1. Introduction
The DNA in each of our cells accumulates thousands of lesions every day. Cells are continuously challenged by DNA damage stimuli from various exogenous environmental factors, such as ultraviolet (UV) radiation, ionizing radiation (IR) and numerous chemical agents, or endogenous sources mainly represented by products of cellular metabolism. DNA damage can interfere with essential cellular processes, such as transcription or replication, and can compromise the viability of the cell. Specific DNA lesions can also induce mutations that cause cancer or other diseases, as well as contribute to the aging process [
1]. DNA damage at the telomeric chromosomal sections definitely has pro-aging consequences [
2]. Fortunately, cells have developed elaborate and efficient response pathways to preserve genomic stability. The DNA damage response (DDR) [
3] is an evolutionarily-conserved signaling cascade activated by DNA damage (DD), which directs cell fate toward DNA repair, senescence or apoptosis [
4]. DDR is a signaling network initiated by lesion recognition and amplified by multiple mediator signaling proteins, which eventually activate downstream effectors to modulate cell fate. Multiple DNA repair pathways have evolved to resolve various DNA lesions, including: base excision repair that removes damaged bases; mismatch repair that recognizes base incorporation errors and base damage; nucleotide excision repair that removes bulky DNA adducts; and cross-link repair that removes inter-strand cross-links. In addition, breaks in the DNA backbone are repaired via double-strand break (DSB) repair pathways, including homologous recombination (HR) and nonhomologous end joining [
5]. If DNA lesions are not properly repaired or amended during replication, they can be converted into permanent mutations. When this occurs at the sites of oncogenes or tumor suppressor genes, the risk of neoplastic cell transformation is significantly increased [
1,
6]. In higher organisms, successful DDR is thought to prevent neoplastic transformation in a cell-autonomous manner, by ensuring removal of cells with severely damaged DNA [
7]. DDR signaling has been suggested as a key mechanism linking DNA damage accumulation, cell senescence and organismal aging. In fact, during the cell lifetime, the genomic DNA is continuously exposed to different exogenous or endogenous factors that destabilize its integrity and functionality. Therefore, genomic instability and decline of DNA repair efficiency is considered one of the main drivers of the aging process [
8]. However, emerging data suggest that DDR signaling can also work through a paracrine/systemic mechanism, shaping the systemic environment through the regulation of tissue repair and immune responses [
9]. Given the fundamental role of DDR in maintaining genome integrity, this complex signaling network requires accurate regulatory mechanisms to respond to different types of DNA lesions in different stages of the cell cycle.
Recently, microRNAs (miRs) have emerged as important players in regulatory networks affecting the DNA damage/repair process in a wide range of physiological and pathological conditions [
5,
10]. miRs are 18–25 nucleotide non-coding RNAs that post-transcriptionally regulate gene expression stalling the translation of the cognate mRNA or promoting its degradation [
11]. miRs have been identified to influence physiological processes, such as development, growth and differentiation [
12], and have also been implicated in a wide range of diseases [
13]. Multiple miRs may target the same miR, and the majority of miRs contain multiple binding sites for miRs, generating a highly complex regulatory network system by which hundreds of genes involved in different signaling pathways can be regulated simultaneously [
14].
Nutrients and their bioactive compounds can modulate the miRs’ expression involved in many physiological and pathological processes [
15]. Nutrition is the process that offers different substances to an organism that can work as energy suppliers (carbohydrate and fat), as cell structure sources (proteins) and on metabolism control (vitamins and minerals), thereby maintaining its homeostasis. The importance of diet and nutrition in human health and disease is well established. Basic laboratory research, clinical trials and epidemiological studies demonstrated that nutrient-rich bioactive foods can induce epigenetic changes and alter genes’ expression by the alteration of the histone structure, DNA methylation and miRs’ modulation [
16].
Epi-gallocatechin-3-gallate (EGCG), resveratrol (RSV), curcumin (CRC) and n3-PUFA (n3-polyunsaturated fatty acids) are among the most studied compounds shown to have beneficial effects on human health [
17,
18]. EGCG, RSV and CRC are polyphenols present in fruits and vegetables. n3-PUFAs, are polyunsaturated fatty acids found in plants, as linolenic acid (ALA), or in fish eicosapentaenoic acid (EPA); and docosahexaenoic acid (DHA). Each of these four dietary compounds, at a concentration that potentially may be achievable in the organism, has been shown to suppress cell proliferation and induce apoptosis in certain types of human cancer cells [
18,
19]. In addition to potential anticancer activity, they possess cardiovascular protective properties [
20,
21] and beneficial effects on degenerative diseases [
22]. Furthermore, an anti-inflammatory and antioxidant activity has been frequently associated with these compounds [
23]. Most importantly, all of them may exert these effects by modulating miRs’ expressions [
24,
25]. In order to investigate and compare the impact of these specific food-derived compounds on DDR processes, we performed a literature search to identify miRs involved in DD/DDR and modulated by these dietary compounds. An
in silico analysis using the DIANA software web-server was applied to identify targets and pathways that play a major role in the DD/DDR modulation by these compounds [
26]. The results of the analysis of the pathways allowed us to speculate how food intervention could modulate DD/DDR.
3. Discussion
The maintenance of genome integrity by an efficient DNA repair is of paramount importance in the prevention of cancer, attenuation of aging processes and age-related degenerative diseases. Effective DDR is the key mechanism providing genome stability. miRs play a critical role in the regulation of DDR. In this in silico study, therefore, we examined the influence of dietary bioactive compounds on the miRs involved in DDR, which potentially may offer clues on the role of these compounds as potential chemopreventive and antiaging (geroprotective) modalities.
