
Immunomodulation Induced by Stem Cell Mobilization and Harvesting in Healthy Donors: Increased Systemic Osteopontin Levels after Treatment with Granulocyte Colony-Stimulating Factor

Abstract
:1. Introduction
2. Results
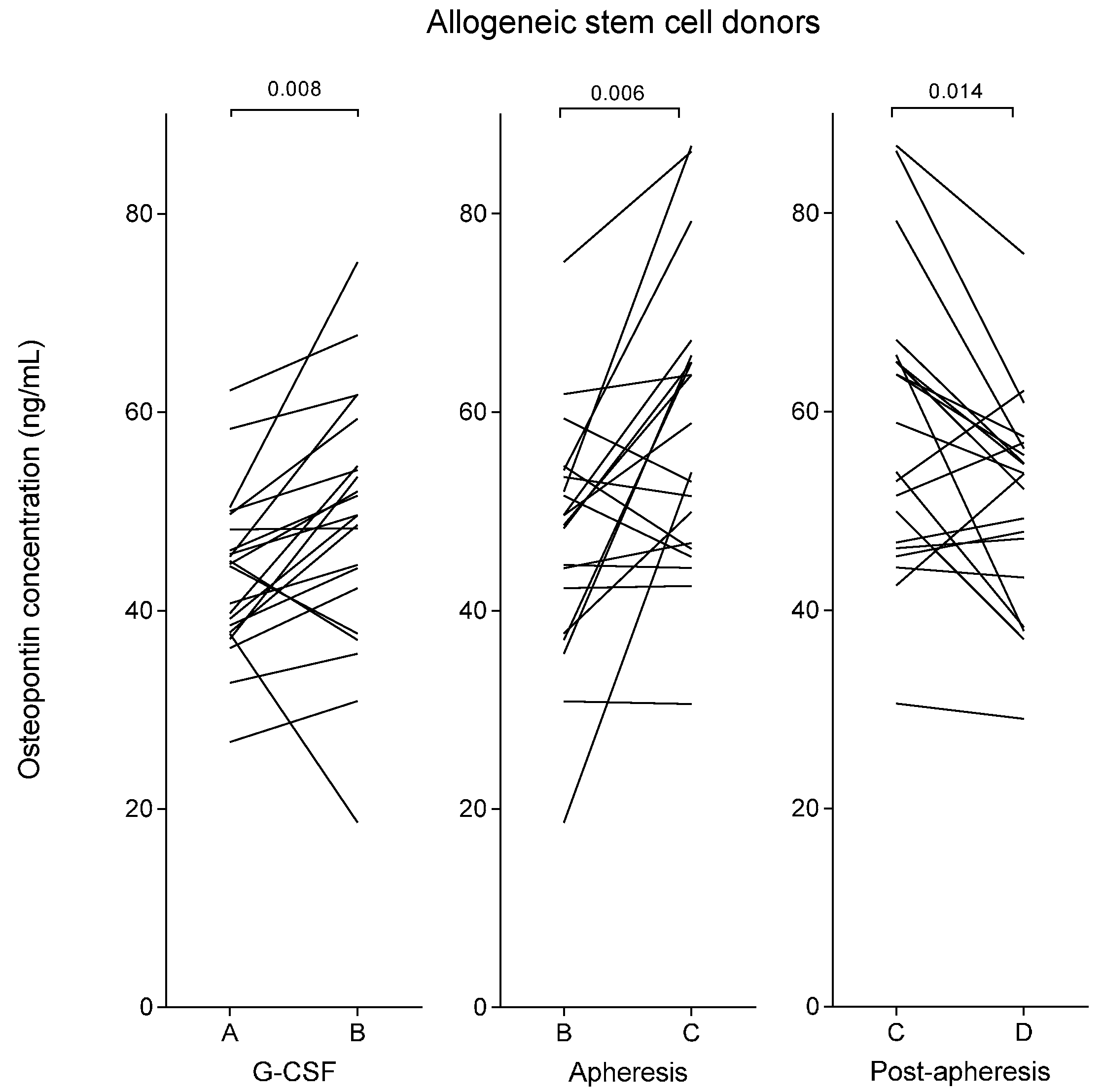
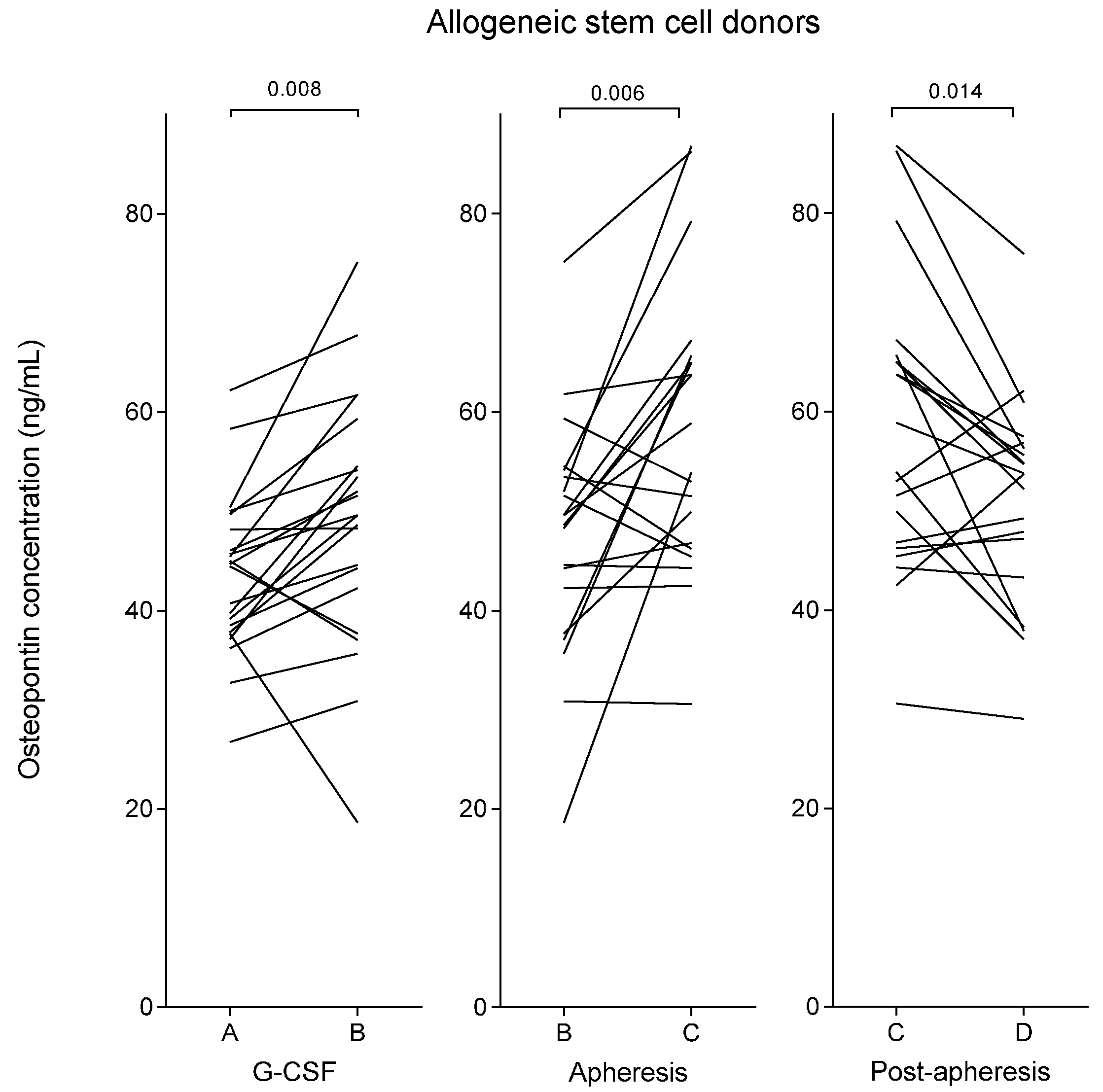
2.1. Plasma Osteopontin Levels of Healthy Stem Cell Donors Increase during Granulocyte Colony-Stimulating Factor (G-CSF) Treatment and Reach a Maximal Level Immediately Following Stem Cell Harvesting by Leukapheresis
2.2. Plasma Osteopontin Levels Show an Inverse Correlation with Peripheral Blood Neutrophil Levels during G-CSF Therapy but No Association with Peripheral Blood Levels or Yields of CD34+ Cells
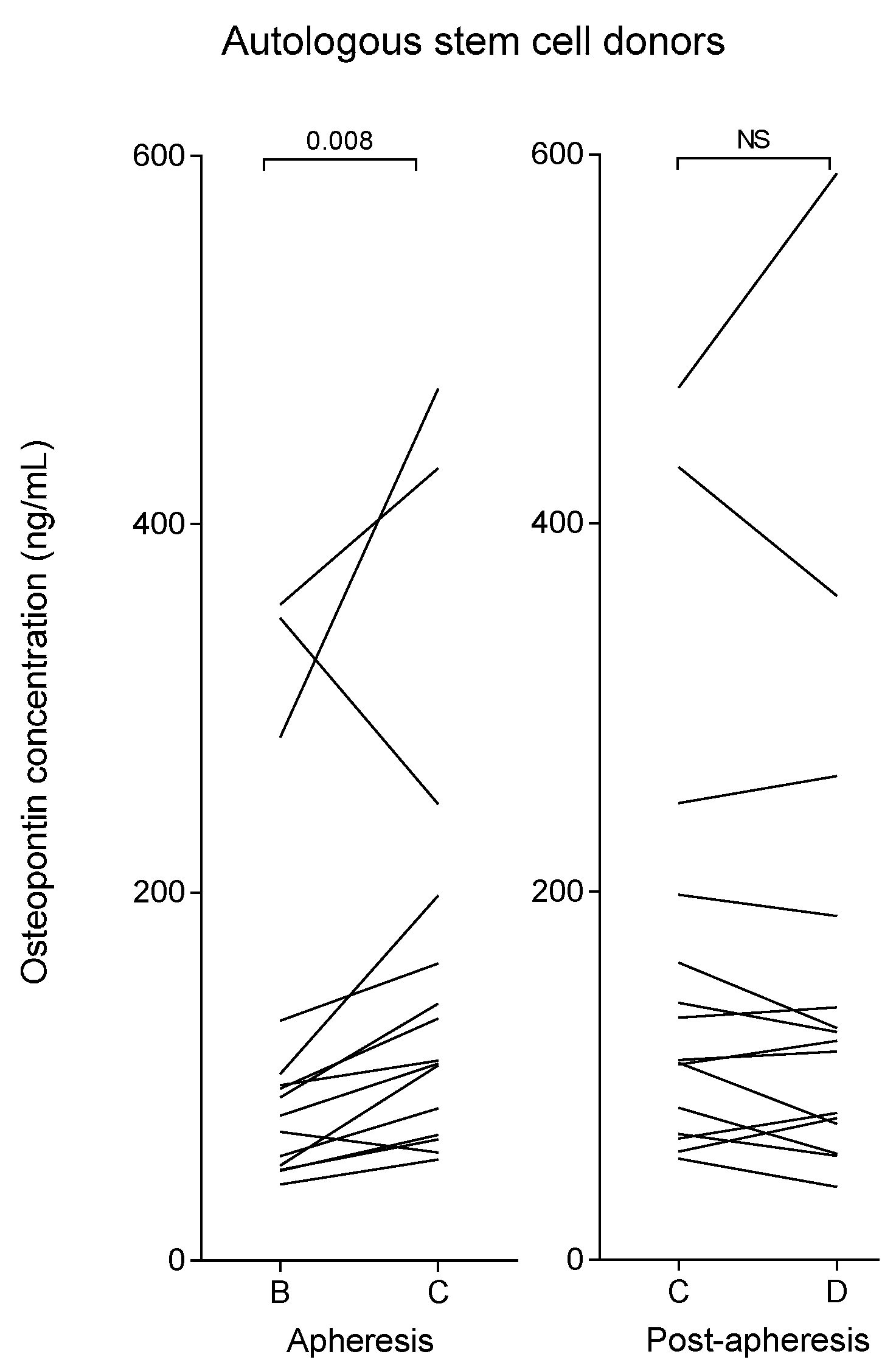
2.3. Myeloma Patients (Autologous Stem Cell Donors) Show Increased Plasma Osteopontin Levels after G-CSF Therapy Compared with Healthy Allogeneic Stem Cell Donors
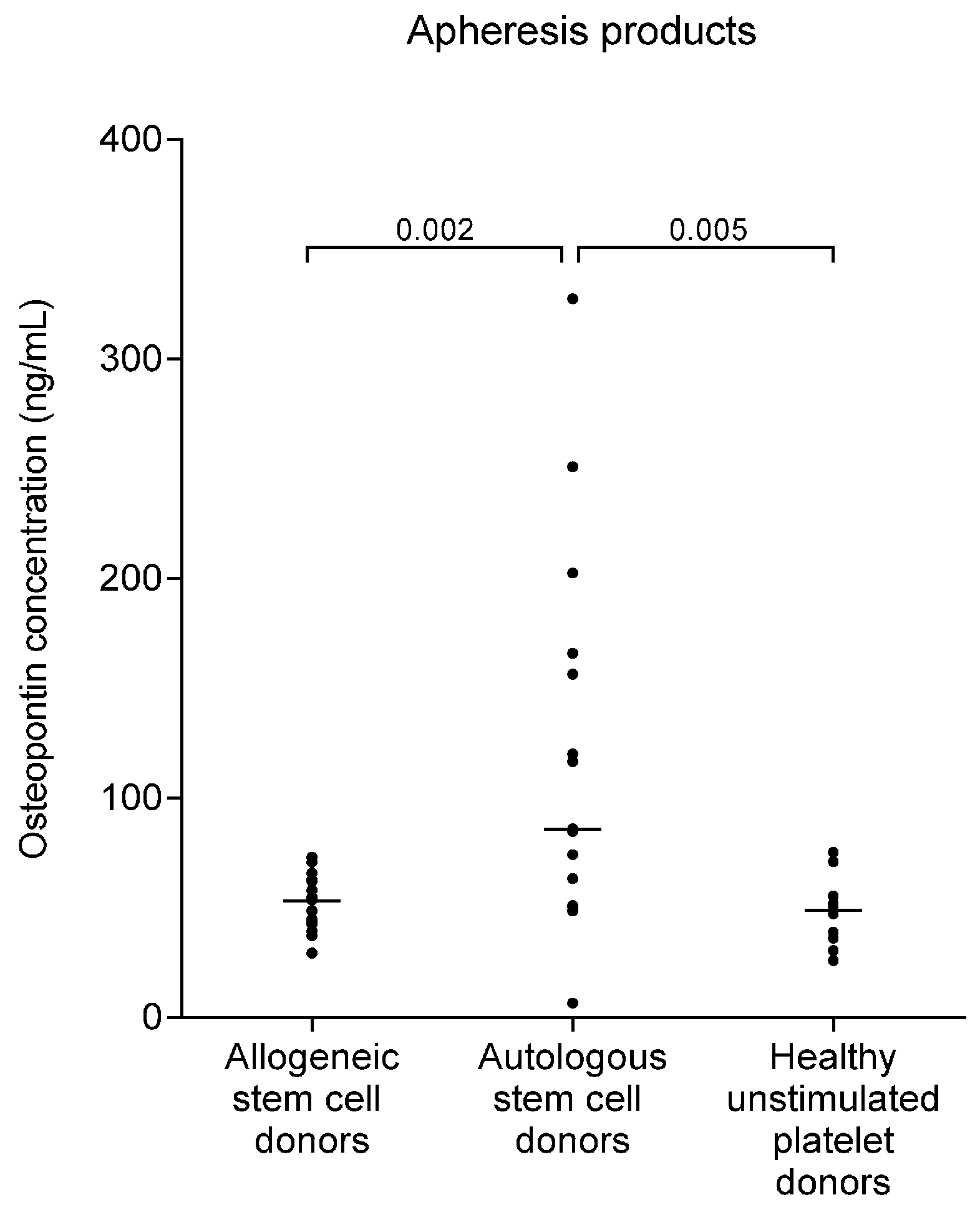
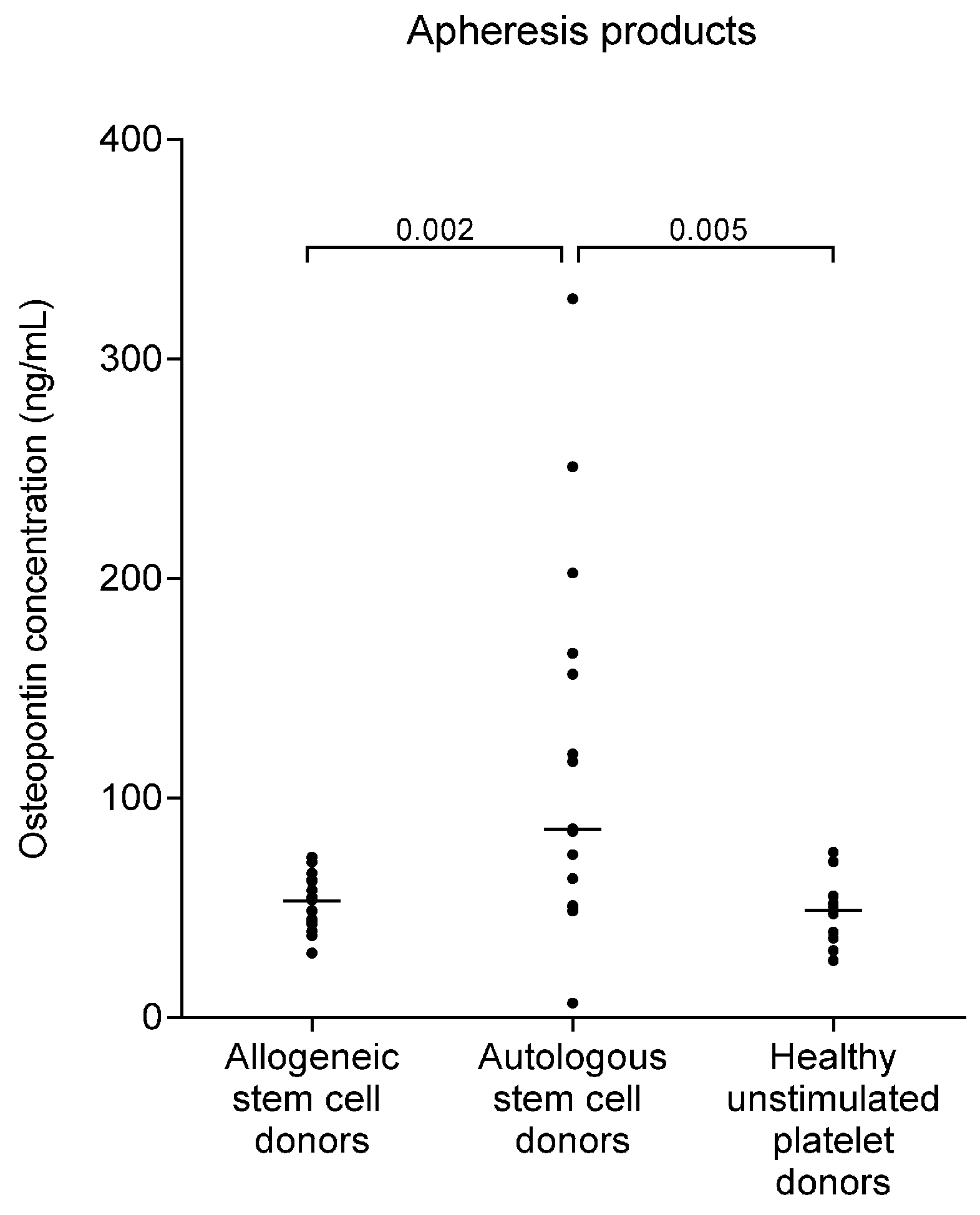
2.4. Osteopontin Levels Are Higher in Autografts from Myeloma Patients than in Allografts from Healthy Stem Cell Donors
2.5. Pretransplant Osteopontin Levels of Allotransplant Recipients Are Increased and the High Levels Are Not Altered Following the Infusion of Osteopontin-Containing Stem Cell Grafts
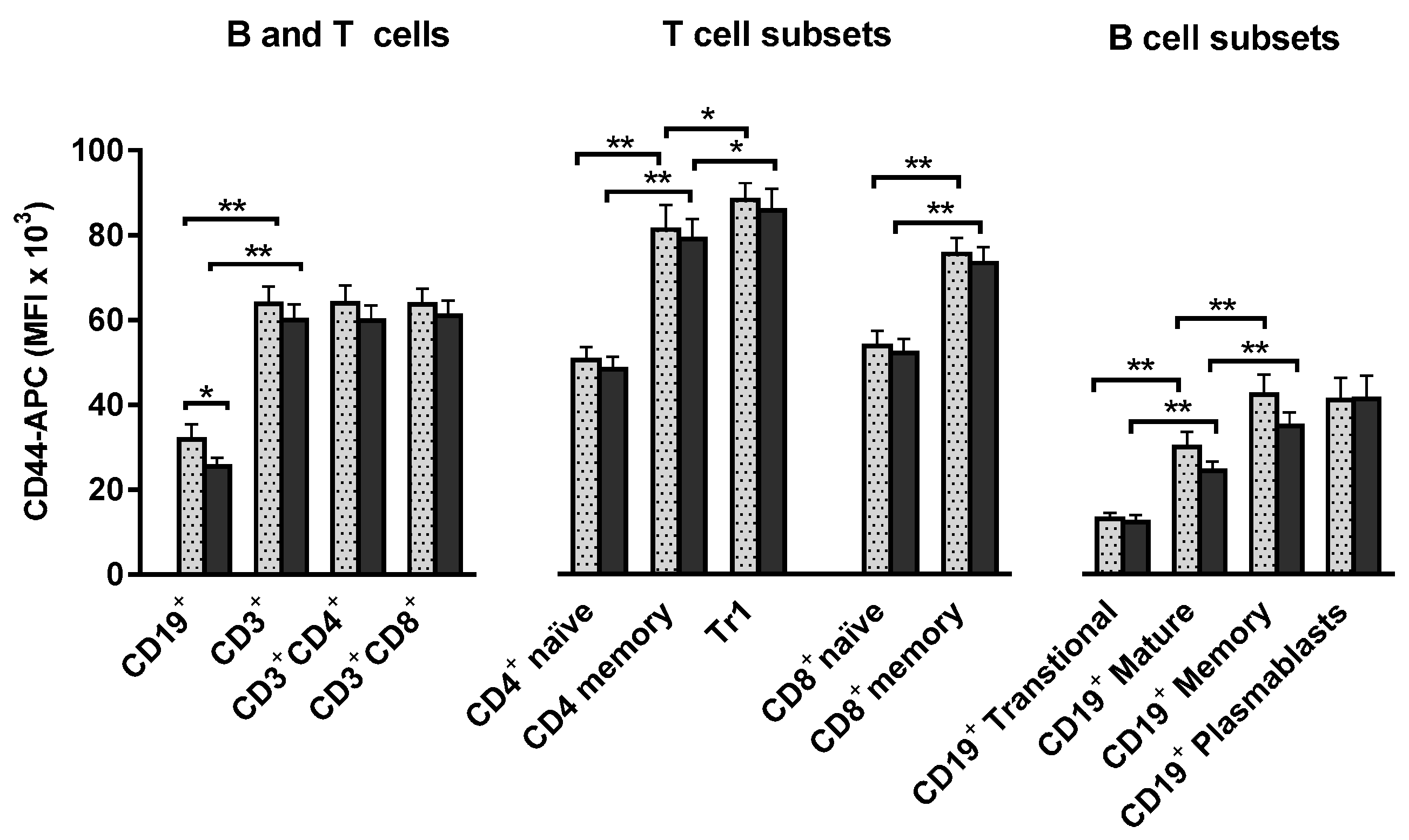
2.6. T and B Lymphocytes Show High Expression of the CD44 Osteopontin Receptor and these High Levels Are Maintained during Stem Cell Mobilization and Harvesting
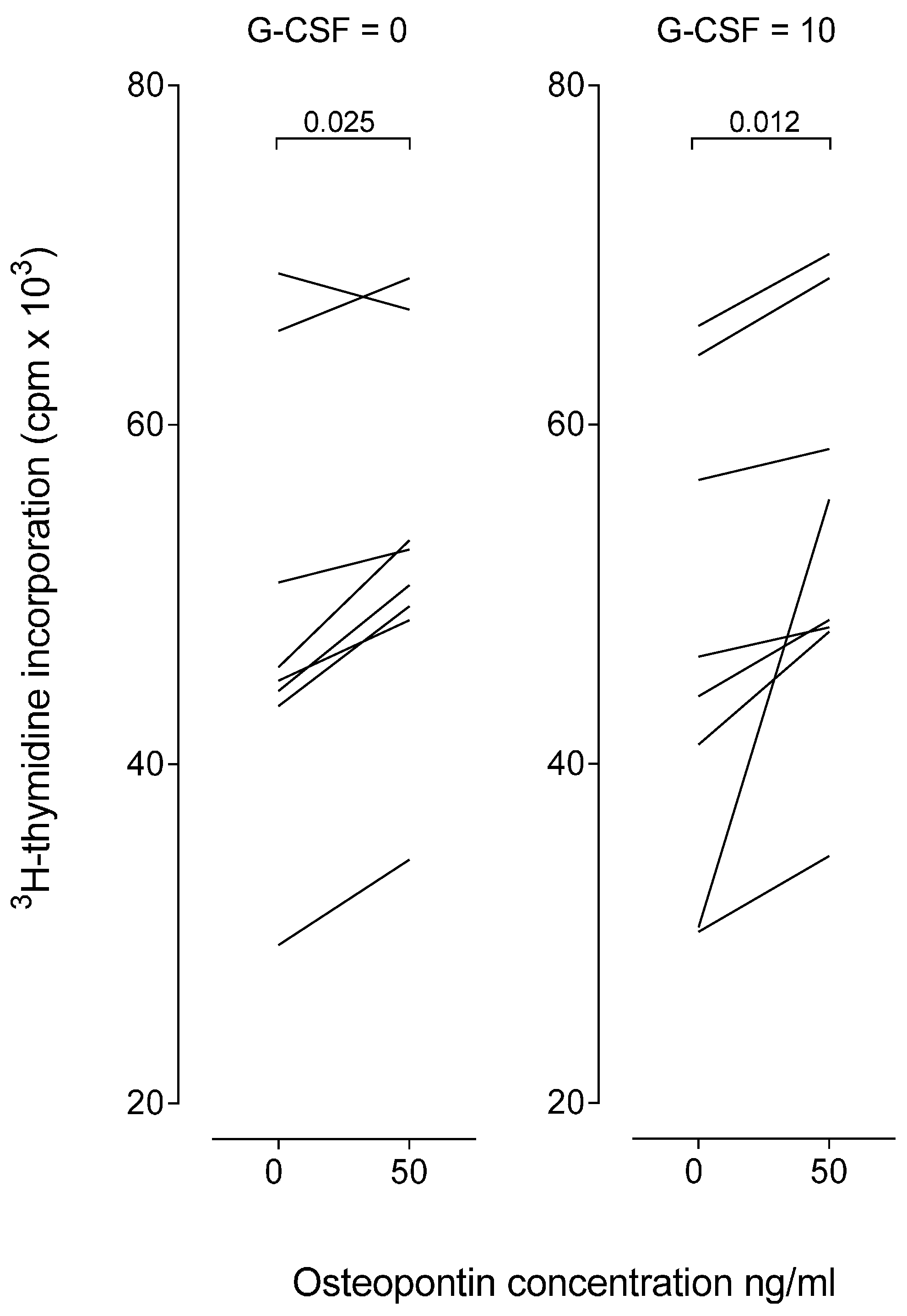
2.7. Osteopontin Causes a Minor Increase of in Vitro Proliferative T Cell Responses
3. Discussion
4. Materials and Methods
4.1. Stem Cell Donors and Allotransplant Recipients
4.2. Stem Cell Mobilization in Healthy Donors and Myeloma Patients
4.3. Apheresis Procedures
4.4. Allogeneic Stem Cell Transplantation
4.5. Preparation of Plasma and Peripheral Blood Mononuclear Cells (PBMC)
4.5.1. Blood Sampling
4.5.2. Preparation of Plasma Samples
4.5.3. Preparation of PBMC Samples
4.6. Analysis of Plasma Osteopontin and G-CSF Concentrations
4.7. Flow Cytometry Analyses
4.8. Analysis of T-Cell Proliferation by 3H-Thymidine Incorporation
4.9. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| G-CSF | Granulocyte colony-stimulating factor |
| CD | Cluster of differentiation |
| HLA | Human leukocyte antigen |
| GVHD | Graft versus host disease |
| OPN | Osteopontin |
| HSC | Hematopoietic stem cell |
| HSCT | Hematopoietic stem cell transplantation |
| MFI | Mean fluorescence intensity |
| PBMC | Peripheral blood mononuclear cells |
| LAG-3 | Lymphocyte activation gene 3 |
| Tr1 cells | T regulatory type 1 cells |
| 3H | Tritiated hydrogen |
| MNC | Mononuclear cells |
| AML | Acute myeloid leukemia |
| B-ALL | B cell lymphoblastic leukemia |
| MDS | Myelodysplastic syndrome |
| ACD-A | Acid-citrate-dextrose solution A |
| FBS | Fetal Bovine Serum |
| DMSO | Dimethyl sulfoxide |
