.; Shun, M.C.; Kaur, S.; Swanson, S.K.; Florens, L.; Washburn, M.P.; Skowronski, J. HIV-1 and HIV-2 exhibit divergent interactions with HLTF and UNG2 DNA repair proteins. Proc. Natl. Acad. Sci. USA 2016, 113, E3921–E3930. [Google Scholar] [CrossRef] [PubMed]
- Sharova, N.; Wu, Y.; Zhu, X.; Stranska, R.; Kaushik, R.; Sharkey, M.; Stevenson, M. Primate lentiviral Vpx commandeers DDB1 to counteract a macrophage restriction. PLoS Pathog. 2008, 4, e1000057. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.L.; Dehart, J.L.; Zimmerman, E.S.; Ardon, O.; Kim, B.; Jacquot, G.; Benichou, S.; Planelles, V. HIV-1 Vpr-induced apoptosis is cell cycle dependent and requires Bax but not ANT. PLoS Pathog. 2006, 2, e127. [Google Scholar] [CrossRef] [PubMed]
- Stromajer-Racz, T.; Gazdag, Z.; Belagyi, J.; Vagvolgyi, C.; Zhao, R.Y.; Pesti, M. Oxidative stress induced by HIV-1 F34IVpr in Schizosaccharomyces pombe is one of its multiple functions. Exp. Mol. Pathol. 2010, 88, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Aida, Y. Visualizing Vpr-induced G2 arrest and apoptosis. PLoS ONE 2014, 9, e86840. [Google Scholar] [CrossRef] [PubMed]
- Felzien, L.K.; Woffendin, C.; Hottiger, M.O.; Subbramanian, R.A.; Cohen, E.A.; Nabel, G.J. HIV transcriptional activation by the accessory protein, Vpr, is mediated by the p300 co-activator. Proc. Natl. Acad. Sci. USA 1998, 95, 5281–5286. [Google Scholar] [CrossRef] [PubMed]
- Vodicka, M.A.; Koepp, D.M.; Silver, P.A.; Emerman, M. HIV-1 Vpr interacts with the nuclear transport pathway to promote macrophage infection. Genes Dev. 1998, 12, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, G.; Le, R.E.; David, A.; Mazzolini, J.; Bouchet, J.; Bouaziz, S.; Niedergang, F.; Pancino, G.; Benichou, S. Localization of HIV-1 Vpr to the nuclear envelope: Impact on Vpr functions and virus replication in macrophages. Retrovirology 2007, 4, 84. [Google Scholar] [CrossRef] [PubMed]
- Iijima, S.; Nitahara-Kasahara, Y.; Kimata, K.; Zhong, Z.W.; Kamata, M.; Isogai, M.; Miwa, M.; Tsunetsugu-Yokota, Y.; Aida, Y. Nuclear localization of Vpr is crucial for the efficient replication of HIV-1 in primary CD4+ T cells. Virology 2004, 327, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, S.; Ayyavoo, V.; Patel, M.; Kieber-Emmons, T.; Weiner, D.B. Nuclear import, virion incorporation, and cell cycle arrest/differentiation are mediated by distinct functional domains of human immunodeficiency virus type 1 Vpr. J. Virol. 1997, 71, 6339–6347. [Google Scholar] [PubMed]
- Levy, D.N.; Fernandes, L.S.; Williams, W.V.; Weiner, D.B. Induction of cell differentiation by human immunodeficiency virus 1 Vpr. Cell 1993, 72, 541–550. [Google Scholar] [CrossRef]
- Igarashi, T.; Brown, C.R.; Endo, Y.; Buckler-White, A.; Plishka, R.; Bischofberger, N.; Hirsch, V.; Martin, M.A. Macrophage are the principal reservoir and sustain high virus loads in rhesus macaques after the depletion of CD4+ T cells by a highly pathogenic simian immunodeficiency virus/HIV type 1 chimera (SHIV): Implications for HIV-1 infections of humans. Proc. Natl. Acad. Sci. USA 2001, 98, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, A.; Nonnemacher, M.R.; Wigdahl, B. Extracellular HIV-1 viral protein R affects astrocytic glyceraldehyde 3-phosphate dehydrogenase activity and neuronal survival. J. Neurovirol. 2013, 19, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Laguette, N.; Bregnard, C.; Hue, P.; Basbous, J.; Yatim, A.; Larroque, M.; Kirchhoff, F.; Constantinou, A.; Sobhian, B.; Benkirane, M. Premature activation of the SLX4 complex by Vpr promotes G2/M arrest and escape from innate immune sensing. Cell 2014, 156, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Lindl, K.A.; Marks, D.R.; Kolson, D.L.; Jordan-Sciutto, K.L. HIV-associated neurocognitive disorder: Pathogenesis and therapeutic opportunities. J. Neuroimmune Pharmacol. 2010, 5, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, M.A.; Zhao, M.L.; Lee, S.C. HIV-1 expression protects macrophages and microglia from apoptotic death. Neuropathol. Appl. Neurobiol. 2004, 30, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Dumas, A.; Le-Bury, G.; Marie-Anais, F.; Herit, F.; Mazzolini, J.; Guilbert, T.; Bourdoncle, P.; Russell, D.G.; Benichou, S.; Zahraoui, A.; et al. The HIV-1 protein Vpr impairs phagosome maturation by controlling microtubule-dependent trafficking. J. Cell Biol. 2015, 211, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Jambo, K.C.; Banda, D.H.; Kankwatira, A.M.; Sukumar, N.; Allain, T.J.; Heyderman, R.S.; Russell, D.G.; Mwandumba, H.C. Small alveolar macrophages are infected preferentially by HIV and exhibit impaired phagocytic function. Mucosal Immunol. 2014, 7, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, Y.; Sanchez, P.V.; Meyer, B.E.; Malim, M.H. Nuclear export of human immunodeficiency virus type 1 Vpr is not required for virion packaging. J. Virol. 2001, 75, 8348–8352. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.A.; Poon, B.; Jowett, J.B.; Chen, I.S. Human immunodeficiency virus type 1 Vpr induces apoptosis following cell cycle arrest. J. Virol. 1997, 71, 5579–5592. [Google Scholar] [PubMed]
