
Knockdown of AMPKα2 Promotes Pulmonary Arterial Smooth Muscle Cells Proliferation via mTOR/Skp2/p27Kip1 Signaling Pathway

Abstract
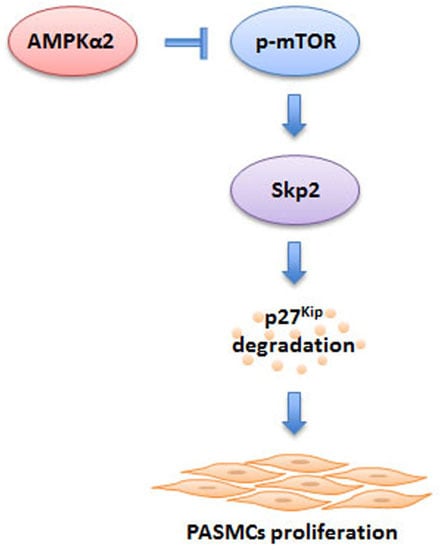
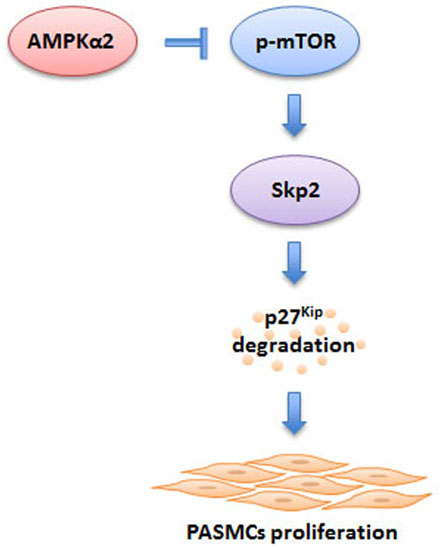
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
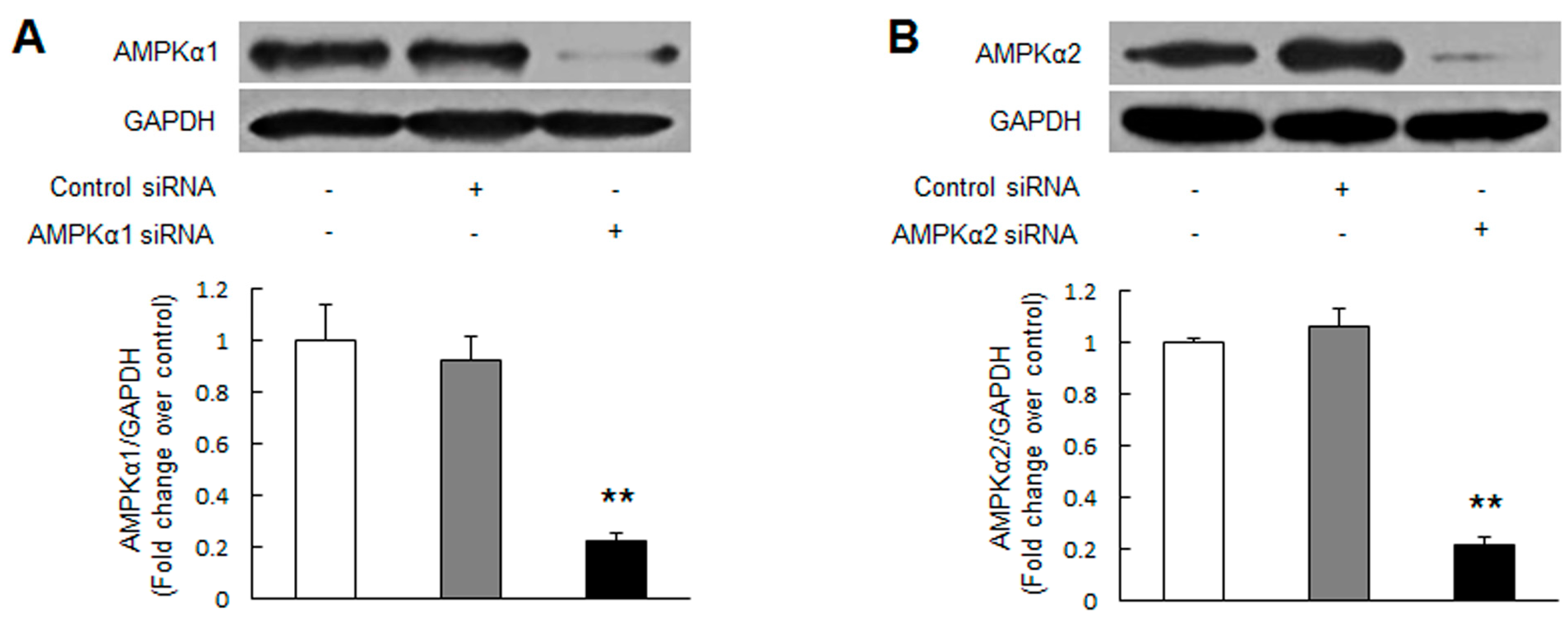
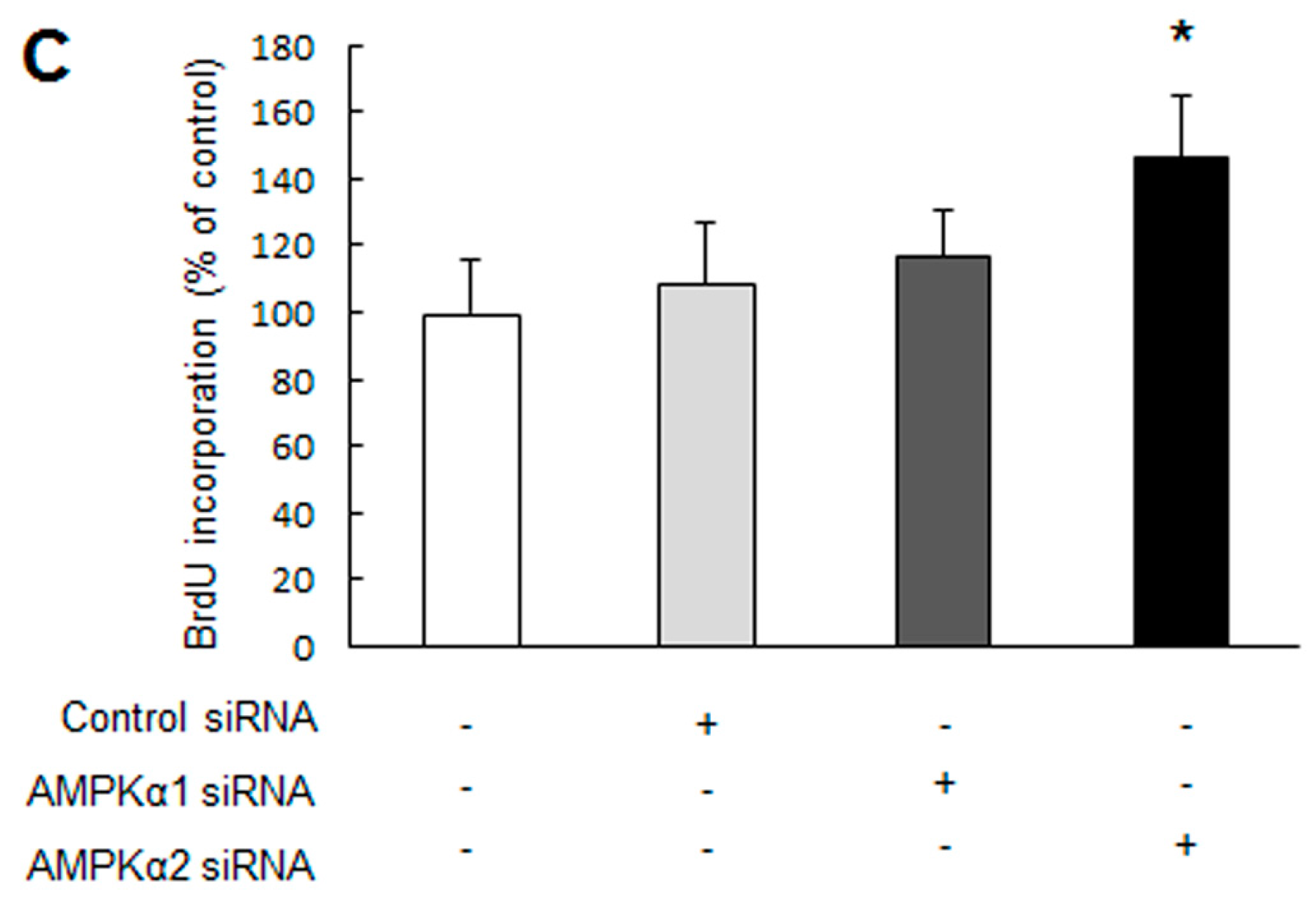
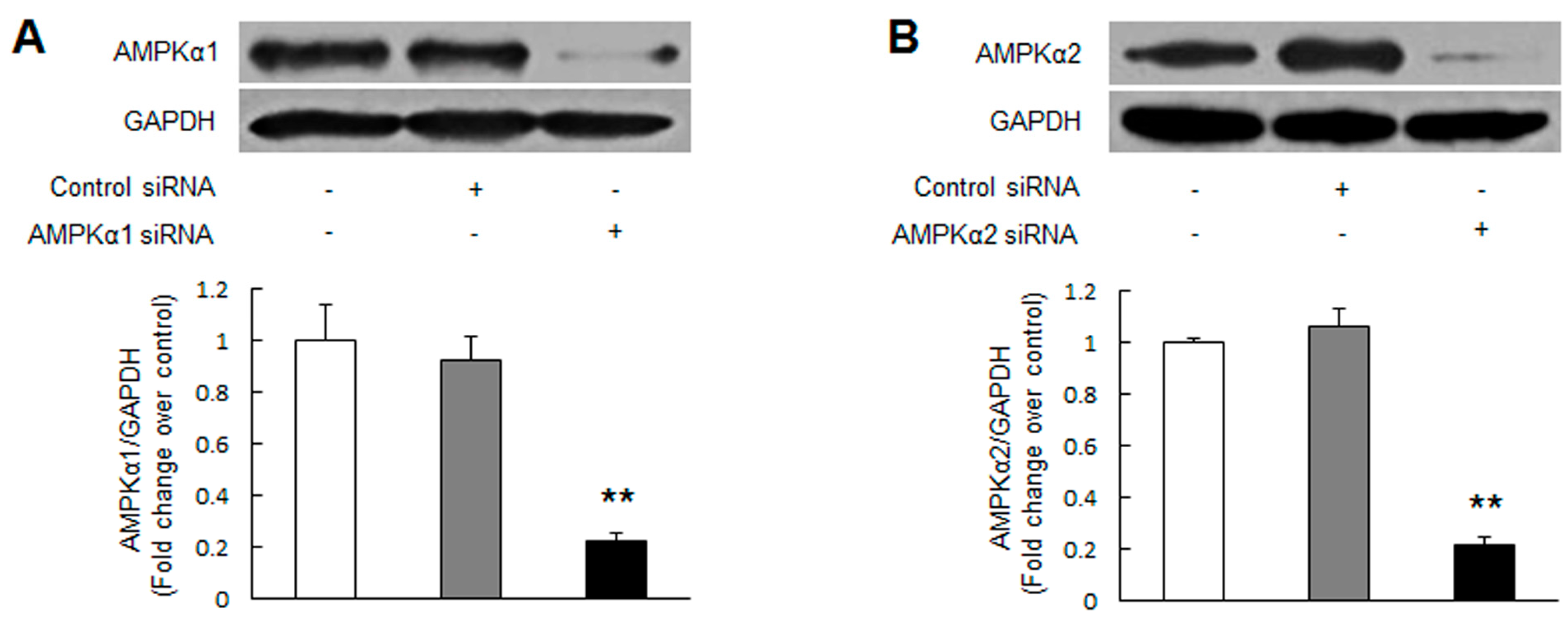
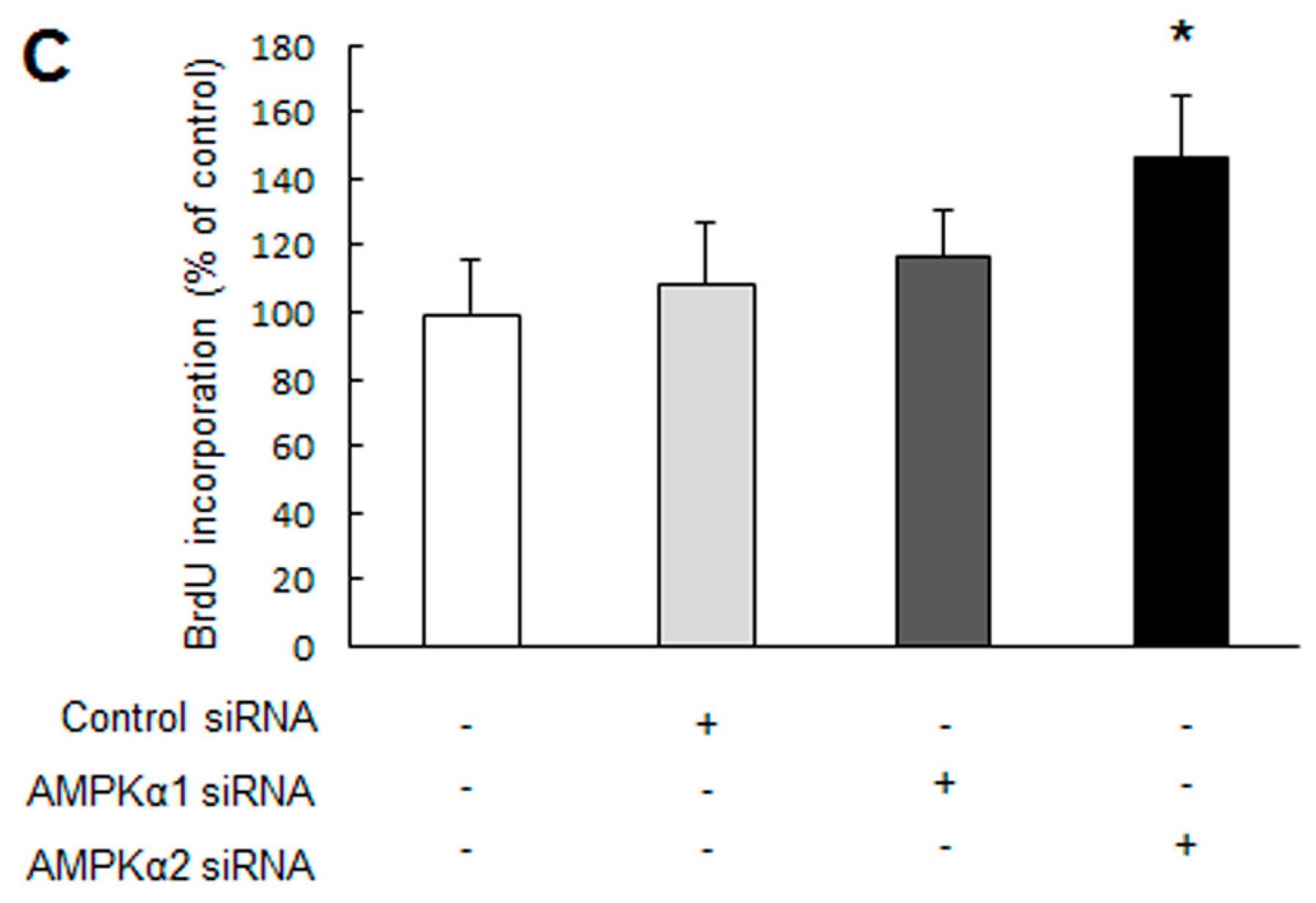
2.1. Knockdown of Adenosine Monophosphate-Activated Protein Kinase α2 (AMPKα2) Promotes Pulmonary Arterial Smooth Muscle Cells (PASMCs) Proliferation
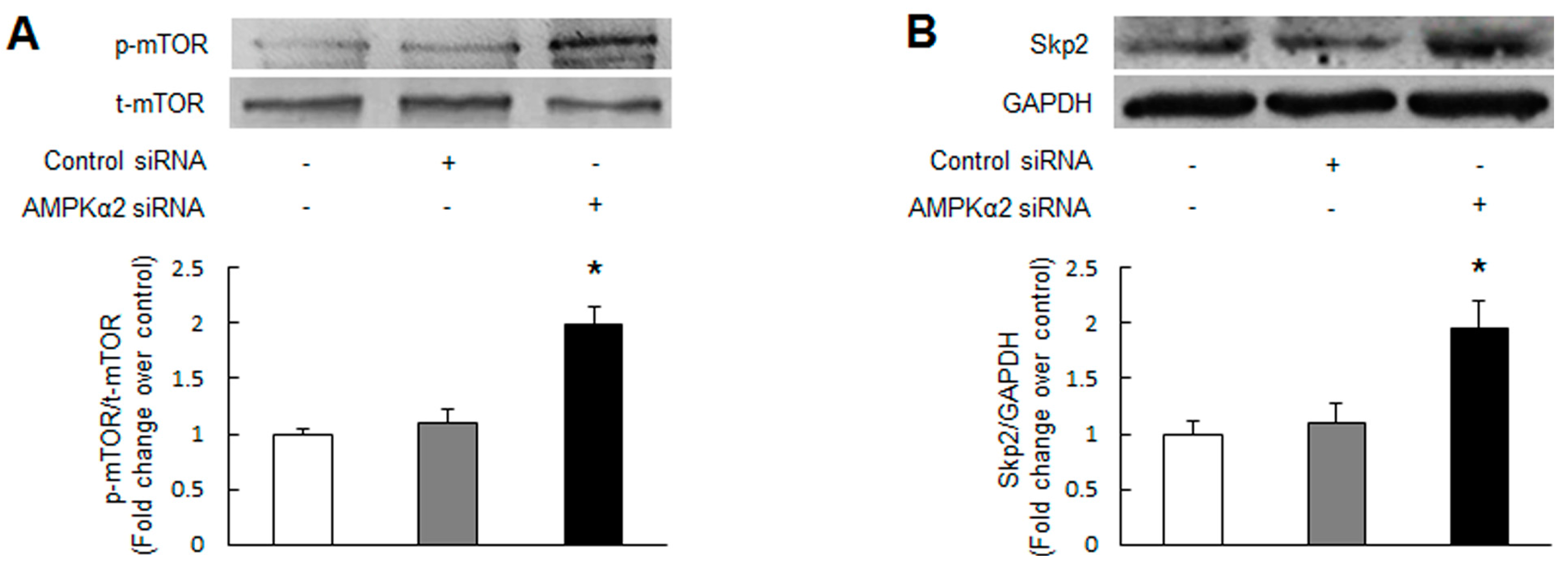
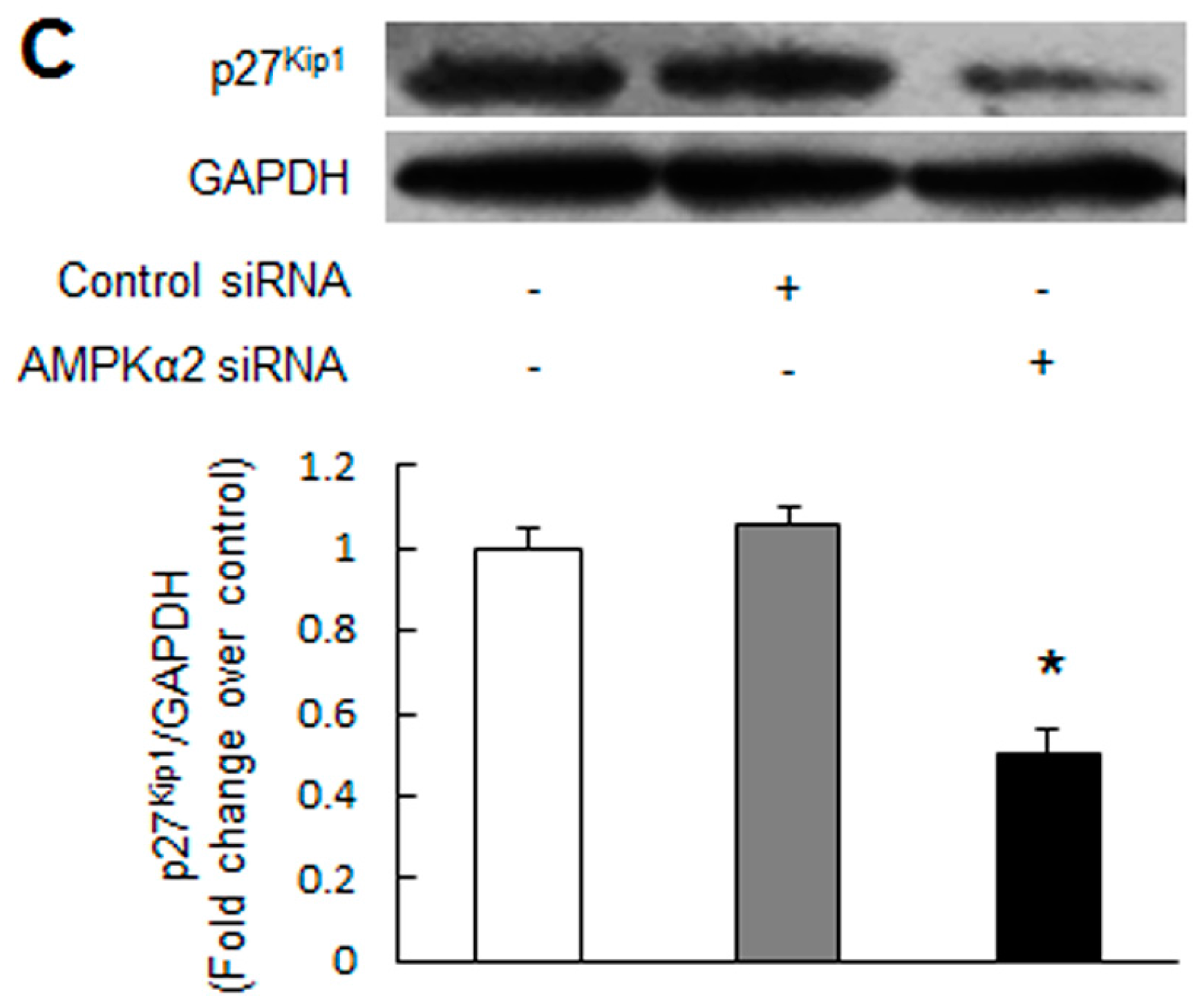
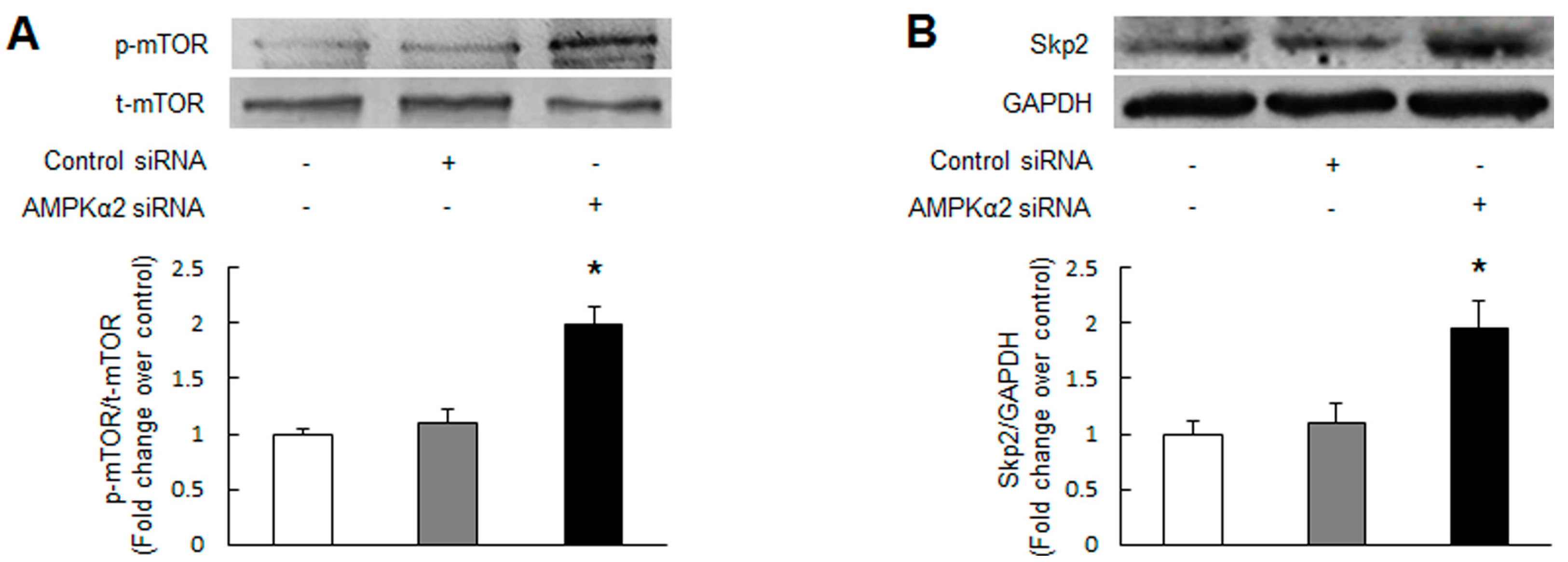
2.2. Knockdown of AMPKα2 Increases the Phosphorylation of Mammalian Target of Rapamycin (mTOR), Upregulates S-Phase Kinase-Associated Protein 2 (Skp2) and Downregulates p27Kip1
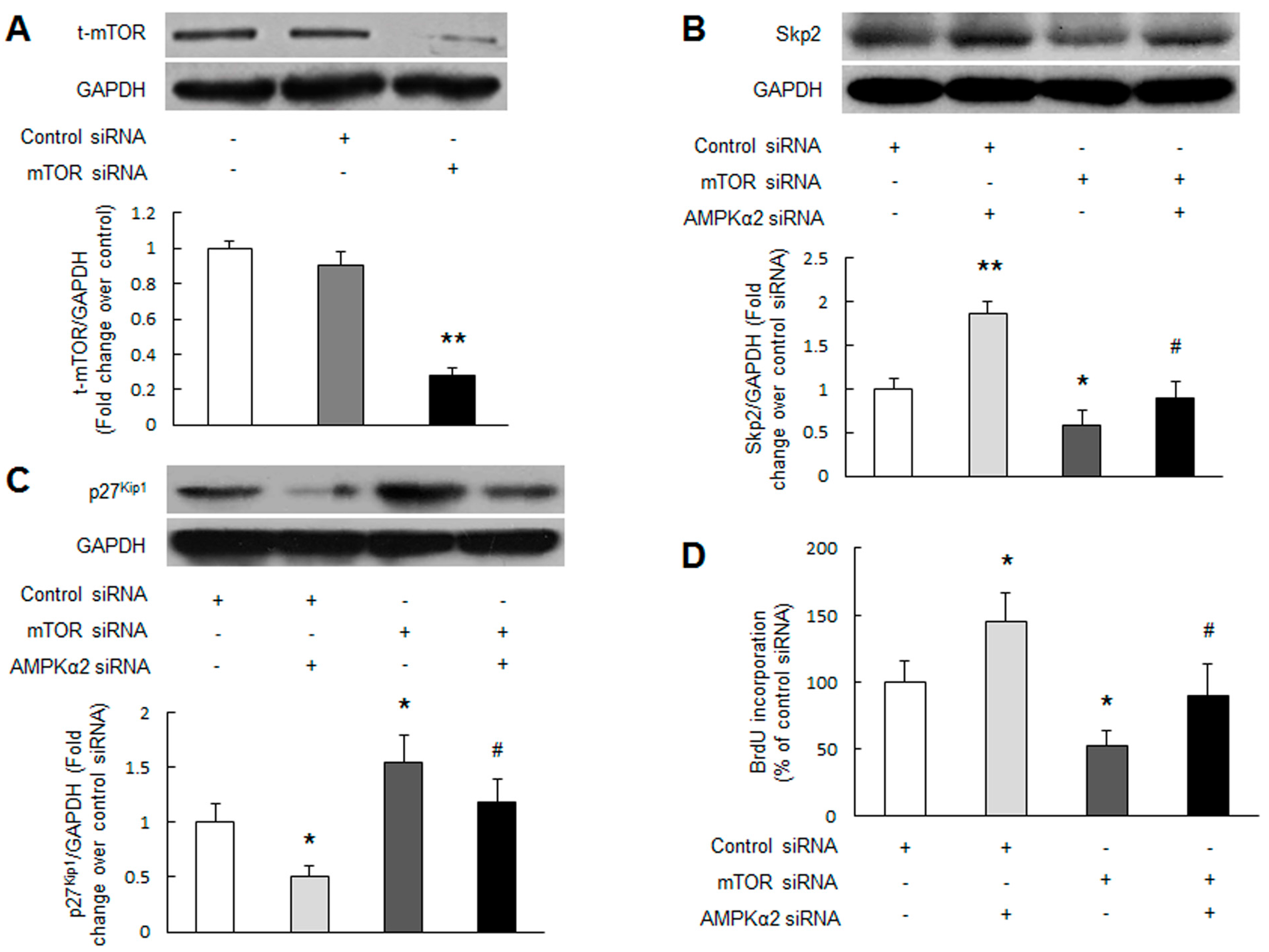
2.3. mTOR Mediates AMPKα2 Knockdown-Induced Skp2 Upregulation, p27Kip1 Downregulation and PASMCs Proliferation
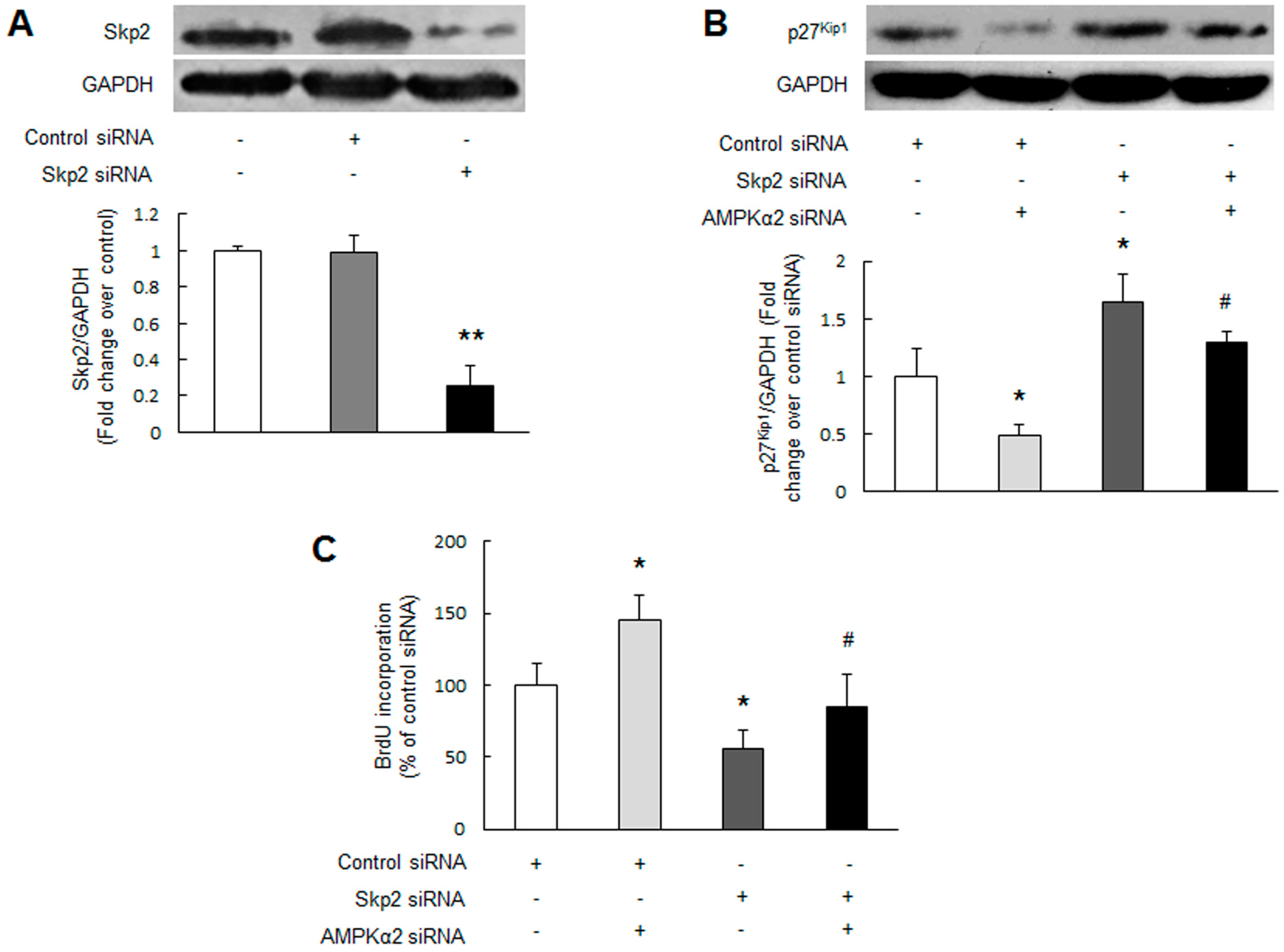
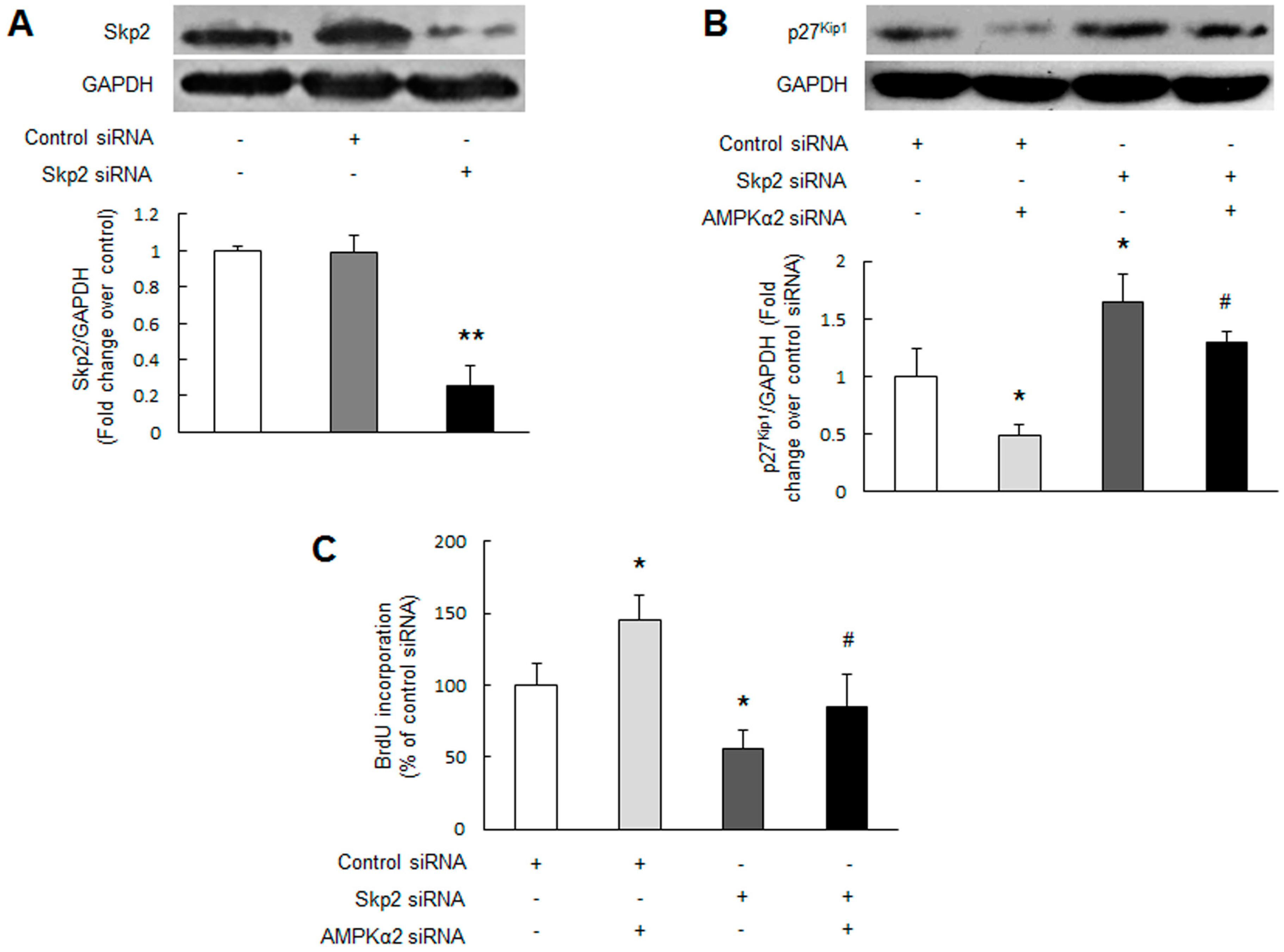
2.4. Skp2 Mediates AMPKα2 Knockdown-Induced p27Kip1 Reduction and PASMCs Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. siRNA Transfection
4.3. BrdU Incorporation Assay
4.4. Immunoblotting
4.5. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AICAR | 5-aminoimidazole-4-carboxamide ribonucleoside |
| AMPK | adenosine monophosphate-activated protein kinase |
| CDKs | cyclin-dependent kinases |
| mTOR | mammalian target of rapamycin |
| PAH | pulmonary arterial hypertension |
| PASMCs | pulmonary arterial smooth muscle cells |
| RNAi | RNA interference |
| Skp2 | S-phase kinase-associated protein 2 |
| siRNA | small interfering RNA |
| TSC | tuberous sclerosis complex |