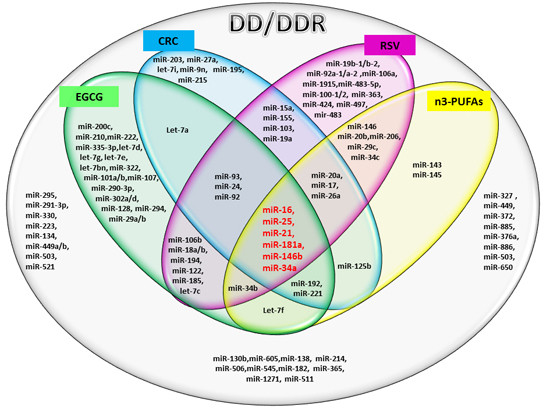
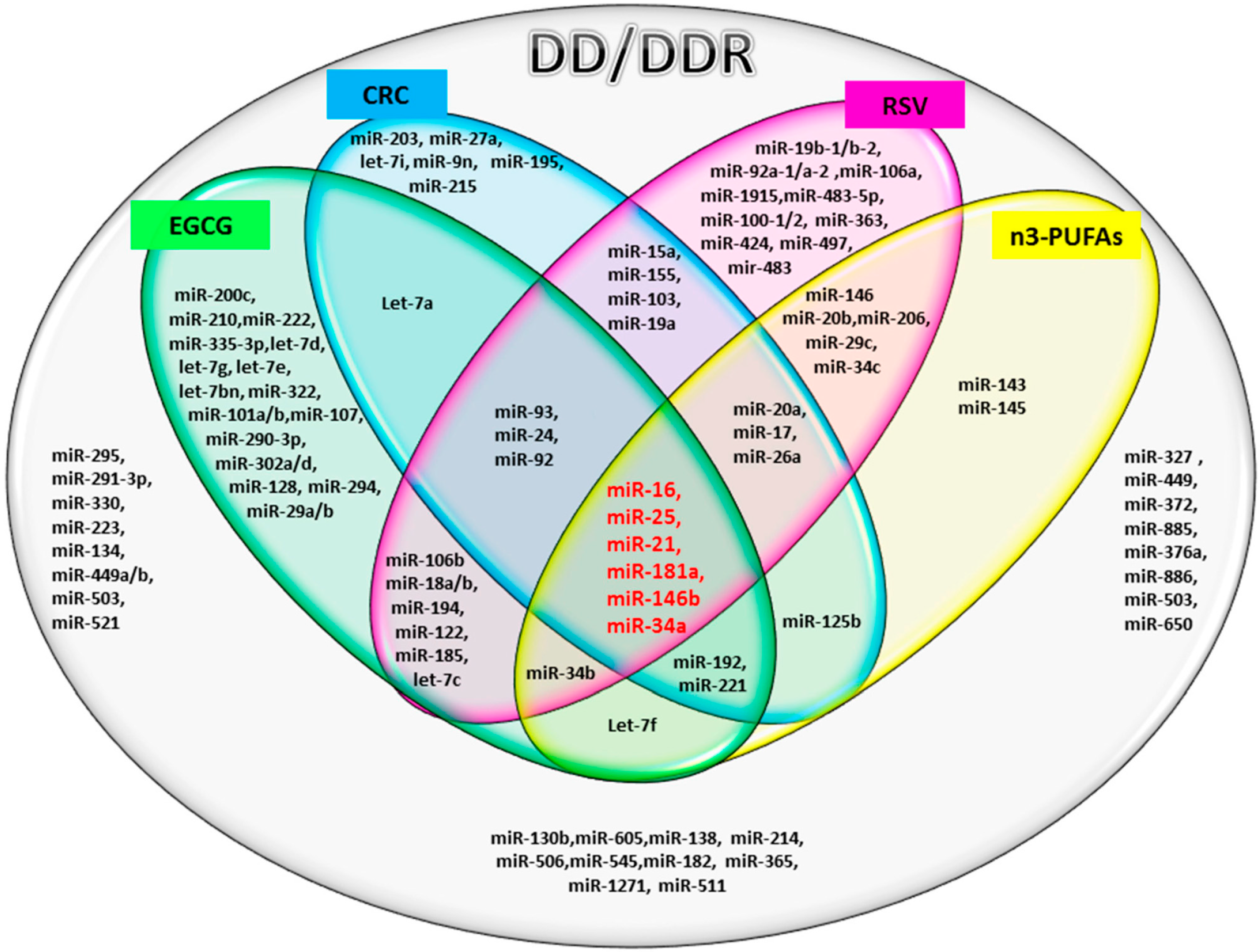
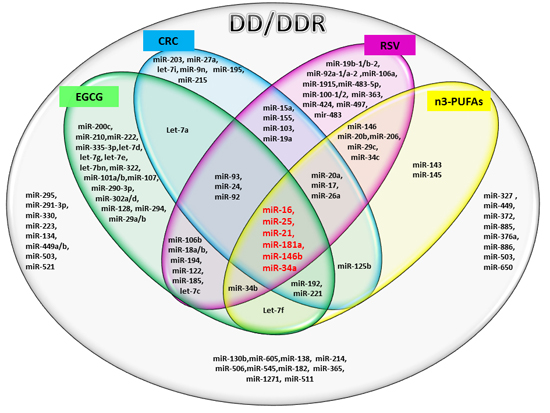
The results of our analysis show a strong impact of the compounds considered on important miRs involved in DD/DDR pathways. In particular, the analysis suggests a synergic action of all compounds in the modulation of six miRs. Among these common miRs, we found miR-21 and miR-146. It has recently been highlighted that miR-146, miR-155 and miR-21 have a kay role in the interplay among DDR, cell senescence, inflammation and age-related diseases [
9]. This trio of miRNAs, termed “inflamma-miRs”, has been primarily associated with chronic, low-grade inflammation known to characterize human aging and predisposing to age-related diseases [
67]. We here find that two of these miRs, DD/DDR-associated (miR-146 and miR-21), are modulated by all four studied compounds and one of them, miR-155, by CRC and RSV [
57]. miR-146 is one of the major miRs involved in orchestrating immune and inflammatory signaling via modulation of NF-kB activation [
9]. Both miR-146a and miR-146b-5p have been found to target the DSB repair key protein BRCA1 [
10].
miR-21 is a key modulator in many inflammatory pathways, and its aberrant expression in numerous cancers has led to its designation as an “onco-miR” [
68]. miR-21 is induced by DD, negatively regulating G1/S transition. It also participates in the DNA damage-induced G2/M checkpoint [
5]. It has been also shown that mR-21 negatively regulates Cdc25A and cell cycle progression in colon cancer cells [
69]. In addition, miR-21 is upregulated during hepatitis C virus infection and negatively regulates IFN-α signaling through MyD88 and IRAK1; it may thus be a potential therapeutic target for antiviral intervention [
70]. Recent observations suggest that miR-21 in cooperation with miR-145 (modulated by n3-PUFAs) play critical roles in the regulation of colon cancer stem cells [
71]. miR-155 has been reported to regulate inflammation and immune responses [
72]. miR-155 may have different functions in innate and adaptive immune responses, and the systemic diffusion of this DDR/-related miR may have either adverse or beneficial effects, depending on overall senescence/immunological host condition [
9]. In DDR network, miR-155 modulates cell cycle after DD by targeting key genes involved in cell cycle control, such as WEE1 [
73]. miR-155 has been shown to control the expression of TRF1, the protein that negatively regulates telomere length [
74]. Indeed, the TRF1 gene is known to be a target of miR-155 [
75]. Telomere length may also be regulated by miR-34a [
76], which is in turn modulated by all four compounds, as demonstrated by our
in silico analysis. miR-34b and miR-34c are also modulated by the three and two studied compounds, respectively. The miR-34 family, is a direct transcriptional target of p53, whose induction by DD and oncogenic stress pervades in diverse aspects of the DD response pathway [
27]. Noteworthy, the compounds being evaluated here modulate members of the miR-17/92 cluster and miR-106a/b clusters (DDR modulated), having an important role in cancer and other numerous diseases [
77]. In particular, our analysis reveals that RSV is able to modulate all of the miRs of these tree important clusters.