- Zhao, Y.; Elder, R.T.; Chen, M.; Cao, J. Fission yeast expression vectors adapted for positive identification of gene insertion and green fluorescent protein fusion. Biotechniques 1998, 25, 438–440. [Google Scholar] [PubMed]
- Sato, K.; Misawa, N.; Iwami, S.; Satou, Y.; Matsuoka, M.; Ishizaka, Y.; Ito, M.; Aihara, K.; An, D.S.; Koyanagi, Y. HIV-1 Vpr accelerates viral replication during acute infection by exploitation of proliferating CD4+ T cells in vivo. PLoS Pathog. 2013, 9, e1003812. [Google Scholar] [CrossRef] [PubMed]
- Roesch, F.; Richard, L.; Rua, R.; Porrot, F.; Casartelli, N.; Schwartz, O. Vpr enhances tumor necrosis factor production by HIV-1-infected T cells. J. Virol. 2015, 89, 12118–12130. [Google Scholar] [CrossRef] [PubMed]
- Aukrust, P.; Liabakk, N.B.; Muller, F.; Lien, E.; Espevik, T.; Froland, S.S. Serum levels of tumor necrosis factor-alpha (TNFα) and soluble TNF receptors in human immunodeficiency virus type 1 infection-correlations to clinical, immunologic, and virologic parameters. J. Infect. Dis. 1994, 169, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.A.; Korner, C.; Sirignano, M.N.; Amero, M.; Bazner, S.; Rychert, J.; Allen, T.M.; Rosenberg, E.S.; Bosch, R.J.; Altfeld, M. Tumor necrosis factor alpha is associated with viral control and early disease progression in patients with HIV type 1 infection. J. Infect. Dis. 2014, 210, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.; Passmore, J.A.; Williamson, C.; Little, F.; Bebell, L.M.; Mlisana, K.; Burgers, W.A.; van Loggerenberg, F.; Walzl, G.; Djoba Siawaya, J.F.; et al. Plasma cytokine levels during acute HIV-1 infection predict HIV disease progression. AIDS 2010, 24, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Gangwani, M.R.; Kumar, A. Multiple protein kinases via activation of transcription factors NF-κB, AP-1 and C/EBP-γ regulate the IL-6/IL-8 production by HIV-1 Vpr in astrocytes. PLoS ONE 2015, 10, e0135633. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.; Alfieri, C.; Hrimech, M.; Cohen, E.A.; Tanner, J.E. Activation of transcription factors NF-κB and NF-IL-6 by human immunodeficiency virus type 1 protein R (Vpr) induces interleukin-8 expression. J. Virol. 2000, 74, 4658–4665. [Google Scholar] [CrossRef] [PubMed]
- Mirani, M.; Elenkov, I.; Volpi, S.; Hiroi, N.; Chrousos, G.P.; Kino, T. HIV-1 protein Vpr suppresses IL-12 production from human monocytes by enhancing glucocorticoid action: Potential implications of Vpr coactivator activity for the innate and cellular immunity deficits observed in HIV-1 infection. J. Immunol. 2002, 169, 6361–6368. [Google Scholar] [CrossRef] [PubMed]
- Macreadie, I.G.; Arunagiri, C.K.; Hewish, D.R.; White, J.F.; Azad, A.A. Extracellular addition of a domain of HIV-1 Vpr containing the amino acid sequence motif H(S/F)RIG causes cell membrane permeabilization and death. Mol. Microbiol. 1996, 19, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Jacotot, E.; Ravagnan, L.; Loeffler, M.; Ferri, K.F.; Vieira, H.L.; Zamzami, N.; Costantini, P.; Druillennec, S.; Hoebeke, J.; Briand, J.P.; et al. The HIV-1 viral protein R induces apoptosis via a direct effect on the mitochondrial permeability transition pore. J. Exp. Med. 2000, 191, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Mukerjee, R.; Chang, J.R.; del, V.L.; Bagashev, A.; Gayed, M.M.; Lyde, R.B.; Hawkins, B.J.; Brailoiu, E.; Cohen, E.; Power, C.; et al. Deregulation of microRNAs by HIV-1 Vpr protein leads to the development of neurocognitive disorders. J. Biol. Chem. 2011, 286, 34976–34985. [Google Scholar] [CrossRef] [PubMed]
- Macreadie, I.G.; Thorburn, D.R.; Kirby, D.M.; Castelli, L.A.; de Rozario, N.L.; Azad, A.A. HIV-1 protein Vpr causes gross mitochondrial dysfunction in the yeast Saccharomyces cerevisiae. FEBS Lett. 1997, 410, 145–149. [Google Scholar] [CrossRef]
- Bolton, D.L.; Hahn, B.I.; Park, E.A.; Lehnhoff, L.L.; Hornung, F.; Lenardo, M.J. Death of CD4+ T cell lines caused by human immunodeficiency virus type 1 does not depend on caspases or apoptosis. J. Virol. 2002, 76, 5094–5107. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Elder, R.T.; Yu, M.; O’Gorman, M.G.; Selig, L.; Benarous, R.; Yamamoto, A.; Zhao, Y. Mutational analysis of Vpr-induced G2 arrest, nuclear localization, and cell death in fission yeast. J. Virol. 1999, 73, 3236–3245. [Google Scholar] [PubMed]
- Chen, R.; Le, R.E.; Kearney, J.A.; Mansky, L.M.; Benichou, S. Vpr-mediated incorporation of UNG2 into HIV-1 particles is required to modulate the virus mutation rate and for replication in macrophages. J. Biol. Chem. 2004, 279, 28419–28425. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.B.; Weeks, O.; Zhao, L.J.; Saltarelli, M.; Bond, V.C. Effects of extracellular human immunodeficiency virus type 1 Vpr protein in primary rat cortical cell cultures. J. Neurovirol. 2000, 6, 202–220. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M.; Preveral, S.; Selig, L.; Benarous, R.; Benichou, S. The interaction of Vpr with uracil DNA glycosylase modulates the human immunodeficiency virus type 1 in vivo mutation rate. J. Virol. 2000, 74, 7039–7047. [Google Scholar] [CrossRef] [PubMed]