References
- Hoeper, M.M.; Bogaard, H.J.; Condliffe, R.; Frantz, R.; Khanna, D.; Kurzyna, M.; Langleben, D.; Manes, A.; Satoh, T.; Torres, F.; et al. Definitions and diagnosis of pulmonary hypertension. Turk Kardiyol. Dern. Arsivi 2014, 42, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Hardegree, E.L.; Sachdev, A.; Fenstad, E.R.; Villarraga, H.R.; Frantz, R.P.; McGoon, M.D.; Oh, J.K.; Ammash, N.M.; Connolly, H.M.; Eidem, B.W.; et al. Impaired left ventricular mechanics in pulmonary arterial hypertension: Identification of a cohort at high risk. Circ. Heart Fail. 2013, 6, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, M.; Ruffenach, G.; Meloche, J.; Bonnet, S. Adaptation and remodelling of the pulmonary circulation in pulmonary hypertension. Can. J. Cardiol. 2015, 31, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Tajsic, T.; Morrell, N.W. Smooth muscle cell hypertrophy, proliferation, migration and apoptosis in pulmonary hypertension. Compr. Physiol. 2011, 1, 295–317. [Google Scholar] [PubMed]
- Schermuly, R.T.; Ghofrani, H.A.; Wilkins, M.R.; Grimminger, F. Mechanisms of disease: Pulmonary arterial hypertension. Nat. Rev. Cardiol. 2011, 8, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, Y.; Shi, H.; Zhang, Y.; Wang, G.; Xu, J.; Lu, J.; Zhang, D.; Xie, X.; Han, D.; et al. Statins inhibit pulmonary artery smooth muscle cell proliferation by upregulation of HO-1 and p21WAF1. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. AMP-activated protein kinase: A key regulator of energy balance with many roles in human disease. J. Intern. Med. 2014, 276, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. Minireview: The AMP-activated protein kinase cascade: The key sensor of cellular energy status. Endocrinology 2003, 144, 5179–5183. [Google Scholar] [CrossRef] [PubMed]
