
Incretin-Based Therapies for Diabetic Complications: Basic Mechanisms and Clinical Evidence



Abstract
:
1. Introduction
2. Incretins as Therapeutic Targets of Diabetes
3. Organ Protective Effects of Incretins
4. Effects of Incretin-Based Therapies on Diabetic Nephropathy
4.1. Experimental Studies
4.2. Clinical Studies
5. Effects of Incretin-Based Agents on Diabetic Retinopathy
5.1. Experimental Studies
5.2. Clinical Studies
6. Effects of Incretin-Based Agents on Diabetic Neuropathy
6.1. Experimental Studies
6.2. Clinical Studies
7. Diabetic Macrovascular Complications
7.1. Experimental Studies
7.2. Clinical Studies
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Joao, A.L.; Reis, F.; Fernandes, R. The incretin system ABCs in obesity and diabetes-novel therapeutic strategies for weight loss and beyond. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2016, 17, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A. Incretin-based therapies for type 2 diabetes mellitus: Properties, functions, and clinical implications. Am. J. Med. 2011, 124 (Suppl. 1), S3–S18. [Google Scholar] [CrossRef] [PubMed]
- Chia, C.W.; Carlson, O.D.; Kim, W.; Shin, Y.K.; Charles, C.P.; Kim, H.S.; Melvin, D.L.; Egan, J.M. Exogenous glucose-dependent insulinotropic polypeptide worsens post prandial hyperglycemia in type 2 diabetes. Diabetes 2009, 58, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Heimesaat, M.M.; Behle, K.; Holst, J.J.; Nauck, M.S.; Ritzel, R.; Hufner, M.; Schmiegel, W.H. Effects of glucagon-like peptide 1 on counterregulatory hormone responses, cognitive functions, and insulin secretion during hyperinsulinemic, stepped hypoglycemic clamp experiments in healthy volunteers. J. Clin. Endocrinol. Metab. 2002, 87, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Orskov, C.; Holst, J.J.; Nielsen, O.V. Effect of truncated glucagon-like peptide-1 (proglucagon-(78-107) amide) on endocrine secretion from pig pancreas, antrum, and nonantral stomach. Endocrinology 1988, 123, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- De Heer, J.; Rasmussen, C.; Coy, D.H.; Holst, J.J. Glucagon-like peptide-1, but not glucose-dependent insulinotropic peptide, inhibits glucagon secretion via somatostatin (receptor subtype 2) in the perfused rat pancreas. Diabetologia 2008, 51, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Seino, Y.; Yabe, D. Glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1: Incretin actions beyond the pancreas. J. Diabetes Investig. 2013, 4, 108–130. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, G.; Zhong, J.; Peters, M.; Rajagopalan, S. Incretin-based therapy for diabetes: What a cardiologist needs to know. J. Am. Coll. Cardiol. 2016, 67, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Rajagopalan, S. Dipeptidyl peptidase-4 regulation of SDF-1/CXCR4 axis: Implications for cardiovascular disease. Front. Immunol. 2015, 6, 477. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Kanasaki, K.; Koya, D. Linagliptin but not Sitagliptin inhibited transforming growth factor-β2-induced endothelial DPP-4 activity and the endothelial-mesenchymal transition. Biochem. Biophys. Res. Commun. 2016, 471, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Deacon, C.F. What do we know about the secretion and degradation of incretin hormones? Regul. Pept. 2005, 128, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Knop, F.K.; Vilsboll, T.; Hojberg, P.V.; Larsen, S.; Madsbad, S.; Volund, A.; Holst, J.J.; Krarup, T. Reduced incretin effect in type 2 diabetes: Cause or consequence of the diabetic state? Diabetes 2007, 56, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Knop, F.K.; Aaboe, K.; Vilsboll, T.; Volund, A.; Holst, J.J.; Krarup, T.; Madsbad, S. Impaired incretin effect and fasting hyperglucagonaemia characterizing type 2 diabetic subjects are early signs of dysmetabolism in obesity. Diabetes Obes. Metab. 