| Near-IR | Near-infrared |
| PBS | Phosphate-buffered saline |
| BSA | Bovine Serum Albumin |
References
- Wang, K.X.; Denhardt, D.T. Osteopontin: Role in immune regulation and stress responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Zohar, R.; Zhu, B.; Liu, P.; Sodek, J.; McCulloch, C.A. Increased cell death in osteopontin-deficient cardiac Fibroblasts Occurs by a Caspase-3-Independent Pathway. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1730–H1739. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Zhu, B.; Rittling, S.R.; Denhardt, D.T.; Goldberg, H.A.; McCulloch, C.A.; Sodek, J. Colocalization of intracellular osteopontin with CD44 is associated with migration, cell fusion, and resorption in osteoclasts. J. Bone Miner. Res. 2002, 17, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.L.; Lu, L.; Bu, J.; Werneck, M.B.; Kobayashi, K.S.; Glimcher, L.H.; Cantor, H. Osteopontin expression is essential for interferon-α production by plasmacytoid dendritic cells. Nat. Immunol. 2006, 7, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.H.; Juliana, M.M.; Hicks, P.H.; Feng, G.; Elmets, C.; Liaw, L.; Chang, P.L. Papilloma development is delayed in osteopontin-null mice: Implicating an antiapoptosis role for osteopontin. Cancer Res. 2006, 66, 7119–7127. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Shinohara, M.L. Intracellular osteopontin (iOPN) and immunity. Immunol. Res. 2011, 49, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K.; Johnston, H.M.; Whitty, G.A.; Williams, B.; Webb, R.J.; Denhardt, D.T.; Bertoncello, I.; Bendall, L.J.; Simmons, P.J.; Haylock, D.N. Osteopontin, a key component of the hematopoietic stem cell niche and regulator of primitive hematopoietic progenitor cells. Blood 2005, 106, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhai, Q.; Zou, D.; Meng, H.; Xie, Z.; Li, C.; Wang, Y.; Qi, J.; Cheng, T.; Qiu, L. A pivotal role of bone remodeling in granulocyte colony stimulating factor induced hematopoietic stem/progenitor cells mobilization. J. Cell. Physiol. 2013, 228, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Sodek, J.; Batista Da Silva, A.P.; Zohar, R. Osteopontin and mucosal protection. J. Dent. Res. 2006, 85, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ambrosi, J.; Catalan, V.; Ramirez, B.; Rodriguez, A.; Colina, I.; Silva, C.; Rotellar, F.; Mugueta, C.; Gil, M.J.; Cienfuegos, J.A.; et al. Plasma osteopontin levels and expression in adipose tissue are increased in obesity. J. Clin. Endocrinol. Metab. 2007, 92, 3719–3727. [Google Scholar] [CrossRef] [PubMed]