- Yamada, E.; Yoshikawa, R.; Nakano, Y.; Misawa, N.; Koyanagi, Y.; Sato, K. Impacts of humanized mouse models on the investigation of HIV-1 infection: Illuminating the roles of viral accessory proteins in vivo. Viruses 2015, 7, 1373–1390. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.; Sadowska, M.; Denaro, F.; Rao, S.; Foulke, J., Jr.; Hayes, N.; Jones, O.; Doodnauth, D.; Davis, H.; Sill, A.; et al. An HIV-1 transgenic rat that develops HIV-related pathology and immunologic dysfunction. Proc. Natl. Acad. Sci. USA 2001, 98, 9271–9276. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, L.M.; Purnell, P.R.; Stauch, K.L.; Callen, S.E.; Buch, S.J.; Fox, H.S. HIV-1 transgenic rats display mitochondrial abnormalities consistent with abnormal energy generation and distribution. J. Neurovirol. 2016, 22, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Barrero, C.A.; Datta, P.K.; Sen, S.; Deshmane, S.; Amini, S.; Khalili, K.; Merali, S. HIV-1 Vpr modulates macrophage metabolic pathways: A SILAC-based quantitative analysis. PLoS ONE 2013, 8, e68376. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Zeng, Y.; Wu, X.; Ji, Y.; He, X.; Andrus, T.; Zhu, T.; Wang, T. Endogenous HIV-1 Vpr-mediated apoptosis and proteome alteration of human T cell leukemia virus-1 transformed C8166 cells. Apoptosis 2009, 14, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Lahouassa, H.; Blondot, M.L.; Chauveau, L.; Chougui, G.; Morel, M.; Leduc, M.; Guillonneau, F.; Ramirez, B.C.; Schwartz, O.; Margottin-Goguet, F. HIV-1 Vpr degrades the HLTF DNA translocase in T cells and macrophages. Proc. Natl. Acad. Sci. USA 2016, 113, 5311–5316. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.M.; Olson, L.; Coppotelli, G. Mitochondrial and ubiquitin proteasome system dysfunction in ageing and disease: two sides of the same coin? Int. J. Mol. Sci. 2015, 16, 19458–19476. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, G.; Kaiser, P. The 26S proteasome and initiation of gene transcription. Biomolecules 2014, 4, 827–847. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Hollenbaugh, J.A.; Munger, J.; Kim, B. Metabolite profiles of human immunodeficiency virus infected CD4+ T cells and macrophages using LC-MS/MS analysis. Virology 2011, 415, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Kaminiski, R.; Deshmane, S.; Langford, D.; Khalili, K.; Amini, S.; Datta, P.K. Role of hexokinase-1 in the survival of HIV-1-infected macrophages. Cell Cycle 2015, 14, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Masuda, F.; Ishii, M.; Mori, A.; Uehara, L.; Yanagida, M.; Takeda, K.; Saitoh, S. Glucose restriction induces transient G2 cell cycle arrest extending cellular chronological lifespan. Sci. Rep. 2016, 6, 19629. [Google Scholar] [CrossRef] [PubMed]
- Antal, J.; Pesti, M. The dose-dependent H2O2 stress response promotes increased survival for Schizosaccharomyces pombe cells expressing HIV-1 Vpr. Folia Microbiol. 2006, 51, 406–412. [Google Scholar] [CrossRef]
- Baruchel, S.; Wainberg, M.A. The role of oxidative stress in disease progression in individuals infected by the human immunodeficiency virus. J. Leukoc. Biol. 1992, 52, 111–114. [Google Scholar] [PubMed]
- Porter, K.M.; Sutliff, R.L. HIV-1, reactive oxygen species, and vascular complications. Free Radic. Biol. Med. 2012, 53, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Staal, F.J.; Ela, S.W.; Roederer, M.; Anderson, M.T.; Herzenberg, L.A.; Herzenberg, L.A. Glutathione deficiency and human immunodeficiency virus infection. Lancet 1992, 339, 909–912. [Google Scholar] [CrossRef]
- Ferrucci, A.; Nonnemacher, M.R.; Cohen, E.A.; Wigdahl, B. Extracellular human immunodeficiency virus type 1 viral protein R causes reductions in astrocytic ATP and glutathione levels compromising the antioxidant reservoir. Virus Res. 2012, 167, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Monroy, N.; Herrero, L.; Carrasco, L.; Gonzalez, M.E. Influence of glutathione availability on cell damage induced by human immunodeficiency virus type 1 viral protein R. Virus Res. 2016, 213, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhang, X.; Li, S.; Liu, N.; Lian, W.; McDowell, E.; Zhou, P.; Zhao, C.; Guo, H.; Zhang, C.; et al. Physiological levels of ATP negatively regulate proteasome function. Cell Res. 2010, 20, 1372–1385. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.Y.; Sutton, J.N.; Jacobs, J.M.; Bondarenko, A.; Smith, R.D.; Katze, M.G. Dynamic host energetics and cytoskeletal proteomes in human immunodeficiency virus type 1-infected human primary CD4 cells: Analysis by multiplexed label-free mass spectrometry. J. Virol. 2009, 83, 9283–9295. [Google Scholar] [CrossRef] [PubMed]
- Palamara, A.T.; Perno, C.F.; Aquaro, S.; Bue, M.C.; Dini, L.; Garaci, E. Glutathione inhibits HIV replication by acting at late stages of the virus life cycle. AIDS Res. Hum. Retrovir. 1996, 12, 1537–1541. [Google Scholar] [CrossRef] [PubMed]