2012, 14, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Tura, A.; Muscelli, E.; Gastaldelli, A.; Ferrannini, E.; Mari, A. Altered pattern of the incretin effect as assessed by modelling in individuals with glucose tolerance ranging from normal to diabetic. Diabetologia 2014, 57, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Ryskjaer, J.; Deacon, C.F.; Carr, R.D.; Krarup, T.; Madsbad, S.; Holst, J.; Vilsboll, T. Plasma dipeptidyl peptidase-IV activity in patients with type-2 diabetes mellitus correlates positively with HbAlc levels, but is not acutely affected by food intake. Eur. J. Endocrinol. Eur. 2006, 155, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, E.; Pala, L.; Ciani, S.; Bardini, G.; Pezzatini, A.; Sposato, I.; Cremasco, F.; Ognibene, A.; Rotella, C.M. Hyperglycaemia increases dipeptidyl peptidase IV activity in diabetes mellitus. Diabetologia 2005, 48, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Osawa, S.; Kawamori, D.; Katakami, N.; Takahara, M.; Sakamoto, F.; Katsura, T.; Yasuda, T.; Kaneto, H.; Matsuhisa, M.; Matsuoka, T.A.; et al. Significant elevation of serum dipeptidyl peptidase-4 activity in young-adult type 1 diabetes. Diabetes Res. Clin. Pract. 2016, 113, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Heimesaat, M.M.; Orskov, C.; Holst, J.J.; Ebert, R.; Creutzfeldt, W. Preserved incretin activity of glucagon-like peptide 1 (7-36 amide) but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J. Clin. Investig. 1993, 91, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Vilsboll, T.; Krarup, T.; Madsbad, S.; Holst, J.J. Defective amplification of the late phase insulin response to glucose by GIP in obese Type II diabetic patients. Diabetologia 2002, 45, 1111–1119. [Google Scholar] [PubMed]
- Nauck, M.A.; Homberger, E.; Siegel, E.G.; Allen, R.C.; Eaton, R.P.; Ebert, R.; Creutzfeldt, W. Incretin effects of increasing glucose loads in man calculated from venous insulin and C-peptide responses. J. Clin. Endocrinol. Metab. 1986, 63, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Kawanami, D.; Matoba, K.; Utsunomiya, K. Signaling pathways in diabetic nephropathy. Histol. Histopathol. 2016, 11777. [Google Scholar]
- Inoue, T.; Inoguchi, T.; Sonoda, N.; Hendarto, H.; Makimura, H.; Sasaki, S.; Yokomizo, H.; Fujimura, Y.; Miura, D.; Takayanagi, R. GLP-1 analog liraglutide protects against cardiac steatosis, oxidative stress and apoptosis in streptozotocin-induced diabetic rats. Atherosclerosis 2015, 240, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, Y.; Matsui, T.; Takeuchi, M.; Yamagishi, S. Glucagon-like peptide-1 (GLP-1) inhibits advanced glycation end product (AGE)-induced up-regulation of VCAM-1 mRNA levels in endothelial cells by suppressing AGE receptor (RAGE) expression. Biochem. Biophys. Res. Commun. 2010, 391, 1405–1408. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, Y.; Nishino, Y.; Matsui, T.; Takeuchi, M.; Yamagishi, S. Glucagon-like peptide-1 suppresses advanced glycation end product-induced monocyte chemoattractant protein-1 expression in mesangial cells by reducing advanced glycation end product receptor level. Metab. Clin. Exp. 2011, 60, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Luo, P.; Wang, Y.; Li, W.; Wang, C.; Sun, D.; Zhang, R.; Su, T.; Ma, X.; Zeng, C.; et al. Glucagon-like peptide-1 protects against cardiac microvascular injury in diabetes via a cAMP/PKA/Rho-dependent mechanism. Diabetes 2013, 62, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Park, M.S.; Choung, J.S.; Kim, S.S.; Oh, H.H.; Choi, C.S.; Ha, S.Y.; Kang, Y.; Kim, Y.; Jun, H.S. Glucagon-like peptide-1 inhibits adipose tissue macrophage infiltration and inflammation in an obese mouse model of diabetes. Diabetologia 2012, 55, 2456–2468. [Google Scholar] [CrossRef] [PubMed]
- Kodera, R.; Shikata, K.; Kataoka, H.U.; Takatsuka, T.; Miyamoto, S.; Sasaki, M.; Kajitani, N.; Nishishita, S.; Sarai, K.; Hirota, D.; et al. Glucagon-like peptide-1 receptor agonist ameliorates renal injury through its anti-inflammatory action without lowering blood glucose level in a rat model of type 1 diabetes. Diabetologia 2011, 54, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Salles, T.A.; dos Santos, L.; Barauna, V.G.; Girardi, A.C. Potential role of dipeptidyl peptidase IV in the pathophysiology of heart failure. Int. J. Mol. Sci. 2015, 16, 4226–4249. [Google Scholar] [CrossRef] [PubMed]
- Boerrigter, G.; Costello-Boerrigter, L.C.; Harty, G.J.; Lapp, H.; Burnett, J.C., Jr. Des-serine-proline brain natriuretic peptide 3–32 in cardiorenal regulation. Am. J. Physiol. Regul. Integ. Comp. Physiol. 2007, 292, R897–R901. [Google Scholar] [CrossRef] [PubMed]