- Rangaswami, H.; Bulbule, A.; Kundu, G.C. Osteopontin: Role in cell signaling and cancer progression. Trends Cell Biol. 2006, 16, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K.; Rittling, S. Role of osteopontin in regulating hepatic inflammatory responses and toxic liver injury. Expert Opin. Drug Metab. Toxicol. 2007, 3, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.H.; Messenger, M.P.; Gregory, W.M.; Wind, T.C.; Vasudev, N.S.; Cartledge, J.; Thompson, D.; Selby, P.J.; Banks, R.E. Prognostic utility of pre-operative circulating osteopontin, carbonic anhydrase IX and CRP in renal cell carcinoma. Br. J. Cancer 2012, 107, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin—A master regulator of epithelial-mesenchymal transition. J. Clin. Med. 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Rittling, S.R.; Singh, R. Osteopontin in immune-mediated diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Iyonaga, K.; Ichiyasu, H.; Nagano, J.; Suga, M.; Sasaki, Y. Differentiation, maturation, and survival of dendritic cells by osteopontin regulation. Clin. Diagn. Lab. Immunol. 2005, 12, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Tumang, J.R.; Rothstein, T.L. B cell receptor crosstalk: B cells express osteopontin through the combined action of the alternate and classical BCR signaling pathways. Mol. Immunol. 2009, 46, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S.; Hashimoto, S.; Yamashita, T.; Toyoda, N.; Satoh, T.; Suzuki, T.; Matsushima, K. Comprehensive gene expression profile of human activated Th1- and Th2-polarized cells. Int. Immunol. 2001, 13, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.F.; Cantor, H. Differential roles of osteopontin/Eta-1 in early and late lpr disease. Clin. Exp. Immunol. 2001, 126, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, K.; Hoffmann, U.; Witte, E.; Loddenkemper, C.; Sina, C.; Schreiber, S.; Hayford, C.; Holzlohner, P.; Wolk, K.; Tchatchou, E.; et al. Osteopontin as two-sided mediator of intestinal inflammation. J. Cell. Mol. Med. 2009, 13, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nakai, T.; Tamura, N.; Okamoto, S.; Matsuoka, K.; Sakuraba, A.; Fukushima, T.; Uede, T.; Hibi, T. Osteopontin/Eta-1 upregulated in Crohn’s disease regulates the Th1 immune response. Gut 2005, 54, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Zafrir, M.; Yesilkanal, A.E.; Price, T.T.; Hyjek, E.M.; Sipkins, D.A. Adhesion to osteopontin in the bone marrow niche regulates lymphoblastic leukemia cell dormancy. Blood 2013, 121, 4821–4831. [Google Scholar] [CrossRef] [PubMed]
- Liersch, R.; Gerss, J.; Schliemann, C.; Bayer, M.; Schwoppe, C.; Biermann, C.; Appelmann, I.; Kessler, T.; Lowenberg, B.; Buchner, T.; et al. Osteopontin is a prognostic factor for survival of acute myeloid leukemia patients. Blood 2012, 119, 5215–5220. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Lit, L.C.; Tam, L.S.; Li, E.K.; Lam, C.W. Elevation of plasma osteopontin concentration is correlated with disease activity in patients with systemic lupus erythematosus. Rheumatology 2005, 44, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Ostheimer, C.; Bache, M.; Guttler, A.; Kotzsch, M.; Vordermark, D. A pilot study on potential plasma hypoxia markers in the radiotherapy of non-small cell lung cancer. Osteopontin, carbonic anhydrase IX and vascular endothelial growth factor. Strahlenther. Onkol. 2014, 190, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Korbling, M.; Freireich, E.J. Twenty-five years of peripheral blood stem cell transplantation. Blood 2011, 117, 6411–6416. [Google Scholar] [CrossRef] [PubMed]