- Garaci, E.; Palamara, A.T.; Ciriolo, M.R.; D’Agostini, C.; Abdel-Latif, M.S.; Aquaro, S.; Lafavia, E.; Rotilio, G. Intracellular GSH content and HIV replication in human macrophages. J. Leukoc. Biol. 1997, 62, 54–59. [Google Scholar] [PubMed]
- Nordgren, M.; Fransen, M. Peroxisomal metabolism and oxidative stress. Biochimie 2014, 98, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmane, S.L.; Amini, S.; Sen, S.; Khalili, K.; Sawaya, B.E. Regulation of the HIV-1 promoter by HIF-1α and Vpr proteins. Virol. J. 2011, 8, 477. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Mukerjee, R.; Fan, S.; del Valle, L.; Michiels, C.; Sweet, T.; Rom, I.; Khalili, K.; Rappaport, J.; Amini, S.; et al. Activation of the oxidative stress pathway by HIV-1 Vpr leads to induction of hypoxia inducible factor 1 alpha expression. J. Biol. Chem. 2009, 284, 11364–11373. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, A.; Munshi, M.; Khan, S.Z.; Fatima, S.; Arya, R.; Jameel, S.; Singh, A. Measuring glutathione redox potential of HIV-1-infected macrophages. J. Biol. Chem. 2015, 290, 1020–1038. [Google Scholar] [CrossRef] [PubMed]
- Roshal, M.; Kim, B.; Zhu, Y.; Nghiem, P.; Planelles, V. Activation of the ATR-mediated DNA damage response by the HIV-1 viral protein R. J. Biol. Chem. 2003, 278, 25879–25886. [Google Scholar] [CrossRef] [PubMed]
- Bartz, S.R.; Rogel, M.E.; Emerman, M. Human immunodeficiency virus type 1 cell cycle control: Vpr is cytostatic and mediates G2 accumulation by a mechanism which differs from DNA damage checkpoint control. J. Virol. 1996, 70, 2324–2331. [Google Scholar] [PubMed]
- Elder, R.T.; Benko, Z.; Zhao, Y. HIV-1 VPR modulates cell cycle G2/M transition through an alternative cellular mechanism other than the classic mitotic checkpoints. Front. Biosci. 2002, 7, 349–357. [Google Scholar] [CrossRef]
- Lai, M.; Zimmerman, E.S.; Planelles, V.; Chen, J. Activation of the ATR pathway by human immunodeficiency virus type 1 Vpr involves its direct binding to chromatin in vivo. J. Virol. 2005, 79, 15443–15451. [Google Scholar] [CrossRef] [PubMed]
- Marechal, A.; Zou, L. DNA damage sensing by the ATM and ATR kinases. Cold Spring Harb. Perspect. Biol. 2013, 5, a012716. [Google Scholar] [CrossRef] [PubMed]
- Romani, B.; Baygloo, N.S.; Hamidi-Fard, M.; Aghasadeghi, M.R.; Allahbakhshi, E. HIV-1 Vpr protein induces proteasomal degradation of chromatin-associated class I HDACs to overcome latent infection of macrophages. J. Biol. Chem. 2016, 291, 2696–2711. [Google Scholar] [CrossRef] [PubMed]
- Dehart, J.L.; Zimmerman, E.S.; Ardon, O.; Monteiro-Filho, C.M.; Arganaraz, E.R.; Planelles, V. HIV-1 Vpr activates the G2 checkpoint through manipulation of the ubiquitin proteasome system. Virol. J. 2007, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Hrecka, K.; Gierszewska, M.; Srivastava, S.; Kozaczkiewicz, L.; Swanson, S.K.; Florens, L.; Washburn, M.P.; Skowronski, J. Lentiviral Vpr usurps Cul4-DDB1[VprBP] E3 ubiquitin ligase to modulate cell cycle. Proc. Natl. Acad. Sci. USA 2007, 104, 11778–11783. [Google Scholar] [CrossRef] [PubMed]
- Le Rouzic, E.; Belaidouni, N.; Estrabaud, E.; Morel, M.; Rain, J.C.; Transy, C.; Margottin-Goguet, F. HIV1 Vpr arrests the cell cycle by recruiting DCAF1/VprBP, a receptor of the Cul4-DDB1 ubiquitin ligase. Cell Cycle 2007, 6, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Romani, B.; Shaykh, B.N.; Aghasadeghi, M.R.; Allahbakhshi, E. HIV-1 Vpr protein enhances proteasomal degradation of MCM10 DNA replication factor through the Cul4-DDB1[VprBP] E3 ubiquitin ligase to induce G2/M cell cycle arrest. J. Biol. Chem. 2015, 290, 17380–17389. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.J.; Mukherjee, S.; Narayan, O. Biochemical mechanism of HIV-1 Vpr function. Specific interaction with a cellular protein. J. Biol. Chem. 1994, 269, 15577–15582. [Google Scholar] [PubMed]
- Zhang, S.; Feng, Y.; Narayan, O.; Zhao, L.J. Cytoplasmic retention of HIV-1 regulatory protein Vpr by protein–protein interaction with a novel human cytoplasmic protein VprBP. Gene 2001, 263, 131–140. [Google Scholar] [CrossRef]
- Wen, X.; Duus, K.M.; Friedrich, T.D.; de Noronha, C.M. The HIV1 protein Vpr acts to promote G2 cell cycle arrest by engaging a DDB1 and cullin4A containing ubiquitin ligase complex using VPRBP/DCAF1 as an adaptor. J. Biol. Chem. 2007, 282, 27046–27057. [Google Scholar] [CrossRef] [PubMed]
- Belzile, J.P.; Duisit, G.; Rougeau, N.; Mercier, J.; Finzi, A.; Cohen, E.A. HIV-1 Vpr-mediated G2 arrest involves the DDB1-CUL4AVPRBP E3 ubiquitin ligase. PLoS Pathog. 2007, 3, e85. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; DeLucia, M.; Ahn, J. SLX4-SLX1 orotein-independent down-regulation of MUS81-EME1 protein by HIV-1 viral protein R (Vpr). J. Biol. Chem. 2016, 291, 16936–16947. [Google Scholar] [CrossRef] [PubMed]
- Mashiba, M.; Collins, D.R.; Terry, V.H.; Collins, K.L. Vpr overcomes macrophage-specific restriction of HIV-1 Env expression and virion production. Cell Host Microbe 2014, 16, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Fukumori, T.; Akari, H.; Iida, S.; Hata, S.; Kagawa, S.; Aida, Y.; Koyama, A.H.; Adachi, A. The HIV-1 Vpr displays strong anti-apoptotic activity. FEBS Lett. 1998, 432, 17–20. [Google Scholar] [CrossRef]