- Winder, W.W. Energy-sensing and signaling by AMP-activated protein kinase in skeletal muscle. J. Appl. Physiol. 2001, 91, 1017–1028. [Google Scholar] [PubMed]
- Vasamsetti, S.B.; Karnewar, S.; Kanugula, A.K.; Thatipalli, A.R.; Kumar, J.M.; Kotamraju, S. Metformin inhibits monocyte-to-macrophage differentiation via AMPK-mediated inhibition of STAT3 activation: Potential role in atherosclerosis. Diabetes 2015, 64, 2028–2041. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhao, L.; Qin, Y.; Zhang, M.; He, Y. Resveratrol inhibits proliferation and induces apoptosis of nasopharyngeal carcinoma cell line C666-1 through AMPK activation. Pharmazie 2015, 70, 399–403. [Google Scholar] [PubMed]
- Hardie, D.G. The AMP-activated protein kinase pathway—New players upstream and downstream. J. Cell Sci. 2004, 117, 5479–5487. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. AMP-activated protein kinase: An energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, Y.; Yang, Z.; Ahmad, I.; Nixon, C.; Salt, I.P.; Leung, H.Y. AMP-activated protein kinase (AMPK) as a potential therapeutic target independent of PI3K/Akt signaling in prostate cancer. Oncoscience 2014, 1, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.W.; Zhao, L.; Zhang, J.S.; Hou, Y.L.; Yu, Y.R.; Jia, M.Z.; Tang, C.S.; Qi, Y.F. Intermedin1–53 protects against cardiac hypertrophy by inhibiting endoplasmic reticulum stress via activating AMP-activated protein kinase. J. Hypertens. 2015, 33, 1676–1687. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Hayakawa, M.; Koga, H.; Torimura, T. Effects of fucoidan on proliferation, AMP-activated protein kinase, and downstream metabolism- and cell cycle-associated molecules in poorly differentiated human hepatoma HLF cells. Int. J. Oncol. 