- Devine, H.; Tierney, D.K.; Schmit-Pokorny, K.; McDermott, K. Mobilization of hematopoietic stem cells for use in autologous transplantation. Clin. J. Oncol. Nurs. 2010, 14, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Kindwall-Keller, T. Peripheral stem cell collection: From leukocyte growth factor to removal of catheter. J. Clin. Apheresis 2014, 29, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Flommersfeld, S.; Bakchoul, T.; Bein, G.; Wachtel, A.; Loechelt, C.; Sachs, U.J. A single center comparison between three different apheresis systems for autologous and allogeneic stem cell collections. Transfus. Apheresis Sci. 2013, 49, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Brauninger, S.; Bialleck, H.; Thorausch, K.; Felt, T.; Seifried, E.; Bonig, H. Allogeneic donor peripheral blood “stem cell” apheresis: Prospective comparison of two apheresis systems. Transfusion 2012, 52, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, P.; Brauninger, S.; Bialleck, H.; Thorausch, K.; Smith, R.; Schrezenmeier, H.; Seifried, E.; Wiesneth, M.; Bonig, H. Automatic interface-controlled apheresis collection of stem/progenitor cells: Results from an autologous donor validation trial of a novel stem cell apheresis device. Transfusion 2011, 51, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Gennery, A.R.; Slatter, M.A.; Grandin, L.; Taupin, P.; Cant, A.J.; Veys, P.; Amrolia, P.J.; Gaspar, H.B.; Davies, E.G.; Friedrich, W.; et al. Transplantation of hematopoietic stem cells and long-term survival for primary immunodeficiencies in Europe: Entering a new century, do we do better? J. Allergy Clin. Immunol. 2010, 126, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Naumann-Winter, F.; Greb, A.; Borchmann, P.; Bohlius, J.; Engert, A.; Schnell, R. First-line tandem high-dose chemotherapy and autologous stem cell transplantation versus single high-dose chemotherapy and autologous stem cell transplantation in multiple myeloma, a systematic review of controlled studies. Cochrane Database Syst. Rev. 2012, 10. [Google Scholar] [CrossRef] [Green Version]
- Jantunen, E.; Sureda, A. The evolving role of stem cell transplants in lymphomas. Biol. Blood Marrow Transplant. J. Am. Soc. Blood Marrow Transplant. 2012, 18, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Passweg, J.R.; Baldomero, H.; Bader, P.; Bonini, C.; Cesaro, S.; Dreger, P.; Duarte, R.F.; Dufour, C.; Kuball, J.; Farge-Bancel, D.; et al. Hematopoietic stem cell transplantation in Europe 2014: More than 40,000 transplants annually. Bone Marrow Transplant. 2016, 51, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Bader, P.; Cesaro, S.; Dreger, P.; Duarte, R.F.; Dufour, C.; Falkenburg, J.H.; Farge-Bancel, D.; Gennery, A.; Kroger, N.; et al. Indications for allo- and auto-sct for haematological diseases, solid tumours and immune disorders: Current practice in Europe, 2015. Bone Marrow Transplant. 2015, 50, 1037–1056. [Google Scholar] [CrossRef] [PubMed]