- Grouzmann, E.; Bigliardi, P.; Appenzeller, M.; Pannatier, A.; Buclin, T. Substance P-induced skin inflammation is not modulated by a single dose of sitagliptin in human volunteers. Biol. Chem. 2011, 392, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Mentlein, R.; Dahms, P.; Grandt, D.; Kruger, R. Proteolytic processing of neuropeptide Y and peptide YY by dipeptidyl peptidase IV. Regul. Pept. 1993, 49, 133–144. [Google Scholar] [CrossRef]
- Shioda, T.; Kato, H.; Ohnishi, Y.; Tashiro, K.; Ikegawa, M.; Nakayama, E.E.; Hu, H.; Kato, A.; Sakai, Y.; Liu, H.; et al. Anti-HIV-1 and chemotactic activities of human stromal cell-derived factor 1α (SDF-1α) and SDF-1β are abolished by CD26/dipeptidyl peptidase IV-mediated cleavage. Proc. Natl. Acad. Sci. USA 1998, 95, 6331–6336. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, C.; Di Carlo, A.; Facchiano, F.; Senatore, C.; De Cristofaro, R.; Luzi, A.; Federici, M.; Romani, M.; Napolitano, M.; Capogrossi, M.C.; et al. High mobility group box 1 is a novel substrate of dipeptidyl peptidase-IV. Diabetologia 2012, 55, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Makino, Y.; Fujita, Y.; Haneda, M. Dipeptidyl peptidase-4 inhibitors in progressive kidney disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Hocher, B.; Sharkovska, Y.; Mark, M.; Klein, T.; Pfab, T. The novel DPP-4 inhibitors linagliptin and BI 14361 reduce infarct size after myocardial ischemia/reperfusion in rats. Int. J. Cardiol. 2013, 167, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, H.; Binder, R.; Greischel, A. Tissue distribution of the novel DPP-4 inhibitor BI 1356 is dominated by saturable binding to its target in rats. Biopharm. Drug Dispos. 2009, 30, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Mega, C.; de Lemos, E.T.; Vala, H.; Fernandes, R.; Oliveira, J.; Mascarenhas-Melo, F.; Teixeira, F.; Reis, F. Diabetic nephropathy amelioration by a low-dose sitagliptin in an animal model of type 2 diabetes (Zucker diabetic fatty rat). Exp. Diabetes Res. 2011, 2011, 162092. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Mega, C.; Goncalves, A.; Rodrigues-Santos, P.; Teixeira-Lemos, E.; Teixeira, F.; Fontes-Ribeiro, C.; Reis, F.; Fernandes, R. Sitagliptin prevents inflammation and apoptotic cell death in the kidney of type 2 diabetic animals. Mediat. Inflamm. 2014, 2014, 538737. [Google Scholar] [CrossRef] [PubMed]
- Kodera, R.; Shikata, K.; Takatsuka, T.; Oda, K.; Miyamoto, S.; Kajitani, N.; Hirota, D.; Ono, T.; Usui, H.K.; Makino, H. Dipeptidyl peptidase-4 inhibitor ameliorates early renal injury through its anti-inflammatory action in a rat model of type 1 diabetes. Bioche. Biophys. Res. Commun. 2014, 443, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, K.; Shi, S.; Kanasaki, M.; He, J.; Nagai, T.; Nakamura, Y.; Ishigaki, Y.; Kitada, M.; Srivastava, S.P.; Koya, D. Linagliptin-mediated DPP-4 inhibition ameliorates kidney fibrosis in streptozotocin-induced diabetic mice by inhibiting endothelial-to-mesenchymal transition in a therapeutic regimen. Diabetes 2014, 63, 2120–2131. [Google Scholar] [CrossRef] [PubMed]
- Hendarto, H.; Inoguchi, T.; Maeda, Y.; Ikeda, N.; Zheng, J.; Takei, R.; Yokomizo, H.; Hirata, E.; Sonoda, N.; Takayanagi, R. GLP-1 analog liraglutide protects against oxidative stress and albuminuria in streptozotocin-induced diabetic rats via protein kinase A-mediated inhibition of renal NAD(P)H oxidases. Metab. Clin. Exp. 2012, 61, 1422–1434. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Marques, C.; Leal, E.; Ribeiro, C.F.; Reis, F.; Ambrosio, A.F.; Fernandes, R. Dipeptidyl peptidase-IV inhibition prevents blood-retinal barrier breakdown, inflammation and neuronal cell death in the retina of type 1 diabetic rats. Biochim. Biophys. Acta 2014, 1842, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Leal, E.; Paiva, A.; Teixeira Lemos, E.; Teixeira, F.; Ribeiro, C.F.; Reis, F.; Ambrosio, A.F.; Fernandes, R. Protective effects of the dipeptidyl peptidase IV inhibitor sitagliptin in the blood-retinal barrier in a type 2 diabetes animal model. Diabetes Obes. Metab. 