2015, 46, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, L.; Zhang, Y.; Wang, G.; Han, D.; Ke, R.; Li, S.; Feng, W.; Li, M. Activation of AMPK inhibits pulmonary arterial smooth muscle cells proliferation. Exp. Lung Res. 2014, 40, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Fan, R.; Lu, Y.; Yu, C.; Xu, X.; Zhang, X.; Liu, P.; Yan, S.; Chen, C.; Wang, L. Regulatory effect of AMP-activated protein kinase on pulmonary hypertension induced by chronic hypoxia in rats: In vivo and in vitro studies. Mol. Biol. Rep. 2014, 41, 4031–4041. [Google Scholar] [CrossRef] [PubMed]
- Agard, C.; Rolli-Derkinderen, M.; Dumas-de-La-Roque, E.; Rio, M.; Sagan, C.; Savineau, J.P.; Loirand, G.; Pacaud, P. Protective role of the antidiabetic drug metformin against chronic experimental pulmonary hypertension. Br. J. Pharmacol. 2009, 158, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Tian, R. Targeting AMPK for cardiac protection: Opportunities and challenges. J. Mol. Cell. Cardiol. 2011, 51, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Ming, M.; Sinnett-Smith, J.; Wang, J.; Soares, H.P.; Young, S.H.; Eibl, G.; Rozengurt, E. Dose-dependent AMPK-dependent and independent mechanisms of berberine and metformin inhibition of mTORC1, ERK, DNA synthesis and proliferation in pancreatic cancer cells. PLoS ONE 2014, 9, e114573. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zacharia, J.; Sun, X.; Wier, W.G. Effects of siRNA knock-down of TRPC6 and InsP3R1 in vasopressin-induced Ca2+ oscillations of A7r5 vascular smooth muscle cells. Pharmacol. Res. 2008, 58, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. mTOR: From growth signal integration to cancer, diabetes and ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Z.J.; Yi, W.; Rong, Y.W.; Kee, J.D.; Zhong, W.X. MicroRNA-1285 regulates 17β-estradiol-inhibited immature boar Sertoli cell proliferation via adenosine monophosphate-activated protein kinase activation. Endocrinology 2015, 4059–4070. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, L.; Liu, C.; Xia, T.; Zha, X.; Wang, S. Phenformin induces cell cycle change, apoptosis, and mesenchymal-epithelial transition and regulates the AMPK/mTOR/p70s6k and MAPK/ERK pathways in breast cancer cells. PLoS ONE 2015, 10, e0131207. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.K.; Wang, Y.J.; Wang, Y.; Tang, Z.Y.; Tan, P.; Huang, W.; Liu, Y.S. Adiponectin attenuates the osteoblastic differentiation of vascular smooth muscle cells through the AMPK/mTOR pathway. Exp. Cell Res. 2014, 323, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Larrea, M.D.; Wander, S.A.; Slingerland, J.M. p27 as Jekyll and Hyde: Regulation of cell cycle and cell motility. Cell Cycle 2009, 8, 3455–3461. [Google Scholar] [CrossRef] [PubMed]
- Auld, C.A.; Fernandes, K.M.; Morrison, R.F. Skp2-mediated p27Kip1 degradation during S/G2 phase progression of adipocyte hyperplasia. J. Cell. Physiol. 2007, 211, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, T.; Wei, T.Q.; Wei, X.; Li, S.F.; Wang, K.J.; Li, H. Skp2-mediated degradation of p27 regulates cell cycle progression in compressed human bladder smooth muscle cells. Kaohsiung J. Med. Sci. 2014, 30, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Shapira, M.; Kakiashvili, E.; Rosenberg, T.; Hershko, D.D. The mTOR inhibitor rapamycin down-regulates the expression of the ubiquitin ligase subunit Skp2 in breast cancer cells. Breast Cancer Res. 2006, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.R.; Wang, Y.; Wang, Y.; Zhang, J.J.; Zhang, J.H.; Lu, C.; Wang, X.Z. mTOR is involved in 