- Farge, D.; Labopin, M.; Tyndall, A.; Fassas, A.; Mancardi, G.L.; van Laar, J.; Ouyang, J.; Kozak, T.; Moore, J.; Kotter, I.; et al. Autologous hematopoietic stem cell transplantation for autoimmune diseases: An observational study on 12 years’ experience from the European group for blood and marrow transplantation working party on autoimmune diseases. Haematologica 2010, 95, 284–292. [Google Scholar] [CrossRef] [PubMed]
- D’Addio, F.; Vasquez, A.V.; Nasr, M.B.; Franek, E.; Zhu, D.; Li, L.; Ning, G.; Snarski, E.; Fiorina, P. Autologous nonmyeloablative hematopoietic stem cell transplantation in new-onset type 1 diabetes: A multicenter analysis. Diabetes 2014, 63, 3041–3046. [Google Scholar] [CrossRef] [PubMed]
- Al-toma, A.; Nijeboer, P.; Bouma, G.; Visser, O.; Mulder, C.J. Hematopoietic stem cell transplantation for non-malignant gastrointestinal diseases. World J. Gastroenterol. 2014, 20, 17368–17375. [Google Scholar] [CrossRef] [PubMed]
- Pavo, N.; Charwat, S.; Nyolczas, N.; Jakab, A.; Murlasits, Z.; Bergler-Klein, J.; Nikfardjam, M.; Benedek, I.; Benedek, T.; Pavo, I.J.; et al. Cell therapy for human ischemic heart diseases: Critical review and summary of the clinical experiences. J. Mol. Cell. Cardiol. 2014, 75, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Burt, R.K.; Testori, A.; Oyama, Y.; Rodriguez, H.E.; Yaung, K.; Villa, M.; Bucha, J.M.; Milanetti, F.; Sheehan, J.; Rajamannan, N.; et al. Autologous peripheral blood CD133+ cell implantation for limb salvage in patients with critical limb ischemia. Bone Marrow Transplant. 2010, 45, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.K.; Driscoll, T.; Boczkowski, D.; Schmittling, R.; Reynolds, R.; Johnson, L.A.; Grant, G.; Fuchs, H.; Bigner, D.D.; Sampson, J.H.; et al. Ex vivo generation of dendritic cells from cryopreserved, post-induction chemotherapy, mobilized leukapheresis from pediatric patients with medulloblastoma. J. Neuro-Oncol. 2015, 125, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kassis, I.; Zangi, L.; Rivkin, R.; Levdansky, L.; Samuel, S.; Marx, G.; Gorodetsky, R. Isolation of mesenchymal stem cells from G-CSF-mobilized human peripheral blood using fibrin microbeads. Bone Marrow Transplant. 2006, 37, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.L.; Zhou, C.Y.; Yu, J.K. A new source of mesenchymal stem cells for articular cartilage repair: MSCs derived from mobilized peripheral blood share similar biological characteristics in vitro and chondrogenesisin vivo as MSCs from bone marrow in a rabbit model. Am. J. Sports Med. 2014, 42, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Paczesny, S.; Hanauer, D.; Sun, Y.; Reddy, P. New perspectives on the biology of acute GVHD. Bone Marrow Transplant. 