- Zhu, Y.; Roshal, M.; Li, F.; Blackett, J.; Planelles, V. Upregulation of survivin by HIV-1 Vpr. Apoptosis 2003, 8, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Jacotot, E.; Ferri, K.F.; El Hamel, C.; Brenner, C.; Druillennec, S.; Hoebeke, J.; Rustin, P.; Metivier, D.; Lenoir, C.; Geuskens, M.; et al. Control of mitochondrial membrane permeabilization by adenine nucleotide translocator interacting with HIV-1 viral protein rR and Bcl-2. J. Exp. Med. 2001, 193, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Vieira, H.L.; Haouzi, D.; El, H.C.; Jacotot, E.; Belzacq, A.S.; Brenner, C.; Kroemer, G. Permeabilization of the mitochondrial inner membrane during apoptosis: Impact of the adenine nucleotide translocator. Cell Death Differ. 2000, 7, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Muthumani, K.; Hwang, D.S.; Desai, B.M.; Zhang, D.; Dayes, N.; Green, D.R.; Weiner, D.B. HIV-1 Vpr induces apoptosis through caspase 9 in T cells and peripheral blood mononuclear cells. J. Biol. Chem. 2002, 277, 37820–37831. [Google Scholar] [CrossRef] [PubMed]
- Rom, I.; Deshmane, S.L.; Mukerjee, R.; Khalili, K.; Amini, S.; Sawaya, B.E. HIV-1 Vpr deregulates calcium secretion in neural cells. Brain Res. 2009, 1275, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Rainaldi, G.; Matarrese, P.; Varano, B.; Rivabene, R.; Columba, S.; Sato, A.; Belardelli, F.; Malorni, W.; Gessani, S. The HIV-1 Vpr protein acts as a negative regulator of apoptosis in a human lymphoblastoid T cell line: Possible implications for the pathogenesis of AIDS. J. Exp. Med. 1998, 187, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Hilton, B.A.; Li, Z.; Musich, P.R.; Wang, H.; Cartwright, B.M.; Serrano, M.; Zhou, X.Z.; Lu, K.P.; Zou, Y. ATR plays a direct antiapoptotic role at mitochondria, which is regulated by prolyl isomerase Pin1. Mol. Cell 2015, 60, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Gragerov, A.; Kopp, J.B.; Stauber, R.H.; Pavlakis, G.N.; Chrousos, G.P. The HIV-1 virion-associated protein Vpr is a coactivator of the human glucocorticoid receptor. J. Exp. Med. 1999, 189, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Vanitharani, R.; Mahalingam, S.; Rafaeli, Y.; Singh, S.P.; Srinivasan, A.; Weiner, D.B.; Ayyavoo, V. HIV-1 Vpr transactivates LTR-directed expression through sequences present within −278 to −176 and increases virus replication in vitro. Virology 2001, 289, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mukherjee, S.; Jia, F.; Narayan, O.; Zhao, L.J. Interaction of virion protein Vpr of human immunodeficiency virus type 1 with cellular transcription factor Sp1 and trans-activation of viral long terminal repeat. J. Biol. Chem. 1995, 270, 25564–25569. [Google Scholar] [CrossRef] [PubMed]
- Agostini, I.; Navarro, J.M.; Rey, F.; Bouhamdan, M.; Spire, B.; Vigne, R.; Sire, J. The human immunodeficiency virus type 1 Vpr transactivator: Cooperation with promoter-bound activator domains and binding to TFIIB. J. Mol. Biol. 1996, 261, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, B.E.; Khalili, K.; Mercer, W.E.; Denisova, L.; Amini, S. Cooperative actions of HIV-1 Vpr and p53 modulate viral gene transcription. J. Biol. Chem. 1998, 273, 20052–20057. [Google Scholar] [CrossRef] [PubMed]
- Selig, L.; Benichou, S.; Rogel, M.E.; Wu, L.I.; Vodicka, M.A.; Sire, J.; Benarous, R.; Emerman, M. Uracil DNA glycosylase specifically interacts with Vpr of both human immunodeficiency virus type 1 and simian immunodeficiency virus of sooty mangabeys, but binding does not correlate with cell cycle arrest. J. Virol. 1997, 71, 4842–4846. [Google Scholar] [PubMed]
- Bouhamdan, M.; Benichou, S.; Rey, F.; Navarro, J.M.; Agostini, I.; Spire, B.; Camonis, J.; Slupphaug, G.; Vigne, R.; Benarous, R.; et al. Human immunodeficiency virus type 1 Vpr protein binds to the uracil DNA glycosylase DNA repair enzyme. J. Virol. 1996, 70, 697–704. [Google Scholar] [PubMed]
- Schrofelbauer, B.; Yu, Q.; Zeitlin, S.G.; Landau, N.R. Human immunodeficiency virus type 1 Vpr induces the degradation of the UNG and SMUG uracil-DNA glycosylases. J. Virol. 2005, 79, 10978–10987. [Google Scholar] [CrossRef] [PubMed]
- Schrofelbauer, B.; Hakata, Y.; Landau, N.R. HIV-1 Vpr function is mediated by interaction with the damage-specific DNA-binding protein DDB1. Proc. Natl. Acad. Sci. USA 2007, 104, 4130–4135. [Google Scholar] [CrossRef] [PubMed]
- Weil, A.F.; Ghosh, D.; Zhou, Y.; Seiple, L.; McMahon, M.A.; Spivak, A.M.; Siliciano, R.F.; Stivers, J.T. Uracil DNA glycosylase initiates degradation of HIV-1 cDNA containing misincorporated dUTP and prevents viral integration. Proc. Natl. Acad. Sci. USA 2013, 110, E448–E457. [Google Scholar] [CrossRef] [PubMed]
- Los Alamos HIV Sequence Database and Analysis Staff. HIV Sequence Compendium 2016; Theoretical Biology and Biophysics Group, Los Alamos National Laboratory: Los Alamos, NM, USA. Available online: https://www.hiv.lanl.gov/content/sequence/HIV/COMPENDIUM/2016compendium.html (accessed on 20 October 2016).