Our previous studies have demonstrated that RSV decreased the level of constitutive DD signaling by the reduced expression of γH2AX in proliferating A549, TK6 and WI-38 cells and in mitogenically-stimulated human lymphocytes [
78]. The reduction of γ-H2AX was paralleled by a drop in the level of endogenous ROS and a decline in mTOR/S6K1 signaling [
79]. H2AX is the initial sensor protein in the DSB response that can detect and mark DD by its phosphorylated form (γ-H2AX) [
80,
81]. H2AX is a target of miR-24 [
80]. The present analysis suggests that RSV, as well as CRC and EGCG might regulate γH2AX expression by miR-24 mediation.
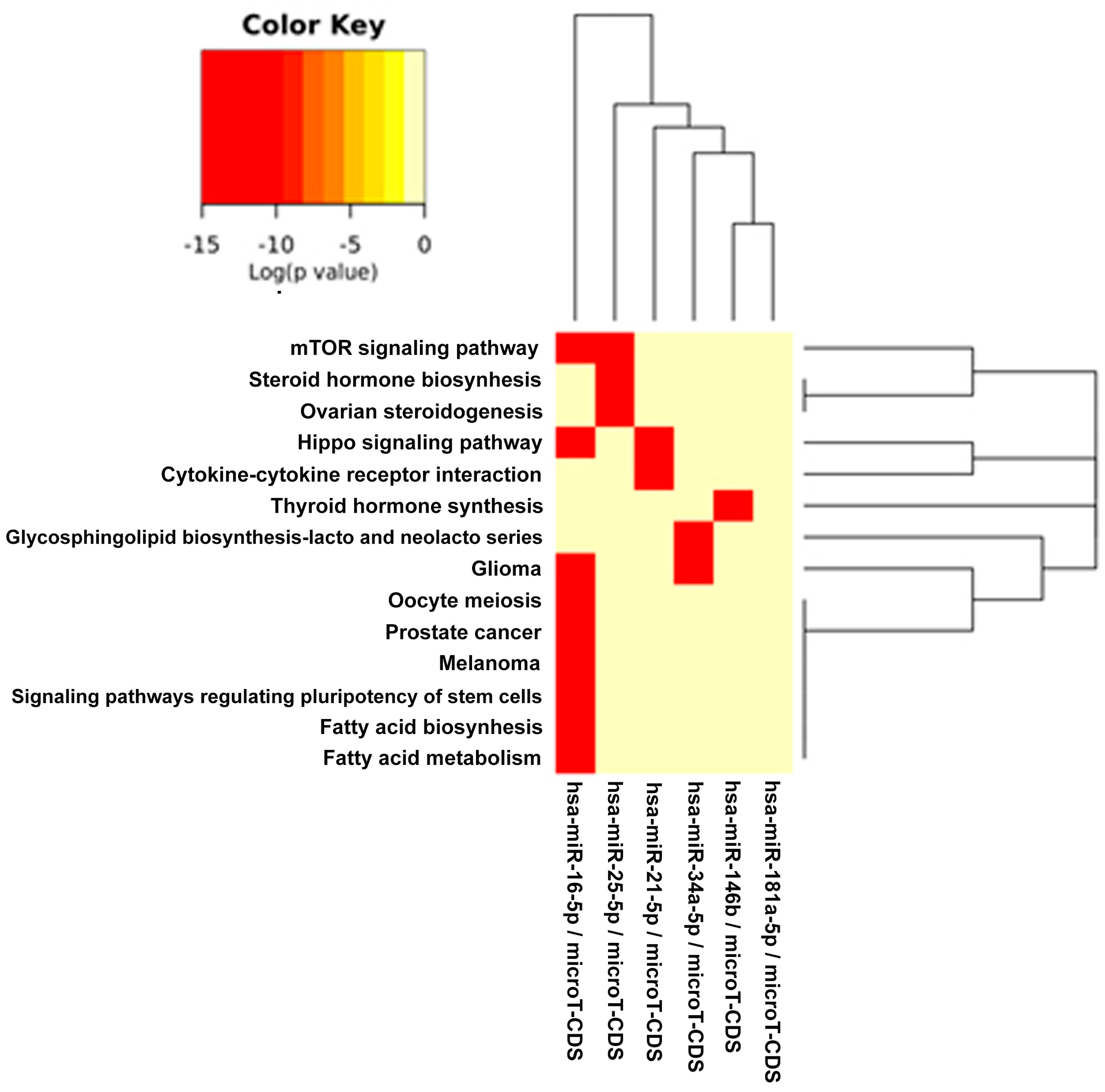
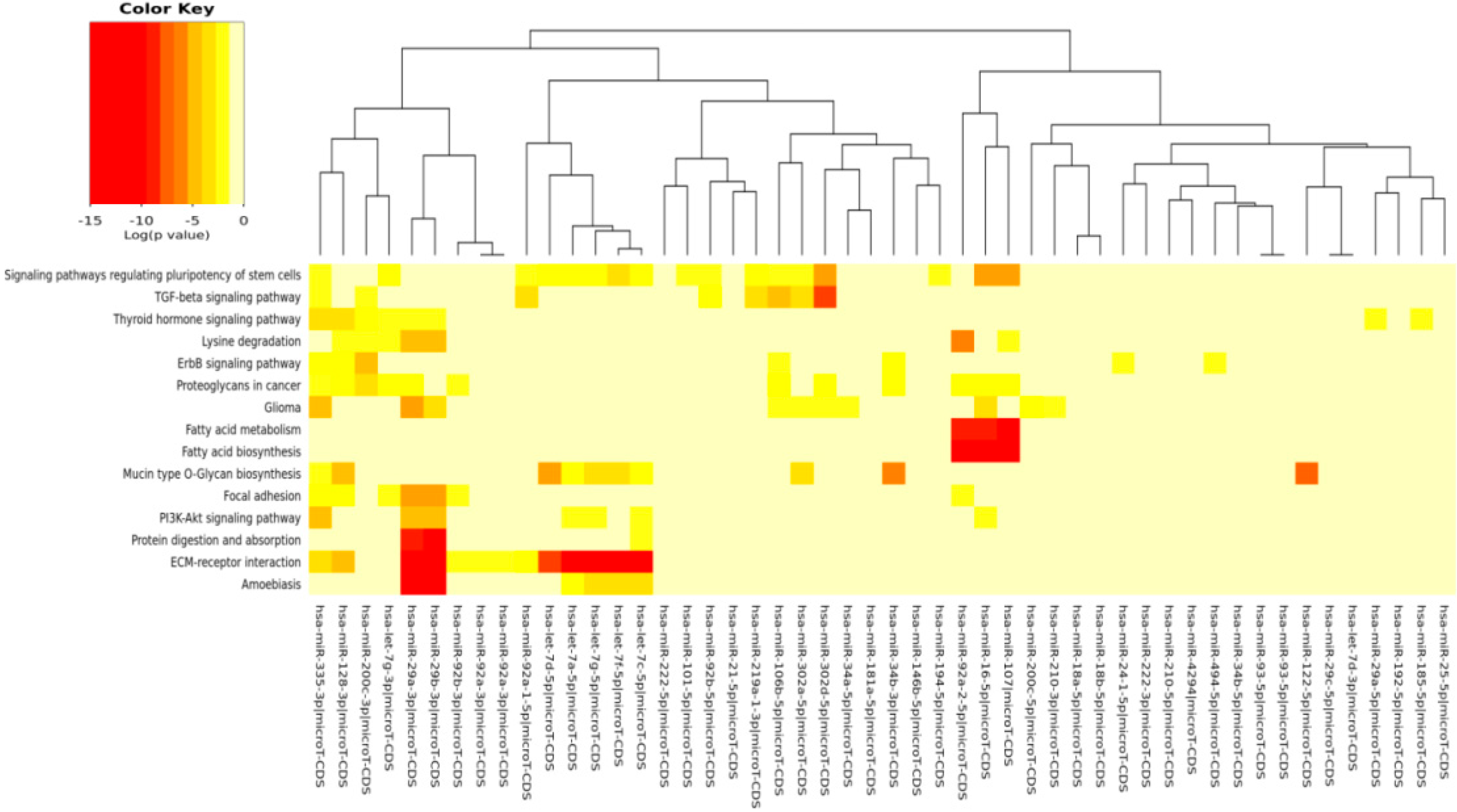
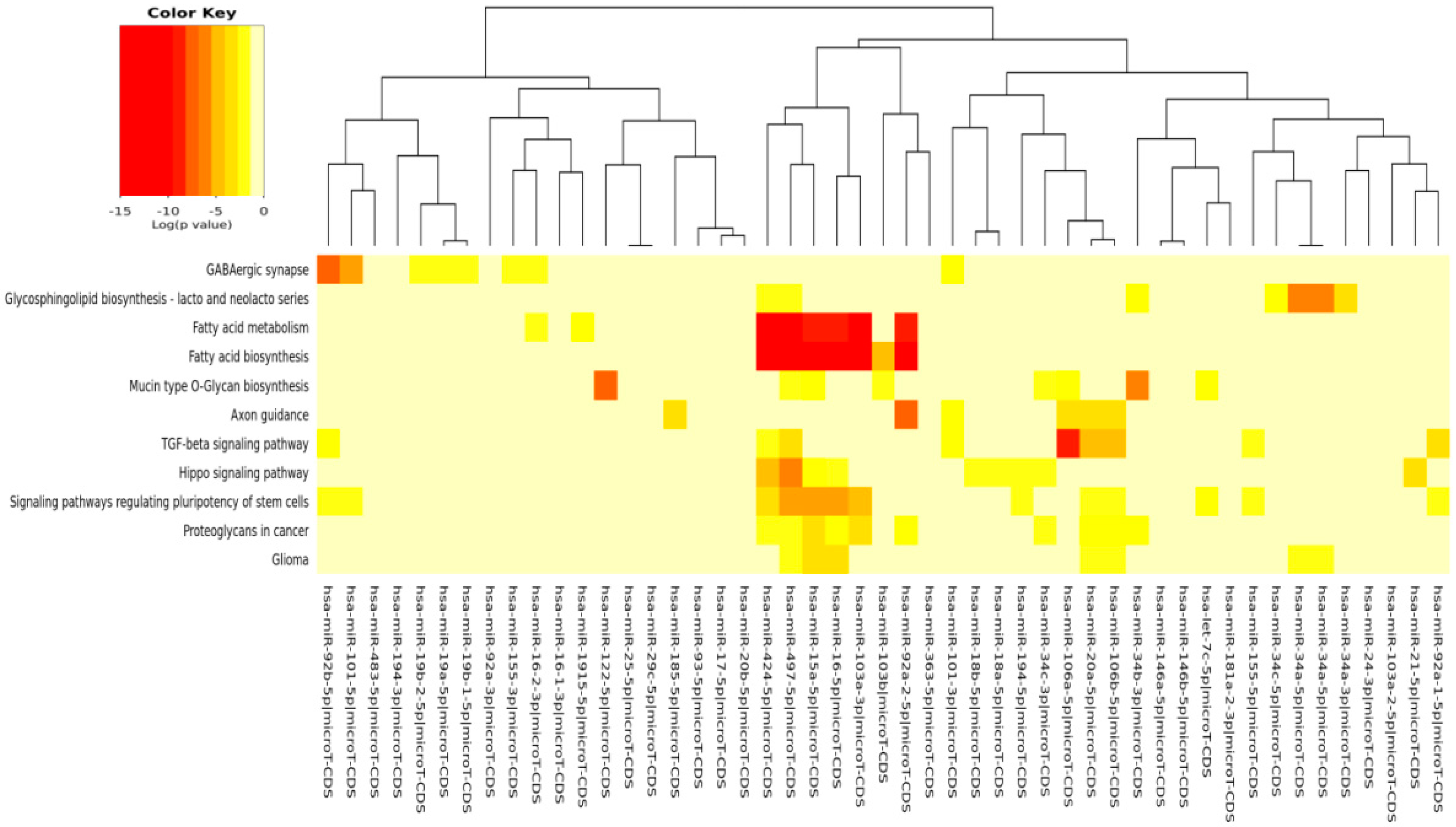
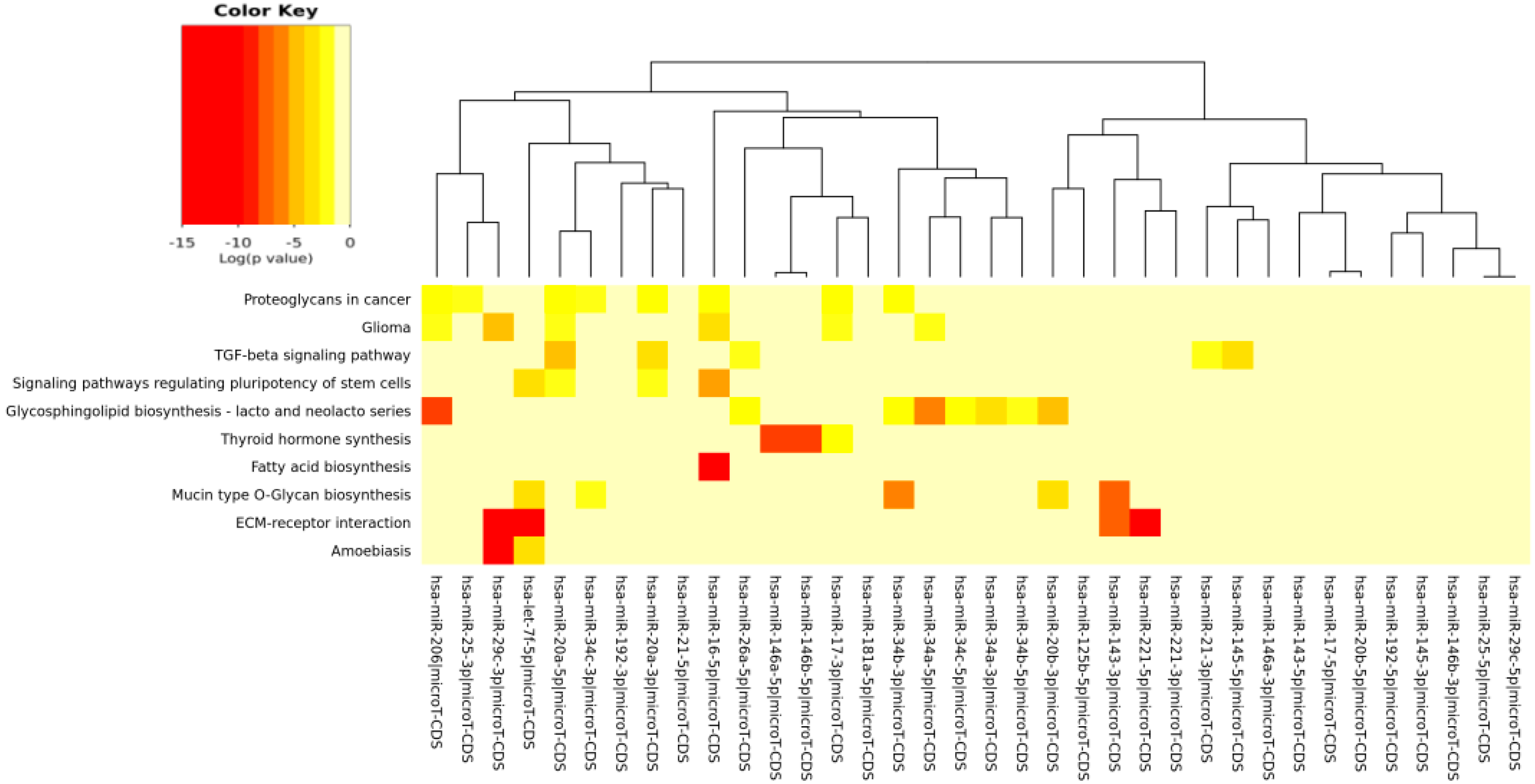
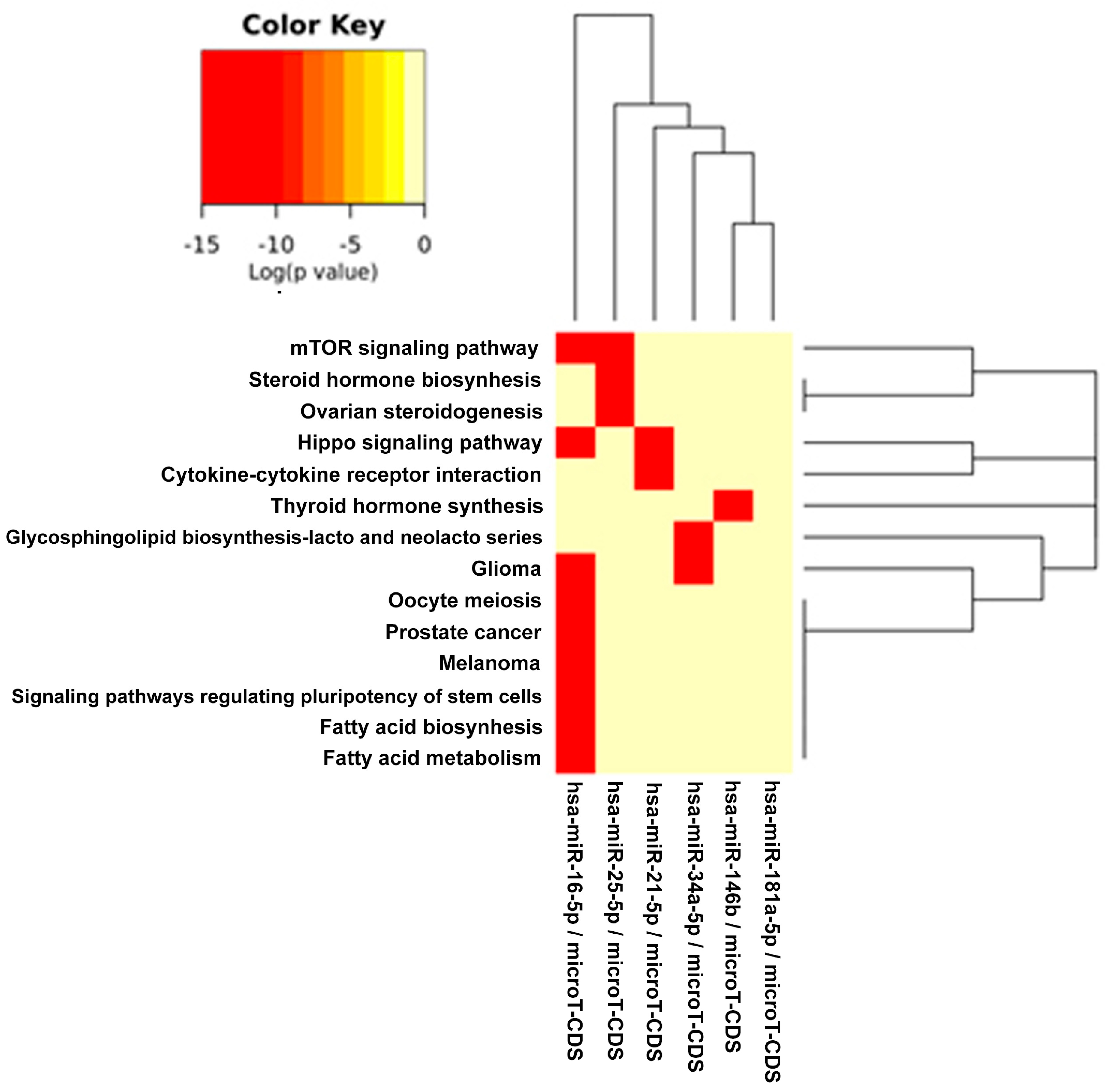
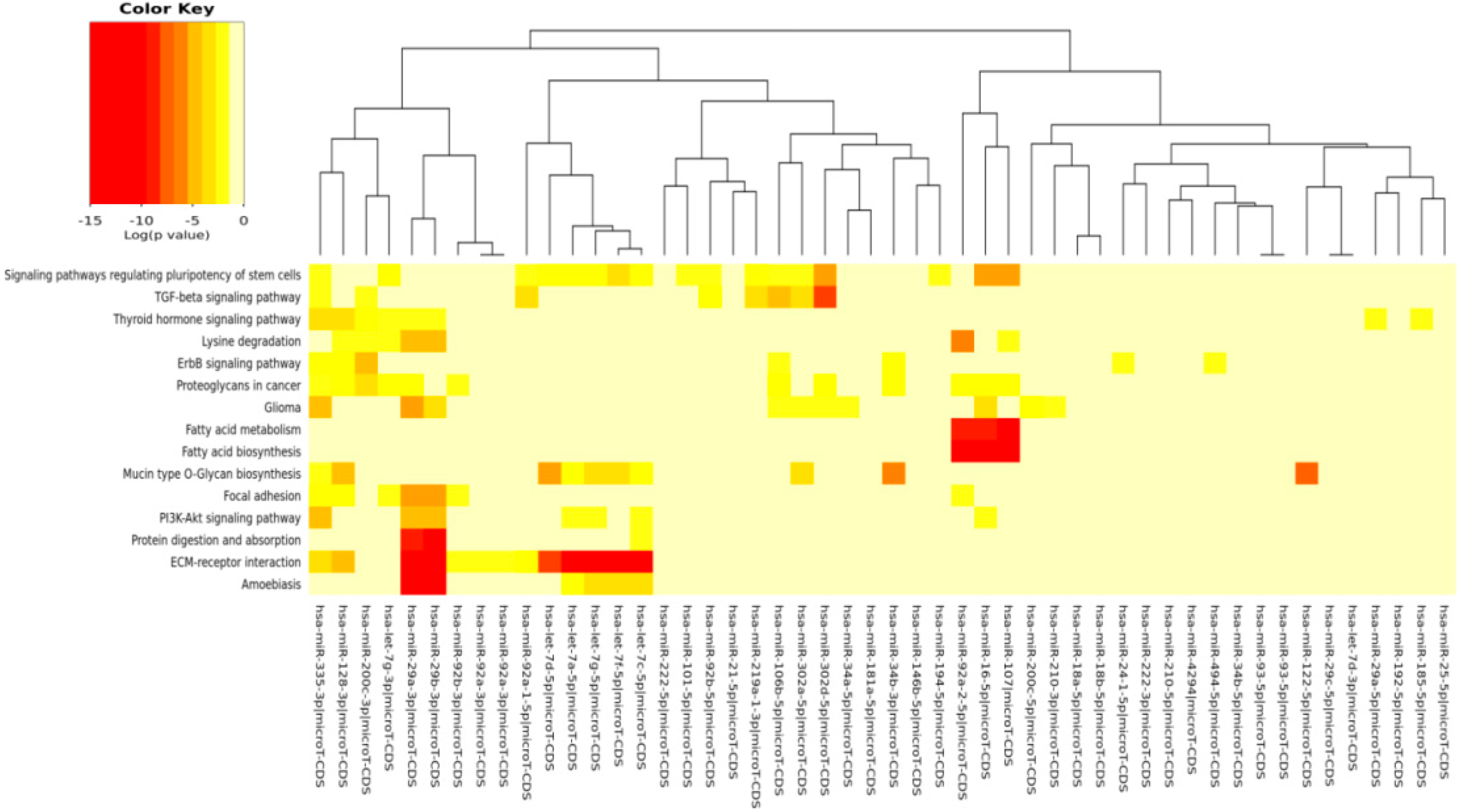
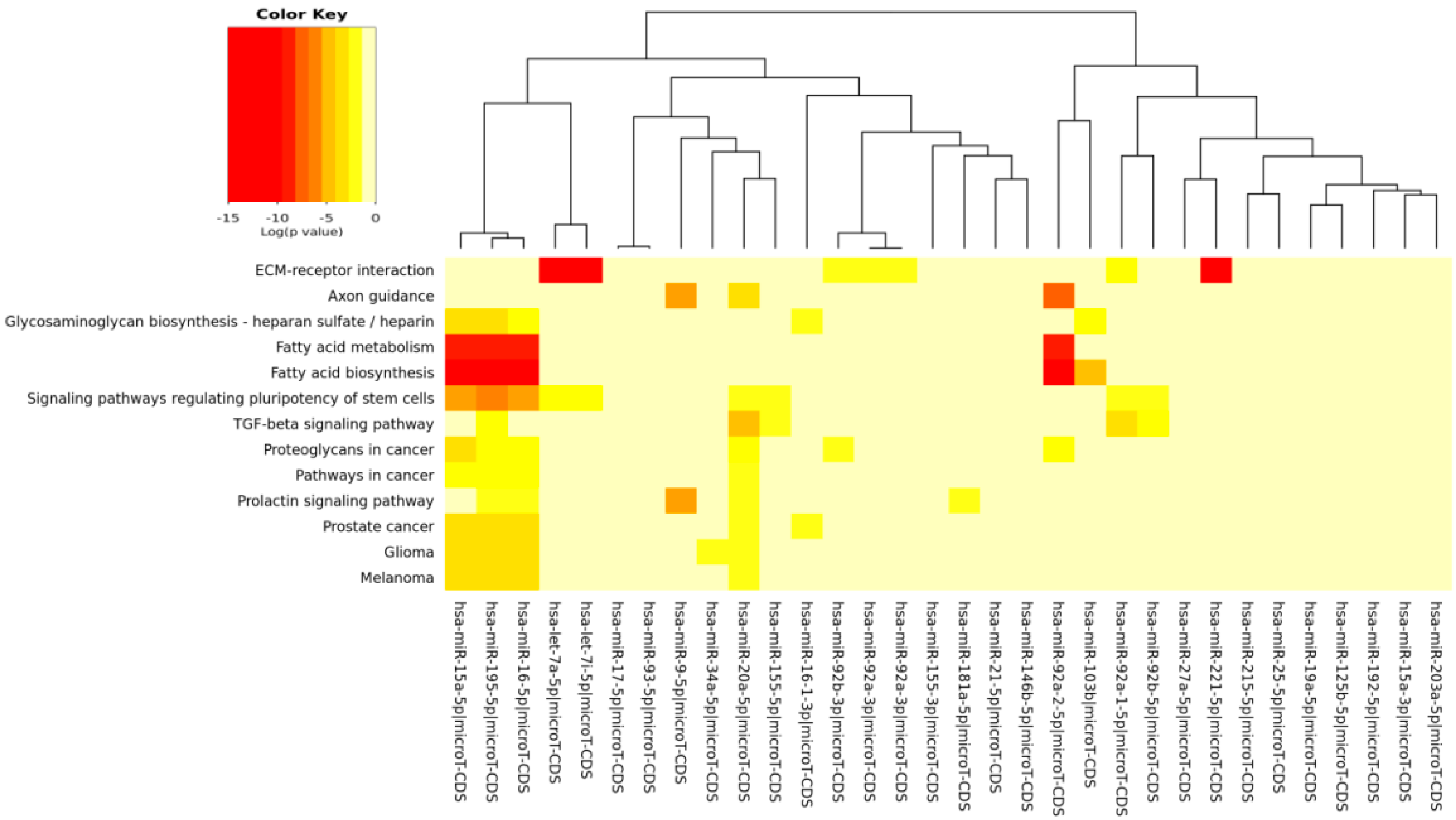
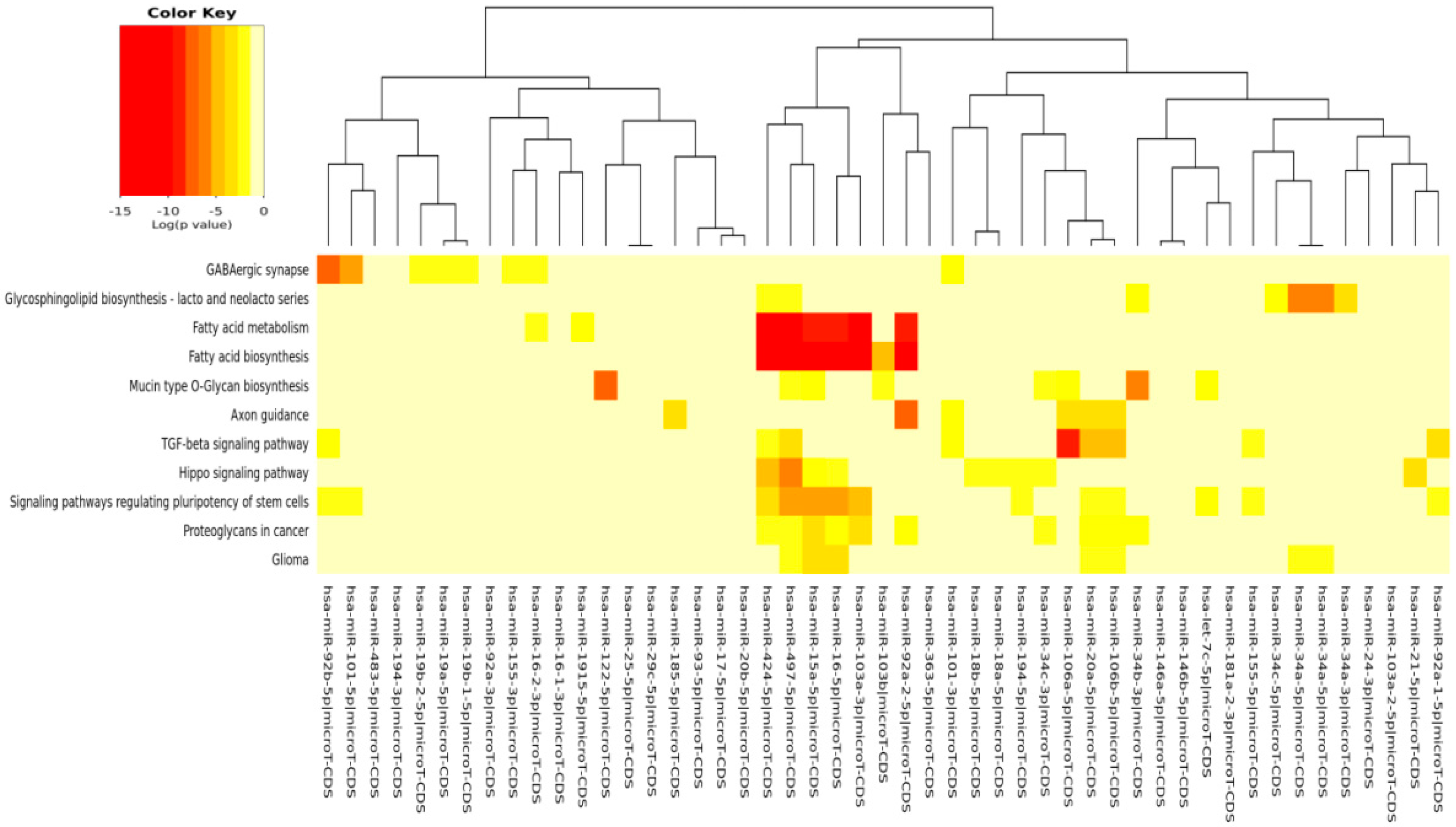
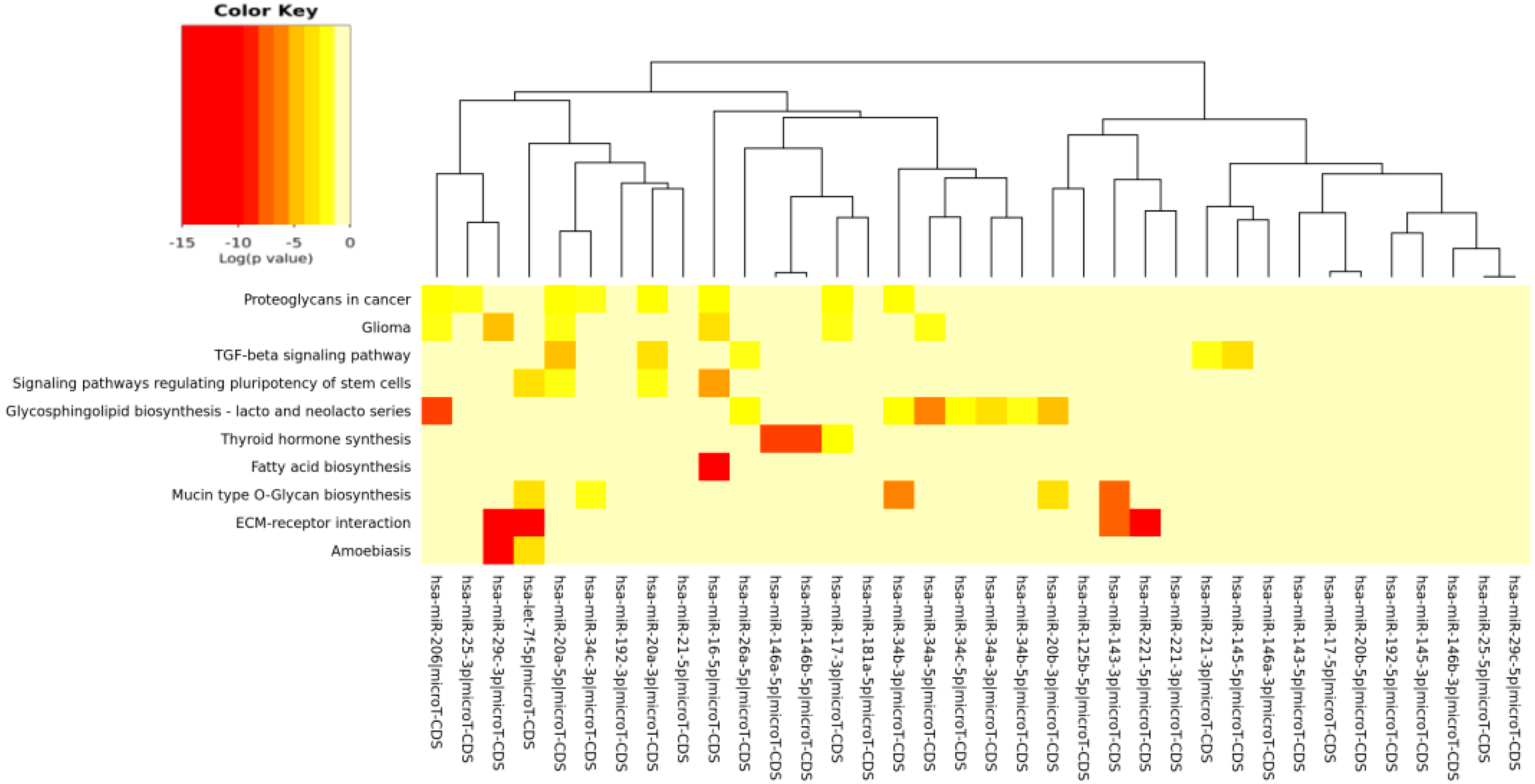
The DIANA program allowed us to investigate the common pathways associated with miRs. Among the most popular pathways involved in DD and DDR influenced by the compounds considered in this study, we found the fatty acids’ biosynthesis pathway and fatty acids’ metabolism. Alterations in fatty acid metabolism in cancer cells have received less attention, but are increasingly being recognized. Indeed, some studies have suggested that the DNA damage response is involved in the regulation of metabolic homeostasis. DNA damage could impair metabolic organ functions by causing cell death or senescence [