2010, 45, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, Y.; Wang, H.; Jin, M.; He, S.; Shi, Y.; Guo, Y.; Zhang, Y. Blockade of osteopontin reduces alloreactive CD8+ T cell-mediated graft-versus-host disease. Blood 2011, 117, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-ligand interaction between CD44 and osteopontin (Eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Gagliani, N.; Magnani, C.F.; Huber, S.; Gianolini, M.E.; Pala, M.; Licona-Limon, P.; Guo, B.; Herbert, D.R.; Bulfone, A.; Trentini, F.; et al. Coexpression of CD49b and LAG-3 identifies human and mouse T regulatory type 1 cells. Nat. Med. 2013, 19, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Carsetti, R.; Rosado, M.M.; Wardmann, H. Peripheral development of B cells in mouse and man. Immunol. Rev. 2004, 197, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Agematsu, K.; Nagumo, H.; Yang, F.C.; Nakazawa, T.; Fukushima, K.; Ito, S.; Sugita, K.; Mori, T.; Kobata, T.; Morimoto, C.; et al. B cell subpopulations separated by CD27 and crucial collaboration of CD27+ B cells and helper T cells in immunoglobulin production. Eur. J. Immunol. 1997, 27, 2073–2079. [Google Scholar] [CrossRef] [PubMed]
- Sennels, H.P.; Jacobsen, S.; Jensen, T.; Hansen, M.S.; Ostergaard, M.; Nielsen, H.J.; Sorensen, S. Biological variation and reference intervals for circulating osteopontin, osteoprotegerin, total soluble receptor activator of nuclear factor κB ligand and high-sensitivity C-reactive protein. Scand. J. Clin. Lab. Investig. 2007, 67, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.; Wan, T.M.; Lam, C.S.; Chow, A.K.; Wong, S.K.; Man, J.H.; Li, H.S.; Cheng, N.S.; Pak, R.C.; Cheung, A.H.; et al. Post-operative plasma osteopontin predicts distant metastasis in human colorectal cancer. PLoS ONE 2015, 10, e0126219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurata, M.; Okura, T.; Irita, J.; Enomoto, D.; Nagao, T.; Jotoku, M.; Miyoshi, K.; Desilva, V.R.; Higaki, J. Angiotensin II receptor blockade with valsartan decreases plasma osteopontin levels in patients with essential hypertension. J. Hum. Hypertens. 2011, 25, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Azeez, H.A.; Al-Zaky, M. Plasma osteopontin as a predictor of coronary artery disease: Association with echocardiographic characteristics of atherosclerosis. J. Clin. Lab. Anal. 2010, 24, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Sfiridaki, A.; Miyakis, S.; Pappa, C.; Tsirakis, G.; Alegakis, A.; Kotsis, V.; Stathopoulos, E.; Alexandrakis, M. Circulating osteopontin: A dual marker of bone destruction and angiogenesis in patients with multiple myeloma. J. Hematol. Oncol. 2011, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Patel, C.G.; Yee, A.J.; Scullen, T.A.; Nemani, N.; Santo, L.; Richardson, P.G.; Laubach, J.P.; Ghobrial, I.M.; Schlossman, R.L.; Munshi, N.C.; et al. Biomarkers of bone remodeling in multiple myeloma patients to tailor bisphosphonate therapy. Clin. Cancer Res. 2014, 20, 3955–3961. [Google Scholar] [CrossRef] [PubMed]