- Barnitz, R.A.; Chaigne-Delalande, B.; Bolton, D.L.; Lenardo, M.J. Exposed hydrophobic residues in human immunodeficiency virus type 1 Vpr helix-1 are important for cell cycle arrest and cell death. PLoS ONE 2011, 6, e24924. [Google Scholar] [CrossRef] [PubMed]
- Somasundaran, M.; Sharkey, M.; Brichacek, B.; Luzuriaga, K.; Emerman, M.; Sullivan, J.L.; Stevenson, M. Evidence for a cytopathogenicity determinant in HIV-1 Vpr. Proc. Natl. Acad. Sci. USA 2002, 99, 9503–9508. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ratner, L. Phosphorylation of human immunodeficiency virus type 1 Vpr regulates cell cycle arrest. J. Virol. 2000, 74, 6520–6527. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M.P.; de Noronha, C.M.; Pearce, D.; Greene, W.C. Human immunodeficiency virus type 1 Vpr contains two leucine-rich helices that mediate glucocorticoid receptor coactivation independently of its effects on G(2) cell cycle arrest. J. Virol. 2000, 74, 8159–8165. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.J.; Subbramanian, R.A.; Rougeau, N.; Boisvert, F.; Bergeron, D.; Cohen, E.A. Mutagenic analysis of human immunodeficiency virus type 1 Vpr: Role of a predicted N-terminal α-helical structure in Vpr nuclear localization and virion incorporation. J. Virol. 1995, 69, 7032–7044. [Google Scholar] [PubMed]
- Jian, H.; Zhao, L.J. Pro-apoptotic activity of HIV-1 auxiliary regulatory protein Vpr is subtype-dependent and potently enhanced by nonconservative changes of the leucine residue at position 64. J. Biol. Chem. 2003, 278, 44326–44330. [Google Scholar] [CrossRef] [PubMed]
- Macreadie, I.G.; Castelli, L.A.; Hewish, D.R.; Kirkpatrick, A.; Ward, A.C.; Azad, A.A. A domain of human immunodeficiency virus type 1 Vpr containing repeated H(S/F)RIG amino acid motifs causes cell growth arrest and structural defects. Proc. Natl. Acad. Sci. USA 1995, 92, 2770–2774. [Google Scholar] [CrossRef] [PubMed]
- Maudet, C.; Bertrand, M.; Le, R.E.; Lahouassa, H.; Ayinde, D.; Nisole, S.; Goujon, C.; Cimarelli, A.; Margottin-Goguet, F.; Transy, C. Molecular insight into how HIV-1 Vpr protein impairs cell growth through two genetically distinct pathways. J. Biol. Chem. 2011, 286, 23742–23752. [Google Scholar] [CrossRef] [PubMed]
- Paxton, W.; Connor, R.I.; Landau, N.R. Incorporation of Vpr into human immunodeficiency virus type 1 virions: Requirement for the p6 region of gag and mutational analysis. J. Virol. 1993, 67, 7229–7237. [Google Scholar] [PubMed]
- Caly, L.; Saksena, N.K.; Piller, S.C.; Jans, D.A. Impaired nuclear import and viral incorporation of Vpr derived from a HIV long-term non-progressor. Retrovirology 2008, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Hadi, K.; Walker, L.A.; Guha, D.; Murali, R.; Watkins, S.C.; Tarwater, P.; Srinivasan, A.; Ayyavoo, V. Human immunodeficiency virus type 1 Vpr polymorphisms associated with progressor and nonprogressor individuals alter Vpr-associated functions. J. Gen. Virol. 2014, 95, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Lum, J.J.; Cohen, O.J.; Nie, Z.; Weaver, J.G.; Gomez, T.S.; Yao, X.J.; Lynch, D.; Pilon, A.A.; Hawley, N.; Kim, J.E.; et al. Vpr R77Q is associated with long-term nonprogressive HIV infection and impaired induction of apoptosis. J. Clin. Investig. 2003, 111, 1547–1554. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, B.E.; Khalili, K.; Rappaport, J.; Serio, D.; Chen, W.; Srinivasan, A.; Amini, S. Suppression of HIV-1 transcription and replication by a Vpr mutant. Gene Ther. 1999, 6, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Tcherepanova, I.; Starr, A.; Lackford, B.; Adams, M.D.; Routy, J.P.; Boulassel, M.R.; Calderhead, D.; Healey, D.; Nicolette, C. The immunosuppressive properties of the HIV Vpr protein are linked to a single highly conserved residue, R90. PLoS ONE 2009, 4, e5853. [Google Scholar] [CrossRef] [PubMed]
- Tzitzivacos, D.B.; Tiemessen, C.T.; Stevens, W.S.; Papathanasopoulos, M.A. Viral genetic determinants of nonprogressive HIV type 1 subtype C infection in antiretroviral drug-naive children. AIDS Res. Hum. Retrovir. 2009, 25, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Kamori, D.; Hasan, Z.; Ohashi, J.; Kawana-Tachikawa, A.; Gatanaga, H.; Oka, S.; Ueno, T. Identification of two unique naturally occurring Vpr sequence polymorphisms associated with clinical parameters in HIV-1 chronic infection. J. Med. Virol. 2017, 89, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Dampier, W.; Antell, G.C.; Aiamkitsumrit, B.; Nonnemacher, M.R.; Jacobson, J.M.; Pirrone, V.; Zhong, W.; Kercher, K.; Passic, S.; Williams, J.W.; et al. Specific amino acids in HIV-1 Vpr are significantly associated with differences in patient neurocognitive status. J. Neurovirol. 2016. [Google Scholar] [CrossRef]