2012, 14, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Yamagishi, S.; Matsui, T.; Nakashima, S.; Ojima, A.; Maeda, S.; Nishino, Y.; Ishibashi, Y.; Yoshida, Y.; Yamakawa, R. Beneficial effects of vildagliptin on retinal injury in obese type 2 diabetic rats. Ophthalmic Res. 2013, 50, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.; Bogdanov, P.; Corraliza, L.; Garcia-Ramirez, M.; Sola-Adell, C.; Arranz, J.A.; Arroba, A.I.; Valverde, A.M.; Simo, R. Topical Administration of GLP-1 receptor agonists prevents retinal neurodegeneration in experimental diabetes. Diabetes 2016, 65, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.Y.; Liu, W.J.; Park, J.H.; Baek, H.S.; Park, T.S. Effect of dipeptidyl peptidase-IV (DPP-IV) inhibitor (Vildagliptin) on peripheral nerves in streptozotocin-induced diabetic rats. Arch. Med. Res. 2009, 40, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, R.; Cervellini, I.; Porretta-Serapiglia, C.; Oggioni, N.; Burkey, B.; Ghezzi, P.; Cavaletti, G.; Lauria, G. Beneficial effects of PKF275–055, a novel, selective, orally bioavailable, long-acting dipeptidyl peptidase IV inhibitor in streptozotocin-induced diabetic peripheral neuropathy. J. Pharmacol. Exp. Ther. 2012, 340, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Himeno, T.; Kamiya, H.; Naruse, K.; Harada, N.; Ozaki, N.; Seino, Y.; Shibata, T.; Kondo, M.; Kato, J.; Okawa, T.; et al. Beneficial effects of exendin-4 on experimental polyneuropathy in diabetic mice. Diabetes 2011, 60, 2397–2406. [Google Scholar] [CrossRef] [PubMed]
- Jolivalt, C.G.; Fineman, M.; Deacon, C.F.; Carr, R.D.; Calcutt, N.A. GLP-1 signals via ERK in peripheral nerve and prevents nerve dysfunction in diabetic mice. Diabetes Obes. Metab. 2011, 13, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Ta, N.N.; Schuyler, C.A.; Li, Y.; Lopes-Virella, M.F.; Huang, Y. DPP-4 (CD26) inhibitor alogliptin inhibits atherosclerosis in diabetic apolipoprotein E-deficient mice. J. Cardiovasc. Pharmacol. 2011, 58, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Takai, S.; Sakonjo, H.; Jin, D. Significance of vascular dipeptidyl peptidase-4 inhibition on vascular protection in Zucker diabetic fatty rats. J. Pharmacol. Sci. 2014, 125, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.T.; Zhang, Q.; Tang, H.Q.; Wang, C.J.; Su, H.; Zhou, Q.; Wei, W.; Zhu, H.Q.; Wang, Y. Effects of glucagon-like peptide-1 on advanced glycation endproduct-induced aortic endothelial dysfunction in streptozotocin-induced diabetic rats: Possible roles of Rho kinase- and AMP kinase-mediated nuclear factor κB signaling pathways. Endocrine 2016, 53, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Hattori, S. Sitagliptin reduces albuminuria in patients with type 2 diabetes. Endocr. J. 2011, 58, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Okada, Y.; Arao, T.; Tanaka, Y. Sitagliptin improves albuminuria in patients with type 2 diabetes mellitus. J. Diabetes Investig. 2014, 5, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Scirica, B.M.; Bhatt, D.L.; Braunwald, E.; Steg, P.G.; Davidson, J.; Hirshberg, B.; Ohman, P.; Frederich, R.; Wiviott, S.D.; Hoffman, E.B.; et al. Saxagliptin and cardiovascular outcomes in patients with type 2 diabetes mellitus. N. Engl. J. Med. 2013, 369, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Groop, P.H.; Cooper, M.E.; Perkovic, V.; Emser, A.; Woerle, H.J.; von Eynatten, M. Linagliptin lowers albuminuria on top of recommended standard treatment in patients with type 2 diabetes and renal dysfunction. Diabetes Care 2013, 36, 3460–3468. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Taniai, H.; Murayama, H.; Ohshiro, H.; Hayashi, H.; Sato, S.; Kikuchi, N.; Komatsu, T.; Komatsu, K.; Komatsu, K.; et al. DPP-4 inhibition with alogliptin on top of angiotensin II type 1 receptor blockade ameliorates albuminuria via up-regulation of SDF-1α in type 2 diabetic patients with incipient nephropathy. Endocr. J. 2014, 61, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Zavattaro, M.; Caputo, M.; Sama, M.T.; Mele, C.; Chasseur, L.; Marzullo, P.; Pagano, L.; Mauri, M.G.; Ponziani, M.C.; Aimaretti, G.; et al. One-year treatment with liraglutide improved renal function in patients with type 2 diabetes: A pilot prospective study. Endocrine 2015, 50, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Hu, C.; Lu, W. Exenatide reduces urinary transforming growth factor-β1 and type IV collagen excretion in patients with type 2 diabetes and microalbuminuria. Kidney Blood Press. Res. 2012, 35, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Marso, S.P.; Daniels, G.H.; Brown-Frandsen, K.; Kristensen, P.; Mann, J.F.; Nauck, M.A.; Nissen, S.E.; Pocock, S.; Poulter, N.R.; Ravn, L.S.; et al. Liraglutide and cardiovascular outcomes in type 2 diabetes. N. Engl. J. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Raff, U.; Schmidt, S.; Kistner, I.; Friedrich, S.; Bramlage, P.; Harazny, J.M.; Schmieder, R.E. Effects of saxagliptin on early microvascular changes in patients with type 2 diabetes. Cardiovasc. Diabetol. 2014, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Varadhan, L.; Humphreys, T.; Hariman, C.; Walker, A.B.; Varughese, G.I. GLP-1 agonist treatment: Implications for diabetic retinopathy screening. Diabetes Res. Clin. Pract. 2011, 94, e68–e71. [Google Scholar] [CrossRef] [PubMed]
- Varadhan, L.; Humphreys, T.; Walker, A.B.; Varughese, G.I. The impact of improved glycaemic control with GLP-1 receptor agonist therapy on diabetic retinopathy. Diabetes Res. Clin. Pract. 2014, 103, e37–e39. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.; Martin, C.L.; Brown, M.B.; Callaghan, B.; Albers, J.W.; Feldman, E.L.; Pop-Busui, R. Effects of exenatide on measures of diabetic neuropathy in subjects with type 2 diabetes: Results from an 18-month proof-of-concept open-label randomized study. J. Diabetes Complic. 2015, 29, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- White, W.B.; Cannon, C.P.; Heller, S.R.; Nissen, S.E.; Bergenstal, R.M.; Bakris, G.L.; Perez, A.T.; Fleck, P.R.; Mehta, C.R.; Kupfer, S.; et al. Alogliptin after acute coronary syndrome in patients with type 2 diabetes. N. Engl. J. Med. 2013, 369, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Green, J.B.; Bethel, M.A.; Armstrong, P.W.; Buse, J.B.; Engel, S.S.; Garg, J.; Josse, R.; Kaufman, K.D.; Koglin, J.; Korn, S.; et al. Effect of sitagliptin on cardiovascular outcomes in type 2 diabetes. N. Engl. J. Med. 2015, 373, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.A.; Claggett, B.; Diaz, R.; Dickstein, K.; Gerstein, H.C.; Kober, L.V.; Lawson, F.C.; Ping, L.; Wei, X.; Lewis, E.F.; et al. Lixisenatide in patients with type 2 diabetes and acute coronary syndrome. N. Engl. J. Med. 2015, 373, 2247–2257. [Google Scholar] [CrossRef] [PubMed]
- Crajoinas, R.O.; Oricchio, F.T.; Pessoa, T.D.; Pacheco, B.P.; Lessa, L.M.; Malnic, G.; Girardi, A.C. Mechanisms mediating the diuretic and natriuretic actions of the incretin hormone glucagon-like peptide-1. Am. J. Physiol. Ren. Physiol. 2011, 301, F355–F363. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Morii, T.; Fujishima, H.; Sato, T.; Shimizu, T.; Hosoba, M.; Tsukiyama, K.; Narita, T.; Takahashi, T.; Drucker, D.J.; et al. The protective roles of GLP-1R signaling in diabetic nephropathy: Possible mechanism and therapeutic potential. Kidney Int. 2014, 85, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Korner, M.; Stockli, M.; Waser, B.; Reubi, J.C. GLP-1 receptor expression in human tumors and human normal tissues: Potential for in vivo targeting. J. Nucl. Med. 2007, 48, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, P.; Beglinger, C.; Drewe, J.; Gutmann, H. Glucagon-like peptide 1 receptor expression in primary porcine proximal tubular cells. Regul. Pept. 2007, 141, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Campitelli, J.; Hu, G.; Lin, Y.; Luo, J.; Xue, C. Increase in DPP-IV in the intestine, liver and kidney of the rat treated with high fat diet and streptozotocin. Life Sci. 2007, 81, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Xie, S.H.; Liu, Y.N.; Kim, W.; Jin, H.Y.; Park, S.K.; Shao, Y.M.; Park, T.S. Dipeptidyl peptidase IV inhibitor attenuates kidney injury in streptozotocin-induced diabetic rats. J. Pharmacol. Exp. Ther. 2012, 340, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Mima, A.; Hiraoka-Yamomoto, J.; Li, Q.; Kitada, M.; Li, C.; Geraldes, P.; Matsumoto, M.; Mizutani, K.; Park, K.; Cahill, C.; et al. Protective effects of GLP-1 on glomerular endothelium and its inhibition by PKCβ activation in diabetes. Diabetes 2012, 61, 2967–2979. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, S.; Villar, V.A.; Jose, P.A.; Armando, I. Renal dopamine receptors, oxidative stress, and hypertension. Int. J. Mol. Sci. 2013, 14, 17553–17572. [Google Scholar] [CrossRef] [PubMed]