82]. There is a strong relationship between DD and energy metabolism [
83], and an implication for its role in tumorigenesis has been indicated. p53, the key factor in the DD/DDR network, has been reported to regulate fatty acid oxidation [
84], and genes involved in fatty acid metabolism are regulated by p53 in different cell types and in response to both oncogenic stress and DD [
85]. mTOR holds also an important role in lipid biosynthesis and metabolism [
86,
87] demonstrating a cross-talk among these pathways, and this fact has a significant meaning in cancerogenesis and chemoprevention. For example, by downregulating mTOR signaling and energy metabolism, it is possible to suppress the malignant phenotype of colorectal cancer cells [
88,
89]. Indeed, fatty acid synthase is a potential therapeutic target in cancer. There is strong evidence that constitutive mTOR signaling is the driving force of cellular and organismal aging and induction of senescence [
90,
91]. Extracellular matrix (ECM)-receptor interaction is also shown to be strongly involved in the DD/DDR and compounds interaction (EGCG, n3-PUFA, CRC). This is of interest in light of the evidence that ECM proteins are involved in cancer progression and outcome. The initiation of cell transformation is generally associated with genetic alterations in normal cells that lead to the loss of intercellular- and/or extracellular-matrix (ECM)-mediated cell adhesion [
92]. There is a connection between ECM proteins, such integrin signaling and DNA repair [
93]. There is also strong evidence that hyaluronic acid (HA; hyaluronate), the key component of ECM, by neutralizing the ROS, is reducing the induction of DNA damage by endogenous and exogenous oxidants [