- Szmigielska-Kaplon, A.; Krawczynska, A.; Czemerska, M.; Pluta, A.; Cebula-Obrzut, B.; Szmigielska, K.; Stepka, K.; Smolewski, P.; Robak, T.; Wierzbowska, A. Angiopoietins in haematopoietic stem cell mobilisation in patients with haematological malignancies. Blood Transfus. 2015, 13, 102–108. [Google Scholar] [PubMed]
- Hagberg, I.A.; Akkok, C.A.; Lyberg, T.; Kjeldsen-Kragh, J. Apheresis-induced platelet activation: Comparison of three types of cell separators. Transfusion 2000, 40, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Macher, S.; Sipurzynski-Budrass, S.; Rosskopf, K.; Rohde, E.; Griesbacher, A.; Groselj-Strele, A.; Lanzer, G.; Schallmoser, K. Function and activation state of platelets in vitro depend on apheresis modality. Vox Sang. 2010, 99, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; da Silva, A.P.; Bansal, A.K.; Bansal, M.; Sun, C.; Lee, H.; Glogauer, M.; Sodek, J.; Zohar, R. Role of osteopontin in neutrophil function. Immunology 2007, 122, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Szmigielska-Kaplon, A.; Szemraj, J.; Hamara, K.; Robak, M.; Wolska, A.; Pluta, A.; Czemerska, M.; Krawczynska, A.; Jamroziak, K.; Szmigielska, K.; et al. Polymorphism of CD44 influences the efficacy of CD34+ cells mobilization in patients with hematological malignancies. Biol. Blood Marrow Transplantat. J. Am. Soc. Blood Marrow Transplant. 2014, 20, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Melve, G.K.; Ersvssr, E.; Kittang, A.O.; Bruserud, O. The chemokine system in allogeneic stem-cell transplantation: A possible therapeutic target? Expert Rev. Hematol. 2011, 4, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Apelseth, T.O.; Hervig, T.A.; Wentzel-Larsen, T.; Bruserud, O. Cytokine accumulation in photochemically treated and γ-irradiated platelet concentrates during storage. Transfusion 2006, 46, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Apelseth, T.; Hervig, T. Comments on “release of immune modulation factors from platelet concentrates during storage after photochemical pathogen inactivation treatment”. Transfusion 2009, 49, 603–604. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients/Donors | Procedure | Pre-Procedure OPN (ng/mL) | Post-Procedure OPN (ng/mL) | p Value | Apheresis Product OPN (ng/mL) |
|---|---|---|---|---|---|
| Allogeneic stem cell donors | G-CSF stimulation | 45 (27–62) | 50 (19–75) | 0.008 | - |
| Stem cell apheresis | 50 (19–75) | 56 (31–87) | 0.006 | 53 (29–73) | |
| Autologous stem cell donors | Stem cell apheresis | 89 (41–356) | 109 (55–473) | 0.008 | 86 (7–328) |
| Healthy platelet donors | Platelet apheresis | 44 (28–60) | 46 (33–56) | NS | 48 (25–75) 1 |