17β-estradiol-induced, cultured immature boar Sertoli cell proliferation via regulating the expression of SKP2, CCND1, and CCNE1. Mol. Reprod. Dev. 2015, 82, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Totary-Jain, H.; Sanoudou, D.; Dautriche, C.N.; Schneller, H.; Zambrana, L.; Marks, A.R. Rapamycin resistance is linked to defective regulation of Skp2. Cancer Res. 2012, 72, 1836–1843. [Google Scholar] [CrossRef] [PubMed]
- Motoshima, H.; Goldstein, B.J.; Igata, M.; Araki, E. AMPK and cell proliferation—AMPK as a therapeutic target for atherosclerosis and cancer. J. Physiol. 2006, 574, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Gogol, M.; Gaudenz, K.; Gerton, J.L. Improved transcription and translation with l-leucine stimulation of mTORC1 in Roberts syndrome. BMC Genom. 2016, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Houssaini, A.; Abid, S.; Mouraret, N.; Wan, F.; Rideau, D.; Saker, M.; Marcos, E.; Tissot, C.M.; Dubois-Rande, J.L.; Amsellem, V.; et al. Rapamycin reverses pulmonary artery smooth muscle cell proliferation in pulmonary hypertension. Am. J. Respir. Cell Mol. Biol. 2013, 48, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.P.; Li, X.H.; Gong, S.X.; Li, W.Q.; Hu, C.P.; Zhang, Z.; Li, Y.J. miR-100 suppresses mTOR signaling in hypoxia-induced pulmonary hypertension in rats. Eur. J. Pharmacol. 2015, 765, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Lee, K.K.; Zhang, L.; Gerton, J.L. Stimulation of mTORC1 with l-leucine rescues defects associated with Roberts syndrome. PLoS Genet. 2013, 9, e1003857. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Ouyang, H.; Zhu, T.; Lindvall, C.; Wang, Y.; Zhang, X.; Yang, Q.; Bennett, C.; Harada, Y.; Stankunas, K.; et al. TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth. Cell 2006, 126, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.I.; Nakayama, K. Ubiquitin ligases: Cell-cycle control and cancer. Nat. Rev. Cancer 2006, 6, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Nagahama, H.; Minamishima, Y.A.; Miyake, S.; Ishida, N.; Hatakeyama, S.; Kitagawa, M.; Iemura, S.; Natsume, T.; Nakayama, K.I. Skp2-mediated degradation of p27 regulates progression into mitosis. Dev. Cell 2004, 6, 661–672. [Google Scholar] [CrossRef]
- Gstaiger, M.; Jordan, R.; Lim, M.; Catzavelos, C.; Mestan, J.; Slingerland, J.; Krek, W. Skp2 is oncogenic and overexpressed in human cancers. Proc. Natl. Acad. Sci. USA 2001, 98, 5043–5048. [Google Scholar] [CrossRef] [PubMed]
- Signoretti, S.; di Marcotullio, L.; Richardson, A.; Ramaswamy, S.; Isaac, B.; Rue, M.; Monti, F.; Loda, M.; Pagano, M. Oncogenic role of the ubiquitin ligase subunit Skp2 in human breast cancer. J. Clin. Investig. 2002, 110, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Ayala, G.; de Marzo, A.; Tian, W.; Frolov, A.; Wheeler, T.M.; Thompson, T.C.; Harper, J.W. Elevated Skp2 protein expression in human prostate cancer: Association with loss of the cyclin-dependent kinase inhibitor p27 and PTEN and with reduced recurrence-free survival. Clin. Cancer Res. 2002, 8, 3419–3426. [Google Scholar] [PubMed]
- Chen, H.; Mo, X.; Yu, J.; Huang, S.; Huang, Z.; Gao, L. Interference of Skp2 effectively inhibits the development and metastasis of colon carcinoma. Mol. Med. Rep. 2014, 10, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Bond, M.; Wu, Y.J. Proliferation unleashed: The role of Skp2 in vascular smooth muscle cell proliferation. Front. Biosci. 2011, 16, 1517–1535. [Google Scholar] [CrossRef]