- Udell, J.A.; Bhatt, D.L.; Braunwald, E.; Cavender, M.A.; Mosenzon, O.; Steg, P.G.; Davidson, J.A.; Nicolau, J.C.; Corbalan, R.; Hirshberg, E.B.; et al. Saxagliptin and cardiovascular outcomes in patients with type 2 diabetes and moderate or severe renal impairment: Observations from the SAVOR-TIMI 53 Trial. Diabetes Care 2015, 38, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Groop, P.H.; Cooper, M.E.; Perkovic, V.; Sharma, K.; Schernthaner, G.; Haneda, M.; Hocher, B.; Gordat, M.; Cescutti, J.; Woerle, H.J.; et al. Dipeptidyl peptidase-4 inhibition with linagliptin and effects on hyperglycaemia and albuminuria in patients with type 2 diabetes and renal dysfunction: Rationale and design of the MARLINA-T2D trial. Diabetes Vasc. Dis. Res. 2015, 12, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Von Scholten, B.J.; Hansen, T.W.; Goetze, J.P.; Persson, F.; Rossing, P. Glucagon-like peptide 1 receptor agonist (GLP-1 RA): Long-term effect on kidney function in patients with type 2 diabetes. J. Diabetes Complicat. 2015, 29, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Boulton, M. The pathogenesis of diabetic retinopathy: Old concepts and new questions. Eye 2002, 16, 242–260. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Ambrosio, A.F.; Fernandes, R. Regulation of claudins in blood-tissue barriers under physiological and pathological states. Tissue Barriers 2013, 1, e24782. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.I.; Park, M.J.; Choi, J.H.; Lim, S.K.; Choi, H.J.; Park, S.H. Hyperglycemia-induced GLP-1R downregulation causes RPE cell apoptosis. Int. J. Biochem. Cell Biol. 2015, 59, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Lin, C.M.; Muthusamy, A.; Fontes-Ribeiro, C.; Ambrosio, A.F.; Abcouwer, S.F.; Fernandes, R.; Antonetti, D.A. Protective Effect of a GLP-1 Analog on ischemia-reperfusion induced blood-retinal barrier breakdown and inflammation. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2584–2592. [Google Scholar] [CrossRef] [PubMed]
- Sango, K.; Utsunomiya, K. Efficacy of glucagon-like peptide-1 mimetics for neural regeneration. Neural Regen. Res. 2015, 10, 1723–1724. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, M.; Niimi, N.; Sango, K.; Takaku, S.; Kanazawa, Y.; Utsunomiya, K. Neurotrophic and neuroprotective properties of exendin-4 in adult rat dorsal root ganglion neurons: Involvement of insulin and RhoA. Histochem. Cell Biol. 2015, 144, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Yorek, M.A.; Coppey, L.J.; Gellett, J.S.; Davidson, E.P. Sensory nerve innervation of epineurial arterioles of the sciatic nerve containing calcitonin gene-related peptide: Effect of streptozotocin-induced diabetes. Exp. Diabesity Res. 2004, 5, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Mizukami, H.; Inaba, W.; Baba, M.; Yagihashi, S. The dipeptidyl peptidase IV inhibitor vildagliptin suppresses development of neuropathy in diabetic rodents: Effects on peripheral sensory nerve function, structure and molecular changes. J. Neurochem. 2016, 136. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.P.; Coppey, L.J.; Dake, B.; Yorek, M.A. Treatment of streptozotocin-induced diabetic rats with alogliptin: Effect on vascular and neural complications. Exp. Diabetes Res. 2011, 2011, 810469. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Mori, Y. Anti-atherogenic and anti-inflammatory properties of glucagon-like peptide-1, glucose-dependent insulinotropic polypepide, and dipeptidyl peptidase-4 inhibitors in experimental animals. J. Diabetes Investig. 2016, 7 (Suppl. 1), 80–86. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Nagashima, M.; Nohtomi, K.; Kohashi, K.; Tomoyasu, M.; Sinmura, K.; Nogi, Y.; Katayama, Y.; Sato, K.; Itoh, F.; et al. Preventive effect of dipeptidyl peptidase-4 inhibitor on atherosclerosis is mainly attributable to incretin’s actions in nondiabetic and diabetic apolipoprotein E-null mice. PLoS ONE 2013, 8, e70933. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, M.; Watanabe, T.; Terasaki, M.; Tomoyasu, M.; Nohtomi, K.; Kim-Kaneyama, J.; Miyazaki, A.; Hirano, T. Native incretins prevent the development of atherosclerotic lesions in apolipoprotein E knockout mice. Diabetologia 2011, 54, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Nagashima, M.; Terasaki, M.; Nohtomi, K.; Watanabe, T.; Hirano, T. Glucose-dependent insulinotropic polypeptide prevents the progression of macrophage-driven atherosclerosis in diabetic apolipoprotein E-null mice. PLoS ONE 2012, 7, e35683. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.; Kampfrath, T.; Deiuliis, J.A.; Zhong, J.; Pineda, C.; Ying, Z.; Xu, X.; Lu, B.; Moffatt-Bruce, S.; Durairaj, R.; et al. Long-term dipeptidyl-peptidase 4 inhibition reduces atherosclerosis and inflammation via effects on monocyte recruitment and chemotaxis. Circulation 2011, 124, 2338–2349. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, M.; Mita, T.; Azuma, K.; Ebato, C.; Goto, H.; Nomiyama, T.; Fujitani, Y.; Hirose, T.; Kawamori, R.; Watada, H. Inhibition of monocyte adhesion to endothelial cells and attenuation of atherosclerotic lesion by a glucagon-like peptide-1 receptor agonist, exendin-4. Diabetes 2010, 59, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Yang, S.; Lee, J.I.; Chang, M.J. Cardiovascular effect of incretin-based therapy in patients with type 2 diabetes mellitus: Systematic review and meta-analysis. PLoS ONE 2016, 11, e0153502. [Google Scholar] [CrossRef] [PubMed]
- Mita, T.; Katakami, N.; Yoshii, H.; Onuma, T.; Kaneto, H.; Osonoi, T.; Shiraiwa, T.; Kosugi, K.; Umayahara, Y.; Yamamoto, T.; et al. Alogliptin, a Dipeptidyl Peptidase 4 Inhibitor, Prevents the Progression of Carotid Atherosclerosis in Patients With Type 2 Diabetes: The Study of Preventive Effects of Alogliptin on Diabetic Atherosclerosis (SPEAD-A). Diabetes Care 2016, 39, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Mita, T.; Katakami, N.; Shiraiwa, T.; Yoshii, H.; Onuma, T.; Kuribayashi, N.; Osonoi, T.; Kaneto, H.; Kosugi, K.; Umayahara, Y.; et al. Sitagliptin attenuates the progression of carotid intima-media thickening in insulin-treated patients with type 2 diabetes: The Sitagliptin Preventive Study of Intima-Media Thickness Evaluation (SPIKE): A randomized controlled trial. Diabetes Care 2016, 39, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Jia, H.; Jiang, Y.; Wang, L.; Zhang, Y.; Mu, Y.; Liu, Y. Anti-atherosclerotic effects of the glucagon-like peptide-1 (GLP-1) based therapies in patients with type 2 Diabetes Mellitus: A meta-analysis. Sci. Rep. 2015, 5, 10202. [Google Scholar] [CrossRef] [PubMed]
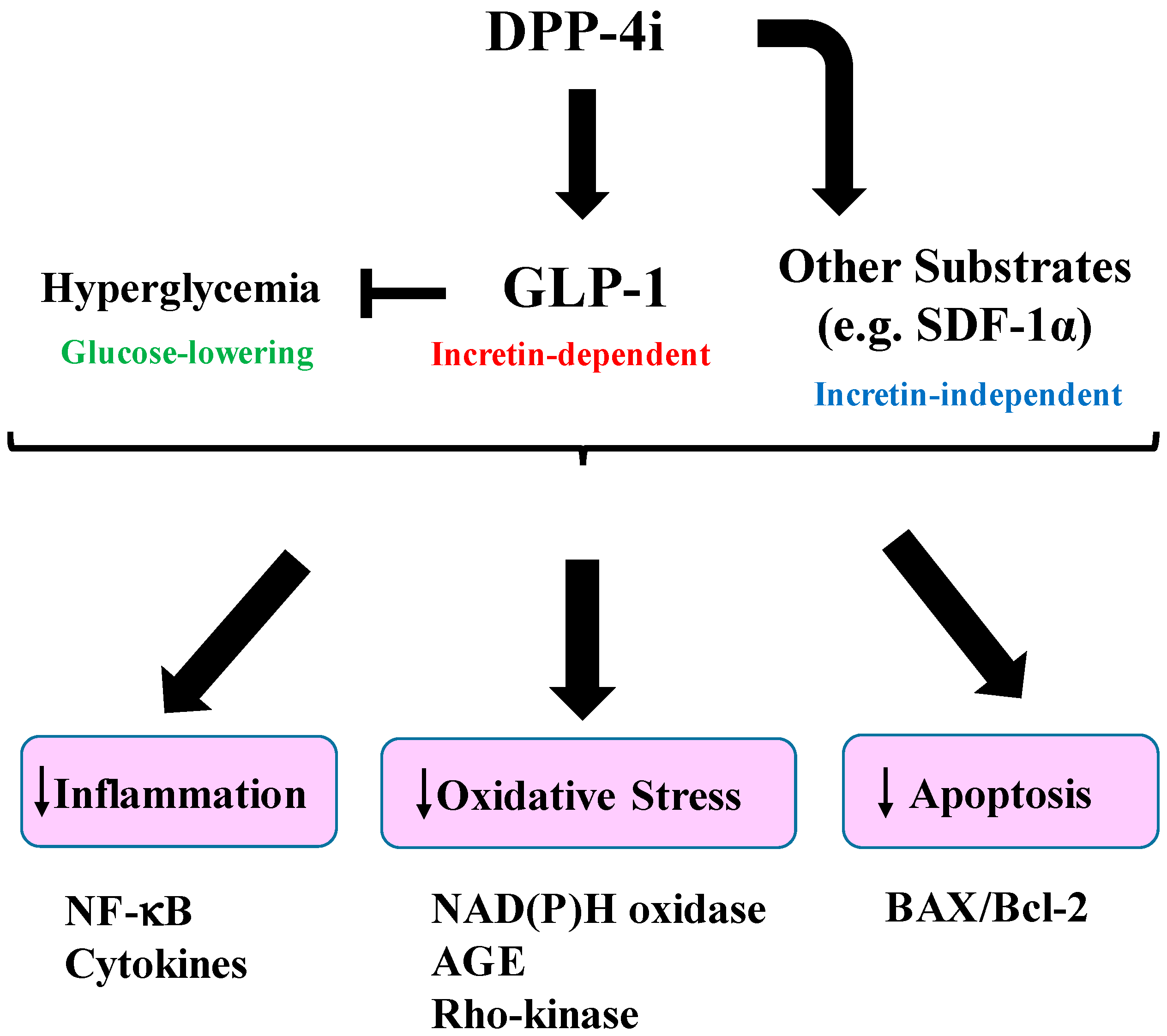


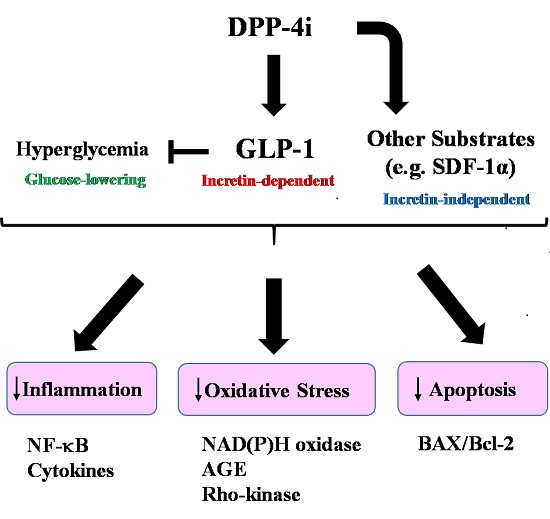{kind=link}
{kind=link}
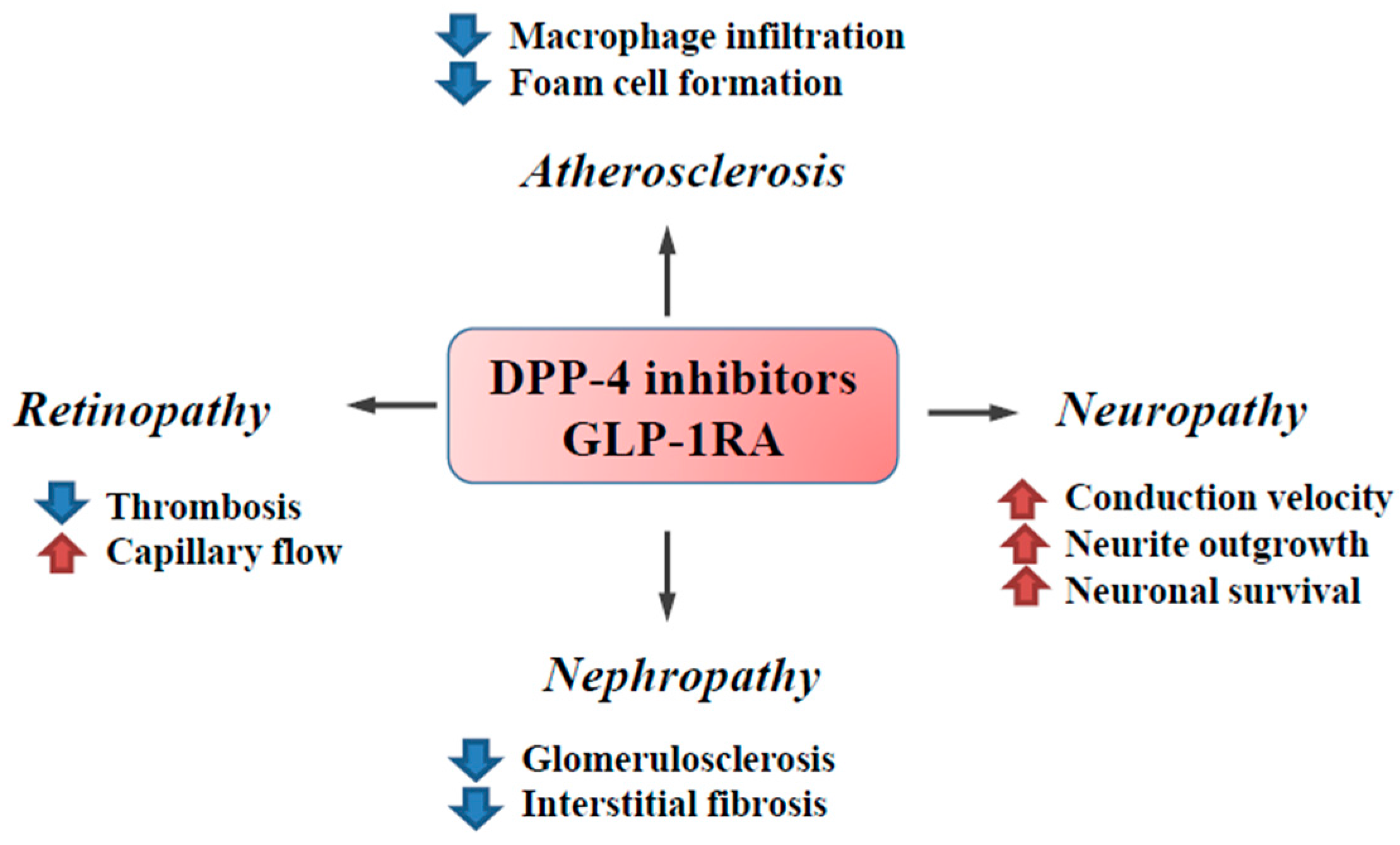{kind=link}
| Complication | Model | Drug/Dose/Duration | Major Effects |
|---|---|---|---|
| Nephropathy | ZDF rats [39] | Sitagliptin,10 mg/kg, 6 weeks | ↓Glomerular lesion |
| ZDF rats [40] | Sitagliptin,10 mg/kg, 6 weeks | ↓Glomerulosclerosis | |
| ↓Tubulointerstitial fibrosis | |||
| STZ-diabetic rats [41] | PKF275-055, 3 mg/kg, 8 weeks | ↓Inflammation | |
| STZ-diabetic mice [42] | Linagliptin, 5 mg/kg, 4 weeks | ↓Kidney fibrosis | |
| STZ-diabetic rats [29] | Exendin-4, 10 mg/kg, 8 weeks | ↓Inflammation | |
| STZ-diabetic rats [43] | Liragltuide, 0.3 mg/kg, 8 weeks | ↓Oxidative stress | |
| Retinopathy | STZ-diabetic rats [44] | Sitagliptin, 5 mg/kg, 2 weeks | ↓Blood-retinal barrier breakdown |
| ↓Inflammation | |||
| ↓Neuronal apoptosis | |||
| ZDF-rats [45] | Sitaglitpin 10 mg/kg, 6 weeks | ↓Inflammation | |
| ↓Retinal cell apoptosis | |||
| OLETF rats [46] | Vildagliptin 3 mg/kg, 10 weeks | ↓Thrombogenic reactions | |
| db/db mice [47] | Liraglutide 400 μg/kg, 15 days | ↓Retinal neurodegenartion | |
| Neuropathy | STZ-diabetic rats [48] | Vildagliptin 0.3 or 10 mg/kg, 32 weeks | ↓Peripheral nerve degeneration |
| STZ-diabetic rats [49] | PKF275-055 3 mg/kg, 4 or 5 weeks | ↑NCV | |
| STZ-diabetic mice [50] | Exendin-4 10 nmol/kg, 4 weeks | ↑Neurite DRG outgrowth | |
| ↑MNCV, SNCV | |||
| STZ-diabetic mice [51] | Exenatide 0.3 pmoles/kg/min, 8 weeks (infusion) | ↑MNCV | |