- Collins, D.R.; Lubow, J.; Lukic, Z.; Mashiba, M.; Collins, K.L. Vpr promotes macrophage-dependent HIV-1 infection of CD4+ T lymphocytes. PLoS Pathog. 2015, 11, e1005054. [Google Scholar] [CrossRef] [PubMed]
- Mologni, D.; Citterio, P.; Menzaghi, B.; Zanone, P.B.; Riva, C.; Broggini, V.; Sinicco, A.; Milazzo, L.; Adorni, F.; Rusconi, S.; et al. Vpr and HIV-1 disease progression: R77Q mutation is associated with long-term control of HIV-1 infection in different groups of patients. AIDS 2006, 20, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Cavert, W.; Webb, C.H.; Balfour, H.H. Alterations in the C-terminal region of the HIV-1 accessory gene vpr do not confer clinical advantage to subjects receiving nucleoside antiretroviral therapy. J. Infect. Dis. 2004, 189, 2181–2184. [Google Scholar] [CrossRef] [PubMed]
- Chui, C.; Cheung, P.K.; Brumme, C.J.; Mo, T.; Brumme, Z.L.; Montaner, J.S.; Badley, A.D.; Harrigan, P.R. HIV VprR77Q mutation does not influence clinical response of individuals initiating highly active antiretroviral therapy. AIDS Res. Hum. Retrovir. 2006, 22, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, G.; Le, R.E.; Maidou-Peindara, P.; Maizy, M.; Lefrere, J.J.; Daneluzzi, V.; Monteiro-Filho, C.M.; Hong, D.; Planelles, V.; Morand-Joubert, L.; et al. Characterization of the molecular determinants of primary HIV-1 Vpr proteins: Impact of the Q65R and R77Q substitutions on Vpr functions. PLoS ONE 2009, 4, e7514. [Google Scholar] [CrossRef] [PubMed]
- Yedavalli, V.R.; Chappey, C.; Ahmad, N. Maintenance of an intact human immunodeficiency virus type 1 vpr gene following mother-to-infant transmission. J. Virol. 1998, 72, 6937–6943. [Google Scholar] [PubMed]
- Romani, B.; Glashoff, R.H.; Engelbrecht, S. Functional integrity of naturally occurring mutants of HIV-1 subtype C Vpr. Virus Res. 2010, 153, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H.; Sarver, N. HIV accessory proteins as therapeutic targets. Nat. Med. 1997, 3, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.E. Vpu protein: The viroporin encoded by HIV-1. Viruses 2015, 7, 4352–4368. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Ao, Z.; Yao, X. Apobec3G-based strategies to defeat HIV infection. Curr. HIV Res. 2016, 14, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Smithgall, T.E.; Thomas, G. Small molecule inhibitors of the HIV-1 virulence factor, Nef. Drug Discov. Today Technol. 2013, 10, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Weydert, C.; Rijck, J.D.; Christ, F.; Debyser, Z. Targeting virus-host interactions of HIV replication. Curr. Top. Med. Chem. 2016, 16, 1167–1190. [Google Scholar] [CrossRef] [PubMed]
- Poon, B.; Grovit-Ferbas, K.; Stewart, S.A.; Chen, I.S. Cell cycle arrest by Vpr in HIV-1 virions and insensitivity to antiretroviral agents. Science 1998, 281, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Gavegnano, C.; Schinazi, R.F. Antiretroviral therapy in macrophages: Implication for HIV eradication. Antivir. Chem. Chemother. 2009, 20, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Herbein, G. The macrophage: A therapeutic target in HIV-1 infection. Mol. Cell Ther. 2014, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Serio, D.; Rizvi, T.A.; Cartas, M.; Kalyanaraman, V.S.; Weber, I.T.; Koprowski, H.; Srinivasan, A. Development of a novel anti-HIV-1 agent from within: Effect of chimeric Vpr-containing protease cleavage site residues on virus replication. Proc. Natl. Acad. Sci. USA 1997, 94, 3346–3351. [Google Scholar] [CrossRef] [PubMed]
- Poon, B.; Jowett, J.B.; Stewart, S.A.; Armstrong, R.W.; Rishton, G.M.; Chen, I.S. Human immunodeficiency virus type 1 vpr gene induces phenotypic effects similar to those of the DNA alkylating agent, nitrogen mustard. J. Virol. 1997, 71, 3961–3971. [Google Scholar] [PubMed]