- Conradie, R.; Bruggeman, F.J.; Ciliberto, A.; Csikasz-Nagy, A.; Novak, B.; Westerhoff, H.V.; Snoep, J.L. Restriction point control of the mammalian cell cycle via the cyclin E/Cdk2:p27 complex. FEBS J. 2010, 277, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; James, M.K.; Larochelle, S.; Fisher, R.P.; Blain, S.W. p27Kip1 inhibits cyclin d-cyclin-dependent kinase 4 by two independent modes. Mol. Cell. Biol. 2009, 29, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Di Gennaro, E.; Bruzzese, F.; Avallone, A.; Budillon, A. New perspective for an old antidiabetic drug: Metformin as anticancer agent. Cancer Treat. Res. 2014, 159, 355–376. [Google Scholar] [PubMed]
- Song, Y.; Wu, Y.; Su, X.; Zhu, Y.; Liu, L.; Pan, Y.; Zhu, B.; Yang, L.; Gao, L.; Li, M. Activation of AMPK inhibits PDGF-induced pulmonary arterial smooth muscle cells proliferation and its potential mechanisms. Pharmacol. Res. 2016, 107, 117–124. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, R.; Liu, L.; Zhu, Y.; Li, S.; Xie, X.; Li, F.; Song, Y.; Yang, L.; Gao, L.; Li, M. Knockdown of AMPKα2 Promotes Pulmonary Arterial Smooth Muscle Cells Proliferation via mTOR/Skp2/p27Kip1 Signaling Pathway. Int. J. Mol. Sci. 2016, 17, 844. https://doi.org/10.3390/ijms17060844
Ke R, Liu L, Zhu Y, Li S, Xie X, Li F, Song Y, Yang L, Gao L, Li M. Knockdown of AMPKα2 Promotes Pulmonary Arterial Smooth Muscle Cells Proliferation via mTOR/Skp2/p27Kip1 Signaling Pathway. International Journal of Molecular Sciences. 2016; 17(6):844. https://doi.org/10.3390/ijms17060844
Chicago/Turabian StyleKe, Rui, Lu Liu, Yanting Zhu, Shaojun Li, Xinming Xie, Fangwei Li, Yang Song, Lan Yang, Li Gao, and Manxiang Li. 2016. "Knockdown of AMPKα2 Promotes Pulmonary Arterial Smooth Muscle Cells Proliferation via mTOR/Skp2/p27Kip1 Signaling Pathway" International Journal of Molecular Sciences 17, no. 6: 844. https://doi.org/10.3390/ijms17060844