94,
95,
96]. Particularly interesting is the role of HA, which is a large constituent of stem cells’ niche, in protecting the integrity of genome of stem cells against reactive oxidants [
95]. Our
in silico results evidence that signaling regulating the pluripotency of stem cells pathway is clearly targeted by many miRs selected in this study, as evidenced by the heat maps of specific compounds. miRs represent an important layer of regulation for stem cell self-renewal and differentiation [
97,
98]. Among DDR/compound-modulated miRs, in our analysis, let-7, miR-302 and miR-17-92 have been found in previous studies as regulators of cellular pluripotency [
99]. Signaling regulating the pluripotency of stem cells converges towards the activation of the transcriptional network and of many different pathways, which often show cross-talk in the determination of stem cell function. Indeed, FOXO, e.g., transcription factors, is required for DNA damage-induced growth arrest checkpoints [
100], and as a further example, the mTOR pathway has been shown to be a major regulator of both ROS levels and autophagy in human stem cells [
101]. The life-long persistence of stem cells in the body makes them particularly susceptible to the accumulation of cellular damage, which ultimately can lead to cell death, senescence or loss of regenerative function. Indeed, stem cells in many tissues have been found to undergo profound changes with age, exhibiting blunted responsiveness to tissue injury, dysregulation of proliferative activities and declining functional capacities. These changes translate into reduced effectiveness of cell replacement and tissue regeneration in aged organisms [