- Kamata, M.; Wu, R.P.; An, D.S.; Saxe, J.P.; Damoiseaux, R.; Phelps, M.E.; Huang, J.; Chen, I.S. Cell-based chemical genetic screen identifies damnacanthal as an inhibitor of HIV-1 Vpr induced cell death. Biochem. Biophys. Res. Commun. 2006, 348, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Win, N.N.; Ito, T.; Matsui, T.; Aimaiti, S.; Kodama, T.; Ngwe, H.; Okamoto, Y.; Tanaka, M.; Asakawa, Y.; Abe, I.; et al. Isopimarane diterpenoids from Kaempferia pulchra rhizomes collected in Myanmar and their Vpr inhibitory activity. Bioorg. Med. Chem. Lett. 2016, 26, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Shimura, M.; Zhou, Y.; Asada, Y.; Yoshikawa, T.; Hatake, K.; Takaku, F.; Ishizaka, Y. Inhibition of Vpr-induced cell cycle abnormality by quercetin: A novel strategy for searching compounds targeting Vpr. Biochem. Biophys. Res. Commun. 1999, 261, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Nishihara, Y.; Yamaguchi, T.; Koito, A.; Miyoshi, H.; Kakeya, H.; Osada, H. Fumagillin suppresses HIV-1 infection of macrophages through the inhibition of Vpr activity. FEBS Lett. 2006, 580, 2598–2602. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Kopp, J.B.; Chrousos, G.P. Glucocorticoids suppress human immunodeficiency virus type-1 long terminal repeat activity in a cell type-specific, glucocorticoid receptor-mediated fashion: Direct protective effects at variance with clinical phenomenology. J. Steroid Biochem. Mol. Biol. 2000, 75, 283–290. [Google Scholar] [CrossRef]
- Refaeli, Y.; Levy, D.N.; Weiner, D.B. The glucocorticoid receptor type II complex is a target of the HIV-1 vpr gene product. Proc. Natl. Acad. Sci. USA 1995, 92, 3621–3625. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.J.; Lemay, J.; Rougeau, N.; Clement, M.; Kurtz, S.; Belhumeur, P.; Cohen, E.A. Genetic selection of peptide inhibitors of human immunodeficiency virus type 1 Vpr. J. Biol. Chem. 2002, 277, 48816–48826. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, N.; Nonaka, M.; Hashimoto, Y.; Matsuda, G.; Takeshima, S.N.; Matsuyama, M.; Igarashi, T.; Miura, T.; Tanaka, R.; et al. Inhibition of human immunodeficiency virus type 1 (HIV-1) nuclear import via Vpr-importin alpha interactions as a novel HIV-1 therapy. Biochem. Biophys. Res. Commun. 2009, 380, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, K.; Ishii, H.; Murakami, T.; Takeshima, S.N.; Chutiwitoonchai, N.; Kodama, E.N.; Kawaji, K.; Kondoh, Y.; Honda, K.; Osada, H.; et al. Synthesis of a Vpr-binding derivative for use as a novel HIV-1 inhibitor. PLoS ONE 2015, 10, e0145573. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, K.; Murakami, T.; Xue, G.; Shimizu, Y.; Takeda, E.; Hashimoto, Y.; Honda, K.; Kondoh, Y.; Osada, H.; Tsunetsugu-Yokota, Y.; et al. Identification of a novel Vpr-binding compound that inhibits HIV-1 multiplication in macrophages by chemical array. Biochem. Biophys. Res. Commun. 2010, 403, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Ong, E.B.; Watanabe, N.; Saito, A.; Futamura, Y.; Abd El Galil, K.H.; Koito, A.; Najimudin, N.; Osada, H. Vipirinin, a coumarin-based HIV-1 Vpr inhibitor, interacts with a hydrophobic region of Vpr. J. Biol. Chem. 2011, 286, 14049–14056. [Google Scholar] [CrossRef] [PubMed]
- Mamik, M.K.; Hui, E.; Branton, W.G.; McKenzie, B.A.; Chisholm, J.; Cohen, E.A.; Power, C. HIV-1 viral protein R activates NLRP3 inflammasome in microglia: Implications for HIV-1 associated neuroinflammation. J. Neuroimmune Pharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, H.; Miura, Y.; Ando, Y.; Hoshino, S.; Ishizaka, Y.; Koyanagi, Y. Human immunodeficiency virus type 1 Vpr inhibits axonal outgrowth through induction of mitochondrial dysfunction. J. Virol. 2008, 82, 2528–2542. [Google Scholar] [CrossRef] [PubMed]
- James, T.; Nonnemacher, M.R.; Wigdahl, B.; Krebs, F.C. Defining the roles for Vpr in HIV-1-associated neuropathogenesis. J. Neurovirol. 2016, 22, 403–415. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Function | Cellular Protein | Effect | Reference |
|---|---|---|---|
| Mitochondria/reactive oxigen species (ROS) | ANT | + | [83] |
| VDAC | + | [83] | |
| PMCA | − | [138] | |
| GLUD2 | − | [94] | |
| Glucose metabolism | HK1 | + | [94] |
| Pentose phosphate pathway | G6PDH | + | [94] |
| Glycolysis | GAPDH | − | [94] |
| Stress response | ATR | + | [117] |
| Proteasome | DCAF1 | + | [128] |
| Transcription | NFAT | + | [10] |
| NF-ĸB | +/− | [79]/[39] | |
| C/EBP | + | [79] | |
| AP-1 | + | [79] | |
| HIF-1α | + | [114] | |
| HDAC1 | − | [122] | |
| GR | + | [141] | |
| SP1 | + | [143] | |
| DNA metabolism | SLX4 | + | [66] |
| HLTF | − | [59] | |
| UNG2 | − | [148] |
© 2017 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, M.E. The HIV-1 Vpr Protein: A Multifaceted Target for Therapeutic Intervention. Int. J. Mol. Sci. 2017, 18, 126. https://doi.org/10.3390/ijms18010126
González ME. The HIV-1 Vpr Protein: A Multifaceted Target for Therapeutic Intervention. International Journal of Molecular Sciences. 2017; 18(1):126. https://doi.org/10.3390/ijms18010126
Chicago/Turabian StyleGonzález, María Eugenia. 2017. "The HIV-1 Vpr Protein: A Multifaceted Target for Therapeutic Intervention" International Journal of Molecular Sciences 18, no. 1: 126. https://doi.org/10.3390/ijms18010126