| Allogeneic HSC recipients | Allogeneic stem cell transplantation | 126 (80–438) | 103 (72–260) | NS | Not applicable |
| Patients/Donors | Procedure | Pre-Procedure G-CSF (pg/mL) | Post-Procedure G-CSF (pg/mL) | p Value | Apheresis Product G-CSF (pg/mL) |
| Allogeneic stem cell donors | G-CSF stimulation | 50 (22–241) | 10,780 (3687–31,947) | 0.0003 | 6673 (1704–21,152) |
| Autologous stem cell donors | G-CSF stimulation | Not determined | 18,366 (9861–46,314) | Not determined | 12,906 (8863–41,139) |
| Group | Age | Gender (M/F) | Total White Blood Cell Count in the Grafts | CD34+ Cells after G-CSF Treatment | ||
|---|---|---|---|---|---|---|
| Baseline Level (×109/L) | After G-CSF (×109/L) | PB Level (×103/mL) | Yield (×106/kg) | |||
| Allogeneic stem cell donors (n = 22) | 51 (25–77) | 14/8 | 5.9 (3.1–13.4) | 46.0 (30.1–76.3) | 44.1 (16.7–147.8) | 5.4 (0.8–22.4) |
| Autologous stem cell donors (n = 15) | 57 (44–67) | 9/6 | 5.4 (2.5–9.0) | 10.8 (2.7–43.7) | 39.9 (9.7–175.0) | 5.3 (1.1–27.9) |
| Platelet donors (n = 15) | 47 (26–62) | 8/7 | 6.0 (4.7–13.5) | - | - | - |
| Allogeneic HSCT recipients (n = 16) | 47 (35–63) | 7/9 | - | - | - | - |
| Cell Type | Graft Content (×108) | Infused Cells (×106/kg) | Post-Transplant Course 1 | |
|---|---|---|---|---|
| Total WBC | 791 (342–2495) | 109 (376–3054) | Neutrophil reconstitution | 17 (13–28) |
| CD34+ stem cells | 4.6 (2.4–6.7) | 5.5 (3.3–6.8) | Platelet reconstitution | 15 (11–39) |
| CD3+ T cells | 278 (71–490) | 39 (10–61) | aGVHD | 2/16 |
| Neutrophils | 285 (112–1048) | 45 (15–133) | cGVHD | 9/16 |
| Monocytes | 127 (18–563) | 16 (3–69) | Early death | 4/16 |
| Lymphocytes | 346 (105–759) | 50 (14–96) | Relapse | 4/16 |
| Platelets | 7068 (3176–11,449) | 9607 (3655–14,260) | - | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melve, G.K.; Ersvaer, E.; Akkök, Ç.A.; Ahmed, A.B.; Kristoffersen, E.K.; Hervig, T.; Bruserud, Ø. Immunomodulation Induced by Stem Cell Mobilization and Harvesting in Healthy Donors: Increased Systemic Osteopontin Levels after Treatment with Granulocyte Colony-Stimulating Factor. Int. J. Mol. Sci. 2016, 17, 1158. https://doi.org/10.3390/ijms17071158
Melve GK, Ersvaer E, Akkök ÇA, Ahmed AB, Kristoffersen EK, Hervig T, Bruserud Ø. Immunomodulation Induced by Stem Cell Mobilization and Harvesting in Healthy Donors: Increased Systemic Osteopontin Levels after Treatment with Granulocyte Colony-Stimulating Factor. International Journal of Molecular Sciences. 2016; 17(7):1158. https://doi.org/10.3390/ijms17071158
Chicago/Turabian StyleMelve, Guro Kristin, Elisabeth Ersvaer, Çiğdem Akalın Akkök, Aymen Bushra Ahmed, Einar K. Kristoffersen, Tor Hervig, and Øystein Bruserud. 2016. "Immunomodulation Induced by Stem Cell Mobilization and Harvesting in Healthy Donors: Increased Systemic Osteopontin Levels after Treatment with Granulocyte Colony-Stimulating Factor" International Journal of Molecular Sciences 17, no. 7: 1158. https://doi.org/10.3390/ijms17071158