| Macrovasculopathy | STZ-diabetic apoE-null mice [52] | Alogliptin 15 mg/kg, 24 weeks | ↓Atherosclerotic plaque |
| ZDF rats [53] | Sitaglitpin 10 mg/kg or Linaglitpin 3 mg/kg, 4 weeks | ↑Vascular relaxation, ↓Oxidative stress | |
| STZ-diabetic rats [54] | Sitagliptin 30 mg/kg or | ↓Inflammation | |
| Exenatide 30 μg/kg/12h (infusion), 12 weeks | ↑Endothelial function |
| Complication | Drug | Doses (Duration) | Patients | Endpoint |
|---|---|---|---|---|
| Nephropathy | Sitagliptin [55] | 50 mg/day (6 months) | T2D patients (n = 36) | ↓Albuminuria |
| Sitagliptin [56] | 50 mg/day (6 months) | T2D patients (n = 85) | ↓Albuminuria | |
| Saxagliptin [57] | 2.5 or 5 mg/day (2 years) | T2D patients (n = 16,492) | ↓Albuminuria | |
| Linagliptin [58] | 5 mg/day (6 months) | T2D patients (n = 217) | ↓Albuminuria | |
| Alogliptin [59] | 25 mg/day (4 weeks) (vs. Sitagliptin 50 mg/day) (cross over) | T2D patients (n = 12) | ↓Albuminuria | |
| Liraglutide [60] | 0.6-1.8 mg/day (1 year) | T2D patients (n = 84) | ↓Albuminuria | |
| Exenatide [61] | 10 μg twice daily (16 weeks) (5 μg twice daily (first 4 weeks) | T2D patients (n = 31) | ↓Albuminuria | |
| Liraglutide [62] | 1.8 mg/day (3.8 years) | T2D patients (n = 9340) | ↓Composite outcome of renal and retinal microvascular events | |
| Retinopathy | Saxagliptin [63] | 5 mg/day (6 weeks) | T2D patients (n = 50) | Normalization of retinal capillary flow |
| Exenatide [64] | N/A (300 days) | T2D patients (n = 165) | Transient worsening of diabetic retinopathy (DR) | |
| Exenatide [65] | N/A (430 days) | T2D patients (n = 39) | Improvement of DR | |
| Neuropathy | Exenatide [66] | 10 μg twice daily (18 months) (5 μg twice daily (first 4 weeks)) | T2D patients (n = 46) | No changes in confirmed clinical neuropathy, cardiovascular autonomic neuropathy |
| Trial | Drug/Doses | Patients | Primary Composite Outcome | Result (Risk of Cardiovascular Events) |
|---|---|---|---|---|
| SAVOR-TIMI53 [57] (2.1 years) | Saxagliptin 2.5 mg or 5 mg/day (on the basis of estimated glomerular filtration rate (eGFR) at baseline) | T2D patients who had a history of, or were at risk for, cardiovascular events (n = 16,492) | Cardiovascular death, myocardial infarction, or ischemic stroke |  (no change) |
| EXAMINE [67] (1.5 years) | Alogliptin 6.25 mg or 12.5 mg or 25 mg (same as above) | T2D patients with either an acute myocardial infarction or unstable angina requiring hospitalization within the previous 15 to 90 days (n = 5380) | Cardiovascular death, nonfatal myocardial infarction, or nonfatal stroke | |
| TECOS [68] (3.0 years) | Sitagliptin 50 mg or 100 mg/day (same as above) | T2D patients who had a history of major coronary artery disease, ischemic cerebrovascular disease, or atherosclerotic peripheral arterial disease (n = 14,671) | Cardiovascular death, nonfatal myocardial infarction, nonfatal stroke, or hospitalization for unstable angina | |
| ELIXA [69] (2.1 years) | Lixisenatide 20 μg/day | T2D patients who had had a myocardial infarction or who had been hospitalized for unstable angina within the previous 180 days (n = 6068) | Cardiovascular death, myocardial infarction, stroke, or hospitalization for unstable angina | |
| LEADER [62] (3.8 years) | Liraglutide 1.8 mg/day | T2D patients ≥50 years of age with at least one cardiovascular coexisting condition or ≥60 years of age with at least one cardiovascular risk factor (n = 9340) | Cardiovascular death, nonfatal myocardial infarction, or nonfatal stroke |  (decrease) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawanami, D.; Matoba, K.; Sango, K.; Utsunomiya, K. Incretin-Based Therapies for Diabetic Complications: Basic Mechanisms and Clinical Evidence. Int. J. Mol. Sci. 2016, 17, 1223. https://doi.org/10.3390/ijms17081223
Kawanami D, Matoba K, Sango K, Utsunomiya K. Incretin-Based Therapies for Diabetic Complications: Basic Mechanisms and Clinical Evidence. International Journal of Molecular Sciences. 2016; 17(8):1223. https://doi.org/10.3390/ijms17081223
Chicago/Turabian StyleKawanami, Daiji, Keiichiro Matoba, Kazunori Sango, and Kazunori Utsunomiya. 2016. "Incretin-Based Therapies for Diabetic Complications: Basic Mechanisms and Clinical Evidence" International Journal of Molecular Sciences 17, no. 8: 1223. https://doi.org/10.3390/ijms17081223