102]. Noteworthy, miR-146a/b and miR-195, which we found modulated by all compounds and by CRC, respectively, have been reported as age-induced miRNAs involved in stem cell senescence [
103]. These recent findings have demonstrated that silencing miR-195 reverses the senescence clock in aged stem cells by telomerase reactivation. The regulation of telomere length performed by the above-mentioned miR-155 may also have an important role in the cancer stem cell fate and cancer tissue homeostasis. miR-155 is upregulated in many cancers and can promote cancer stem cell phenotypes in liver [
104]. RSV has been demonstrated to be able to decrease its levels, supporting our prediction analysis [
57].
The availability of nutritional agents that can target specific miRs regulating stem cells’ function can improve regenerative potential in the body and, finally, counteract aging and prevent diseases, such as cancer.
It must be considered that most of the literature here analyzed refers to experimental research carried out on
in vitro models. The results indicated that exposure time and concentrations of compounds can differently affect miRNA expression in different human cells. In addition another important issue evident in the literature [
41] is the poor bioavailability of phytochemicals due to their metabolism, absorption and, consequently, the fact that most phytochemical compounds reach the targeted sites at sub-therapeutic concentrations [
105]. These observations pave the way for
in vivo dietary intervention studies and clinical trials focused on setting the appropriate dosage of the bioactive compounds that can effectively affect the miRs’ expression in humans.
The comparison of pathways identified, with our in silico analysis, to those identified experimentally in the study examples utilized here interestingly demonstrated that new pathways were evidenced from our prediction. Even more interesting, however, is that some of our predicted pathways were indeed found as directly implicated in some cancer studies and modulated by the compounds here considered.
The fact that experimental literature results have found that pathways, such as, for example, “fatty acid biosynthesis” and “fatty acid metabolism”, were actually affected by the compounds taken into account in this paper validated our prediction. This positive inter-comparison proves that the pathways computationally found by us were, indeed, the same found experimentally involved in some cancer models and in some senescence studies, confirming the validity of our prediction study. For examples, in breast and lung cancer cells, EGCG affected fatty acid metabolism [
106,
107]. Resveratrol was found to suppress cancer cell proliferation by inhibiting the fatty acid synthase signaling pathway [
108]. Curcumin, as well, induces apoptosis, inhibiting intracellular fatty acid synthase in human breast cancer [
109]. Clinical studies corroborate these findings [
110], and trials were also carried out [
111]. Resveratrol and EGCG are also found to improve the functional activity of the membrane lipids in aged liver models by influencing lipid composition and metabolism [
112,
113].
Presently, miRs have been already found to be modulated by the compounds here considered. On the other hand, other studies found that some of the pathways here evidenced are modulated by our compounds, but we provide here the link between miRs’ expression and the pathways’ modulation by the respective compounds. Thus, the importance of our results consists, indeed, of the possibility of detecting experimentally the link between miRs’ expression and pathways’ modulation.
Some authors suggested an interplay among miRNA, DD/DDR, specific signaling pathways and cancer promotion and invasion [
114,
115], and bioactive compounds play an important role in this interplay.
Taken together, the clinical relevance of such observations could be related to the bioactive compounds’ chemopreventive value in some type of tumors, as for example in breast cancers or lung cancers, where enough evidence corroborates this hypothesis.
In conclusion, since DDR and these pathways are strongly related to aging and carcinogenesis, the in silico results of our study suggest that monitoring the induction of specific miRs may provide the means to assess the antiaging and chemopreventive properties of particular bioactive compounds. Furthermore, the results of this in silico study indicate the most important pathways potentially modulated by dietary compounds and provide a framework for the generation of new strategies to design experimental studies and, finally, new anti-cancer and anti-aging therapies.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}